Journal of Ecology ( IF 5.3 ) Pub Date : 2022-05-27 , DOI: 10.1111/1365-2745.13930 Laura García-Velázquez 1, 2 , Antonio Gallardo 1, 3 , Victoria Ochoa 2 , Beatriz Gozalo 2 , Roberto Lázaro 4 , Fernando T Maestre 2, 5
|
1 INTRODUCTION
Water and nitrogen (N) availability have been traditionally considered the main limiting resources for life in dryland ecosystems (Delgado-Baquerizo et al., 2013b; Robertson and Groffman, 2015; Sardans et al., 2008; Schlesinger et al., 1995), which cover 41% of terrestrial surface (Cherlet et al., 2018). Thus, it is not surprising to find that most research conducted so far on the impacts of ongoing climate change on dryland biogeochemistry have focused on the responses of C and N (Delgado-Baquerizo et al., 2013a; Jiao et al., 2016; Nielsen and Ball, 2015). In recent years, however, there is a growing interest in understanding the role of P in drylands, particularly after studies suggesting that increases in aridity, a hallmark of climate change in drylands worldwide (Huang et al., 2015), imbalance the proportion of N and P in the soil (Delgado-Baquerizo et al., 2013a; Jiao et al., 2016). Despite so, the impacts of climate change drivers such as warming and altered rainfall patterns on the complex diversity of P forms in dryland soils remain largely unexplored (García-Velázquez et al., 2020) and have not, to the best of our knowledge, been assessed experimentally yet. This lack of knowledge limits our ability to incorporate changes in the availability of P and its impacts on processes such as productivity into models aiming to forecast ecosystem responses to climate change.
The inputs of P into the soils come mainly from parent rock weathering (Lajtha and Schlesinger, 1988; Walker and Syers, 1976) and, to a lesser extent, from deposition of the P present in atmospheric dust (Okin et al., 2004). The latter is quite insoluble in the soil solution (Chen et al., 2006; Shafqat et al., 2016), but it could be an essential source of P in ecosystems limited by this macronutrient (Chadwick et al., 1999; Okin et al., 2004). Over 95% of soil total P is associated with primary or secondary minerals, constituted in organic forms or occluded by soil minerals and immediately unavailable for plants (Condron and Newman, 2011; Shen et al., 2011). Organic P has a key role in plant nutrition since it is transformed to inorganic P fractions through mineralization mediated by the release of phosphatase enzymes from microbes and plant roots (Condron et al., 2005; Turner and Engelbrecht, 2011). Both biological (i.e. mineralization/immobilization) and geochemical (i.e. precipitation/dissolution and sorption/desorption) processes contribute to P availability (Cross and Schlesinger, 2001; Hou et al., 2018a; Vitousek et al., 2010), and are strongly dependent on precipitation and temperature (Belnap, 2011; Feng et al., 2016; Hou et al., 2018a). Increases in temperature have been shown to favour P sorption on secondary minerals and increase the occluded P (Hou et al., 2018a), and to reduce P availability due to the loss of soil moisture (Sardans et al., 2006; Sardans and Peñuelas, 2004). Moreover, low precipitation blocks microbial activity, ion diffusion and nutrient uptake by plants (Belnap, 2011; Sardans and Peñuelas, 2004). Drier conditions thus promote a slowdown in P release due to decreased litter decomposition rate and enzymatic activity, contributing to increases in soil humic compounds that keep P in less available forms (Sardans and Peñuelas, 2004). Finally, the synergistic effects of temperature and precipitation control the rates of solubilization and release of labile P bound to carbonates into the soil solution (Belnap, 2011). Thus, high temperatures and drier conditions, such as those expected for drylands under climate change (Greve et al., 2014; Huang et al., 2015), would deter both the increases in concentrations of soil carbonic acid and the decrease in soil pH, reducing the dissolution rate of carbonates and the transition of P to soil solution phase (Devau et al., 2010). However, we have no experimental evidence of temperature and precipitation effects on P availability in dryland soils.
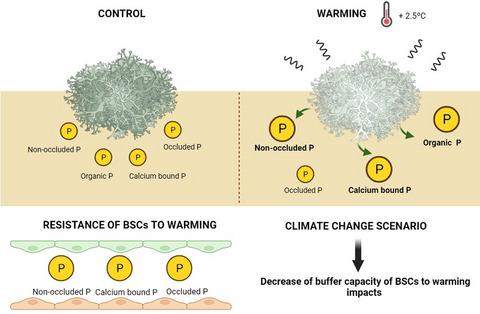
The low moisture content of dryland soils favours that P inputs (from rock weathering, dry deposition and decomposition of litter and organic matter) are not washed to deeper horizons, and thus concentrate in the topsoil (Verrecchia et al., 1995; White et al., 2004). This makes soil P fractions sensitive to the activity of organisms living on the soil surface, such as biocrusts (communities composed by lichens, mosses, bacteria, cyanobacteria, fungi, algae and liverworts that are a major biotic feature of drylands worldwide; Weber et al., 2016). Biocrusts are known to influence soil P availability through secretion of organic acids, chelating agents and the excretion of H+ during respiration, which solubilize mineral bound P increasing available P (Baumann et al., 2017, 2019; Belnap, 2011). Furthermore, the microtopography created by biocrust constituents like lichens could facilitate the capture of silt and clay particles from dust (Fick et al., 2020), increasing soil water and nutrient contents (Delgado-Baquerizo et al., 2015; Eldridge et al., 2020), promoting higher rates of microbial activity (Delgado-Baquerizo et al., 2015) and avoiding P losses via lixiviation (Belnap, 2011; Belnap et al., 2004). Different experiments have shown how warming and altered rainfall regimes such as those forecasted under climate change can dramatically decline the cover and abundance of lichen- and moss-dominated biocrusts (Ferrenberg et al., 2015; Ladrón de Guevara et al., 2018; Maestre et al., 2015), with cascading effects on the C and N cycles driven by this decline (Delgado-Baquerizo et al., 2014; Liu et al., 2016, 2017; Maestre et al., 2013; Reed et al., 2012). However, there is no experimental evidence about whether dryland biocrusts can modulate all P soil fractions, which will determine its availability for plants at short, medium and long terms, in the topsoil under climate change.
Understanding how the interactions between biocrusts and climate change affect soil P fractions in drylands is essential to better comprehend its impacts on these ecosystems and their capacity to provide essential ecosystem services in a warmer world. However, few studies so far have evaluated how simulated climate change affect soil P fractions in drylands, and these have been focused on a few P fractions (e.g. Olsen-P, NaOH-EDTA), have evaluated the effects of warming in isolation from other climate change drivers and have been conducted on dry-subhumid shrublands and forests (e.g. Sardans et al., 2006, 2008; Sardans and Peñuelas, 2004; Zhang et al., 2014). Here we aimed to evaluate how warming (~2.5°C increase), rainfall reduction (~30% reduction) and biocrust cover (<20% vs. >50%) affect soil P pools (non-occluded P, organic P, calcium bound P, occluded P and total P) on the top 1 cm of the soil and their resistance to climate change in two semi-arid grasslands from central and southeastern Spain. We focus on the effects of warming and precipitation on soil depths which biocrust have the potential to alter. Still, we are aware that deeper depths have also the potential to be altered by these climatic variables. We tested the general hypothesis that climate change and biocrusts will directly or indirectly affect to all major organic and mineral P pools because they are interconnected by microbial processes (García-Velázquez et al., 2020). More specifically, we hypothesized that (a) soils under biocrusts will have higher concentration of both labile and stable P pools (Baumann et al., 2017, 2019); (b) warming and, to a lesser extent, rainfall reduction will positively affect both more labile (i.e. non-occluded P, organic P) and stable (i.e. calcium bound P and occluded P) P pools of surface soil layer influenced by biocrusts (Hou et al., 2018a; Sardans and Peñuelas, 2004); and (c) well-developed biocrusts will increase the resistance (according the Orwin and Wardle index; Orwin and Wardle, 2004) of all P pools to warming and, to a lesser extent, to rainfall reduction (Belnap, 2011; Delgado-Baquerizo et al., 2014).
中文翻译:
生物结皮增强了对变暖引起的表土磷库增加的抵抗力
1 简介
水和氮 (N) 的可用性传统上被认为是旱地生态系统生命的主要限制资源(Delgado-Baquerizo 等, 2013b ;Robertson 和 Groffman, 2015 ;Sardans 等, 2008 ;Schlesinger 等, 1995 ) ,覆盖了 41% 的陆地表面(Cherlet 等人, 2018 )。因此,毫不奇怪地发现,迄今为止,大多数关于持续气候变化对旱地生物地球化学影响的研究都集中在碳和氮的响应上(Delgado-Baquerizo等, 2013a ;Jiao等, 2016 ;尼尔森和鲍尔, 2015 )。然而,近年来,人们越来越有兴趣了解磷在旱地中的作用,特别是在研究表明干旱加剧(全球旱地气候变化的一个标志)后(Huang et al., 2015 ),磷的比例失衡。土壤中的氮和磷(Delgado-Baquerizo 等, 2013a ;Jiao 等, 2016 )。尽管如此,气候变化驱动因素(例如变暖和降雨模式改变)对旱地土壤中磷形态复杂多样性的影响在很大程度上仍未得到探索(García-Velázquez et al., 2020 ),而且据我们所知,尚未得到研究。尚未经过实验评估。这种知识的缺乏限制了我们将磷的可用性变化及其对生产力等过程的影响纳入旨在预测生态系统对气候变化响应的模型的能力。
进入土壤的磷主要来自母岩风化(Lajtha 和 Schlesinger, 1988 ;Walker 和 Syers, 1976 ),还有少量来自大气尘埃中磷的沉积(Okin 等, 2004 ) 。后者在土壤溶液中相当难溶(Chen 等人, 2006 年;Shafqat 等人, 2016 年),但它可能是受这种大量营养素限制的生态系统中磷的重要来源(Chadwick 等人, 1999 年;Okin 等人)等, 2004 )。超过 95% 的土壤总磷与原生或次生矿物质相关,以有机形式构成或被土壤矿物质吸留,并且立即无法被植物利用(Condron 和 Newman, 2011 ;Shen 等, 2011 )。有机磷在植物营养中具有关键作用,因为它通过微生物和植物根部释放磷酸酶介导的矿化作用转化为无机磷部分(Condron 等, 2005 ;Turner 和 Engelbrecht, 2011 )。生物(即矿化/固定)和地球化学(即沉淀/溶解和吸附/解吸)过程都有助于磷的有效性(Cross和Schlesinger, 2001 ;Hou等, 2018a ;Vitousek等, 2010 ),并且强烈取决于降水量和温度(Belnap, 2011 ;Feng 等, 2016 ;Hou 等, 2018a )。温度升高已被证明有利于次生矿物上的 P 吸附并增加吸留的 P(Hou 等,2016)。, 2018a ),并由于土壤水分流失而减少磷的有效性(Sardans 等, 2006 ;Sardans 和 Peñuelas, 2004 )。此外,低降水量会阻碍微生物活动、离子扩散和植物对养分的吸收(Belnap, 2011 ;Sardans 和 Peñuelas, 2004 )。因此,由于凋落物分解率和酶活性降低,干燥条件促进磷释放减慢,导致土壤腐殖质化合物增加,使磷保持较少的可用形式(Sardans和Peñuelas, 2004年)。最后,温度和降水的协同效应控制与碳酸盐结合的不稳定磷溶解和释放到土壤溶液中的速率(Belnap, 2011 )。因此,高温和干燥条件,例如气候变化下旱地的预期条件(Greve等, 2014 ;Huang等, 2015 ),将阻止土壤碳酸浓度的增加和土壤pH值的降低,降低碳酸盐的溶解速率和磷向土壤溶液相的转变(Devau et al., 2010 )。然而,我们没有实验证据表明温度和降水对旱地土壤磷有效性的影响。
旱地土壤的低含水量有利于磷的输入(来自岩石风化、干沉降以及凋落物和有机物的分解)不会被冲刷到更深的地平线,从而集中在表土中(Verrecchia等, 1995 ;White等., 2004 )。这使得土壤磷组分对生活在土壤表面的生物体的活动敏感,例如生物结皮(由地衣、苔藓、细菌、蓝细菌、真菌、藻类和地钱组成的群落,是全世界旱地的主要生物特征;Weber 等人) ., 2016 )。众所周知,生物结皮通过有机酸、螯合剂的分泌和呼吸过程中 H+ 的排泄来影响土壤磷的有效性,从而溶解矿物质结合的磷,增加有效磷(Baumann 等人, 2017 年, 2019 年;Belnap, 2011 年)。此外,地衣等生物结皮成分形成的微地形可以促进从灰尘中捕获淤泥和粘土颗粒(Fick 等人, 2020 ),增加土壤水分和养分含量(Delgado-Baquerizo 等人, 2015 ;Eldridge 等人,2015)。 , 2020 ),促进更高的微生物活性(Delgado-Baquerizo 等, 2015 )并避免通过浸出造成的磷损失(Belnap, 2011 ;Belnap 等, 2004 )。不同的实验表明,变暖和降雨状况的改变(例如气候变化下预测的降雨状况)如何显着降低以地衣和苔藓为主的生物结皮的覆盖范围和丰度(Ferrenberg 等人, 2015 年;Ladrón de Guevara 等人, 2018 年;Maestre)等人。, 2015 ),这种下降对碳和氮循环产生了连锁效应(Delgado-Baquerizo等, 2014 ;Liu等, 2016,2017 ; Maestre等, 2013 ;Reed等, 2012 ) 。然而,没有实验证据表明旱地生物结皮是否可以调节所有土壤磷组分,这将决定气候变化下表土中植物短期、中期和长期的可用性。
了解生物结皮和气候变化之间的相互作用如何影响旱地土壤磷含量对于更好地理解其对这些生态系统的影响及其在变暖的世界中提供基本生态系统服务的能力至关重要。然而,迄今为止,很少有研究评估模拟气候变化如何影响旱地土壤磷组分,并且这些研究都集中在少数磷组分(例如 Olsen-P、NaOH-EDTA)上,并独立于其他组分评估了变暖的影响。气候变化驱动因素,并已在干燥半湿润灌木丛和森林中进行了研究(例如 Sardans 等人, 2006 年、 2008 年;Sardans 和 Peñuelas, 2004 年;Zhang 等人, 2014 年)。在这里,我们的目的是评估变暖(约增加 2.5°C)、降雨减少(减少约 30%)和生物结皮覆盖(<20% 与 >50%)如何影响土壤磷库(非吸留磷、有机磷、钙)西班牙中部和东南部两个半干旱草原土壤表层 1 厘米的束缚磷、滞留磷和总磷及其对气候变化的抵抗力。我们关注变暖和降水对生物结皮可能改变的土壤深度的影响。尽管如此,我们意识到更深的深度也有可能被这些气候变量改变。我们测试了一般假设,即气候变化和生物结皮将直接或间接影响所有主要有机和矿物磷库,因为它们通过微生物过程相互关联(García-Velázquez 等人, 2020 )。更具体地说,我们假设(a)生物结皮下的土壤将具有更高浓度的不稳定和稳定磷库(Baumann 等人,2017)。、 2017 年、 2019 年); (b) 气候变暖以及较小程度的降雨量减少将对受生物结皮影响的表土层的更不稳定(即非吸留磷、有机磷)和稳定(即钙结合磷和吸留磷)磷库产生积极影响( Hou 等人, 2018a ;Sardans 和 Peñuelas, 2004 ); (c) 发育良好的生物结皮将增加所有磷库对变暖的抵抗力(根据 Orwin 和 Wardle 指数;Orwin 和 Wardle, 2004 年),并在较小程度上增加对降雨量减少的抵抗力(Belnap, 2011 年;Delgado-Baquerizo)等人, 2014 )。










































 京公网安备 11010802027423号
京公网安备 11010802027423号