当前位置:
X-MOL 学术
›
J. Zool. Syst. Evol. Res.
›
论文详情
Our official English website, www.x-mol.net, welcomes your
feedback! (Note: you will need to create a separate account there.)
Phylogenetic topology and timing of New Zealand olive shells are consistent with punctuated equilibrium
Journal of Zoological Systematics and Evolutionary Research ( IF 2.0 ) Pub Date : 2019-11-22 , DOI: 10.1111/jzs.12342 Michael R. Gemmell 1 , Steven A. Trewick 1 , Simon F. K. Hills 1 , Mary Morgan‐Richards 1
Journal of Zoological Systematics and Evolutionary Research ( IF 2.0 ) Pub Date : 2019-11-22 , DOI: 10.1111/jzs.12342 Michael R. Gemmell 1 , Steven A. Trewick 1 , Simon F. K. Hills 1 , Mary Morgan‐Richards 1
Affiliation
|
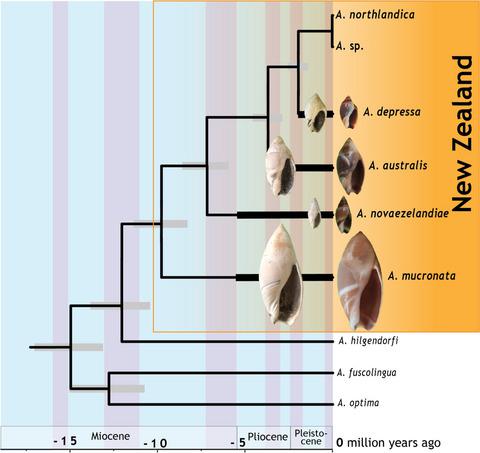
The olive shells (Amalda H. & A. Adams, 1853, Ancillariidae) are marine gastropods largely confined to temperate regions (Olson, 1956) with almost 100 species throughout the Pacific and Atlantic oceans. The highest species diversity is in tropical waters with few species in the southern Pacific (Kantor, Fedosov, Puillandre, Bonillo, & Bouchet, 2017). Amalda generally live in sandy near‐shore envi‐ ronments where they are predators, primarily feeding on bivalves (Powell, 1979). The benthic, soft‐sediment habitat of Amalda lends itself to high probability of fossilization, and in New Zealand, Amalda has a fossil record dating back 45 million years to Eocene fossil beds (Beu & Maxwell, 1990). Of the seven currently recognized extant species in New Zealand waters, four have lineages that extend into the fossil record: A. mucronata Sowerby I, 1830, A. australis Sowerby I, 1830, A. depressa Sowerby II, 1859, and A. novaezelandiae Sowerby II, 1859 (Beu & Maxwell, 1990). New Zealand Amalda have featured in debate about models of morphological evolution since they were presented as examples in support of punctuated equilibrium (Gould, 1991, 2009; Michaux, 1989). Fossil forms identified as “species” were inferred to be equiv‐ alent to reproductively isolated lineages identified with genetic markers and provided strong evidence of morphological stasis over millions of years (Michaux, 1987, 1989). However, accurate identi‐ fication of lineages and morphological stasis are both important elements of the punctuated equilibrium theory and to date, no Received: 5 November 2018 | Revised: 10 August 2019 | Accepted: 20 August 2019 DOI: 10.1111/jzs.12342
中文翻译:

新西兰橄榄壳的系统发育拓扑和时间与间断平衡一致
橄榄壳 (Amalda H. & A. Adams, 1853, Ancillariidae) 是海洋腹足类动物,主要分布在温带地区 (Olson, 1956),在太平洋和大西洋有近 100 种。物种多样性最高的是热带水域,南太平洋的物种很少(Kantor、Fedosov、Puillandre、Bonillo 和 Bouchet,2017 年)。Amalda 通常生活在沙质近岸环境中,在那里它们是捕食者,主要以双壳类动物为食(Powell,1979)。Amalda 的底栖软沉积物栖息地很容易形成化石,而在新西兰,Amalda 的化石记录可以追溯到始新世化石床 (Beu & Maxwell, 1990) 的 4500 万年前。在新西兰水域目前公认的七种现存物种中,有四种具有延伸到化石记录的谱系:A. mucronata Sowerby I, 1830, A. australis Sowerby I, 1830, A. depressa Sowerby II, 1859, 和 A. novaezelandiae Sowerby II, 1859 (Beu & Maxwell, 1990)。新西兰 Amalda 在关于形态进化模型的辩论中占有重要地位,因为它们被用作支持间断平衡的例子(Gould, 1991, 2009; Michaux, 1989)。被确定为“物种”的化石形式被推断为等同于通过遗传标记确定的生殖隔离谱系,并提供了数百万年来形态停滞的有力证据(Michaux,1987,1989)。然而,谱系的准确识别和形态停滞都是间断平衡理论的重要元素,迄今为止,没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342 australis Sowerby I, 1830, A. depressa Sowerby II, 1859, 和 A. novaezelandiae Sowerby II, 1859 (Beu & Maxwell, 1990)。新西兰 Amalda 在关于形态进化模型的辩论中占有重要地位,因为它们被用作支持间断平衡的例子(Gould,1991 年,2009 年;Michaux,1989 年)。被确定为“物种”的化石形式被推断为等同于通过遗传标记确定的生殖隔离谱系,并提供了数百万年来形态停滞的有力证据(Michaux,1987,1989)。然而,谱系的准确识别和形态停滞都是间断平衡理论的重要元素,迄今为止,没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342 australis Sowerby I, 1830, A. depressa Sowerby II, 1859, 和 A. novaezelandiae Sowerby II, 1859 (Beu & Maxwell, 1990)。新西兰 Amalda 在关于形态进化模型的辩论中占有重要地位,因为它们被用作支持间断平衡的例子(Gould,1991 年,2009 年;Michaux,1989 年)。被确定为“物种”的化石形式被推断为等同于通过遗传标记确定的生殖隔离谱系,并提供了数百万年来形态停滞的有力证据(Michaux,1987,1989)。然而,谱系的准确识别和形态停滞都是间断平衡理论的重要元素,迄今为止,没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342 和 A. novaezelandiae Sowerby II, 1859 (Beu & Maxwell, 1990)。新西兰 Amalda 在关于形态进化模型的辩论中占有重要地位,因为它们被用作支持间断平衡的例子(Gould,1991 年,2009 年;Michaux,1989 年)。被确定为“物种”的化石形式被推断为等同于通过遗传标记确定的生殖隔离谱系,并提供了数百万年来形态停滞的有力证据(Michaux,1987,1989)。然而,谱系的准确识别和形态停滞都是间断平衡理论的重要元素,迄今为止,没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342 和 A. novaezelandiae Sowerby II, 1859 (Beu & Maxwell, 1990)。新西兰 Amalda 在关于形态进化模型的辩论中占有重要地位,因为它们被用作支持间断平衡的例子(Gould,1991 年,2009 年;Michaux,1989 年)。被确定为“物种”的化石形式被推断为等同于通过遗传标记确定的生殖隔离谱系,并提供了数百万年来形态停滞的有力证据(Michaux,1987,1989)。然而,谱系的准确识别和形态停滞都是间断平衡理论的重要元素,迄今为止,没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342 新西兰 Amalda 在关于形态进化模型的辩论中占有重要地位,因为它们被用作支持间断平衡的例子(Gould,1991 年,2009 年;Michaux,1989 年)。被确定为“物种”的化石形式被推断为等同于通过遗传标记确定的生殖隔离谱系,并提供了数百万年来形态停滞的有力证据(Michaux,1987,1989)。然而,谱系的准确识别和形态停滞都是间断平衡理论的重要元素,迄今为止,没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342 新西兰 Amalda 在关于形态进化模型的辩论中占有重要地位,因为它们被用作支持间断平衡的例子(Gould,1991 年,2009 年;Michaux,1989 年)。被确定为“物种”的化石形式被推断为等同于通过遗传标记确定的生殖隔离谱系,并提供了数百万年来形态停滞的有力证据(Michaux,1987,1989)。然而,谱系的准确识别和形态停滞都是间断平衡理论的重要元素,迄今为止,没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342 被确定为“物种”的化石形式被推断为等同于通过遗传标记确定的生殖隔离谱系,并提供了数百万年来形态停滞的有力证据(Michaux,1987,1989)。然而,谱系的准确识别和形态停滞都是间断平衡理论的重要元素,迄今为止,没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342 被确定为“物种”的化石形式被推断为等同于通过遗传标记确定的生殖隔离谱系,并提供了数百万年来形态停滞的有力证据(Michaux,1987,1989)。然而,谱系的准确识别和形态停滞都是间断平衡理论的重要元素,迄今为止,没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342 没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342 没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342
更新日期:2019-11-22
中文翻译:
新西兰橄榄壳的系统发育拓扑和时间与间断平衡一致
橄榄壳 (Amalda H. & A. Adams, 1853, Ancillariidae) 是海洋腹足类动物,主要分布在温带地区 (Olson, 1956),在太平洋和大西洋有近 100 种。物种多样性最高的是热带水域,南太平洋的物种很少(Kantor、Fedosov、Puillandre、Bonillo 和 Bouchet,2017 年)。Amalda 通常生活在沙质近岸环境中,在那里它们是捕食者,主要以双壳类动物为食(Powell,1979)。Amalda 的底栖软沉积物栖息地很容易形成化石,而在新西兰,Amalda 的化石记录可以追溯到始新世化石床 (Beu & Maxwell, 1990) 的 4500 万年前。在新西兰水域目前公认的七种现存物种中,有四种具有延伸到化石记录的谱系:A. mucronata Sowerby I, 1830, A. australis Sowerby I, 1830, A. depressa Sowerby II, 1859, 和 A. novaezelandiae Sowerby II, 1859 (Beu & Maxwell, 1990)。新西兰 Amalda 在关于形态进化模型的辩论中占有重要地位,因为它们被用作支持间断平衡的例子(Gould, 1991, 2009; Michaux, 1989)。被确定为“物种”的化石形式被推断为等同于通过遗传标记确定的生殖隔离谱系,并提供了数百万年来形态停滞的有力证据(Michaux,1987,1989)。然而,谱系的准确识别和形态停滞都是间断平衡理论的重要元素,迄今为止,没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342 australis Sowerby I, 1830, A. depressa Sowerby II, 1859, 和 A. novaezelandiae Sowerby II, 1859 (Beu & Maxwell, 1990)。新西兰 Amalda 在关于形态进化模型的辩论中占有重要地位,因为它们被用作支持间断平衡的例子(Gould,1991 年,2009 年;Michaux,1989 年)。被确定为“物种”的化石形式被推断为等同于通过遗传标记确定的生殖隔离谱系,并提供了数百万年来形态停滞的有力证据(Michaux,1987,1989)。然而,谱系的准确识别和形态停滞都是间断平衡理论的重要元素,迄今为止,没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342 australis Sowerby I, 1830, A. depressa Sowerby II, 1859, 和 A. novaezelandiae Sowerby II, 1859 (Beu & Maxwell, 1990)。新西兰 Amalda 在关于形态进化模型的辩论中占有重要地位,因为它们被用作支持间断平衡的例子(Gould,1991 年,2009 年;Michaux,1989 年)。被确定为“物种”的化石形式被推断为等同于通过遗传标记确定的生殖隔离谱系,并提供了数百万年来形态停滞的有力证据(Michaux,1987,1989)。然而,谱系的准确识别和形态停滞都是间断平衡理论的重要元素,迄今为止,没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342 和 A. novaezelandiae Sowerby II, 1859 (Beu & Maxwell, 1990)。新西兰 Amalda 在关于形态进化模型的辩论中占有重要地位,因为它们被用作支持间断平衡的例子(Gould,1991 年,2009 年;Michaux,1989 年)。被确定为“物种”的化石形式被推断为等同于通过遗传标记确定的生殖隔离谱系,并提供了数百万年来形态停滞的有力证据(Michaux,1987,1989)。然而,谱系的准确识别和形态停滞都是间断平衡理论的重要元素,迄今为止,没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342 和 A. novaezelandiae Sowerby II, 1859 (Beu & Maxwell, 1990)。新西兰 Amalda 在关于形态进化模型的辩论中占有重要地位,因为它们被用作支持间断平衡的例子(Gould,1991 年,2009 年;Michaux,1989 年)。被确定为“物种”的化石形式被推断为等同于通过遗传标记确定的生殖隔离谱系,并提供了数百万年来形态停滞的有力证据(Michaux,1987,1989)。然而,谱系的准确识别和形态停滞都是间断平衡理论的重要元素,迄今为止,没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342 新西兰 Amalda 在关于形态进化模型的辩论中占有重要地位,因为它们被用作支持间断平衡的例子(Gould,1991 年,2009 年;Michaux,1989 年)。被确定为“物种”的化石形式被推断为等同于通过遗传标记确定的生殖隔离谱系,并提供了数百万年来形态停滞的有力证据(Michaux,1987,1989)。然而,谱系的准确识别和形态停滞都是间断平衡理论的重要元素,迄今为止,没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342 新西兰 Amalda 在关于形态进化模型的辩论中占有重要地位,因为它们被用作支持间断平衡的例子(Gould,1991 年,2009 年;Michaux,1989 年)。被确定为“物种”的化石形式被推断为等同于通过遗传标记确定的生殖隔离谱系,并提供了数百万年来形态停滞的有力证据(Michaux,1987,1989)。然而,谱系的准确识别和形态停滞都是间断平衡理论的重要元素,迄今为止,没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342 被确定为“物种”的化石形式被推断为等同于通过遗传标记确定的生殖隔离谱系,并提供了数百万年来形态停滞的有力证据(Michaux,1987,1989)。然而,谱系的准确识别和形态停滞都是间断平衡理论的重要元素,迄今为止,没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342 被确定为“物种”的化石形式被推断为等同于通过遗传标记确定的生殖隔离谱系,并提供了数百万年来形态停滞的有力证据(Michaux,1987,1989)。然而,谱系的准确识别和形态停滞都是间断平衡理论的重要元素,迄今为止,没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342 没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342 没有收到:2018 年 11 月 5 日 | 修订日期:2019 年 8 月 10 日 | 接受:2019 年 8 月 20 日 DOI:10.1111/jzs.12342










































 京公网安备 11010802027423号
京公网安备 11010802027423号