

Extensive Countrywide Molecular Identification and High Genetic Diversity of Haemonchus spp. in Domestic Ruminants in Greece
 , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Parasite Material
2.2. Morphological Identification
2.3. Isolation of Genomic DNA
2.4. PCR Amplification and Sequencing
2.5. Data Analysis
2.6. Statistical Analysis
3. Results
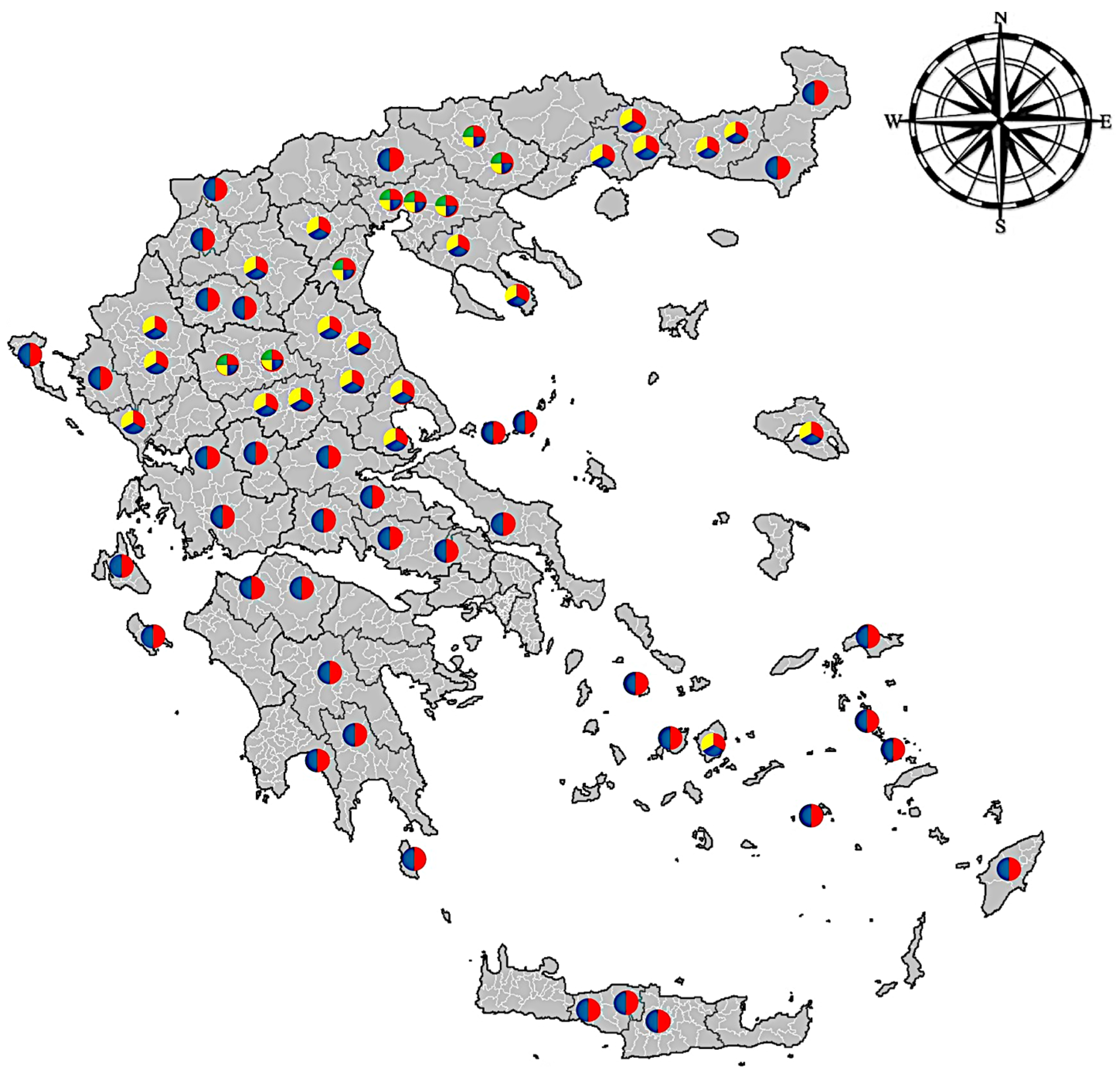
3.1. Morphological Identification
3.2. Molecular Identification of Haemonchus Population, at the Species Level, Based on ITS2 Sequence
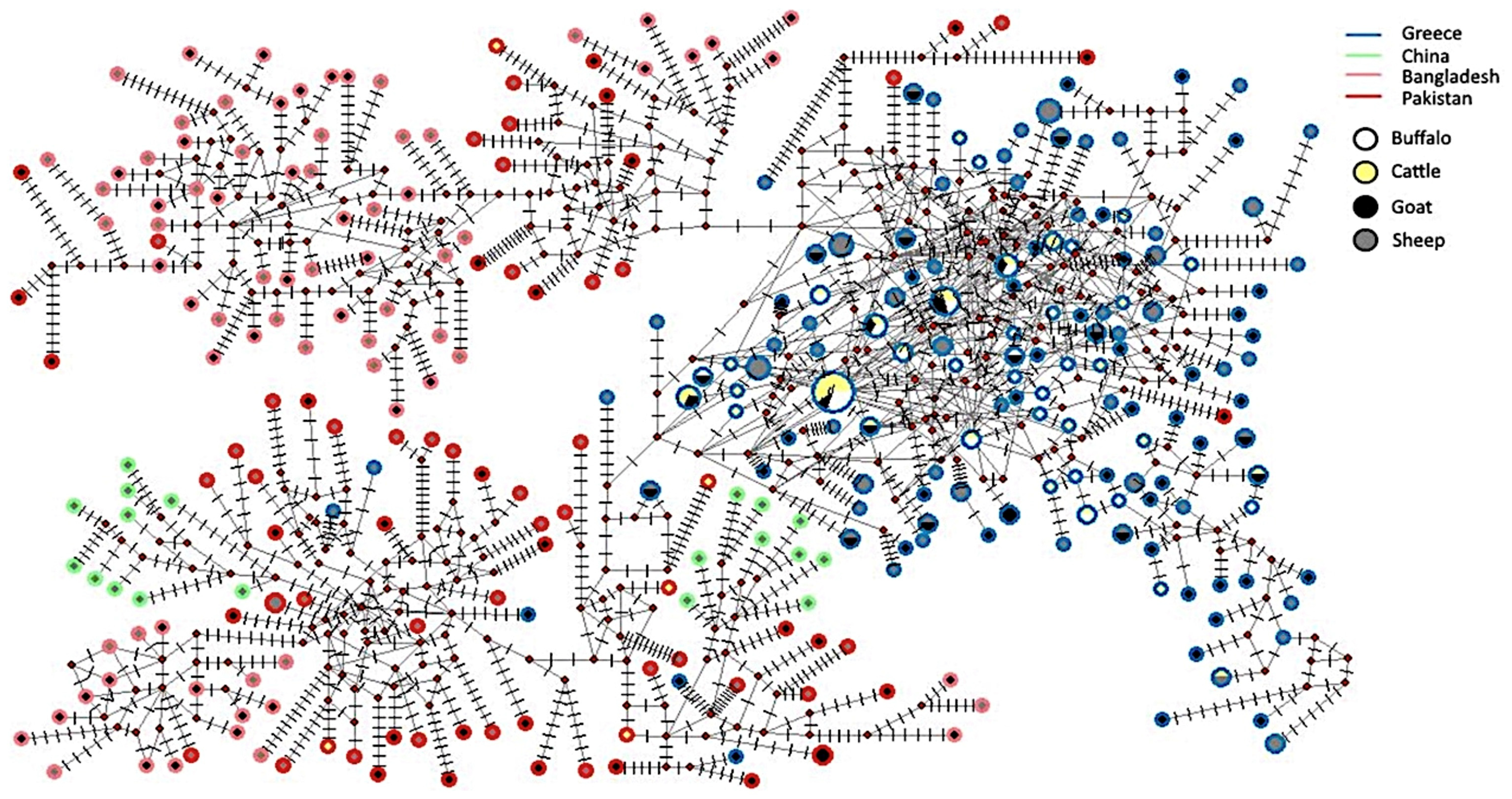
3.3. Estimation of Genetic Diversity of H. contortus Population Based on ND4 Gene
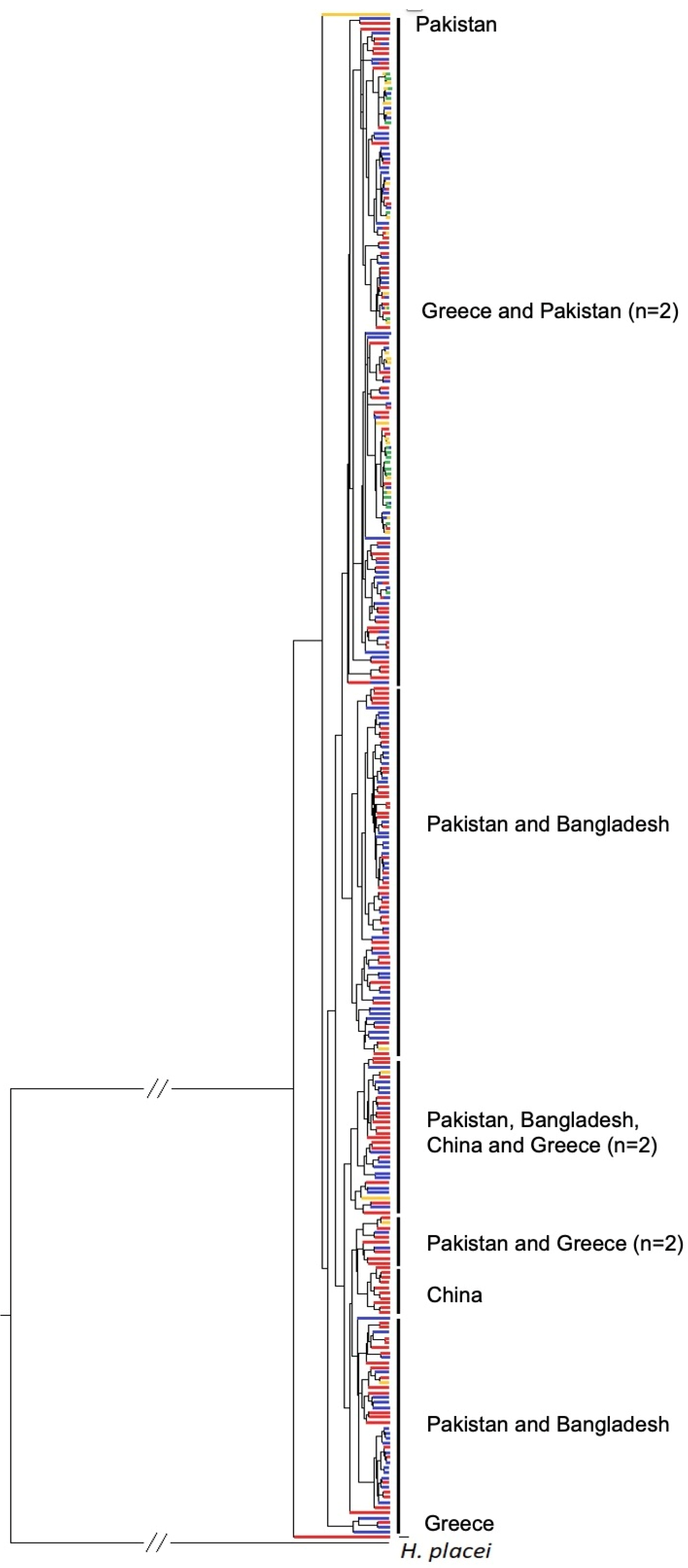
3.4. Phylogenetic Analysis of ND4 Gene
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Soulsby, E.J.L. Helminths, Arthropods and Protozoa of Domestic Animals, 7th ed.; Bailliere Tindall: London, UK, 1982; p. 809. [Google Scholar]
- Perry, B.D.; Randolph, T.F.; McDermott, J.J.; Sones, K.R.; Thorton, P.K. Investing in Animal Health Research to Alleviate Poverty; International Livestock Research Institute: Nairobi, Kenya, 2002; p. 148. [Google Scholar]
- Urquhart, G.M.; Armour, J.; Duncan, J.L.; Dunn, A.M.; Jennings, F.W. Veterinary Parasitology, 8th ed.; Blackwell Science Ltd.: Oxford, UK, 1996; p. 287. [Google Scholar]
- Hoberg, E.P.; Lichtenfels, J.R.; Gibbons, L. Phylogeny for species of the genus Haemonchus (Nematoda: Trichostrongyloidea): Considerations of their evolutionary history and global biogeography among Camelidae and Pecora (Artiodactyla). J. Parasitol. 2004, 90, 1085–1102. [Google Scholar] [CrossRef] [PubMed]
- Riley, D.G.; Van Wyk, J.A. Genetic parameters for FAMACHA score and related traits for host resistance/resilience and production at differing severities of worm challenge in a Merino flock in South Africa. Vet. Parasitol. 2009, 164, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Scheuerle, M.; Mahling, M.; Muntwyler, J.; Pfister, K. The accuracy of the FAMACHA method in detecting anaemia and haemonchosis in goat flocks in Switzerland under field conditions. Vet. Parasitol. 2012, 170, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.E.; Kaplan, R.M.; Pugh, D.G. Internal Parasites. In Sheep and Goat Medicine, 2nd ed.; Pugh, D.G., Baird, A.N., Eds.; Elsevier: London, UK, 2002; pp. 106–125. [Google Scholar]
- McRae, K.; McEwan, J.C.; Dodds, K.G.; Gemmell, N.J. Signatures of selection in sheep bred for resistance or susceptibility to gastrointestinal nematodes. Genomics 2014, 15, 637. [Google Scholar] [CrossRef] [PubMed]
- Papazahariadou, M.G.; Papadopoulos, E.G.; Frydas, S.E.; Mavrovouniotis, C.; Constantinidis, T.C.; Antoniadou-Sotiriadou, K.; Siochu, A.E. Prevalence of gastrointestinal parasites in the Greek population: Local people and refugees. Ann. Gastroenterol. 2004, 17, 194–198. [Google Scholar]
- Githigia, S.M.; Thamsborg, S.M.; Munya, W.K.; Maingi, N. Impact of gastrointestinal helminths on production in goats in Kenya. Small Rumin. Res. 2001, 42, 21–29. [Google Scholar] [CrossRef]
- Theodoridis, Y.; Himonas, C.; Papazahariadou, M. Helminths parasites of digestive tract of sheep and goats in Macedonian region. Bull. Hellenic. Vet. Med. Soc. 2000, 51, 195–199. (In Greek) [Google Scholar] [CrossRef]
- Charalambidis, S.T. Veterinary Parasitology; University Studio Press: Thessaloniki, Greece, 2001; p. 460. (In Greek) [Google Scholar]
- Himonas, C.A. Veterinary Parasitology; AUTH Publication Services: Thessaloniki, Greece, 1970; p. 230. (In Greek) [Google Scholar]
- McCoy, K.D.; Boulinier, T.; Tirard, C.; Mochalakis, Y. Host-dependent genetic structure of parasite populations: Differential dispersal of seabird tick host races. Evolution 2003, 57, 288–296. [Google Scholar]
- Troell, K.; Engstrom, A.; Morriso, D.A.; Mattsson, J.G.; Hoglund, J. Global patterns reveal strong population structure in Haemonchus contortus, a nematode parasite of domesticated ruminants. Int. J. Parasitol. 2006, 36, 1305–1316. [Google Scholar] [CrossRef]
- Lichtenfels, J.R.; Pilitt, P.A.; Hobere, E.P. New morphological characters for identifying individual specimens of Haemonchus spp (Nematoda. trichostrongyloidea) and a key to species in ruminants of North America. J. Parasitol. 1994, 80, 107–119. [Google Scholar]
- Hillis, D.; Mable, B.K.; Larson, A.; Davis, S.K.; Zimmer, E.A. Nucleic acids IV: Sequencing and cloning. In Molecular Systematics; Hills, D., Moritz, C., Mable, B.K., Eds.; Sinauer Associates: Sunderland, UK, 1996; pp. 321–382. [Google Scholar]
- Brasil, B.S.A.F.; Ronaldo, L.; Nunes, R.L.; Bastianetto, E.; Drummond, M.G.; Carvalho, D.C.; Leite, R.C.; Molento, M.B.; Oliveira, D.A.A. Genetic diversity patterns of Haemonchus placei and Haemonchus contortus populations isolated from domestic ruminants in Brazil. Int. J. Parasitol. 2021, 42, 469–479. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Thomson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Rozas, R. DnaSP version 3: An integrated program for molecular population genetics and molecular evolution analysis. Bioinformatics 1999, 15, 174–175. [Google Scholar] [CrossRef]
- Dey, A.R.; Zhang, Z.; Begum, N.; Abdul-Alim, M.D.; Hu, M.; Alam, M.Z. Genetic diversity patterns of Haemonchus contortus isolated from sheep and goats in Bangladesh. Infect. Genet. Evol. 2019, 68, 177–184. [Google Scholar] [CrossRef]
- Hussain, T.; Periasamy, K.; Nadeem, A.; Ellahi, M.B.; Pichler, R.; Diallo, A. Sympatric species distribution, genetic diversity and population structure of Haemonchus isolates from domestic ruminants in Pakistan. Vet. Parasitol. 2014, 206, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Shen, D.; Wang, J.; Zhang, D.; Peng, Z.; Yang, T.; Wang, Z.; Bowman, D.D.; Hou, Z.; Liu, Z. Genetic diversity of Haemonchus contortus isolated from sympatric wild blue sheep (Pseudois nayaur) and sheep in Helan Mountains, China. Parasites Vectors 2017, 10, 437. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Gascuel, O. A simple, fast and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef]
- Bandelt, H.; Forster, P.; Rohl, A. Median Joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, L.A.; Chilton, N.B.; Gasser, R.B. Differentiation of Haemonchus placei from H. contortus (Nematoda: Trichostrongylidae) by the ribosomal DNA second internal transcribed spacer. Int. J. Parasitol. 1995, 25, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.C.; Amarante, M.R.V.; Amarante, A.F.T. Is there competition between Haemonchus contortus and Haemonchus placei in a pasture grazed by only sheep? Vet. Parasitol. 2020, 279, 109054. [Google Scholar] [CrossRef]
- Hoberg, E.P.; Zarlenga, D.S. Evolution and biogeography of Haemonchus contortus: Linking faunal dynamics in space and time. Adv. Parasitol. 2016, 93, 1–30. [Google Scholar]
- Chaundry, U.; Miller, M.; Yazwinski, T.; Kaplan, R.; Gilleard, J. The presence of benzimidazole resistance mutations in Haemonchus placei from US cattle. Vet. Parasitol. 2014, 204, 411–415. [Google Scholar]
- Akkari, H.; Jebali, J.; Gharbi, M.; Mhadhbi, M.; Awadi, S.; Darghouth, M.A. Epidemiological study of sympatric Haemonchus species and genetic characterization of Haemonchus contortus in domestic ruminants in Tunisia. Vet. Parasitol. 2013, 193, 118–125. [Google Scholar] [CrossRef]
- Gelasakis, A.I.; Valergakis, G.E.; Fortomaris, P.; Arsenos, G. Farm conditions and production methods in Chios sheep flocks. J. Hellenic. Vet. Med. Soc. 2010, 61, 111–119. [Google Scholar] [CrossRef]
- Gharamah, A.A.; Siti Azizah, M.N.; Rahmanv, W.A. Genetic variation of Haemonchus contortus (Trichostrongylidae) in sheep and goats from Malaysia and Yemen. Vet. Parasitol. 2012, 188, 268–276. [Google Scholar] [CrossRef]
- Yin, F.; Gasser, R.B.; Li, F.; Bao, M.; Huang, W.; Zou, F.; Zhao, G.; Wang, C.; Yang, X.; Zhou, Y.; et al. Genetic variability within and among Haemonchus contortus isolates from goats and sheep in China. Parasites Vectors 2013, 6, 279. [Google Scholar] [CrossRef]
- Cerutti, M.C.; Citterio, C.V.; Bazzocchi, C.; Epis, S.; D’Amelio, S.; Ferrari, N.; Lanfranchi, P. Genetic variability of Haemonchus contortus (Nematoda: Trichostrongyloidea) in alpine ruminant host species. J. Helminthol. 2012, 84, 276–283. [Google Scholar] [CrossRef]
- Blouin, M.S.; Yowell, C.A.; Courtney, C.H.; Dame, J.B. Host movement and the genetic structure of populations of parasitic nematodes. Genetics 1995, 141, 1007–1014. [Google Scholar] [CrossRef]
- Charlesworth, B. Fundamental concepts in genetics: Effective population size and patterns of molecular evolution and variation. Nat. Rev. Genet. 2009, 10, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Prichard, R. Genetic variability following selection of Haemonchus contortus with anthelmintics. Trends Parasitol. 2001, 17, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Silvestre, A.; Sauve, C.; Cortet, J.; Cabaret, J. Contrasting genetic structures of two parasitic nematodes, determined on the basis of neutral microsatellite markers and selected anthelmintic resistance markers. Mol. Ecol. 2009, 18, 5086–5100. [Google Scholar] [CrossRef]
- Chaundry, U.; Redman, E.M.; Raman, M.; Gilleard, J.S. Genetic evidence for the spread of a benzimidazole resistance mutation across southern India from a single origin in the parasitic nematode Haemonchus contortus. Int. J. Parasitol. 2015, 45, 721–728. [Google Scholar]
- Redman, E.; Whitelaw, F.; Tait, A.; Burgess, C.; Bartley, Y.; Skuce, P.J.; Jackson, F.; Gilleard, J.S. The emergence of resistance to the benzimidazole anthelmintics in parasitic nematodes of livestock is characterised by multiple independent hard and soft selective sweeps. PLoS Neglected Trop. Dis. 2015, 9, e0003494. [Google Scholar] [CrossRef]
- Gennari, S.M.; Vieira Bressan, M.C.R.; Rogero, J.R.; MacLean, J.M.; Duncan, J.L. Pathophysiology of Haemonchus placei infection in calves. Vet. Parasitol. 1991, 38, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Bremner, K.C. The parasitic life-cycle of Haemonchus placei (Place) (Nematoda: Trichostrongyloidea). Aust. J. Zool. 1956, 4, 146–151. [Google Scholar] [CrossRef]
- Riggs, N.L. Experimental cross-infections of Haemonchus placei (Place, 1983) in sheep and cattle. Vet. Parasitol. 2001, 94, 191–197. [Google Scholar] [CrossRef]
- Arsenopoulos, K.V.; Minoudi, S.; Symeonidou, I.; Triantafyllidis, A.; Katsafadou, A.I.; Lianou, D.T.; Fthenakis, G.C.; Papadopoulos, E. Frequency of resistance to benzimidazoles of Haemonchus contortus helminths from dairy sheep, goats, cattle and buffaloes in Greece. Pathogens 2020, 9, 347. [Google Scholar] [CrossRef]
- Nikolakaki, S.V.; Nomikou, K.; Koumbati, M.; Mangana, O.; Papanastassopoulou, M.; Mertens, P.P.C.; Papadopoulos, O. Molecular analysis of the NS3/NS3A gene of Bluetongue virus isolates from the 1979 and 1998–2001 epizootics in Greece and their segregation into two distinct groups. Virus Res. 2005, 114, 6–14. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | Product Size (bp) |
|---|---|---|
| NC1-F | forward: 5′- ACGTCTGGTTCAGGGTTGTT -3′ | 321 |
| NC2-R | reverse: 5′- TTAGTTTCTTTTCCTCCGCT -3′ | |
| OP1-F | forward: 5′- GGATTTGGTCAGCAAATTGAA -3′ | 820 |
| OP2-R | reverse: 5′- TCATTTGTGGTTACCTAAAGC -3′ |
| Accession Numbers | Number of Sequences | Hosts | Origin | Reference | Dataset |
|---|---|---|---|---|---|
| LC361049–LC361102, LC376827–LC376849 | 77 | Sheep & goats | Bangladesh | Dey et al. [22] | 2 |
| KJ724439–KJ724511 | 73 | Sheep, goats & cattle | Pakistan | Hussain et al. [23] | 2 |
| KY305790–KY305808 | 19 | Sheep | China | Shen et al. [24] | 2 |
| Genotype | Position | Number of Parasites | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 59 | 63 | 78 | 123 | 187 | 196 | B | C | G | S | Total | |
| X78803 | T | C | T | C | G | A | - | - | - | - | - |
| GT1 | . | . | . | T | G | T | 3 a | 3 a | 17 b | 14 b | 37 |
| GT2 | . | . | . | T | R | T | 25 a | 28 a | 17 b | 25 a | 95 |
| GT3 | . | . | . | T | R | W | 1 a | 1 a | 2 b | - | 4 |
| GT4 | . | . | . | T | . | W | 1 a | - | 1 a | - | 2 |
| GT5 | . | . | . | Y | . | W | - | 3 a | 4 b | 2 a | 9 |
| GT6 | . | . | Y | Y | . | T | - | - | 3 | - | 3 |
| GT7 | . | . | Y | Y | . | W | - | 1 a | 2 a | 4 b | 7 |
| GT8 | . | . | Y | T | . | T | - | - | 1 a | 3 a | 4 |
| GT9 | . | . | . | Y | . | T | 1 a | - | 2 a | - | 3 |
| GT10 | . | . | Y | Y | R | W | - | 1 a | 2 a | 2 a | 5 |
| GT11 | . | Y | Y | Y | R | W | - | - | 1 | - | 1 |
| GT12 | . | . | . | Y | R | W | 1 a | - | 2 a | 1 a | 4 |
| GT13 | K | Y | Y | Y | . | T | 4 a | 1 b | 2 b | 2 b | 9 |
| GT14 | . | . | Y | T | R | T | - | - | 1 | - | 1 |
| GT15 | K | Y | . | Y | . | W | 1 a | - | 1 a | - | 2 |
| GT16 | K | Y | Y | Y | R | W | 1 a | - | 1 a | - | 2 |
| GT17 | . | Y | . | Y | . | W | - | 1 | - | - | 1 |
| GT18 | . | . | . | Y | R | T | 1 | - | - | - | 1 |
| GT19 | . | Y | Y | Y | . | W | - | - | - | 1 | 1 |
| Total | 39 a | 39 a | 59 b | 54 b | 191 | ||||||
| Ruminants | Dataset | Base Pairs (bp) | Ν | H | Hp | V (%) | W (%) | Hd (SD) | Nd (SD) |
|---|---|---|---|---|---|---|---|---|---|
| Buffaloes | 1 | 690 | 24 | 22 | 13 | 91.7 | 54.2 | 0.989 a (0.017) | 0.007 a (0.001) |
| 2 | 673 | - | - | - | - | - | - | - | |
| Cattle | 1 | 690 | 33 | 25 | 14 | 75.8 | 42.4 | 0.966 b (0.022) | 0.007 a (0.001) |
| 2 | 673 | 38 | 30 | 18 | 78.9 | 47.4 | 0.974 A (0.017) | 0.012 A (0.002) | |
| Goats | 1 | 690 | 63 | 61 | 42 | 96.8 | 66.7 | 0.999 c (0.003) | 0.019 b (0.001) |
| 2 | 673 | 134 | 131 | 112 | 97.8 | 83.6 | 0.999 B (0.001) | 0.029 B (0.001) | |
| Sheep | 1 | 690 | 74 | 59 | 48 | 79.7 | 64.9 | 0.997 c (0.003) | 0.021 c (0.001) |
| 2 | 673 | 167 | 151 | 140 | 90.4 | 83.8 | 0.999 B (0.001) | 0.029 B (0.001) | |
| Total samples | 1 | 690 | 194 | 140 | 117 | 72.2 | 60.3 | 0.995 c (0.002) | 0.017 b (0.001) |
| 2 | 673 | 363 | 306 | 282 | 84.3 | 77.7 | 0.998 B (0.001) | 0.026 B (0.001) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arsenopoulos, K.V.; Minoudi, S.; Symeonidou, I.; Triantafyllidis, A.; Fthenakis, G.C.; Papadopoulos, E. Extensive Countrywide Molecular Identification and High Genetic Diversity of Haemonchus spp. in Domestic Ruminants in Greece. Pathogens 2024, 13, 238. https://doi.org/10.3390/pathogens13030238
Arsenopoulos KV, Minoudi S, Symeonidou I, Triantafyllidis A, Fthenakis GC, Papadopoulos E. Extensive Countrywide Molecular Identification and High Genetic Diversity of Haemonchus spp. in Domestic Ruminants in Greece. Pathogens. 2024; 13(3):238. https://doi.org/10.3390/pathogens13030238
Chicago/Turabian StyleArsenopoulos, Konstantinos V., Styliani Minoudi, Isaia Symeonidou, Alexandros Triantafyllidis, George C. Fthenakis, and Elias Papadopoulos. 2024. "Extensive Countrywide Molecular Identification and High Genetic Diversity of Haemonchus spp. in Domestic Ruminants in Greece" Pathogens 13, no. 3: 238. https://doi.org/10.3390/pathogens13030238