
Seminatural Grasslands: An Emblematic Challenge for Nature Conservation in Protected Areas
 , , , ,
, , , , 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
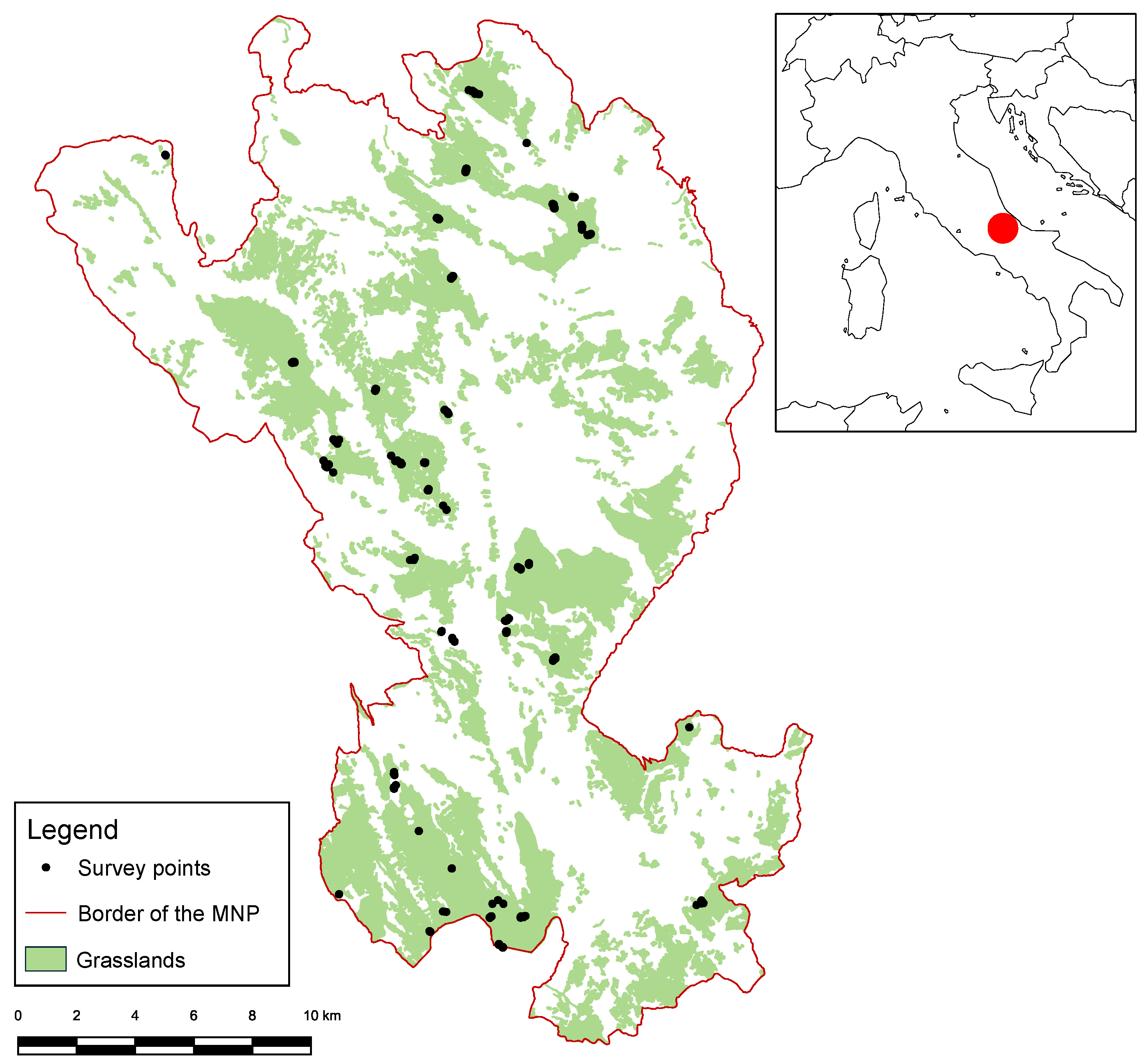
2.1. Study Area
2.2. Botanical Investigation
2.3. Pastoral Value and Biomass Analyses
2.4. The Development of a GEE App
2.5. Statistical Analyses
3. Results
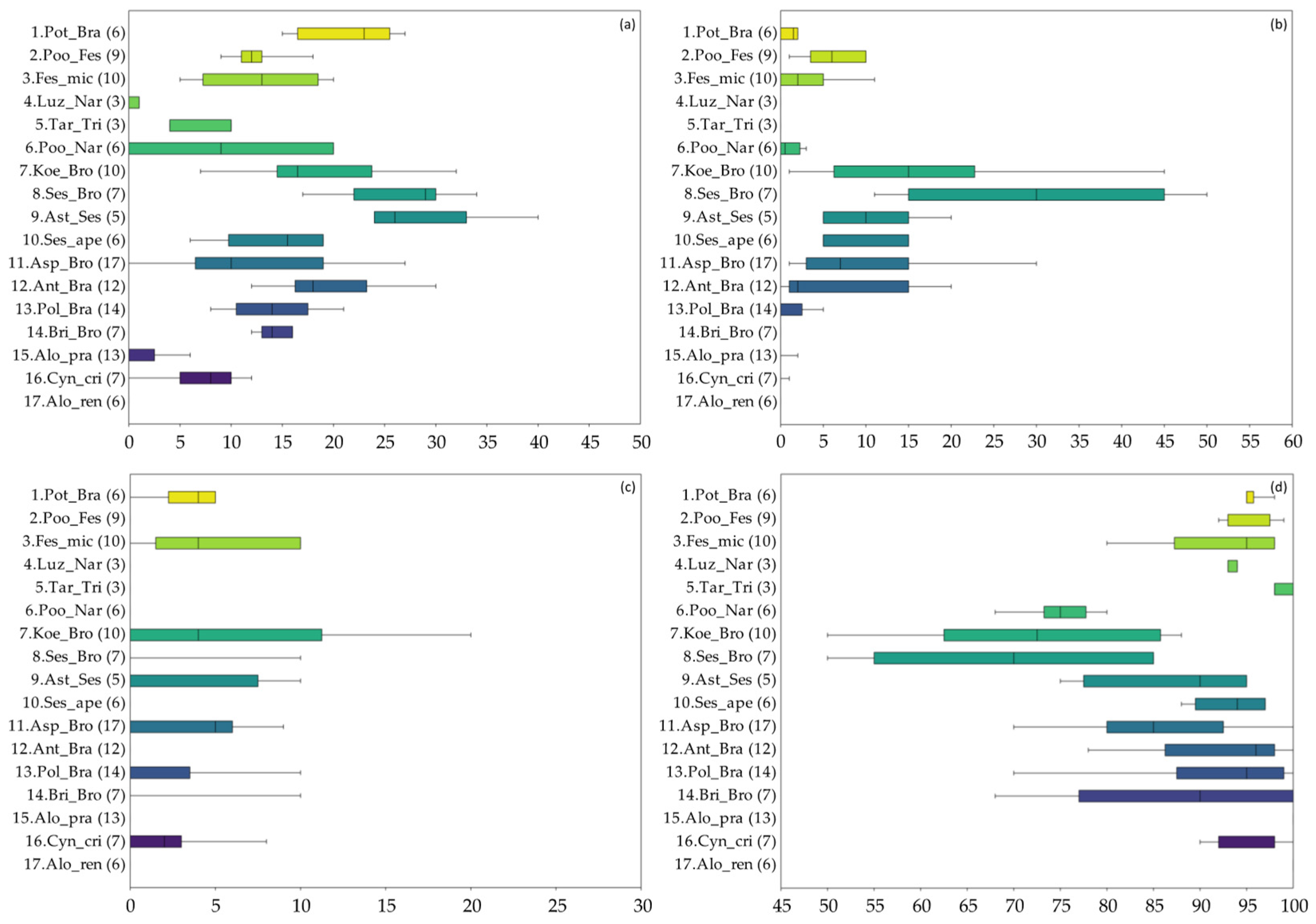
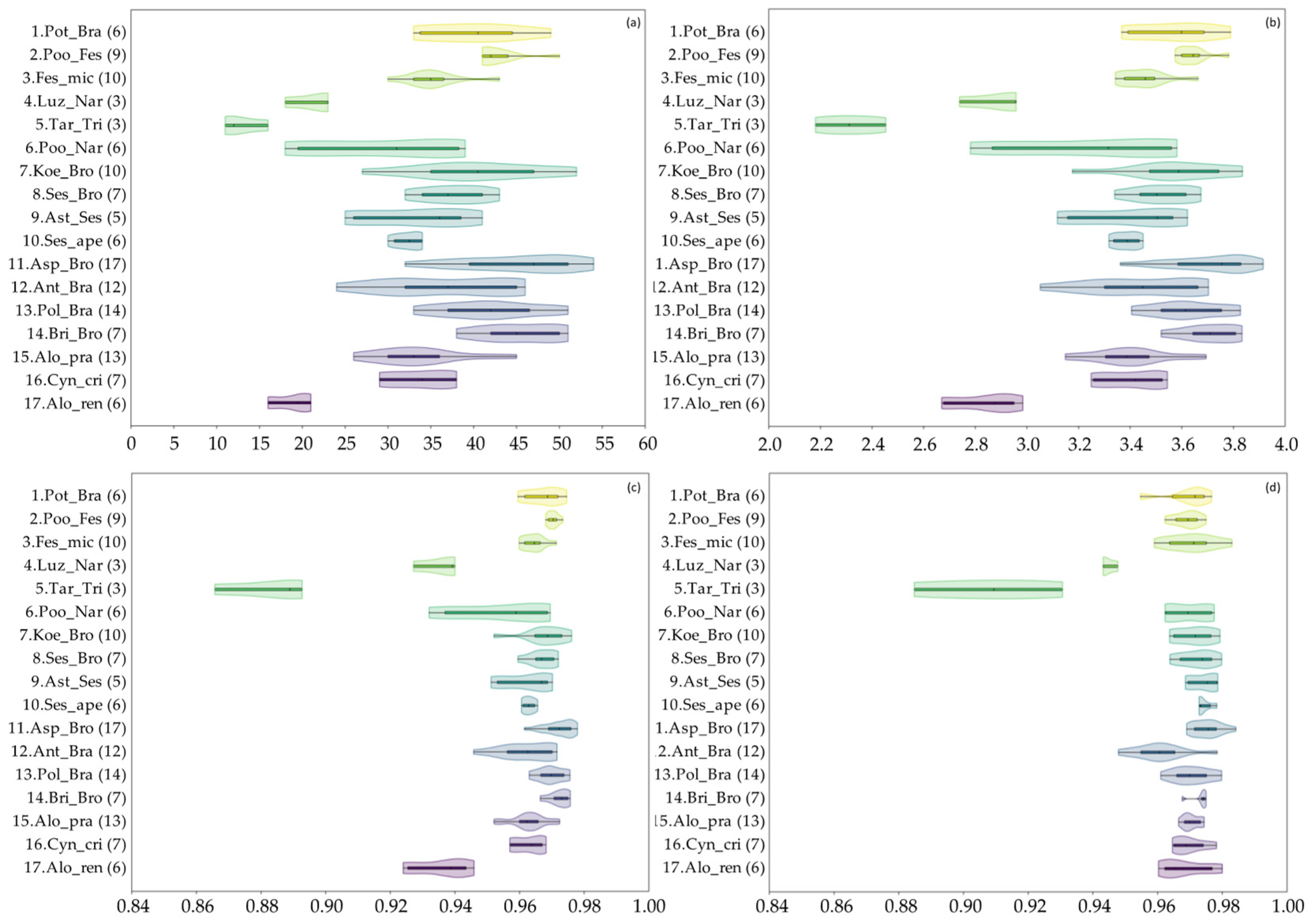
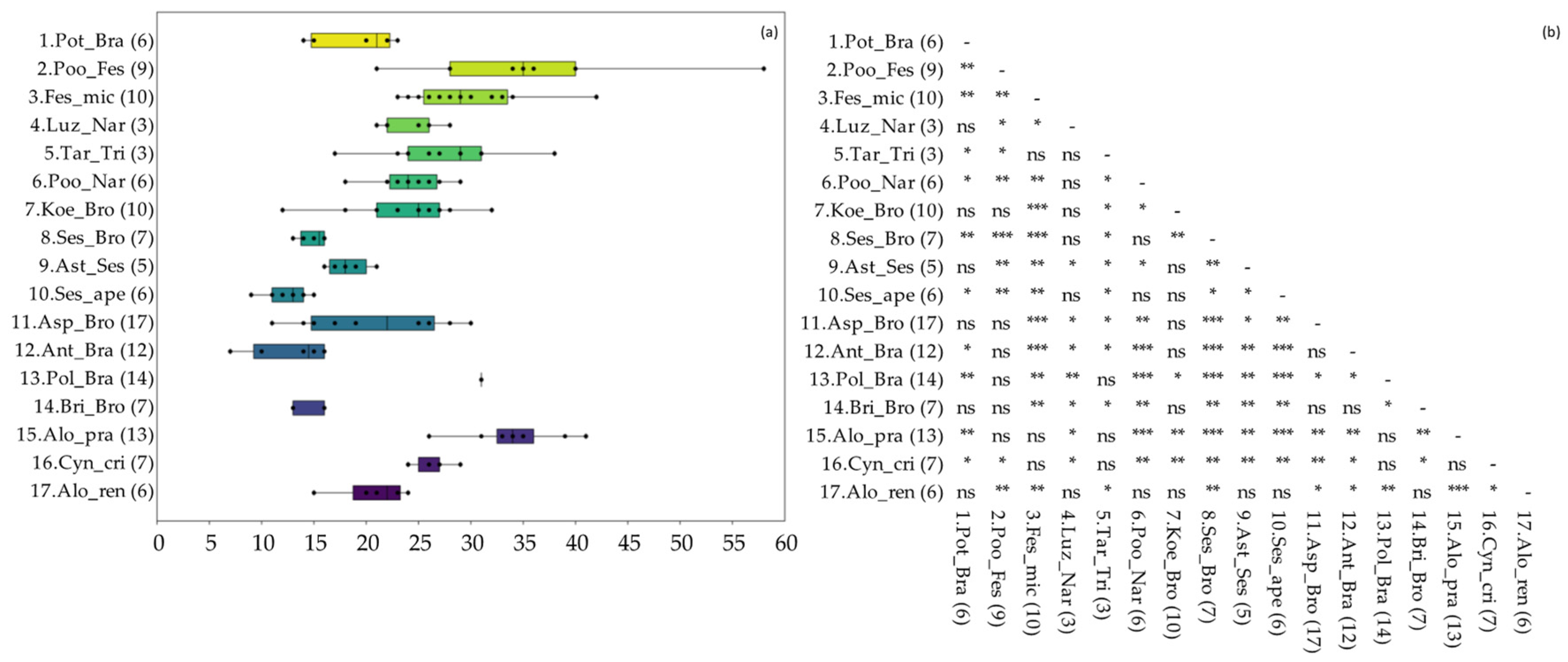
3.1. Vegetation Diversity of the Investigated Grasslands
3.2. Productivity and Nutritional Characteristics of the Grasslands
3.3. WebGIS-Based Tool for the Informed and Sustainable Use of Grasslands
4. Discussion
5. Final Remarks
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hejcman, M.; Hejcmanová, P.; Pavlů, V.; Beneš, J. Origin and History of Grasslands in Central Europe—A Review. Grass Forage Sci. 2013, 68, 345–363. [Google Scholar] [CrossRef]
- Tasser, E.; Tappeiner, U. Impact of Land Use Changes on Mountain Vegetation. Appl. Veg. Sci. 2002, 5, 173–184. [Google Scholar] [CrossRef]
- Navarro, L.M.; Proença, V.; Kaplan, J.O.; Pereira, H.M. Maintaining Disturbance-Dependent Habitats. In Rewilding European Landscapes; Pereira, H.M., Navarro, L.M., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 143–167. [Google Scholar] [CrossRef]
- Valkó, O.; Venn, S.; Żmihorski, M.; Biurrun, I.; Labadessa, R.; Loos, J. The Challenge of Abandonment for the Sustainable Management of Palaearctic Natural and Semi-Natural Grasslands. Hacquetia 2018, 17, 5–16. [Google Scholar] [CrossRef]
- Karlík, P.; Poschlod, P. History or Abiotic Filter: Which Is More Important in Determining the Species Composition of Calcareous Grasslands? Preslia 2009, 81, 321–340. [Google Scholar]
- Klimeš, L.; Dančak, M.; Hájek, M.; Jongepierová, I.; Kučera, T. Scale-dependent Biases in Species Counts in a Grassland. J. Veg. Sci. 2001, 12, 699–704. [Google Scholar] [CrossRef]
- Wilson, J.B.; Peet, R.K.; Dengler, J.; Pärtel, M. Plant Species Richness: The World Records. J. Veg. Sci. 2012, 23, 796–802. [Google Scholar] [CrossRef]
- Habel, J.C.; Dengler, J.; Janišová, M.; Török, P.; Wellstein, C.; Wiezik, M. European Grassland Ecosystems: Threatened Hotspots of Biodiversity. Biodivers. Conserv. 2013, 22, 2131–2138. [Google Scholar] [CrossRef]
- Dengler, J.; Biurrun, I.; Boch, S.; Dembicz, I.; Török, P. Grasslands of the Palaearctic Biogeographic Realm: Introduction and Synthesis. In Encyclopedia of the World’s Biomes; Elsevier: Amsterdam, The Netherlands, 2020; pp. 617–637. [Google Scholar] [CrossRef]
- Díaz, S.; Pascual, U.; Stenseke, M.; Martín-López, B.; Watson, R.T.; Molnár, Z.; Hill, R.; Chan, K.M.A.; Baste, I.A.; Brauman, K.A.; et al. Assessing Nature’s Contributions to People. Science 2018, 359, 270–272. [Google Scholar] [CrossRef] [PubMed]
- Landucci, F.; Panella, L.; Lucarini, D.; Gigante, D.; Donnini, D.; Kell, S.; Maxted, N.; Venanzoni, R.; Negri, V. A Prioritized Inventory of Crop Wild Relatives and Wild Harvested Plants of Italy. Crop Sci. 2014, 54, 1628–1644. [Google Scholar] [CrossRef]
- Tscharntke, T.; Klein, A.M.; Kruess, A.; Steffan-Dewenter, I.; Thies, C. Landscape Perspectives on Agricultural Intensification and Biodiversity—Ecosystem Service Management. Ecol. Lett. 2005, 8, 857–874. [Google Scholar] [CrossRef]
- Zulka, K.P.; Abensperg-Traun, M.; Milasowszky, N.; Bieringer, G.; Gereben-Krenn, B.-A.; Holzinger, W.; Hölzler, G.; Rabitsch, W.; Reischütz, A.; Querner, P.; et al. Species Richness in Dry Grassland Patches of Eastern Austria: A Multi-Taxon Study on the Role of Local, Landscape and Habitat Quality Variables. Agric. Ecosyst. Environ. 2014, 182, 25–36. [Google Scholar] [CrossRef]
- Carboni, M.; Dengler, J.; Mantilla-Contreras, J.; Venn, S.; Török, P. Conservation Value, Management and Restoration of Europe’S Semi-Natural Open Landscapes. Hacquetia 2015, 14, 5–17. [Google Scholar] [CrossRef]
- Bengtsson, J.; Bullock, J.M.; Egoh, B.; Everson, C.; Everson, T.; O’Connor, T.; O’Farrell, P.J.; Smith, H.G.; Lindborg, R. Grasslands—More Important for Ecosystem Services than You Might Think. Ecosphere 2019, 10, e02582. [Google Scholar] [CrossRef]
- Sollenberger, L.E.; Kohmann, M.M.; Dubeux, J.C.B.; Silveira, M.L. Grassland Management Affects Delivery of Regulating and Supporting Ecosystem Services. Crop Sci. 2019, 59, 441–459. [Google Scholar] [CrossRef]
- Isbell, F.; Calcagno, V.; Hector, A.; Connolly, J.; Harpole, W.S.; Reich, P.B.; Scherer-Lorenzen, M.; Schmid, B.; Tilman, D.; van Ruijven, J.; et al. High Plant Diversity Is Needed to Maintain Ecosystem Services. Nature 2011, 477, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Janssen, J.A.M.; Rodwell, J.S.; García Criado, M.; Gubbay, S.; Haynes, T.; Nieto, A.; Sanders, N.; Landucci, F.; Loidi, J.; Ssymank, A.; et al. European Red List of Habitats. Part 2. Terrestrial and Freshwater Habitats; Publications Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Gigante, D.; Acosta, A.T.R.; Agrillo, E.; Armiraglio, S.; Assini, S.; Attorre, F.; Bagella, S.; Buffa, G.; Casella, L.; Giancola, C.; et al. Habitat Conservation in Italy: The State of the Art in the Light of the First European Red List of Terrestrial and Freshwater Habitats. Rend. Fis. Acc. Lincei 2018, 29, 251–265. [Google Scholar] [CrossRef]
- Török, P.; Ambarlı, D.; Kamp, J.; Wesche, K.; Dengler, J. Step(Pe) up! Raising the Profile of the Palaearctic Natural Grasslands. Biodivers. Conserv. 2016, 25, 2187–2195. [Google Scholar] [CrossRef]
- EIONET Portal. Habitat Assessments at EU Biogeographical Level, Grasslands, Overall Assessment, 2013–2018. 2019. Available online: https://nature-art17.eionet.europa.eu/article17/habitat/progress/?period=5&group=Grasslands&conclusion=overall+assessment (accessed on 20 December 2023).
- Baur, B.; Cremene, C.; Groza, G.; Rakosy, L.; Schileyko, A.A.; Baur, A.; Stoll, P.; Erhardt, A. Effects of Abandonment of Subalpine Hay Meadows on Plant and Invertebrate Diversity in Transylvania, Romania. Biol. Conserv. 2006, 132, 261–273. [Google Scholar] [CrossRef]
- Díaz, S.; Lavorel, S.; McIntyre, S.; Falczuk, V.; Casanoves, F.; Milchunas, D.G.; Skarpe, C.; Rusch, G.; Sternberg, M.; Noy-Meir, I.; et al. Plant Trait Responses to Grazing—A Global Synthesis. Glob. Chang. Biol. 2007, 13, 313–341. [Google Scholar] [CrossRef]
- Cramer, V.; Hobbs, R.; Standish, R. What’s New about Old Fields? Land Abandonment and Ecosystem Assembly. Trends Ecol. Evol. 2008, 23, 104–112. [Google Scholar] [CrossRef]
- Halada, L.; Ružičková, H.; David, S.; Halabuk, A. Semi-Natural Grasslands under Impact of Changing Land Use during Last 30 Years: Trollio-Cirsietum Community in the Liptov Region (N Slovakia). Community Ecol. 2008, 9 (Suppl. 1), 115–123. [Google Scholar] [CrossRef]
- Vitasović Kosić, I.; Tardella, F.M.; Ruščić, M.; Catorci, A. Assessment of Floristic Diversity, Functional Composition and Management Strategy of North Adriatic Pastoral Landscape (Croatia). Pol. J. Ecol. 2011, 59, 765–776. [Google Scholar]
- Plieninger, T.; Hui, C.; Gaertner, M.; Huntsinger, L. The Impact of Land Abandonment on Species Richness and Abundance in the Mediterranean Basin: A Meta-Analysis. PLoS ONE 2014, 9, e98355. [Google Scholar] [CrossRef]
- Viciani, D.; Dell’Olmo, L.; Gabellini, A.; Gigante, D.; Lastrucci, L. Landscape Dynamics of Mediterranean Montane Grasslands over 60 Years and Implications for Habitats Conservation. A Case Study in the Northern Apennines (Italy). Landsc. Res. 2018, 43, 952–964. [Google Scholar] [CrossRef]
- Olmeda, C.; Šefferová, V.; Underwood, E.; Millan, L.; Gil, T.; Naumann, S. EU Action Plan to Maintain and Restore to Favourable Conservation Status the Habitat Type 6210 Semi-Natural Dry Grasslands and Scrubland Facies on Calcareous Substrates (Festuco-Brometalia) (*important Orchid Sites). 2019. Available online: https://www.ecologic.eu/17536 (accessed on 25 January 2024).
- van den Pol-van Dasselaar, A.; Bastiaansen-Aantjes, L.; Bogue, F.; O’Donovan, M.; Huyghe, C. Grassland Use in Europe: A Syllabus for Young Farmers; Éditions Quae: Versailles, France, 2019. [Google Scholar]
- Reinermann, S.; Asam, S.; Kuenzer, C. Remote Sensing of Grassland Production and Management—A Review. Remote Sens. 2020, 12, 1949. [Google Scholar] [CrossRef]
- Zarzycki, J.; Korzeniak, J.; Perzanowska, J. Impact of Land Use Changes on the Diversity and Conservation Status of the Vegetation of Mountain Grasslands (Polish Carpathians). Land 2022, 11, 252. [Google Scholar] [CrossRef]
- Fantinato, E.; Lorenzato, L.; Buffa, G. Patterns of Floral Resources and Pollination Interactions along Dry Grassland Succession. Plant Sociol. 2023, 60, 93–103. [Google Scholar] [CrossRef]
- Ceauşu, S.; Carver, S.; Verburg, P.H.; Kuechly, H.U.; Hölker, F.; Brotons, L.; Pereira, H.M. European Wilderness in a Time of Farmland Abandonment. In Rewilding European Landscapes; Pereira, H.M., Navarro, L.M., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 25–46. [Google Scholar] [CrossRef]
- Poschlod, P.; WallisDeVries, M.F. The Historical and Socioeconomic Perspective of Calcareous Grasslands—Lessons from the Distant and Recent Past. Biol. Conserv. 2002, 104, 361–376. [Google Scholar] [CrossRef]
- Pykälä, J.; Luoto, M.; Heikkinen, R.K.; Kontula, T. Plant Species Richness and Persistence of Rare Plants in Abandoned Semi-Natural Grasslands in Northern Europe. Basic Appl. Ecol. 2005, 6, 25–33. [Google Scholar] [CrossRef]
- Klimek, S.; Gen, R.; Kemmermann, A.; Hofmann, M.; Isselstein, J. Plant Species Richness and Composition in Managed Grasslands: The Relative Importance of Field Management and Environmental Factors. Biol. Conserv. 2007, 134, 559–570. [Google Scholar] [CrossRef]
- Schrautzer, J.; Jansen, D.; Breuer, M.; Nelle, O. Succession and Management of Calcareous Dry Grasslands in the Northern Franconian Jura, Germany. Tüxenia 2009, 29, 339–351. [Google Scholar]
- Tardella, F.M.; Catorci, A. Context-Dependent Effects of Abandonment vs. Grazing on Functional Composition and Diversity of Sub-Mediterranean Grasslands. Community Ecol. 2015, 16, 254–266. [Google Scholar] [CrossRef]
- Vitasović Kosić, I.; Tardella, F.M.; Grbeša, D.; Škvorc, Ž.; Catorci, A. Effects of Abandonment on the Functional Composition and Forage Nutritive Value of a North Adriatic Dry Grassland Community (Ćićarija, Croatia). Appl. Ecol. Environ. Res. 2014, 12, 285–299. [Google Scholar] [CrossRef]
- Brooks, T.M.; Mittermeier, R.A.; Da Fonseca, G.A.B.; Gerlach, J.; Hoffmann, M.; Lamoreux, J.F.; Mittermeier, C.G.; Pilgrim, J.D.; Rodrigues, A.S.L. Global Biodiversity Conservation Priorities. Science 2006, 313, 58–61. [Google Scholar] [CrossRef]
- Schmeller, D.S.; Gruber, B.; Budrys, E.; Framsted, E.; Lengyel, S.; Henle, K. National Responsibilities in European Species Conservation: A Methodological Review. Conserv. Biol. 2008, 22, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Schmeller, D.; Maier, A.; Evans, D.; Henle, K. National Responsibilities for Conserving Habitats—A Freely Scalable Method. NC 2012, 3, 21–44. [Google Scholar] [CrossRef]
- Schmeller, D.S.; Evans, D.; Lin, Y.-P.; Henle, K. The National Responsibility Approach to Setting Conservation Priorities—Recommendations for Its Use. J. Nat. Conserv. 2014, 22, 349–357. [Google Scholar] [CrossRef]
- Orsenigo, S.; Montagnani, C.; Fenu, G.; Gargano, D.; Peruzzi, L.; Abeli, T.; Alessandrini, A.; Bacchetta, G.; Bartolucci, F.; Bovio, M.; et al. Red Listing Plants under Full National Responsibility: Extinction Risk and Threats in the Vascular Flora Endemic to Italy. Biol. Conserv. 2018, 224, 213–222. [Google Scholar] [CrossRef]
- Martinasso, B.; Lombardi, G. Remote Sensing in NW Italian Alps for Pastoral Inventory and Improvement of Grassland Management. In Land Use Systems in Grassland Dominated Regions; Luscher, A., Jeangros, B., Kessler, W., Huguenin, O., Lobsiger, M., Millar, N., Suter, D., Eds.; Grassland Science in Europe; ETH Zentrum: Luzern, Switzerland, 2004; Volume 9, pp. 317–319. [Google Scholar]
- Pino-Del-Carpio, A.; Ariño, A.H.; Villarroya, A.; Puig, J.; Miranda, R. The Biodiversity Data Knowledge Gap: Assessing Information Loss in the Management of Biosphere Reserves. Biol. Conserv. 2014, 173, 74–79. [Google Scholar] [CrossRef]
- Urbano, F.; Viterbi, R.; Pedrotti, L.; Vettorazzo, E.; Movalli, C.; Corlatti, L. Enhancing Biodiversity Conservation and Monitoring in Protected Areas through Efficient Data Management. Environ. Monit. Assess. 2024, 196, 12. [Google Scholar] [CrossRef] [PubMed]
- Conde, D.A.; Staerk, J.; Colchero, F.; Da Silva, R.; Schöley, J.; Baden, H.M.; Jouvet, L.; Fa, J.E.; Syed, H.; Jongejans, E.; et al. Data Gaps and Opportunities for Comparative and Conservation Biology. Proc. Natl. Acad. Sci. USA 2019, 116, 9658–9664. [Google Scholar] [CrossRef] [PubMed]
- Demangeot, J. La Montagne de La Majella [The Mountains of Majella]. In Geomorphologie des Abruzzes Adriatiques; Centre National de la Recherche Scientifique (CNRS): Paris, France, 1965; pp. 165–192. [Google Scholar]
- Liberatoscioli, E.; Boscaino, G.; Agostini, S.; Garzarella, A.; Patacca Scandone, E. The Majella National Park: An Aspiring UNESCO Geopark. Geosciences 2018, 8, 256. [Google Scholar] [CrossRef]
- Di Martino, L.; Del Vecchio, S.; Di Cecco, V.; Di Santo, M.; Stanisci, A.; Frattaroli, A.R. The Role of GA3 in the Germination Process of High-Mountain Endemic and Threatened Species: Leontopodium nivalePinguicula Fiorii and Soldanella Minima Subsp. Samnitica (Central Apennines, Italy). Plant Biosyst.—Int. J. Deal. All Asp. Plant Biol. 2014, 148, 1231–1238. [Google Scholar] [CrossRef]
- ISPRA. Meteo 1991–2020. 2023. Available online: https://www.isprambiente.gov.it/it/banche-dati/banche-dati-folder/clima-e-meteo/meteo (accessed on 15 December 2023).
- Conti, F.; Ciaschetti, G.; Di Martino, L.; Bartolucci, F. An Annotated Checklist of the Vascular Flora of Majella National Park (Central Italy). Phytotaxa 2019, 412, 1–90. [Google Scholar] [CrossRef]
- Rossi, G.; Montagnani, C.; Gargano, D.; Peruzzi, L.; Abeli, T.; Ravera, S.; Cogoni, A.; Fenu, G.; Magrini, S.; Gennai, M.; et al. Lista Rossa Della Flora Italiana. 1. Policy Species e Altre Specie Minacciate. 2013. Available online: https://www.iucn.it/pdf/Comitato_IUCN_Lista_Rossa_della_flora_italiana_policy_species.pdf (accessed on 3 January 2024).
- Rossi, G.; Orsenigo, S.; Gargano, D.; Montagnani, C.; Peruzzi, L.; Fenu, G.; Abeli, T.; Alessandrini, A.; Astuti, G.; Bacchetta, G.; et al. Lista Rossa Della Flora Italiana. 2 Endemiti e Altre Specie Minacciate. 2020. Available online: https://www.iucn.it/pdf/LISTAROSSAvol-2-FLORAITALIANA.pdf (accessed on 20 December 2023).
- European Commission. Council Directive 92/43/EEC of 21 May 1992 on the Conservation of Natural Habitats and of Wild Fauna and Flora. 1992. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A01992L0043-20130701 (accessed on 13 March 2024).
- European Commission. Interpretation Manual of European Union Habitats. EUR 28. 2013. Available online: https://circabc.europa.eu/ui/group/3f466d71-92a7-49eb-9c63-6cb0fadf29dc/library/37d9e6d9-b7de-42ce-b789-622e9741b68f/details (accessed on 15 December 2023).
- Biondi, E.; Blasi, C.; Burrascano, S.; Casavecchia, S.; Copiz, R.; Del Vico, E.; Galdenzi, D.; Gigante, D.; Lasen, C.; Spampinato, G.; et al. Manuale Italiano Di Interpretazione Degli Habitat Della Direttiva 92/43/CEE. 2009. Available online: http://vnr.unipg.it/habitat/ (accessed on 10 December 2023).
- Frattaroli, A.-R.; Di Martino, L.; Di Cecco, V.; Catoni, R.; Varoni, L.; Di Santo, M.; Gratani, L. Seed Germination Capability of Four Endemic Species in the Central Apennines (Italy): Relationships with Seed Size. LAZA 2013, 34, 43–53. [Google Scholar] [CrossRef]
- Di Cecco, V.; Di Santo, M.; Di Musciano, M.; Manzi, A.; Di Cecco, M.; Ciaschetti, G.; Marcantonio, G.; Di Martino, L. The Majella National Park: A Case Study for the Conservation of Plant Biodiversity in the Italian Apennines. IB 2020, 10, 1–24. [Google Scholar] [CrossRef]
- Regione Abruzzo. Modello Digitale Del Terreno—Risoluzione 10x10 Metri. 2015. Available online: http://opendata.regione.abruzzo.it/opendata/Modello_digitale_del_terreno_risoluzione_10x10_metri (accessed on 28 December 2023).
- Società Geologica Italiana. Abruzzo. Guida Geologica; BeMa Editrice: Milano, Italy, 2003. [Google Scholar]
- Patacca, E.; Scandone, P. Geological Map of the Majella Mountain, 1:25,000. 2021. Available online: https://www.isprambiente.gov.it/it/attivita/suolo-e-territorio/cartografia/carta-geologica-della-majella/majella_small.jpg/@@images/b984c8c7-89cb-43a2-8640-3ebebfaf37a5.jpeg (accessed on 15 December 2023).
- Pesaresi, S.; Biondi, E.; Casavecchia, S. Bioclimates of Italy. J. Maps 2017, 13, 955–960. [Google Scholar] [CrossRef]
- Pirone, G.; Frattaroli, A.R.; Ciaschetti, G. Le Serie Di Vegetazione Della Regione Abruzzo. In La Vegetazione d’Italia; Blasi, C., Ed.; Palombi & Partner S.r.l.: Roma, Italy; pp. 311–336.
- Braun-Blanquet, J. Plant Sociology. The Study of Plant Communities. Translated, Revised and Edited by George D. Fuller and Herry S. Conard. Authorized English Translations of ‘Pflanzensoziologie’, 1st ed.; McGraw-Hill Book: New York, NY, USA; London, UK, 1932. [Google Scholar]
- Braun-Blanquet, J. Fitosociologia; Blume: Madrid, Spain, 1964. [Google Scholar]
- Dengler, J.; Chytrý, M.; Ewald, J. Phytosociology. In Encyclopedia of Ecology; Jørgensen, S.E., Fath, B.D., Eds.; General Ecology; Elsevier: Oxford, UK, 2008; Volume 4, pp. 2767–2779. [Google Scholar]
- Biondi, E. Phytosociology Today: Methodological and Conceptual Evolution. Plant Biosyst.—Int. J. Deal. All Asp. Plant Biol. 2011, 145 (Suppl. 1), 19–29. [Google Scholar] [CrossRef]
- Gigante, D.; Attorre, F.; Venanzoni, R.; Acosta, A.T.R.; Agrillo, E.; Aleffi, M.; Alessi, N.; Allegrezza, M.; Angelini, P.; Angiolini, C.; et al. A Methodological Protocol for Annex I Habitats Monitoring: The Contribution of Vegetation Science. Plant Sociol. 2016, 53, 77–87. [Google Scholar] [CrossRef]
- Chytrý, M.; Otýpková, Z. Plot Sizes Used for Phytosociological Sampling of European Vegetation. J. Veg. Sci. 2003, 14, 563–570. [Google Scholar] [CrossRef]
- Pignatti, S. Flora d’Italia. Volume 1; Seconda edizione in 4 volumi; Edagricole: Milano, Italy, 2017. [Google Scholar]
- Pignatti, S. Flora d’Italia. Volume 2; Seconda edizione in 4 volumi; Edagricole: Milano, Italy, 2017. [Google Scholar]
- Pignatti, S. Flora d’Italia. Volume 3; Seconda edizione in 4 volumi; Edagricole: Milano, Italy, 2018. [Google Scholar]
- AA.VV. Portal to the Flora of Italy. 2024. Available online: http://dryades.units.it/floritaly (accessed on 10 January 2024).
- Biondi, E.; Guitian, J.; Allegrezza, M.; Ballelli, S. Su Alcuni Pascoli a Sesleria Apennina Ujhelyi Nell’Appennino Centrale. Doc. Phytosoc. N.S. 1988, 11, 417–422. [Google Scholar]
- Biondi, E.; Allegrezza, M.; Frattaroli, A.R. Inquadramento Fitosociologico Di Alcune Formazioni Pascolive Dell’ Appennino Abruzzese-Molisano. Doc. Phytosoc. N.S. 1992, 14, 195–210. [Google Scholar]
- Biondi, E.; Allegrezza, M.; Ballelli, S.; Calandra, R.; Crescente, M.F.; Frattaroli, A.R.; Gratani, L.; Rossi, A.; Taffetani, F. Indagini per Una Cartografia Fitoecologica Dell’altipiano Di Campo Imperatore (Gran Sasso d’Italia). Boll. A.I.C. 1992, 86, 85–98. [Google Scholar]
- Biondi, E.; Ballelli, S. Le Praterie Del Monte Coscerno e Monte Di Civitella (Appennino Umbro- Marchigiano—Italia Centrale). Fitosociologia 1995, 30, 91–121. [Google Scholar]
- Biondi, E.; Ballelli, S.; Allegrezza, M.; Taffetani, F.; Frattaroli, A.R.; Guitian, J.; Zuccarello, V. La Vegetazione Di Campo Imperatore (Gran Sasso d’Italia). Braun-Blanquetia 1999, 16, 53–115. [Google Scholar]
- Manzi, A. La Gestione Dei Pascoli Montani in Abruzzo e La Società Delle Erbe Seconde Di Pescasseroli Ed Opi. Arch. Bot. Ital. 1990, 66, 129–142. [Google Scholar]
- Lucchese, F.; Persia, G.; Pignatti, S. I Pascoli a Bromus Erectus Hudson Dell’ Appennino Laziale. Fitosociologia 1995, 30, 145–180. [Google Scholar]
- Petriccione, B.; Persia, G. Prodromo Delle Praterie Di Altitudine Degli Appennini Su Calcare (Classe Festuco-Seslerietea). Atti dei Convegni Lincei. “La vegetazione italiana” 1995, 115, 361–389. [Google Scholar]
- Pirone, G. Il Paesaggio Vegetale Di Rivisondoli. Aspetti Della Flora e Della Vegetazione. In Azienda Autonoma Soggiorno e Turismo Rivisondoli; Edigrafital S.p.a.: Teramo, Italy, 1997. [Google Scholar]
- Stanisci, A.; Pelino, G.; Blasi, C. Gli Arbusteti Altomontani Dell’Appennino Centrale e Meridionale. Fitosociologia 1997, 34, 3–46. [Google Scholar]
- Ubaldi, D.; Filz, W.; De Santo, G.; Di Cecco, M. Osservazioni Sulla Vegetazione Della Riserva Majella Orientale. Micol. Vegetaz. Medit. 1998, 13, 177–192. [Google Scholar]
- Allegrezza, M. Vegetazione e Paesaggio Vegetale Della Dorsale Del Monte San Vicino (Appennino Centrale). Fitosociologia 2003, 40, 3–118. [Google Scholar]
- Stanisci, A.; Pelino, G.; Blasi, C. Vascular Plant Diversity and Climate Change in the Alpine Belt of the Central Apennines (Italy). Biodivers. Conserv. 2005, 14, 1301–1318. [Google Scholar] [CrossRef]
- Ciaschetti, G.; Pirone, A.R.; Frattaroli, A.R.; Corbetta, F. La Vegetazione Del Piano Di Pezza (Parco Naturale Regionale “Sirente-Velino”—Italia Centrale). Fitosociologia 2006, 43, 67–84. [Google Scholar]
- Blasi, C.; Burrascano, S.; Del Vico, E.; Di Pietro, R.; Iocchi, M.; Rosati, L. Cynosurion Cristati Grasslands in the Central Apennines (Tyrrhenian Sector): A Phytosociological Survey in the Lepini and Prenestini Mountains. Plant Biosyst.—Int. J. Deal. All Asp. Plant Biol. 2009, 143 (Suppl. 1), S69–S77. [Google Scholar] [CrossRef]
- Catorci, A.; Cesaretti, S.; Gatti, R.; Ottaviani, G. Abiotic and Biotic Changes Due to Spread of Brachypodium Genuense (DC.) Roem. & Schult, in Sub-Mediterranean Meadows. Community Ecol. 2011, 12, 117–125. [Google Scholar]
- Di Pietro, R.; Conte, A.L.; Iamonico, D. La Vegetazione Di San Martino D’Agri (Monografia e Carta Della Vegetazione); Grafica Mente, Sant’Arcangelo (PZ): Comune di S. Martino D’Agri, Italy, 2014. [Google Scholar]
- Di Pietro, R.; Theurillat, J.-P.; Capelo, J.; Fernández-González, F.; Terzi, M.; Čarni, A.; Mucina, L. Nomenclature and Syntaxonomic Notes on Some High-Rank Syntaxa of the European Grassland Vegetation. Lazaroa 2015, 36, 79–106. [Google Scholar] [CrossRef]
- Filibeck, G.; Sperandii, M.G.; Bazzichetto, M.; Mancini, L.D.; Rossini, F.; Cancellieri, L. Exploring the Drivers of Vascular Plant Richness at Very Fine Spatial Scale in Sub-Mediterranean Limestone Grasslands (Central Apennines, Italy). Biodivers. Conserv. 2019, 28, 2701–2725. [Google Scholar] [CrossRef]
- Biondi, E.; Blasi, C. Prodromo Della Vegetazione Italiana. 2015. Available online: http://www.prodromo-vegetazione-italia.org (accessed on 28 December 2023).
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.-P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; García, R.G.; et al. Vegetation of Europe: Hierarchical Floristic Classification System of Vascular Plant, Bryophyte, Lichen, and Algal Communities. Appl. Veg. Sci. 2016, 19 (S1), 3–264. [Google Scholar] [CrossRef]
- Theurillat, J.; Willner, W.; Fernández-González, F.; Bültmann, H.; Čarni, A.; Gigante, D.; Mucina, L.; Weber, H. International Code of Phytosociological Nomenclature. 4th Edition. Appl. Veg. Sci. 2021, 24, e12491. [Google Scholar] [CrossRef]
- Biondi, E.; Burrascano, S.; Casavecchia, S.; Copiz, R.; Del Vico, E.; Gigante, D.; Lasen, C.; Spampinato, G.; Venanzoni, R.; Zivkovic, L.; et al. Diagnosis and Syntaxonomic Interpretation of Annex I Habitats (Dir. 92/43/EEC) in Italy at the Alliance Level. Plant Sociol. 2012, 49, 5–37. [Google Scholar] [CrossRef]
- Angelini, P. Manuali per il Monitoraggio di Specie e Habitat di Interesse Comunitario (Direttiva 92/43/CEE) in Italia: Habitat; ISPRA: Roma, Italy, 2016. [Google Scholar]
- Roggero, P.P.; Bagella, S.; Farina, R. Un Archivio Dati Di Indici Specifici per La Valutazione Integrata Del Valore Pastorale. Riv. Agron. 2002, 36, 149–156. [Google Scholar]
- Reiné, R.; Ascaso, J.; Barrantes, O. Nutritional Quality of Plant Species in Pyrenean Hay Meadows of High Diversity. Agronomy 2020, 10, 883. [Google Scholar] [CrossRef]
- Daget, P.; Poissonet, T. Analyse Phytologique Des Praisies; Centre National de la Recherche Scientifique, Centre d’Études Phytosociologiques et Écologiques: Montpellier, France, 1969. [Google Scholar]
- Daget, P.; Poissonet, T. Un Procédé d’estimation de La Valeur Pastorale Des Pâturages. Fourrages 1972, 49, 31–39. [Google Scholar]
- Argenti, G.; Lombardi, G. The Pasture-Type Approach for Mountain Pasture Description and Management. Ital. J. Agron. 2012, 7, e39. [Google Scholar] [CrossRef]
- Argenti, G.; Bianchetto, E.; Ferretti, F. Proposal of a Simplified Method for Pastoral Value Assessment inside Forest Planning. Ann. Silvic. Res. 2017, 41, 67–73. [Google Scholar] [CrossRef]
- Pittarello, M.; Lonati, M.; Gorlier, A.; Perotti, E.; Probo, M.; Lombardi, G. Plant Diversity and Pastoral Value in Alpine Pastures Are Maximized at Different Nutrient Indicator Values. Ecol. Indic. 2018, 85, 518–524. [Google Scholar] [CrossRef]
- Bagella, S. Valore Pastorale Delle Associazioni Vegetali: Un Esempio Di Applicazione neIl’Appennino Umbro-Marchigiano (Italia). Fitosociologia 2001, 38, 153–165. [Google Scholar]
- Bagella, S.; Roggero, P.P. Integrating Phytosociological and Agronomic Analysis to Support the Sustainable Management of Mediterranean Grasslands. Fitosociologia 2004, 41 (Suppl. 1), 101–107. [Google Scholar]
- Bencivenga, M.; Panitti, M.; Taddei, E. Indagine Fitosociologica Nei Prato-Pascoli Del Monte Fausola (Rieti) Sottoposti a Concimazioni Diverse. Nota II—Frequenza e Peso Unitario Delle Specie Più Diffuse. Riv. Agron. 1992, 26, 389–399. [Google Scholar]
- Scotton, M.; Rodaro, P.; Ziliotto, U. Analisi Fitosociologica e Valutazione Agronomica Di Pascoli Subalpini. Stud. Geobotanica. Int. J. 1996, 15, 51–72. [Google Scholar]
- Delpech, R. Criteres de Jugement de La Valeur Agronomique Des Praires. Fourrages 1960, 4, 83–98. [Google Scholar]
- Cavallero, A.; Talamucci, P.; Grignani, C.; Reyneri, A.; Ziliotto, U.; Scotton, M.; Bianchi, A.A.; Santilocchi, R.; Basso, E.; Postiglione, L.; et al. Caratterizzazione Della Dinamica Produttiva Di Pascoli Naturali Italiani. Riv. Agron. 1992, 26, 325–343. [Google Scholar]
- Pacioni, G.; Veri, L. L’ambiente Naturale Dei Monti Simbruini: Contributi Alla Conoscenza. Atti Del Convegno: Per Un’operazione Integrata Di Sviluppo, Il Caso Simbruini. Altipiani Di Arcinazzo, 7 December 1985. [Google Scholar]
- Scoppola, A.; Pelosi, M. I Pascoli Della Riserva Naturale Del Monte Rufeno (Viterbo, Italia Centrale). Fitosociologia 1995, 30, 123–143. [Google Scholar]
- D’Ottavio, P.; Scotton, M.; Ziliotto, U. Pastoral Value of Mountain Pastures of Monti Sibillini (Central Apennines, Italy) Grazed by Sheep. Options Méditerranéennes 2005, 67, 93–98. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC; Association of Official Analytical Chemists (AOAC) International: Washington, DC, USA, 1995. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Zanaga, D.; Van De Kerchove, R.; Daems, D.; De Keersmaecker, W.; Brockmann, C.; Kirches, G.; Wevers, J.; Cartus, O.; Santoro, M.; Fritz, S.; et al. ESA WorldCover 10 m 2021 V200. 2022. Available online: https://doi.org/10.5281/ZENODO.7254221 (accessed on 13 March 2024).
- Tucker, C.J. Red and Photographic Infrared Linear Combinations for Monitoring Vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Chen, J.; Jönsson, P.; Tamura, M.; Gu, Z.; Matsushita, B.; Eklundh, L. A Simple Method for Reconstructing a High-Quality NDVI Time-Series Data Set Based on the Savitzky–Golay Filter. Remote Sens. Environ. 2004, 91, 332–344. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. 2021. Available online: https://www.R-project.org/ (accessed on 13 March 2024).
- De Cáceres, M.; Legendre, P. Associations between Species and Groups of Sites: Indices and Statistical Inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- De Cáceres, M.; Sol, D.; Lapiedra, O.; Legendre, P. A Framework for Estimating Niche Metrics Using the Resemblance between Qualitative Resources. Oikos 2011, 120, 1341–1350. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Magurran, A.E. Ecological Diversity and Its Measurement; Springer: Dordrecht, The Netherlands, 1988. [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. An Analysis of Variance Test for Normality (Complete Samples). Biometrika 1965, 52, 591. [Google Scholar] [CrossRef]
- Harper, W.V. Reduced Major Axis Regression. In Wiley StatsRef: Statistics Reference Online; Kenett, R.S., Longford, N.T., Piegorsch, W.W., Ruggeri, F., Eds.; Wiley: Hoboken, NJ, USA, 2016; pp. 1–6. [Google Scholar] [CrossRef]
- Cavallero, A.; Aceto, P.; Gorlier, A.; Lombardi, G.; Lonati, M.; Martinasso, B.; Tagliatori, C. I Tipi Pastorali Delle Alpi Piemontesi; Gruppo Perdisa Editore: Bologna, Italy, 2007. [Google Scholar]
- Targetti, S.; Staglianò, N.; Messeri, A.; Argenti, G. A State-and-Transition Approach to Alpine Grasslands under Abandonment. iForest 2010, 3, 44–51. [Google Scholar] [CrossRef]
- Purvis, A.; Hector, A. Getting the Measure of Biodiversity. Nature 2000, 405, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Llopis-Belenguer, C.; Blasco-Costa, I.; Balbuena, J.A. Biodiversidad En elSiglo XXI: Más Allá Del Conteo Deespecies. Mètode 2019, 9, 11186. [Google Scholar] [CrossRef]
- Hillebrand, H.; Blasius, B.; Borer, E.T.; Chase, J.M.; Downing, J.A.; Eriksson, B.K.; Filstrup, C.T.; Harpole, W.S.; Hodapp, D.; Larsen, S.; et al. Biodiversity Change Is Uncoupled from Species Richness Trends: Consequences for Conservation and Monitoring. J. Appl. Ecol. 2018, 55, 169–184. [Google Scholar] [CrossRef]
- Blasi, C.; Di Pietro, R.; Pelino, G. The Vegetation of Alpine Belt Karst-Tectonic Basins in the Central Apennines (Italy). Plant Biosyst.—Int. J. Deal. All Asp. Plant Biol. 2005, 139, 357–385. [Google Scholar] [CrossRef]
- Kruess, A.; Tscharntke, T. Grazing Intensity and the Diversity of Grasshoppers, Butterflies, and Trap-Nesting Bees and Wasps. Conserv. Biol. 2002, 16, 1570–1580. [Google Scholar] [CrossRef]
- Fantinato, E.; Sonkoly, J.; Török, P.; Buffa, G. Patterns of Pollination Interactions at the Community Level Are Related to the Type and Quantity of Floral Resources. Funct. Ecol. 2021, 35, 2461–2471. [Google Scholar] [CrossRef]
- Manzi, A. Storia Dell’ambiente Nell’Appennino Centrale: La Trasformazione Della Natura in Abruzzo Dall’ultima Glaciazione Ai Nostri Giorni; Ed. Meta (Treglio): Chieti, Italy, 2012. [Google Scholar]
- Laiolo, P.; Tella, J.L. Fate of Unproductive and Unattractive Habitats: Recent Changes in Iberian Steppes and Their Effects on Endangered Avifauna. Environ. Conserv. 2006, 33, 223–232. [Google Scholar] [CrossRef]
- Manzi, A.; Manzi, G. Pastori, Lanaioli e Contadini. La Pastorizia e La Lavorazione Della Lana Nel Versante Orientale Della Maiella; Ed. Meta (Treglio): Chieti, Italy, 2007. [Google Scholar]
- Colecchia, A. Paesaggi Storici Agro-Silvo-Pastorali Nell’Abruzzo Interno: Dall’analisi Multidisciplinare al Recupero Delle Identità Culturali Locali. Il Capitale Cult.—Stud. Value Cult. Herit. 2015, 12, 743–771. [Google Scholar]
- Micati, E.; Manzi, A.; Di Martino, L.; Monaco, M. Il Paesaggio Agro-Pastorale Del Parco Nazionale Della Majella; Edizioni Parco Nazionale della Majella: Sulmona, Italy, 2019. [Google Scholar]
- Ramírez-Restrepo, C.A.; Barry, T.N. Alternative Temperate Forages Containing Secondary Compounds for Improving Sustainable Productivity in Grazing Ruminants. Anim. Feed Sci. Technol. 2005, 120, 179–201. [Google Scholar] [CrossRef]
- Macheboeuf, D.; Coudert, L.; Bergeault, R.; Lalière, G.; Niderkorn, V. Screening of Plants from Diversified Natural Grasslands for Their Potential to Combine High Digestibility, and Low Methane and Ammonia Production. Animal 2014, 8, 1797–1806. [Google Scholar] [CrossRef] [PubMed]
- Pirhofer-Walzl, K.; Søegaard, K.; Høgh-Jensen, H.; Eriksen, J.; Sanderson, M.A.; Rasmussen, J.; Rasmussen, J. Forage Herbs Improve Mineral Composition of Grassland Herbage. Grass Forage Sci. 2011, 66, 415–423. [Google Scholar] [CrossRef]
- Hammond, K.J.; Humphries, D.J.; Westbury, D.B.; Thompson, A.; Crompton, L.A.; Kirton, P.; Green, C.; Reynolds, C.K. The Inclusion of Forage Mixtures in the Diet of Growing Dairy Heifers: Impacts on Digestion, Energy Utilisation, and Methane Emissions. Agric. Ecosyst. Environ. 2014, 197, 88–95. [Google Scholar] [CrossRef]
- Farruggia, A.; Martin, B.; Baumont, R.; Prache, S.; Doreau, M.; Hoste, H.; Durand, D. Quels Intérêts de La Diversité Floristique Des Prairies Permanentes Pour Les Ruminants et Les Produits Animaux? INRA Prod. Anim. 2008, 21, 181–200. [Google Scholar] [CrossRef]
- Van Vliet, S.; Provenza, F.D.; Kronberg, S.L. Health-Promoting Phytonutrients Are Higher in Grass-Fed Meat and Milk. Front. Sustain. Food Syst. 2021, 4, 555426. [Google Scholar] [CrossRef]
- Corazza, M.; Tardella, F.M.; Ferrari, C.; Catorci, A. Tall Grass Invasion After Grassland Abandonment Influences the Availability of Palatable Plants for Wild Herbivores: Insight into the Conservation of the Apennine Chamois Rupicapra Pyrenaica Ornata. Environ. Manag. 2016, 57, 1247–1261. [Google Scholar] [CrossRef]
- De Simone, W.; Allegrezza, M.; Frattaroli, A.R.; Montecchiari, S.; Tesei, G.; Zuccarello, V.; Di Musciano, M. From Remote Sensing to Species Distribution Modelling: An Integrated Workflow to Monitor Spreading Species in Key Grassland Habitats. Remote Sens. 2021, 13, 1904. [Google Scholar] [CrossRef]
- Lukac, B.; Kramberger, B.; Meclic, V.; Verbic, J. Importance of Non-Leguminous Forbs in Animal Nutrition and Their Ensiling Properties: A Review. Žemdirbyst Agric. 2012, 99, 3–8. [Google Scholar]
- Andueza, D.; Rodrigues, A.M.; Picard, F.; Rossignol, N.; Baumont, R.; Cecato, U.; Farruggia, A. Relationships between Botanical Composition, Yield and Forage Quality of Permanent Grasslands over the First Growth Cycle. Grass Forage Sci. 2016, 71, 366–378. [Google Scholar] [CrossRef]
- Baumont, R.; Aufrère, J.; Niderkorn, V.; Andueza, D.; Surault, F.; Peccatte, J.R.; Delaby, L.; Pelletier, P. La Diversité Spécifique Dans Le Fourrage: Conséquences Sur La Valeur Alimentaire. Fourrages 2008, 194, 189–206. [Google Scholar]
- Loiseau, P. Signification et Limite de l’indice de Valeur Pastorale Pour Le Diagnostic de La Valeur Agricole Des Paturages En Moyenne Montagne Humide. ColI. Phytosoc. 1988, 16, 411–428. [Google Scholar]
- Daget, P.; Godron, M. Pastoralisme: Tropeaux, Espaces et Sociétés; Hatier: Paris, France, 1995. [Google Scholar]
- Pornaro, C.; Basso, E.; Macolino, S. Pasture Botanical Composition and Forage Quality at Farm Scale: A Case Study. Ital. J. Agron. 2019, 14, 214–221. [Google Scholar] [CrossRef]
- Mainetti, A.; Ravetto Enri, S.; Pittarello, M.; Lombardi, G.; Lonati, M. Main Ecological and Environmental Factors Affecting Forage Yield and Quality in Alpine Summer Pastures (NW-Italy, Gran Paradiso National Park). Grass Forage Sci. 2023, 78, 254–267. [Google Scholar] [CrossRef]
- Primi, R.; Filibeck, G.; Amici, A.; Bückle, C.; Cancellieri, L.; Di Filippo, A.; Gentile, C.; Guglielmino, A.; Latini, R.; Mancini, L.D.; et al. From Landsat to Leafhoppers: A Multidisciplinary Approach for Sustainable Stocking Assessment and Ecological Monitoring in Mountain Grasslands. Agric. Ecosyst. Environ. 2016, 234, 118–133. [Google Scholar] [CrossRef]
- Pittarello, M.; Lonati, M.; Ravetto Enri, S.; Lombardi, G. Environmental Factors and Management Intensity Affect in Different Ways Plant Diversity and Pastoral Value of Alpine Pastures. Ecol. Indic. 2020, 115, 106429. [Google Scholar] [CrossRef]
- Napoleone, F.; Probo, M.; Mariotte, P.; Ravetto Enri, S.; Lonati, M.; Argenti, G.; Burrascano, S. Agri-Environmental Payments Drive the Conservation and Forage Value of Semi-Natural Grasslands by Modifying Fine-Scale Grazing Intensity. Biol. Conserv. 2022, 269, 109531. [Google Scholar] [CrossRef]
- Fischer, M.; Stöcklin, J. Local Extinctions of Plants in Remnants of Extensively Used Calcareous Grasslands 1950–1985. Conserv. Biol. 1997, 11, 727–737. [Google Scholar] [CrossRef]
- Luoto, M.; Pykälä, J.; Kuussaari, M. Decline of Landscape-Scale Habitat and Species Diversity after the End of Cattle Grazing. J. Nat. Conserv. 2003, 11, 171–178. [Google Scholar] [CrossRef]
- Kleijn, D.; Rundlöf, M.; Scheper, J.; Smith, H.G.; Tscharntke, T. Does Conservation on Farmland Contribute to Halting the Biodiversity Decline? Trends Ecol. Evol. 2011, 26, 474–481. [Google Scholar] [CrossRef]
- Bricca, A.; Tardella, F.M.; Ferrara, A.; Xinfang, X.; Tolu, F.; Catorci, A. Environmental Heterogeneity Compensates the Potential Homogenising Effect of Abandonment of Grazing in a Sub-Mediterranean Mountain Landscape. Plant Ecol. Divers. 2021, 14, 223–243. [Google Scholar] [CrossRef]
- MacDonald, D.; Crabtree, J.R.; Wiesinger, G.; Dax, T.; Stamou, N.; Fleury, P.; Gutierrez Lazpita, J.; Gibon, A. Agricultural Abandonment in Mountain Areas of Europe: Environmental Consequences and Policy Response. J. Environ. Manag. 2000, 59, 47–69. [Google Scholar] [CrossRef]
- Mottet, A.; Ladet, S.; Coqué, N.; Gibon, A. Agricultural Land-Use Change and Its Drivers in Mountain Landscapes: A Case Study in the Pyrenees. Agric. Ecosyst. Environ. 2006, 114, 296–310. [Google Scholar] [CrossRef]
- Zlinszky, A.; Schroiff, A.; Kania, A.; Deák, B.; Mücke, W.; Vári, Á.; Székely, B.; Pfeifer, N. Categorizing Grassland Vegetation with Full-Waveform Airborne Laser Scanning: A Feasibility Study for Detecting Natura 2000 Habitat Types. Remote Sens. 2014, 6, 8056–8087. [Google Scholar] [CrossRef]
- King, A. Technology: The Future of Agriculture. Nature 2017, 544, S21–S23. [Google Scholar] [CrossRef]
- Tassi, A.; Gigante, D.; Modica, G.; Di Martino, L.; Vizzari, M. Pixel- vs. Object-Based Landsat 8 Data Classification in Google Earth Engine Using Random Forest: The Case Study of Maiella National Park. Remote Sens. 2021, 13, 2299. [Google Scholar] [CrossRef]
- Angelini, F.; Angelini, P.; Angiolini, C.; Bagella, S.; Bonomo, F.; Caccianiga, M.; Santina, C.D.; Gigante, D.; Hutter, M.; Nanayakkara, T.; et al. Robotic Monitoring of Habitats: The Natural Intelligence Approach. IEEE Access 2023, 11, 72575–72591. [Google Scholar] [CrossRef]
- Angelini, F.; Pollayil, M.J.; Bonini, F.; Gigante, D.; Garabini, M. Robotic Monitoring of Grasslands: A Dataset from the EU Natura2000 Habitat 6210* in the Central Apennines (Italy). Sci. Data 2023, 10, 418. [Google Scholar] [CrossRef]
- Kaur, P.; Gigante, D.; Caccianiga, M.; Bagella, S.; Angiolini, C.; Garabini, M.; Angelini, F.; Remagnino, P. Segmentation and Identification of Mediterranean Plant Species. In Advances in Visual Computing; Bebis, G., Ghiasi, G., Fang, Y., Sharf, A., Dong, Y., Weaver, C., Leo, Z., LaViola, J.J., Kohli, L., Eds.; Lecture Notes in Computer Science; Springer Nature: Cham, Switzerland, 2023; Volume 14362, pp. 431–442. [Google Scholar] [CrossRef]
- Pereira, F.C.; Charters, S.; Smith, C.M.S.; Maxwell, T.M.R.; Gregorini, P. A Geospatial Modelling Approach to Assess the Capability of High-Country Stations in Delivering Ecosystem Services. Land 2023, 12, 1243. [Google Scholar] [CrossRef]
- Vizzari, M.; Parracciani, C.; Gigante, D. Enhanced Map Composition and Diachronic Land Cover Classification of Landsat Data in Google Earth Engine. In Computational Science and Its Applications—ICCSA 2023 Workshops; Gervasi, O., Murgante, B., Rocha, A.M.A.C., Garau, C., Scorza, F., Karaca, Y., Torre, C.M., Eds.; Lecture Notes in Computer Science; Springer Nature: Cham, Switzerland, 2023; Volume 14107, pp. 289–299. [Google Scholar] [CrossRef]
- Anderson, N.V.; Petersen, S.L.; Johnson, R.L.; Terry, T.J.; Anderson, V.J. Detecting Floral Resource Availability Using Small Unmanned Aircraft Systems. Land 2024, 13, 99. [Google Scholar] [CrossRef]
- Parracciani, C.; Gigante, D.; Mutanga, O.; Bonafoni, S.; Vizzari, M. Land Cover Changes in Grassland Landscapes: Combining Enhanced Landsat Data Composition, LandTrendr, and Machine Learning Classification in Google Earth Engine with MLP-ANN Scenario Forecasting. GISci. Remote Sens. 2024, 61, 2302221. [Google Scholar] [CrossRef]
- Parracciani, C.; Gigante, D.; Bonini, F.; Grassi, A.; Morbidini, L.; Pauselli, M.; Valenti, B.; Lilli, E.; Antonielli, F.; Vizzari, M. Leveraging Google Earth Engine for a More Effective Grassland Management: A Decision Support Application Perspective. Sensors 2024, 24, 834. [Google Scholar] [CrossRef]
- Michez, A.; Lejeune, P.; Bauwens, S.; Herinaina, A.A.; Blaise, Y.; Castro Muñoz, E.; Lebeau, F.; Bindelle, J. Mapping and Monitoring of Biomass and Grazing in Pasture with an Unmanned Aerial System. Remote Sens. 2019, 11, 473. [Google Scholar] [CrossRef]
- Segarra, J.; Buchaillot, M.L.; Araus, J.L.; Kefauver, S.C. Remote Sensing for Precision Agriculture: Sentinel-2 Improved Features and Applications. Agronomy 2020, 10, 641. [Google Scholar] [CrossRef]
- Santaga, F.S.; Benincasa, P.; Toscano, P.; Antognelli, S.; Ranieri, E.; Vizzari, M. Simplified and Advanced Sentinel-2-Based Precision Nitrogen Management of Wheat. Agronomy 2021, 11, 1156. [Google Scholar] [CrossRef]
- Jiménez-Jiménez, S.I.; Marcial-Pablo, M.D.J.; Ojeda-Bustamante, W.; Sifuentes-Ibarra, E.; Inzunza-Ibarra, M.A.; Sánchez-Cohen, I. VICAL: Global Calculator to Estimate Vegetation Indices for Agricultural Areas with Landsat and Sentinel-2 Data. Agronomy 2022, 12, 1518. [Google Scholar] [CrossRef]
- Amies, A.C.; Dymond, J.R.; Shepherd, J.D.; Pairman, D.; Hoogendoorn, C.; Sabetizade, M.; Belliss, S.E. National Mapping of New Zealand Pasture Productivity Using Temporal Sentinel-2 Data. Remote Sens. 2021, 13, 1481. [Google Scholar] [CrossRef]
- Schino, G.; Borfecchia, F.; De Cecco, L.; Dibari, C.; Iannetta, M.; Martini, S.; Pedrotti, F. Satellite estimate of grass biomass in a mountainous range in central Italy. Agrofor. Syst. 2003, 59, 157–162. [Google Scholar] [CrossRef]
- Edirisinghe, A.; Clark, D.; Waugh, D. Spatio-Temporal Modelling of Biomass of Intensively Grazed Perennial Dairy Pastures Using Multispectral Remote Sensing. Int. J. Appl. Earth Obs. Geoinf. 2012, 16, 5–16. [Google Scholar] [CrossRef]
- Hill, M.J.; Donald, G.E.; Hyder, M.W.; Smith, R.C.G. Estimation of Pasture Growth Rate in the South West of Western Australia from AVHRR NDVI and Climate Data. Remote Sens. Environ. 2004, 93, 528–545. [Google Scholar] [CrossRef]
- Porter, T.F.; Chen, C.; Long, J.A.; Lawrence, R.L.; Sowell, B.F. Estimating Biomass on CRP Pastureland: A Comparison of Remote Sensing Techniques. Biomass Bioenergy 2014, 66, 268–274. [Google Scholar] [CrossRef]
- Barrachina, M.; Cristóbal, J.; Tulla, A.F. Estimating Above-Ground Biomass on Mountain Meadows and Pastures through Remote Sensing. Int. J. Appl. Earth Obs. Geoinf. 2015, 38, 184–192. [Google Scholar] [CrossRef]
- Filippa, G.; Cremonese, E.; Galvagno, M.; Bayle, A.; Choler, P.; Bassignana, M.; Piccot, A.; Poggio, L.; Oddi, L.; Gascoin, S.; et al. On the Distribution and Productivity of Mountain Grasslands in the Gran Paradiso National Park, NW Italy: A Remote Sensing Approach. Int. J. Appl. Earth Obs. Geoinf. 2022, 108, 102718. [Google Scholar] [CrossRef]
- Cabrera-Bosquet, L.; Molero, G.; Stellacci, A.; Bort, J.; Nogués, S.; Araus, J. NDVI as a Potential Tool for Predicting Biomass, Plant Nitrogen Content and Growth in Wheat Genotypes Subjected to Different Water and Nitrogen Conditions. Cereal Res. Commun. 2011, 39, 147–159. [Google Scholar] [CrossRef]
- Li, Z.; Huffman, T.; McConkey, B.; Townley-Smith, L. Monitoring and Modeling Spatial and Temporal Patterns of Grassland Dynamics Using Time-Series MODIS NDVI with Climate and Stocking Data. Remote Sens. Environ. 2013, 138, 232–244. [Google Scholar] [CrossRef]
- Shelestov, A.; Lavreniuk, M.; Kussul, N.; Novikov, A.; Skakun, S. Exploring Google Earth Engine Platform for Big Data Processing: Classification of Multi-Temporal Satellite Imagery for Crop Mapping. Front. Earth Sci. 2017, 5, 232994. [Google Scholar] [CrossRef]
- Tamiminia, H.; Salehi, B.; Mahdianpari, M.; Quackenbush, L.; Adeli, S.; Brisco, B. Google Earth Engine for Geo-Big Data Applications: A Meta-Analysis and Systematic Review. ISPRS J. Photogramm. Remote Sens. 2020, 164, 152–170. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-Scale Geospatial Analysis for Everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Kumar, L.; Mutanga, O. Google Earth Engine Applications Since Inception: Usage, Trends, and Potential. Remote Sens. 2018, 10, 1509. [Google Scholar] [CrossRef]
- Rowland, A.; Folmer, E.; Beek, W. Towards Self-Service GIS—Combining the Best of the Semantic Web and Web GIS. IJGI 2020, 9, 753. [Google Scholar] [CrossRef]
- Pittarello, M.; Probo, M.; Perotti, E.; Lonati, M.; Lombardi, G.; Ravetto Enri, S. Grazing Management Plans Improve Pasture Selection by Cattle and Forage Quality in Sub-Alpine and Alpine Grasslands. J. Mt. Sci. 2019, 16, 2126–2135. [Google Scholar] [CrossRef]
- Catorci, A.; Lulli, R.; Malatesta, L.; Tavoloni, M.; Tardella, F.M. How the Interplay between Management and Interannual Climatic Variability Influences the NDVI Variation in a Sub-Mediterranean Pastoral System: Insight into Sustainable Grassland Use under Climate Change. Agric. Ecosyst. Environ. 2021, 314, 107372. [Google Scholar] [CrossRef]
- Scocco, P.; Piermarteri, K.; Malfatti, A.; Tardella, F.M.; Catorci, A. Increase of Drought Stress Negatively Affects the Sustainability of Extensive Sheep Farming in Sub-Mediterranean Climate. J. Arid Environ. 2016, 128, 50–58. [Google Scholar] [CrossRef]
- Boschetti, M.; Bocchi, S.; Brivio, P.A. Assessment of Pasture Production in the Italian Alps Using Spectrometric and Remote Sensing Information. Agric. Ecosyst. Environ. 2007, 118, 267–272. [Google Scholar] [CrossRef]
- Argenti, G.; Bottai, L.; Chiesi, M.; Maselli, F.; Staglianò, N.; Targetti, S. Analisi e Valutazione Di Pascoli Montani Attraverso l’integrazionedi Dati Multispettrali e Ausiliari. Ital. J. Remote Sens./Riv. Ital.Telerilevam. 2011, 43, 45–57. [Google Scholar] [CrossRef]
- Valkó, O.; Zmihorski, M.; Biurrun, I.; Loos, J.; Labadessa, R.; Venn, S. Ecology and Conservation of Steppes and Semi-Natural Grasslands. Hacquetia 2016, 15, 5–14. [Google Scholar] [CrossRef]
- Karlík, P.; Poschlod, P. Identifying Plant and Environmental Indicators of Ancient and Recent Calcareous Grasslands. Ecol. Indic. 2019, 104, 405–421. [Google Scholar] [CrossRef]
- Ribeiro, P.F.; Santos, J.L. Exploring the Effects of Climate Change on Farming System Choice: A Farm-Level Space-for-Time Approach. Land 2023, 12, 2113. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
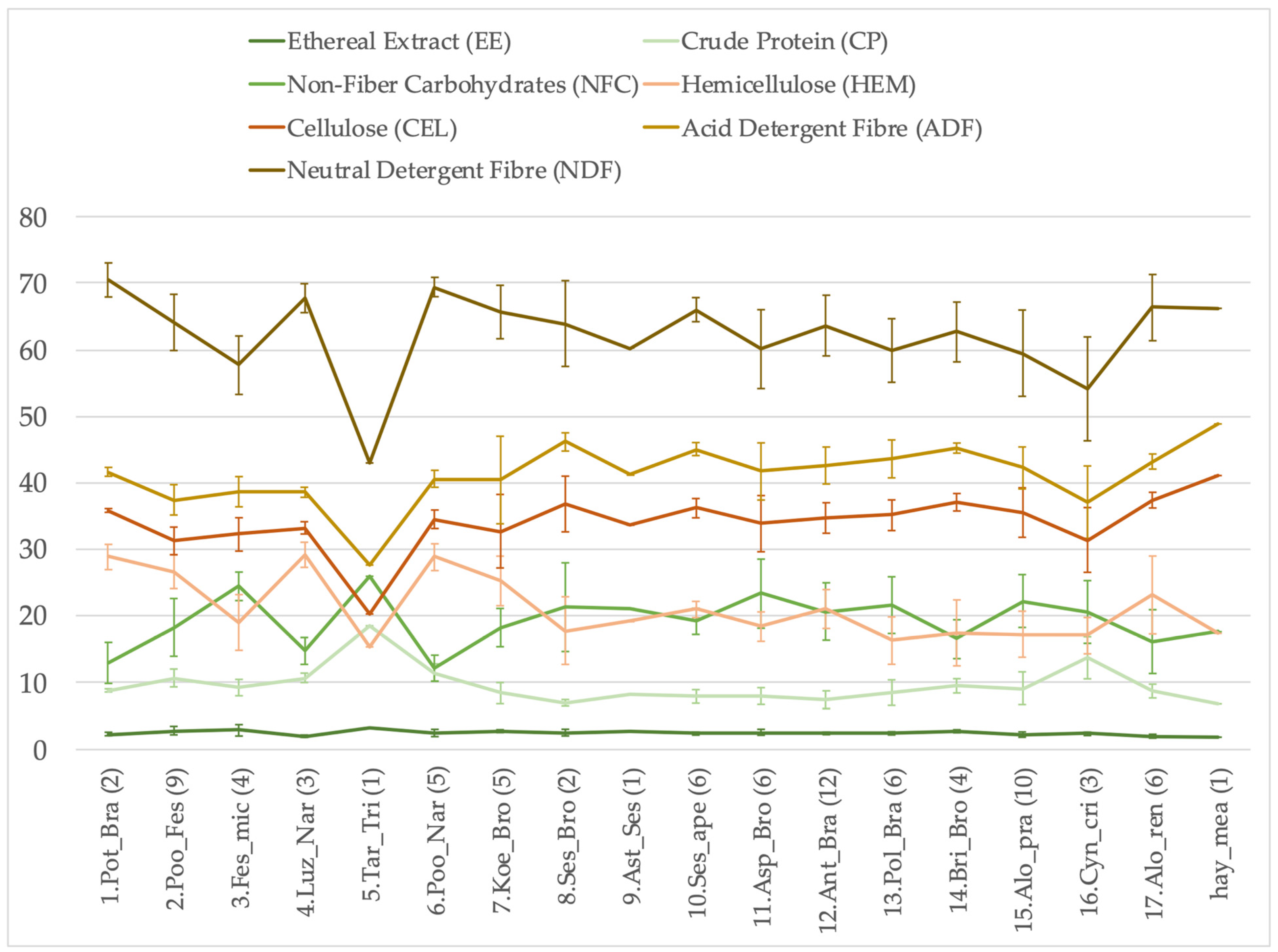
| DM | CEL | HEM | EE | ADF | NDF | ADL | CP | NFC | |
|---|---|---|---|---|---|---|---|---|---|
| H χ2 = 59.20 p < 0.001 | H χ2 = 44.45 p < 0.001 | H χ2 = 44.45 p < 0.001 | H χ2 = 54.21 p < 0.001 | H χ2 = 60.18 p < 0.001 | H χ2 = 56.14 p < 0.001 | H χ2 = 62.68 p < 0.001 | H χ2 = 49.96 p < 0.001 | H χ2 = 62.75 p < 0.001 | |
| 1.Pot_Bra (2) | 98.5 ± 42.4 | 35.2 ± 14.9 | 28.0 ± 10.4 | 2.2 ± 0.7 | 40.9 ± 17.0 | 68.8 ± 27.3 | 5.6 ± 2.1 | 8.6 ± 3.5 | 13.4 ± 8.5 |
| 2.Poo_Fes (9) | 80.2 ± 39.7 | 25.4 ± 12.8 | 21.6 ± 10.7 | 2.3 ± 1.3 | 30.2 ± 14.9 | 51.8 ± 25.6 | 4.8 ± 2.2 | 8.5 ± 4.3 | 14.2 ± 8.0 |
| 3.Fes_mic (4) | 172.4 ± 95.3 | 54.6 ± 27.3 | 31.0 ± 12.6 | 5.2 ± 3.5 | 66.1 ± 34.8 | 97.0 ± 46.7 | 11.5 ± 7.6 | 16.5 ± 10.7 | 43.6 ± 28.6 |
| 4.Luz_Nar (3) | 100.8 ± 23.2 | 33.5 ± 7.5 | 29.2 ± 5.0 | 2.1 ± 0.7 | 38.8 ± 8.4 | 68.0 ± 13.3 | 5.3 ± 0.9 | 10.8 ± 2.3 | 15.1 ± 5.6 |
| 5.Tar_Tri (1) | 26.4 | 5.4 | 4.1 | 0.9 | 7.3 | 11.3 | 1.9 | 4.9 | 6.9 |
| 6.Poo_Nar (5) | 66.3 ± 17.5 | 22.9 ± 6.4 | 19.3 ± 6.0 | 1.6 ± 0.5 | 26.9 ± 7.3 | 46.2 ± 13.1 | 4.0 ± 0.9 | 7.7 ± 2.4 | 7.8 ± 1.4 |
| 7.Koe_Bro (5) | 110.4 ± 32.8 | 35.0 ± 5.6 | 28.8 ± 13.2 | 3.0 ± 0.8 | 43.2 ± 6.9 | 72.0 ± 19.1 | 8.3 ± 1.4 | 9.6 ± 4.4 | 20.4 ± 7.7 |
| 8.Ses_Bro (2) | 209.4 ± 84.6 | 78.8 ± 39.9 | 39.4 ± 25.7 | 5.5 ± 3.2 | 97.2 ± 41.9 | 136.6 ± 67.6 | 18.3 ± 2.0 | 14.4 ± 4.9 | 41.8 ± 4.1 |
| 9.Ast_Ses (1) | 463.8 | 155.9 | 88.7 | 12.5 | 190.8 | 279.5 | 34.9 | 37.9 | 97.9 |
| 10.Ses_ape (6) | 140.0 ± 19.2 | 50.7 ± 7.8 | 29.3 ± 4.1 | 3.3 ± 0.5 | 63.1 ± 9.0 | 92.4 ± 12.9 | 12.4 ± 1.7 | 11.1 ± 1.9 | 26.9 ± 5.0 |
| 11.Asp_Bro (6) | 125.8 ± 76.7 | 44.6 ± 29.7 | 23.6 ± 14.8 | 3.5 ± 2.3 | 54.7 ± 36.2 | 78.2 ± 50.9 | 10.0 ± 6.6 | 9.8 ± 6.1 | 26.7 ± 14.6 |
| 12.Ant_Bra (12) | 183.5 ± 55.1 | 63.6 ± 18.9 | 38.1 ± 10.6 | 4.4 ± 1.3 | 78.5 ± 25.2 | 116.6 ± 34.9 | 14.9 ± 6.7 | 13.8 ± 5.4 | 38.1 ± 15.1 |
| 13.Pol_Bra (6) | 126.9 ± 62.7 | 45.5 ± 25.6 | 22.4 ± 17.6 | 3.1 ± 1.7 | 55.9 ± 29.5 | 78.3 ± 47.0 | 10.5 ± 4.2 | 10.4 ± 4.1 | 25.8 ± 7.5 |
| 14.Bri_Bro (4) | 134.3 ± 34.5 | 49.8 ± 13.2 | 23.3 ± 9.5 | 3.7 ± 0.9 | 60.8 ± 15.9 | 84.0 ± 22.9 | 11.0 ± 3.3 | 12.8 ± 3.7 | 22.2 ± 6.9 |
| 15.Alo_pra (10) | 558.2 ± 301.4 | 207.1 ± 126.2 | 104.4 ± 70.9 | 11.6 ± 4.7 | 242.9 ± 143.1 | 347.3 ± 213.7 | 35.8 ± 17.4 | 45.9 ± 19.0 | 115.4 ± 48.0 |
| 16.Cyn_cri (3) | 162.9 ± 50.6 | 51.3 ± 19.9 | 28.2 ± 12.1 | 3.8 ± 1.0 | 60.3 ± 21.9 | 88.6 ± 34.0 | 9.0 ± 2.1 | 22.0 ± 7.3 | 33.4 ± 12.4 |
| 17.Alo_ren (6) | 528.0 ± 53.9 | 197.0 ± 15.9 | 122.7 ± 35.9 | 10.5 ± 1.3 | 227.7 ± 20.5 | 350.5 ± 47.3 | 30.7 ± 7.9 | 45.7 ± 3.7 | 85.6 ± 26.5 |
| hay_mea (1) | 689.1 | 283.1 | 119.7 | 12.8 | 336.6 | 456.3 | 53.5 | 47 | 121.4 |
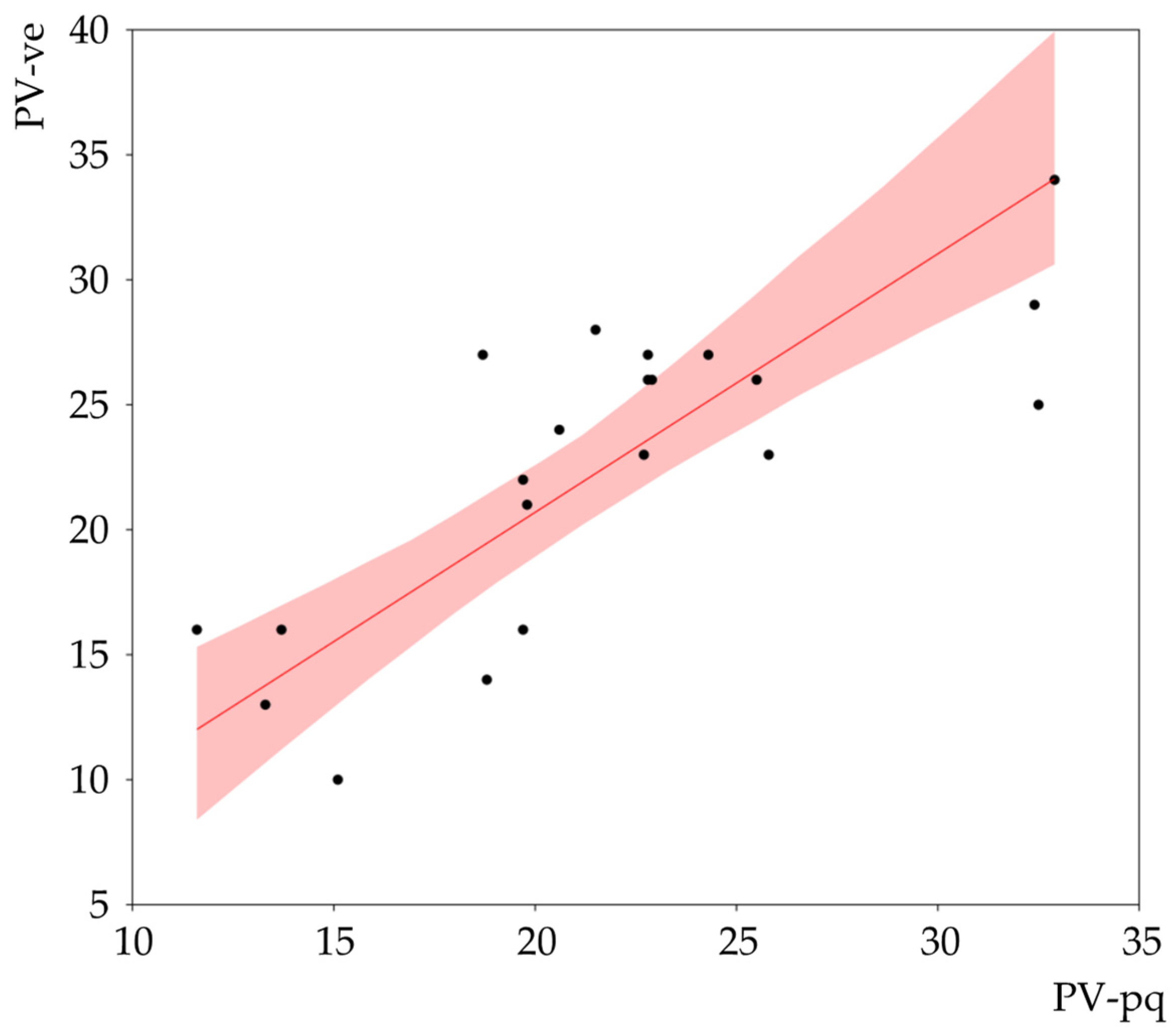
| Parameters | Slope | r | r2 | p |
|---|---|---|---|---|
| DM (%) | 0.06 | 0.15 | 0.02 | n.s. |
| EE (%) | 0.32 | 0.16 | 0.03 | n.s. |
| NDF (%) | −1.84 | −0.52 | 0.27 | *** |
| ADF (%) | −0.78 | −0.22 | 0.05 | * |
| ADL (%) | −0.09 | −0.05 | 0.00 | n.s. |
| HEM (%) | −0.61 | −0.43 | 0.02 | *** |
| CEL (%) | −0.68 | −0.21 | 0.05 | * |
| CP (%) | 0.15 | 0.10 | 0.01 | n.s. |
| NFC (%) | 0.62 | 0.46 | 0.21 | *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gigante, D.; Angelucci, S.; Bonini, F.; Caruso, F.; Di Cecco, V.; Donnini, D.; Morbidini, L.; Pauselli, M.; Valenti, B.; Tassi, A.; et al. Seminatural Grasslands: An Emblematic Challenge for Nature Conservation in Protected Areas. Land 2024, 13, 386. https://doi.org/10.3390/land13030386
Gigante D, Angelucci S, Bonini F, Caruso F, Di Cecco V, Donnini D, Morbidini L, Pauselli M, Valenti B, Tassi A, et al. Seminatural Grasslands: An Emblematic Challenge for Nature Conservation in Protected Areas. Land. 2024; 13(3):386. https://doi.org/10.3390/land13030386
Chicago/Turabian StyleGigante, Daniela, Simone Angelucci, Federica Bonini, Federico Caruso, Valter Di Cecco, Domizia Donnini, Luciano Morbidini, Mariano Pauselli, Bernardo Valenti, Andrea Tassi, and et al. 2024. "Seminatural Grasslands: An Emblematic Challenge for Nature Conservation in Protected Areas" Land 13, no. 3: 386. https://doi.org/10.3390/land13030386