
Etiology of Ulcerative Dermal Necrosis (UDN) in Brown Trout (Salmo trutta Morpha trutta)—Preliminary Results
 , , , , ,
, , , , , {kind=link}
{kind=link}
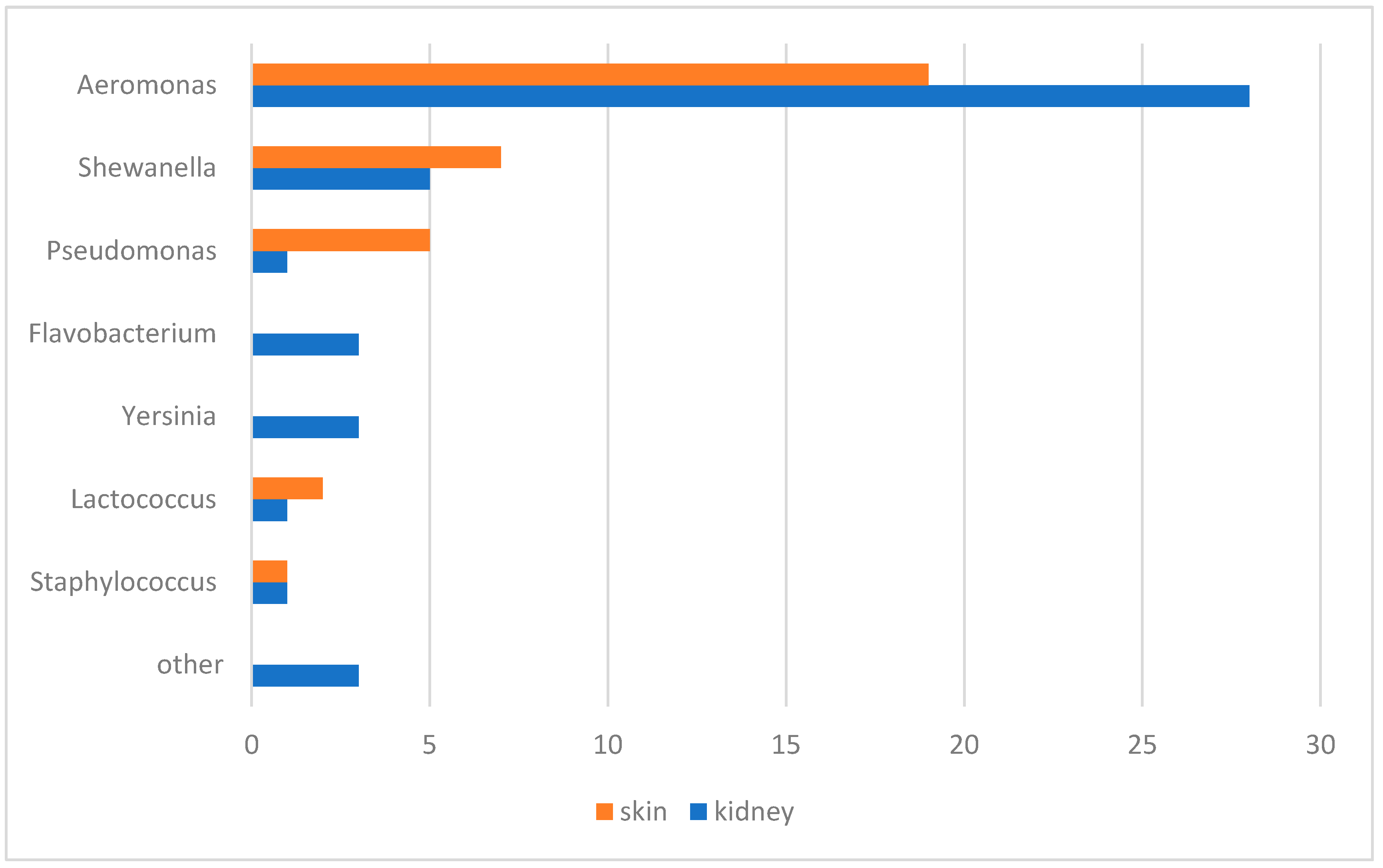{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Delivery and Maintenance
2.2. In Vivo Experiments
2.2.1. First Experiment
2.2.2. Second Experiment
2.2.3. Third Experiment
2.2.4. Fourth Experiment: Cohabitation
2.3. Clinical Examination
2.4. Sample Collection
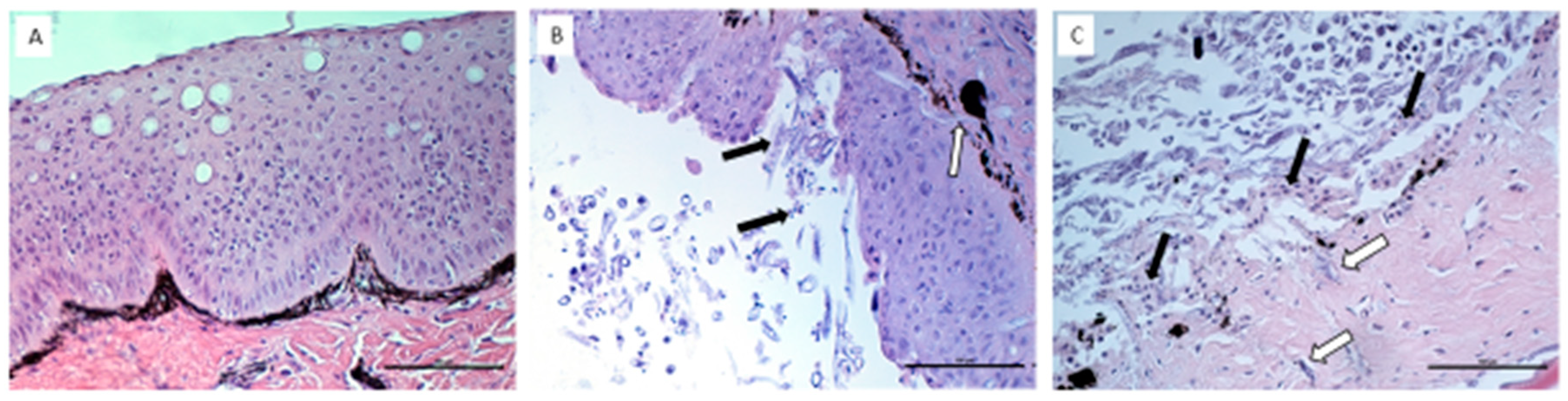
2.5. Histopathological Assessment of UDN Lesions
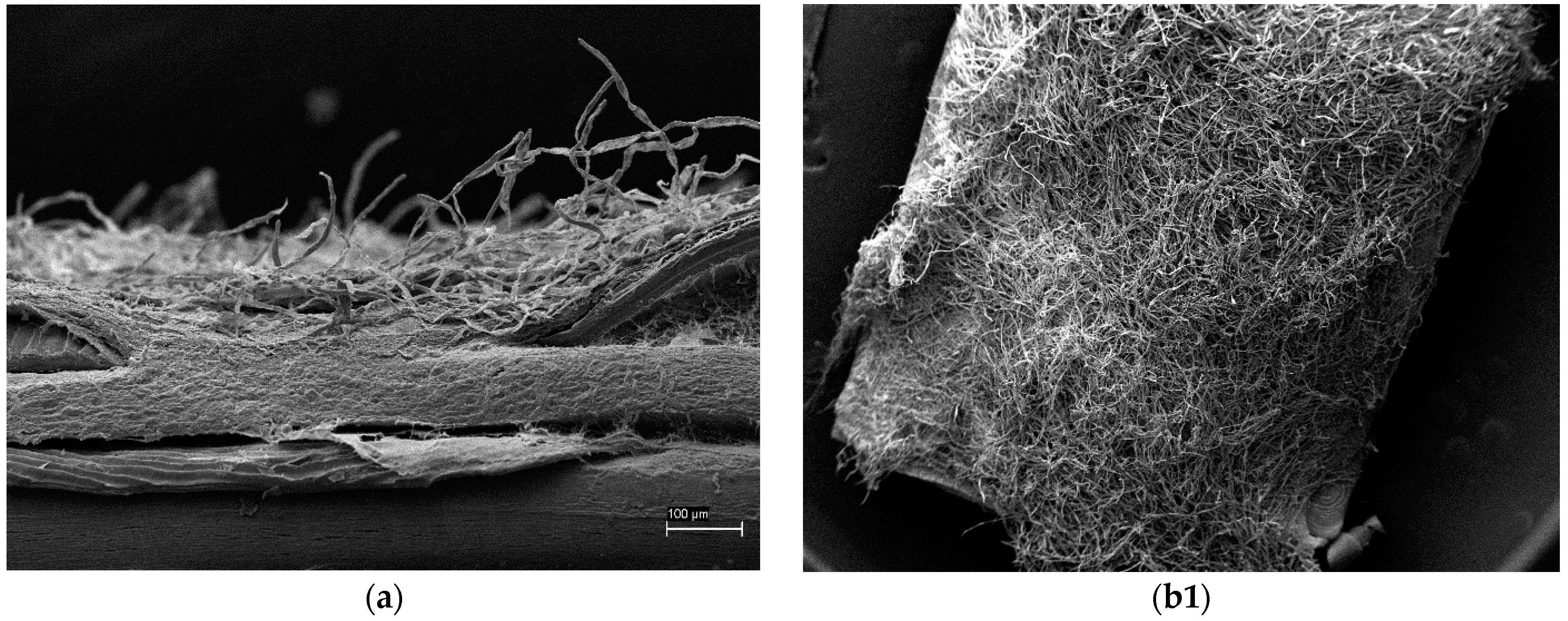
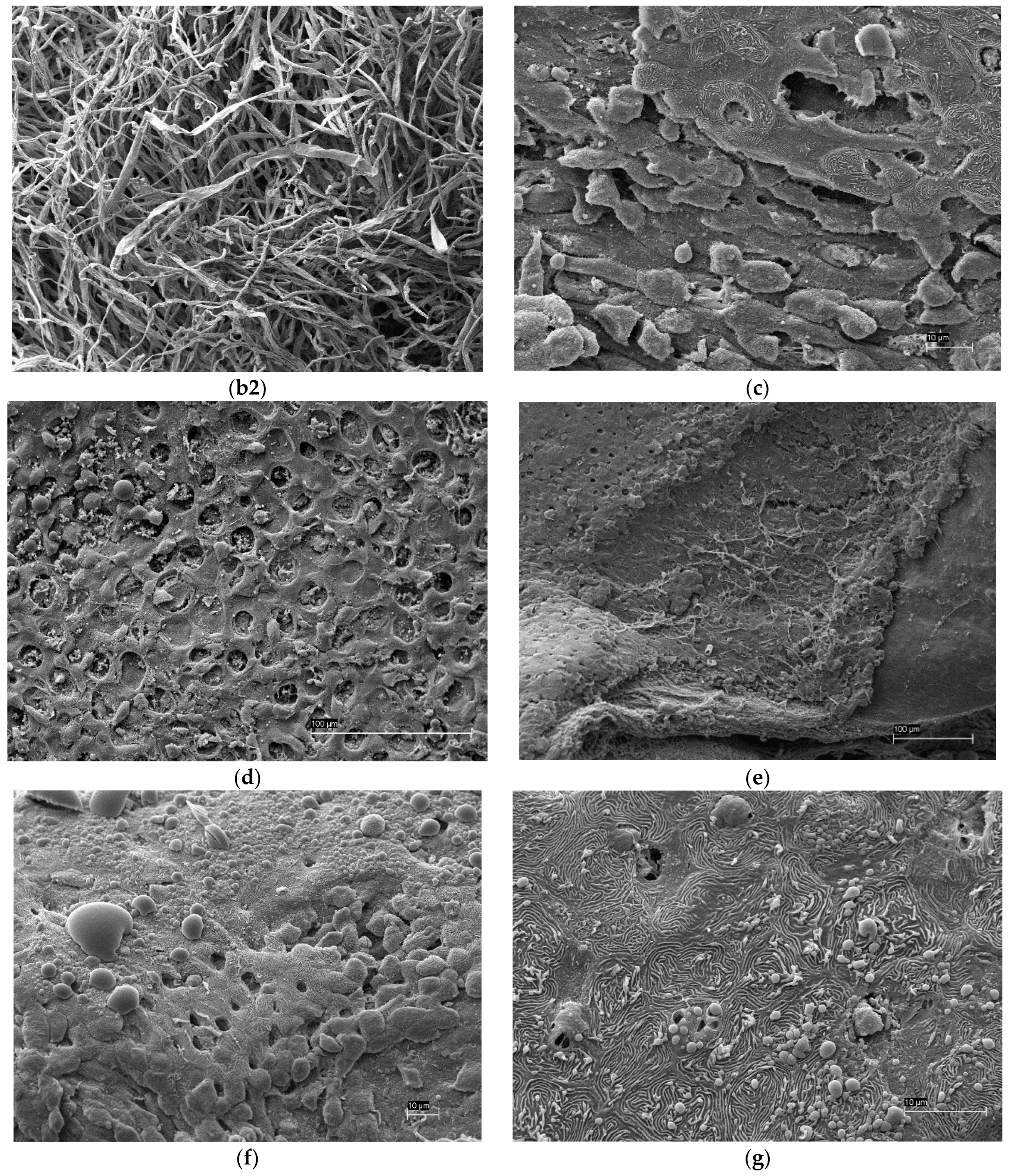
2.6. Scanning Electron Microscopy (SEM)
2.7. Virological Examination
2.7.1. Virus Isolation in Cell Cultures
2.7.2. RNA Extraction
2.7.3. RT-PCR
2.8. Bacteriological and Mycological Examination
3. Results
3.1. In Vivo Experiments
3.2. Histopathological Assessment of UDN Lesions
3.3. Bacteriological and Mycological Examination
3.4. Scanning Electron Microscopy (SEM)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roberts, R.J. Ulcerative dermal necrosis (UDN) in wild salmonids. Fish. Res. 1993, 17, 3–14. [Google Scholar] [CrossRef]
- Mannerla, M.; Andersson, M.; Birzaks, J.; Debowski, P.; Degerman, E.; Huhmarniemi, A.; Häggström, H.; Ikonen, E.; Jokikokko, E.; Jutila, E.; et al. Helcom: Salmon and Sea Trout Populations and Rivers in the Baltic Sea—HELCOM assessment of salmon (Salmo salar) and sea trout (Salmo trutta) populations and habitats in rivers flowing to the Baltic Sea. Balt. Sea Environ. Proc. 2011, 19, 126A. [Google Scholar]
- ICES: Report of the Baltic Salmon and Trout Assessment Working Group (WGBAST), 22–30 March 2011, Riga, Latvia; 2011/ACOM:08; International Council for the Exploration of the Sea location (ICES): Copenhagen, Denmark, 2011; 297p, Available online: https://jukuri.luke.fi/handle/10024/538830 (accessed on 4 January 2024).
- Bartel, R.; Bernaś, R.; Grudniewska, J.; Jesiołowski, M.; Kacperska, B.; Marczyński, A.; Pazda, R.; Pender, R.; Połomski, S.; Skóra, M.; et al. Furunculosis in salmon (Salmo salar) and Sea Trout (Salmo trutta trutta) in Poland in 2007 and 2008. Komun. Rybackie 2009, 3, 7–13. [Google Scholar]
- Carbery, J.T. Ulcerative dermal necrosis (UDN) of salmon: Description, aetiology and differential diagnosis. Bull. Off. Int. Epizoot. 1968, 69, 1401–1410. [Google Scholar] [PubMed]
- Carbery, J.T.; Strickland, K.L. Resistance of Rainbow Trout to Ulcerative Dermal Necrosis. Nature 1968, 217, 1158. [Google Scholar] [CrossRef]
- Bernaś, R.; Wąs-Barcz, A. Genetic structure of important resident brown trout breeding lines in Poland. J. Appl. Genet 2020, 61, 239–247. [Google Scholar] [CrossRef]
- Topić Popović, N.; Strunjak-Perović, I.; Čož-Rakovac, R.; Barišić, J.; Jadan, M.; Beraković, A.P.; Klobučar, R.S. Tricaine methane-sulfonate (MS-222) application in fish anaesthesia. J. Appl. Ichthyol. 2012, 28, 553–564. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Triinu Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic. Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Office International des Épizooties. Manual of Diagnostic Tests for Aquatic Animals, Tenth Edition; Infection with Viral Haemorrhagic Septicaemia Virus (VHSV); WOAH: Paris, France, 2023; Chapter 2.3.10.; pp. 377–401. [Google Scholar]
- Office International des Épizooties. Manual of Diagnostic Tests for Aquatic Animals, Tenth Edition; Infection with Infectious Haematopoietic Necrosis Virus (IHN); WOAH: Paris, France, 2023; Chapter 2.3.5.; pp. 297–313. [Google Scholar]
- Maj-Paluch, J.; Matras, M.; Borzym, E.; Stachnik, M.; Reichert, M. Phylogenetic characterization of Polish isolates of infectious pancreatic necrosis virus in salmonid fish. J. Fish Dis. 2020, 43, 1443–1451. [Google Scholar] [CrossRef] [PubMed]
- Office International des Épizooties. Manual of Diagnostic Tests for Aquatic Animals, Tenth Edition, Infection with HRP-Deleted or HPR0 Infectious Salmon Anaemia Virus; WOAH: Paris, France, 2023; Chapter 2.3.4.; pp. 1–18. [Google Scholar]
- Paździor, E.; Pękala-Safińska, A.; Wasyl, D. Genotypic diversity among Shewanella spp. collected from freshwater fish. J. Fish Dis. 2019, 42, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, C.; Colom, F.S.; Mulet, E.; Abad, J.L.; Alio, A.L. Detection and identification of fungal pathogens by PCR and by ITS2 and 5.8S ribosomal DNA typing in ocular infections. J. Clin. Microbiol. 2001, 39, 2873–2879. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Roberts, S.D.; Powell, M.D. Comparative ionic flux and gill mucous cell histochemistry: Effects of salinity and disease status in Atlantic salmon (Salmo salar L.). Comp. Biochem. Physiol. A Mol. Int. Physiol. 2003, 134, 525–537. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Pickering, A.D.; Pottinger, T.G. Lymphocytopenia and interrenal activity during sexual maturation in the brown trout Salmo trutta L. J. Fish Biol. 1987, 30, 41–50. [Google Scholar] [CrossRef]
- Maule, A.G.; Schrock, R.; Slater, C.; Fitzpatrick, M.S.; Schreck, C.B. Immune and endocrine responses of adult chinook salmon during freshwater immigration and sexual maturation. Fish Shellfish Immunol. 1996, 6, 221–233. [Google Scholar] [CrossRef]
- Hou, Y.; Suzuki, Y.; Aida, K. Changes in immunoglobulin producing cells in response to gonadal maturation in rainbow trout. Fish Sci. 1999, 65, 844–849. [Google Scholar] [CrossRef]
- Suzuki, Y.; Otaka, T.; Sato, S.; Hou, Y.Y.; Aida, K. Reproduction related immunoglobulin changes in rainbow trout. Fish Physiol. Biochem. 1997, 17, 415–421. [Google Scholar] [CrossRef]
- Campbell, J.H.; Dixon, B.; Whitehouse, L.M. The intersection of stress, sex and immunity in fishes. Immunogenetics 2021, 73, 111–129. [Google Scholar] [CrossRef]
- Dolan, B.P.; Fisher, K.M.; Colvin, M.E.; Benda, S.E.; Peterson, J.T.; Kent, M.L.; Schreck, C.B. Innate and adaptive immune responses in migrating spring-run adult chinook salmon Oncorhynchus tshawytscha. Fish Shellfish Immunol. 2016, 48, 136–144. [Google Scholar] [CrossRef]
- Tripp, R.A.; Maule, A.G.; Schreck, C.B.; Kaattari, S.L. Cortisol mediated suppression of salmonid lymphocyte responses in vitro. Dev. Comp. Immunol. 1987, 11, 565–576. [Google Scholar] [CrossRef]
- Dash, S.; Das, S.K.; Samal, J.; Thatoi, H.N. Epidermal mucus, a major determinant in fish health: A review. Iran. J. Vet. Res. 2018, 19, 72–81. [Google Scholar]
- Cordero, H.; Morcillo, P.; Cuesta, A.; Brinchmann, M.F.; Esteban, M.A.J. Differential proteome profile of skin mucus of gilthead seabream (Sparus aurata) after probiotic intake and/or overcrowding stress. Proteomics 2016, 132, 41–50. [Google Scholar] [CrossRef]
- Benhamed, S.; Guardiola, F.A.; Mars, M.; Esteban, M.Á. Pathogen bacteria adhesion to skin mucus of fishes. Vet. Microbiol. 2014, 171, 1–12. [Google Scholar] [CrossRef]
- Guardiola, F.A.; Cuesta, A.; Arizcun, M.; Meseguer, J.; Esteban, M.A. Comparative skin mucus and serum humoral defence mechanisms in the teleost gilthead seabream (Sparus aurata). Fish Shellfish Immunol. 2014, 36, 545–551. [Google Scholar] [CrossRef]
- Scrimshaw, N.S.; SanGiovanni, J.P. Synergism of nutrition, infection, and immunity: An overview. Am. J. Clin. Nutr. 1997, 66, 464S–477S. [Google Scholar] [CrossRef] [PubMed]
- McMurray, D.N. Cell-mediated immunity in nutritional deficiency. Prog. Food Nutr. Sci. 1984, 8, 193–228. [Google Scholar]
- Chandra, R.K.; Kumari, S. Nutrition and immunity: An overview. J. Nutr. 1994, 124 (Suppl. S8), 1433S–1435S. [Google Scholar] [CrossRef] [PubMed]
- Ritz, B.W.; Aktan, I.; Shoko, N.; Gardner, E.M. Energy restriction impairs natural killer cell function and increases the severity of influenza infection in young adult male C57BL/6 mice. J. Nutr. 2008, 138, 2269–2275. [Google Scholar] [CrossRef]
- Echeverría-Bugueño, M.; Irgang, R.; Mancilla-Schulz, J.; Avendaño-Herrera, R. Healthy and infected Atlantic salmon (Salmo salar) skin-mucus response to Tenacibaculum dicentrarchi under in vitro conditions. Fish Shellfish Immunol. 2023, 136, 108747. [Google Scholar] [CrossRef] [PubMed]
- Deng, F.; Wang, D.; Chen, F.; Lu, T.; Li, S. Molecular characterization and expression analysis of claudin-4-like in rainbow trout involved in Flavobacterium psychrophilum infection. Fish Shellfish Immunol. 2022, 130, 244–251. [Google Scholar] [CrossRef]
- Ferguson, H.W.; Morrison, D.; Ostland, V.E.; Lumsden, J.; Byrne, P. Responses of mucus-producing cells in gill disease of rainbow trout (Oncorhynchus mykiss). J. Comp. Pathol. 1992, 106, 255–265. [Google Scholar] [CrossRef]
- Lease, H.M.; Hansen, J.A.; Bergman, H.L.; Meyer, J.S. Structural changes in gills of Lost River suckers exposed to elevated pH and ammonia concentrations. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2003, 134, 491–500. [Google Scholar] [CrossRef]
- Zehra, A.Z.; Oğuz, A. Investigation of gill mucus cells of Lake Van fish (Alburnus tarichi) during reproductive migration. Fish Physiol. Biochem. 2021, 47, 409–419. [Google Scholar]
- Li, Y.; Huang, D.; Sun, W.; Sun, X.; Yan, G.; Gao, W.; Lin, H. Characterizing sediment bacterial community and identifying the biological indicators in a seawater-freshwater transition zone during the wet and dry seasons. Environ. Sci. Pollut. Res. Int. 2022, 29, 41219–41230. [Google Scholar] [CrossRef]
- Kondo, M.; Kawai, K.; Kurohara, K.; Oshima, S. Adherence of Flavobacterium psychrophilum on the body surface of the ayu Plecoglossus altivelis. Microbes Infect. 2002, 4, 279–283. [Google Scholar] [CrossRef]
- Srivastava, A.; Mistri, A.; Mittal, S.; Mittal, A.J. Alterations in the epidermis of the carp, Labeo rohita (Cyprinidae: Cypriniformes), infected by the bacteria, Aeromonas hydrophila: A scanning electron microscopic, histopathological and immunohistochemical investigation. J. Fish Dis. 2020, 43, 941–953. [Google Scholar] [CrossRef] [PubMed]
- Ferri, S. Modification of microridge pattern in teleost (Pimelodus maculatus) epidermal cells induced by NaCl. Gegenbaurs Morphol. Jahrb. 1983, 129, 325–329. [Google Scholar] [PubMed]
- Yamada, J. A study of the structure of surface cell layers in the epidermis of some teleosts. Annot. Zool. Jpn. 1968, 41, 1–8. [Google Scholar]
- Hussain, A.; Shashwati, G.S. Fish Epidermal Mucus as a Source of Diverse Therapeutical Compounds. Int. J. Pept. Res. Ther. 2023, 29, 36. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matras, M.; Stachnik, M.; Kycko, A.; Wasiak, M.; Paździor, E.; Maj-Paluch, J.; Borzym, E.; Reichert, M. Etiology of Ulcerative Dermal Necrosis (UDN) in Brown Trout (Salmo trutta Morpha trutta)—Preliminary Results. Pathogens 2024, 13, 251. https://doi.org/10.3390/pathogens13030251
Matras M, Stachnik M, Kycko A, Wasiak M, Paździor E, Maj-Paluch J, Borzym E, Reichert M. Etiology of Ulcerative Dermal Necrosis (UDN) in Brown Trout (Salmo trutta Morpha trutta)—Preliminary Results. Pathogens. 2024; 13(3):251. https://doi.org/10.3390/pathogens13030251
Chicago/Turabian StyleMatras, Marek, Magdalena Stachnik, Anna Kycko, Magdalena Wasiak, Ewa Paździor, Joanna Maj-Paluch, Ewa Borzym, and Michał Reichert. 2024. "Etiology of Ulcerative Dermal Necrosis (UDN) in Brown Trout (Salmo trutta Morpha trutta)—Preliminary Results" Pathogens 13, no. 3: 251. https://doi.org/10.3390/pathogens13030251