Journal of Ecology ( IF 5.3 ) Pub Date : 2022-07-14 , DOI: 10.1111/1365-2745.13965 Roger Grau‐Andrés 1 , Paul Kardol 1 , Michael J. Gundale 1
|
1 INTRODUCTION
Plant traits are strong drivers of plant function and thus play a key role in ecosystem processes in terrestrial ecosystems (Díaz et al., 2016; Mazziotta et al., 2019; Reich, 2014). As such, plant trait co-variation is seen as a powerful tool to predict how environmental change alters plant traits and ultimately ecosystem functioning (Funk et al., 2017; Suding et al., 2008). Specifically, anthropogenic climate change can alter trait composition of plant communities through affecting species fitness and physiological adaptation (Henn et al., 2018; Springate & Kover, 2014), which can then impact on key ecosystem processes including carbon (C) and nutrient cycling (Cornwell et al., 2008; Madani et al., 2018). In vascular plants, there is ample evidence of trade-offs in their architectural, physiological and chemical trait spectra, which are driven by differences in the allocation of key resources (i.e. C, nutrients and water; Freschet et al., 2010; Reich, 2014). For example, low leaf mass per area tends to be associated with high leaf nutrient concentrations, high photosynthetic capacity and high litter decomposition rates (the ‘leaf economics spectrum’; Wright et al., 2004). Compared to vascular plants, trait co-variation has been much less studied in bryophytes (Cornelissen et al., 2007; Mazziotta et al., 2019; Wang et al., 2017), despite the importance of bryophytes (especially mosses) in C and nutrient dynamics in many ecosystems (Hupperts et al., 2021; Lindo et al., 2013; Street et al., 2013; Turetsky et al., 2012). A better understanding of trait trade-offs in bryophytes is needed to more accurately predict how environmental change may alter bryophyte function (i.e. by altering their trait spectra; Cornelissen et al., 2007; Mazziotta et al., 2019) and impact ecosystem function in bryophyte-dominated ecosystems (Moor et al., 2017).
Research on trait trade-offs in bryophytes has so far provided mixed support for the existence of economics spectra similar to those observed in vascular plants. For example, a positive correlation between nutrient concentration and photosynthetic capacity in bryophyte shoots (in line with the leaf economics spectrum in vascular plants) has been found in some studies (Carriquí et al., 2019; Wang et al., 2014, 2016) but not in others (Bengtsson et al., 2016; Laing et al., 2014; Rice et al., 2008; Waite & Sack, 2010, 2011). Similarly, while some studies support the expectation (based on trait correlations in vascular plants) that lower bryophyte shoot or canopy mass per area is associated with greater nutrient concentrations and photosynthetic rates (Carriquí et al., 2019; Waite & Sack, 2010; Wang et al., 2016), others found limited or no support (Bengtsson et al., 2016; Laing et al., 2014; Waite & Sack, 2011). The discrepancy may be due in part to the fact that architectural traits can be measured at the shoot- or canopy-level. For example, for an analogue of the vascular plant trait ‘leaf mass per area’, some authors have used shoot-based measurements of mass per area (projected on the ground, Wang et al., 2016, or transversal to the shoot, Bond-Lamberty & Gower, 2007, Jonsson et al., 2015), while others have used measurements of canopy mass per ground area (Bansal et al., 2012; Waite & Sack, 2010). In this context, canopy-based measures may better reflect field conditions as they account for effects of self-shading and moisture regulation, which operate at the canopy level (Cornelissen et al., 2007; Elumeeva et al., 2011; Rice & Cornelissen, 2014). Furthermore, much of the research on trait trade-offs in bryophytes has focused on wetlands and on Sphagnum mosses, which limits our understanding of whether and how economics spectra are established across wider bryophyte groups and habitat types.
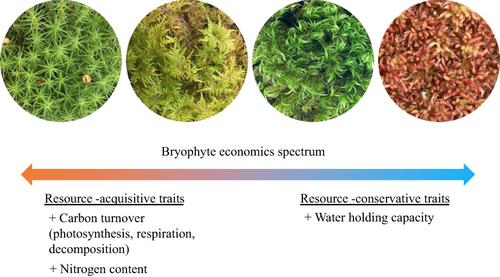
Research on Sphagnum mosses has suggested that traits related to moisture dynamics may be crucial in shaping trait trade-offs, from species forming denser canopies with higher moisture retention capacity, low nutrient concentrations, low photosynthetic capacity and low litter decomposition rates (i.e. resource-conservative) to species forming low-density canopies with the opposite traits (i.e. resource-acquisitive; Bengtsson et al., 2016; Laing et al., 2014; Rice et al., 2008). Given that the functioning of bryophytes is often highly dependent on fast-changing moisture availability (Dilks & Proctor, 1979), traits related to moisture regulation are likely key in driving trait trade-offs also across a wider range of bryophyte functional groups. However, work on trait trade-offs including water traits in non-Sphagnum mosses is very limited and has so far only provided inconclusive evidence of a negative association between water retention capacity and maximum photosynthetic capacity (Wang & Bader, 2018). Moreover, no study has yet tested economics spectra jointly for traits related to C, nutrients and water regulation in a wide range of mosses.
Here, we investigate trait economics spectra in boreal forest mosses. Boreal forests are important drivers of global C dynamics and typically have high abundance of ground mosses (Hupperts et al., 2021; Nilsson & Wardle, 2005; Turetsky et al., 2012), yet research on moss trait co-variation is limited (Jonsson et al., 2015; Rice et al., 2011) and moss trait spectra remain untested in boreal forests. To investigate this, we measured architectural, physiological and chemical trait co-variation in a wide range of boreal forest mosses, including canopy traits, which are most relevant to ecosystem function (Rice & Cornelissen, 2014). We hypothesized that (H1) trait co-variation in bryophytes is structured around a single economics spectrum shaped by C, nutrient and water conservation. We expected to find a bryophyte spectrum similar to what has been shown for vascular plants, ranging from resource-conservative traits (e.g. high canopy mass per area, high moisture retention capacity, low photosynthesis and respiration rates, low shoot nutrient concentration, low litter decomposability) to resource-acquisitive traits. Specifically, we hypothesized that (H1a) C turnover (e.g. photosynthetic and respiratory rates, litter decomposability) would be positively correlated with tissue nitrogen concentration, as has been observed in vascular plants (Wright et al., 2004); (H1b) C turnover would be negatively associated with water-holding capacity of the moss canopy. This is because plant traits that maximize water retention are inherently conservative and cause a trade-off with C uptake and turnover (Mazziotta et al., 2019; Reich, 2014); (H1c) photosynthetic capacity would be negatively correlated with canopy-based architectural traits (e.g. canopy mass per area) but uncorrelated with shoot-based architectural traits (e.g. specific shoot area), because the latter do not account for important regulatory processes (e.g. self-shading) at the canopy level (Cornelissen et al., 2007; Waite & Sack, 2010). Finally, we hypothesized that (H2) resource-conservative strategies would dominate in mosses of open and wet habitats, while resource-acquisitive traits would dominate in dry and shaded habitats. We expected this based on work on Sphagnum mosses showing that reduced light and water availability are associated with resource-acquisitive traits (Mazziotta et al., 2019). By improving our understanding of bryophyte trait co-variation, our study can help predict how environmental changes may affect bryophyte function and thus ecosystem function in bryophyte-dominated ecosystems.
中文翻译:
北方苔藓的性状协调揭示了苔藓植物的经济学谱
1 简介
植物性状是植物功能的强大驱动力,因此在陆地生态系统的生态系统过程中发挥着关键作用(Díaz 等人, 2016 年;Mazziotta 等人, 2019 年;Reich, 2014 年)。因此,植物性状共变被视为预测环境变化如何改变植物性状并最终改变生态系统功能的有力工具(Funk 等人, 2017 年;Suding 等人, 2008 年)。具体来说,人为气候变化可以通过影响物种适应性和生理适应来改变植物群落的性状组成(Henn et al., 2018 ; Springate & Kover, 2014),然后会影响关键的生态系统过程,包括碳 (C) 和养分循环 (Cornwell et al., 2008 ; Madani et al., 2018 )。在维管植物中,有充分的证据表明其结构、生理和化学性状谱是由关键资源(即 C、养分和水)分配的差异驱动的;Freschet 等人, 2010 年;Reich, 2014 年)。例如,单位面积的低叶质量往往与高叶养分浓度、高光合能力和高凋落物分解率相关(“叶经济谱”;Wright 等人, 2004 年))。与维管植物相比 , 尽管苔藓植物(尤其是苔藓 )在 C许多生态系统中的养分动态(Hupperts 等人, 2021 年;Lindo 等人, 2013 年;Street 等人, 2013 年;Turetsky 等人, 2012 年)。需要更好地了解苔藓植物的性状权衡,以更准确地预测环境变化如何改变苔藓植物的功能(即通过改变其性状谱;Cornelissen 等人, 2007 年;Mazziotta 等人, 2019 年)) 并影响以苔藓植物为主的生态系统中的生态系统功能(Moor 等人, 2017 年)。
迄今为止,关于苔藓植物性状权衡的研究为存在类似于在维管植物中观察到的经济学光谱提供了不同的支持。例如,在一些研究中发现了苔藓植物芽的养分浓度和光合能力之间的正相关(与维管植物的叶片经济学谱一致)(Carriquí 等人, 2019 年;Wang 等人, 2014 年,2016 年)但其他人则不然(Bengtsson 等人, 2016 年;Laing 等人, 2014 年;Rice 等人, 2008 年;Waite & Sack, 2010 年,2011 年)。同样,虽然一些研究支持这样的预期(基于维管植物的性状相关性),即较低的苔藓植物枝条或每面积冠层质量与较高的养分浓度和光合速率相关(Carriquí 等人, 2019 年;Waite & Sack, 2010 年;Wang等人, 2016 年),其他人发现支持有限或没有支持(Bengtsson 等人, 2016 年;Laing 等人, 2014 年;Waite & Sack, 2011 年)。这种差异可能部分是由于建筑特征可以在枝条或冠层水平测量。例如,对于维管植物性状“每面积叶质量”的类似物,一些作者使用了基于枝条的每面积质量测量(投影在地面上,Wang 等人, 2016 年,或横向于枝条,邦德-Lamberty & Gower, 2007 , Jonsson et al., 2015 ),而其他人则使用了单位面积冠层质量的测量值 (Bansal et al., 2012 ; Waite & Sack, 2010 )。在这种情况下,基于冠层的措施可以更好地反映田间条件,因为它们考虑了在冠层水平上运行的自遮蔽和湿度调节的影响(Cornelissen 等人, 2007 年;Elumeeva 等人, 2011 年;赖斯和科内利森, 2014 年)。此外,关于苔藓植物性状权衡的大部分研究都集中在湿地和泥炭藓上,这限制了我们对是否以及如何在更广泛的苔藓植物群和栖息地类型中建立经济学谱的理解。
对泥炭藓的研究表明,与水分动力学相关的性状可能对形成性状权衡至关重要,从物种形成具有更高水分保持能力、低营养浓度、低光合能力和低凋落物分解率(即资源节约型)的密集冠层) 对形成具有相反特征的低密度树冠的物种(即资源获取性;Bengtsson 等人, 2016 年;Laing 等人, 2014 年;Rice 等人, 2008 年)。鉴于苔藓植物的功能通常高度依赖于快速变化的水分可用性(Dilks & Proctor, 1979),与水分调节相关的性状可能是推动更广泛的苔藓植物功能组的性状权衡的关键。然而,包括非泥炭藓的水分性状在内的性状权衡工作非常有限,迄今为止仅提供了关于保水能力和最大光合能力之间负相关关系的不确定证据(Wang & Bader, 2018 年)。此外,还没有研究联合测试与广泛的苔藓中的 C、养分和水分调节相关的性状的经济学光谱。
在这里,我们研究了北方森林苔藓的性状经济学谱。北方森林是全球碳动态的重要驱动因素,通常具有高丰度的地面苔藓(Hupperts 等人, 2021 年;Nilsson 和 Wardle, 2005 年;Turetsky 等人, 2012 年),但对苔藓特征共变的研究有限( Jonsson et al., 2015 ; Rice et al., 2011 ) 和苔藓性状光谱在北方森林中仍未得到测试。为了研究这一点,我们测量了广泛的北方森林苔藓的建筑、生理和化学性状的协变,包括与生态系统功能最相关的冠层性状 (Rice & Cornelissen, 2014)。我们假设苔藓植物中的(H1)性状共变是围绕由 C、养分和水分保护形成的单一经济谱构建的。我们希望找到与维管植物相似的苔藓植物谱,范围从资源保守性状(例如,单位面积的高冠层质量、高水分保持能力、低光合作用和呼吸速率、低枝条营养浓度、低凋落物分解性) ) 到资源获取性特征。具体来说,我们假设 (H1a) C 周转率(例如光合和呼吸速率、凋落物分解能力)与组织氮浓度呈正相关,正如在维管植物中观察到的那样(Wright 等, 2004); (H1b) C 周转率与苔藓树冠的持水能力呈负相关。这是因为最大化保水性的植物性状本质上是保守的,会导致与 C 吸收和周转的权衡(Mazziotta 等人, 2019 年;Reich, 2014 年);(H1c) 光合能力与基于冠层的建筑特征(例如每面积的冠层质量)呈负相关,但与基于枝条的建筑特征(例如特定枝条面积)不相关,因为后者不考虑重要的调节过程(例如自我-shading) 在树冠层 (Cornelissen et al., 2007 ; Waite & Sack, 2010)。最后,我们假设(H2)资源保护策略将在开放和潮湿生境的苔藓中占主导地位,而资源获取性特征将在干燥和阴影生境中占主导地位。我们预计这是基于对泥炭藓的研究,表明减少的光和水的可用性与资源获取性状有关(Mazziotta 等人, 2019 年)。通过提高我们对苔藓植物性状共变的理解,我们的研究可以帮助预测环境变化如何影响苔藓植物功能,从而影响以苔藓植物为主的生态系统中的生态系统功能。










































 京公网安备 11010802027423号
京公网安备 11010802027423号