
Integration of Metabolomics and Transcriptomics to Reveal the Metabolic Characteristics of Exercise-Improved Bone Mass

Abstract
:1. Introduction
2. Materials and Methods
2.1. Zebrafish Husbandry
2.2. Zebrafish Exercise Training
2.3. Staining of Bone Tissue and Quantification of Bone Mineralization
2.4. Quantification of Mineralization in Scales
2.5. Biochemical Analyses of Alp and Trap Activities in Scales
2.6. Histological Staining of Alp and Trap in Scales
2.7. Statistical Analysis of Bone Phenotypes
2.8. Metabolomics
2.8.1. Metabolite Extraction from Bone Tissue
2.8.2. Bone Tissue Metabolic Profiling Analysis by Ultra-High-Performance Liquid Chromatography-Tandem Mass Spectrometry (UPLC-MS/MS)
2.8.3. Bone Tissue Metabolomic Data Processing and Annotation
2.9. Transcriptomics
2.9.1. Sample Preparation and RNA Extraction
2.9.2. Library Construction and RNA Sequencing
2.9.3. Read Filtering and Mapping
2.9.4. Analysis of Differentially Expressed Genes (DEGs)
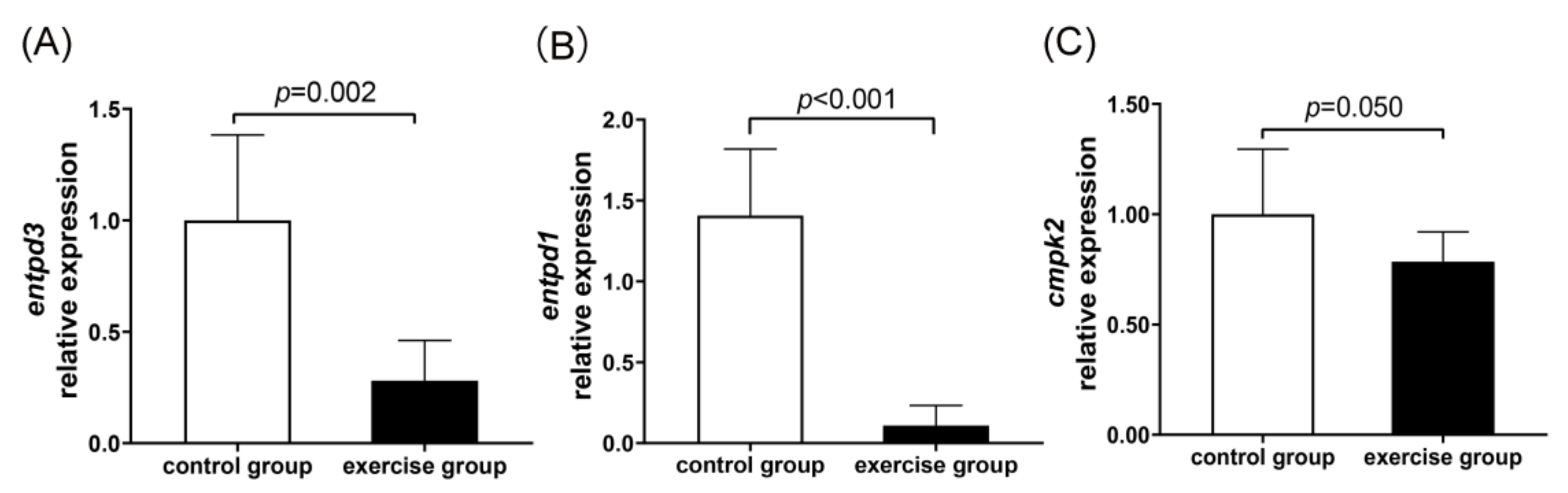
2.9.5. Validation of Transcriptome Analysis Using Quantitative Real-Time PCR (qRT-PCR)
3. Results
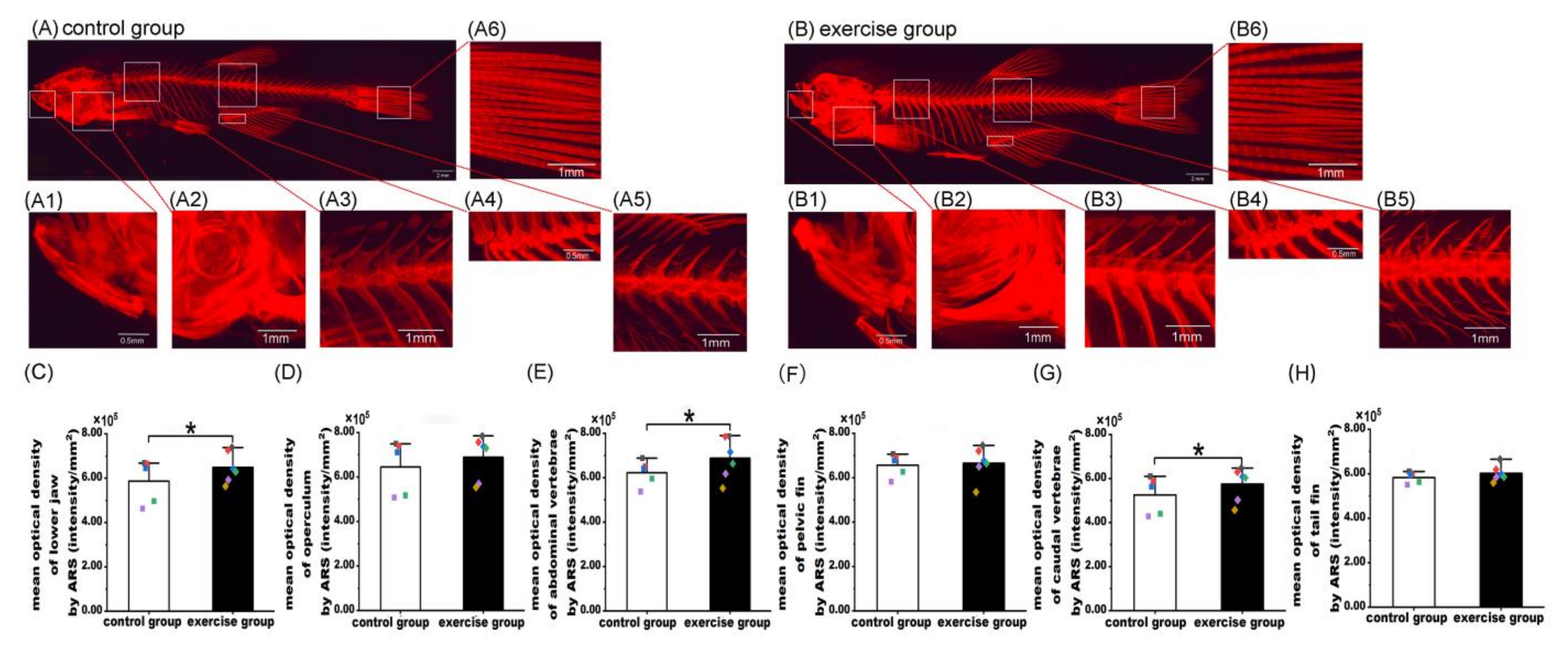
3.1. Exercise Increased Zebrafish Bone Mass
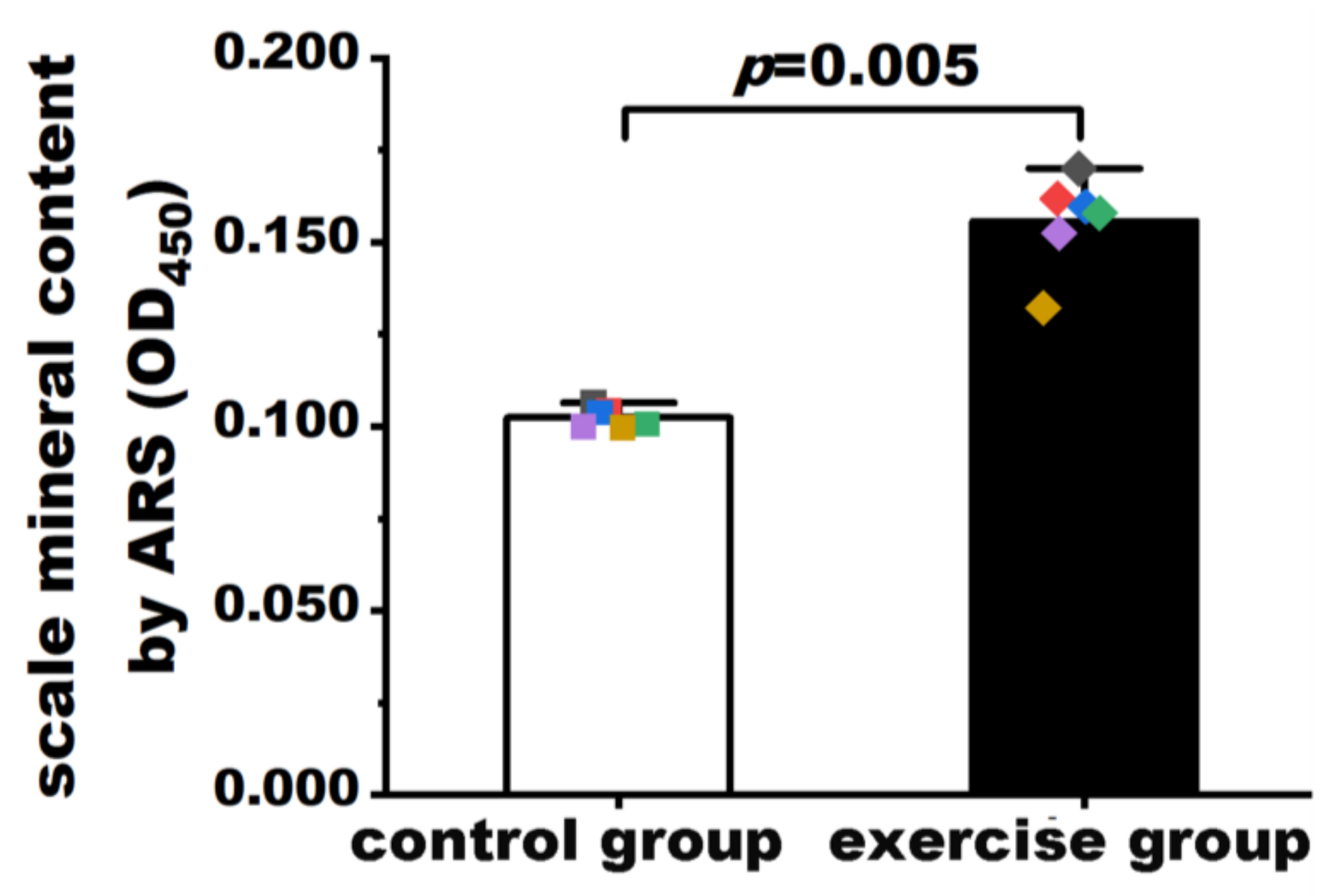
3.2. Exercise Increased Mineralization in Zebrafish Scales
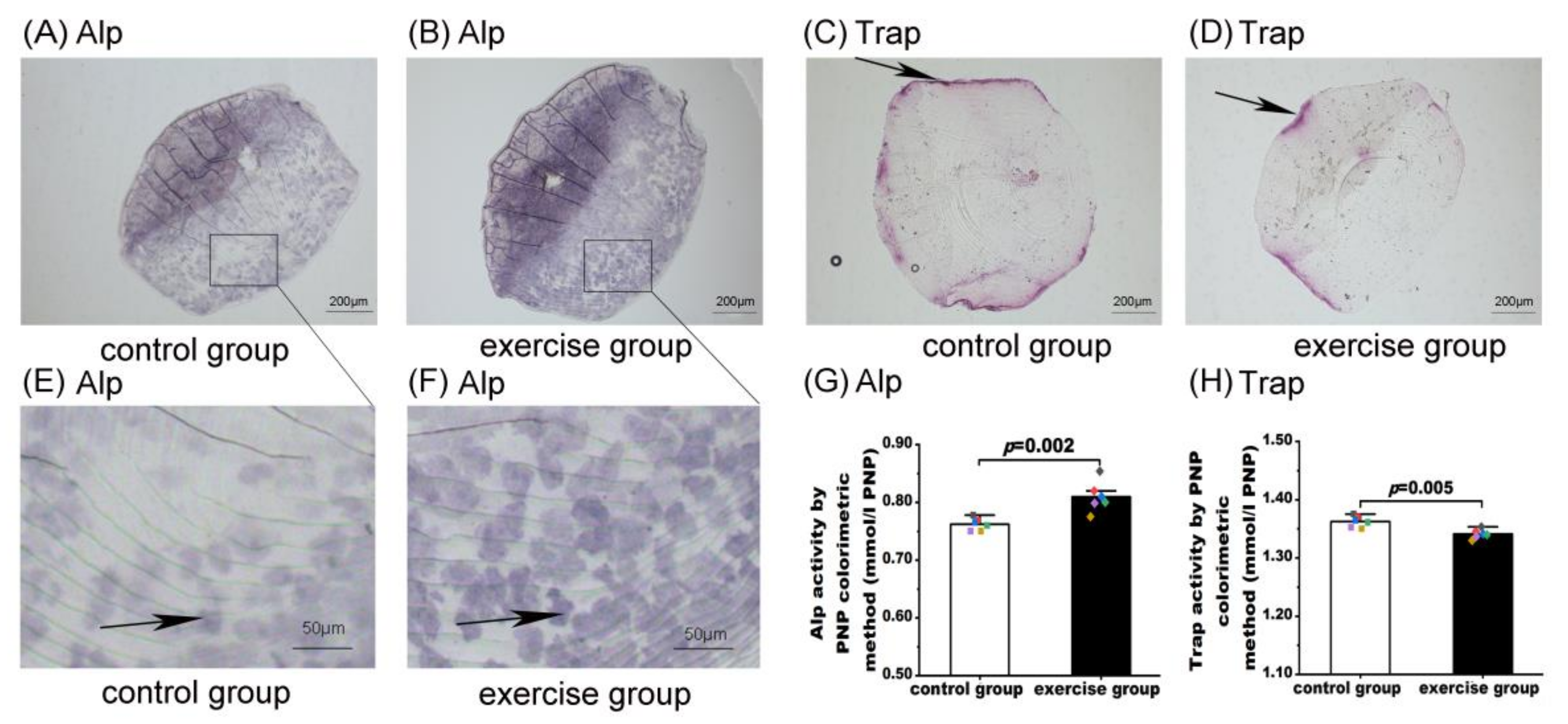
3.3. Exercise Induced Alp Activity and Inhibited Trap Activity in Zebrafish
3.4. Metabolic Profiling of the Two Groups
3.5. DMs between the Two Groups
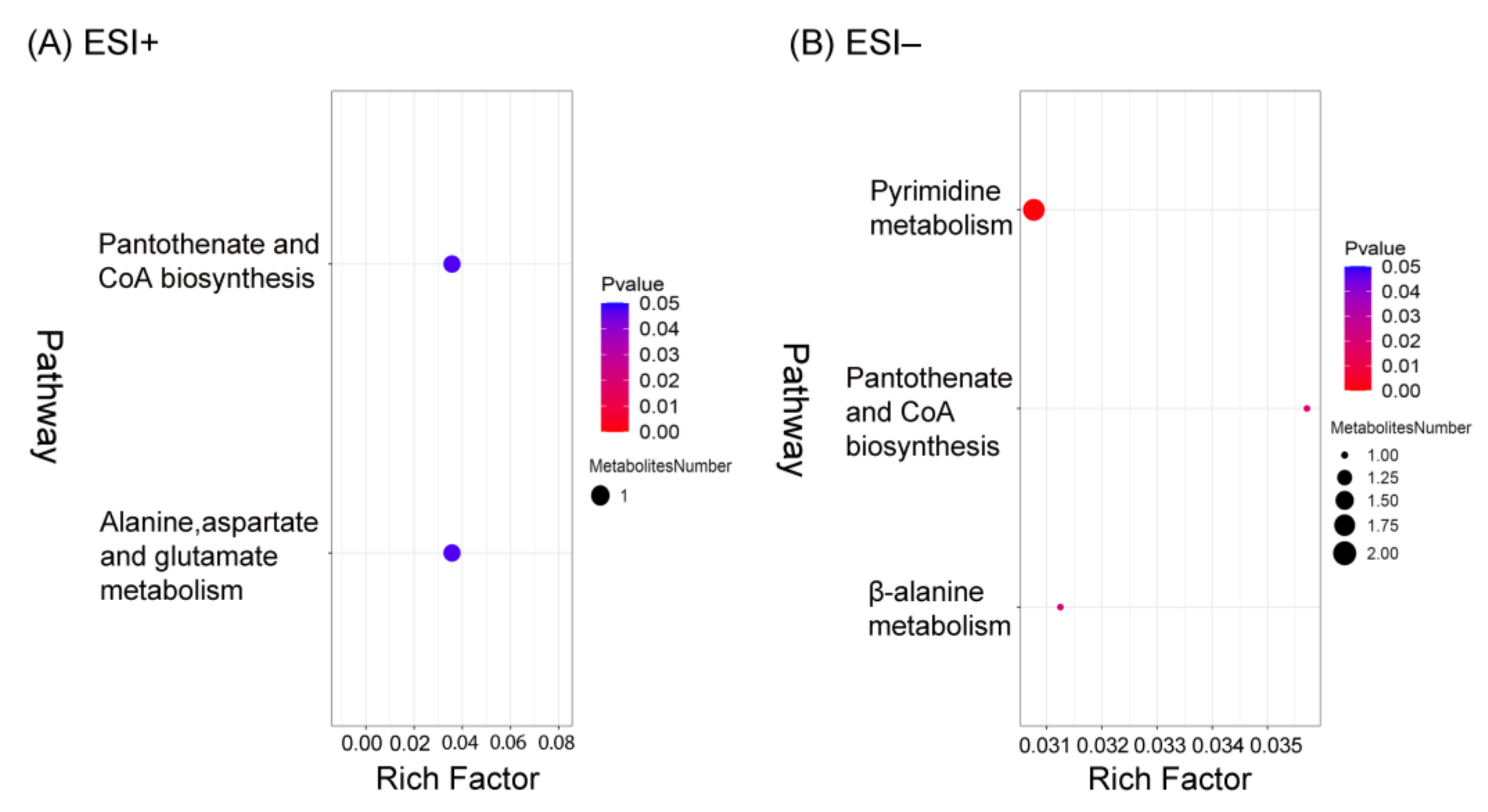
3.6. Pathway Analysis
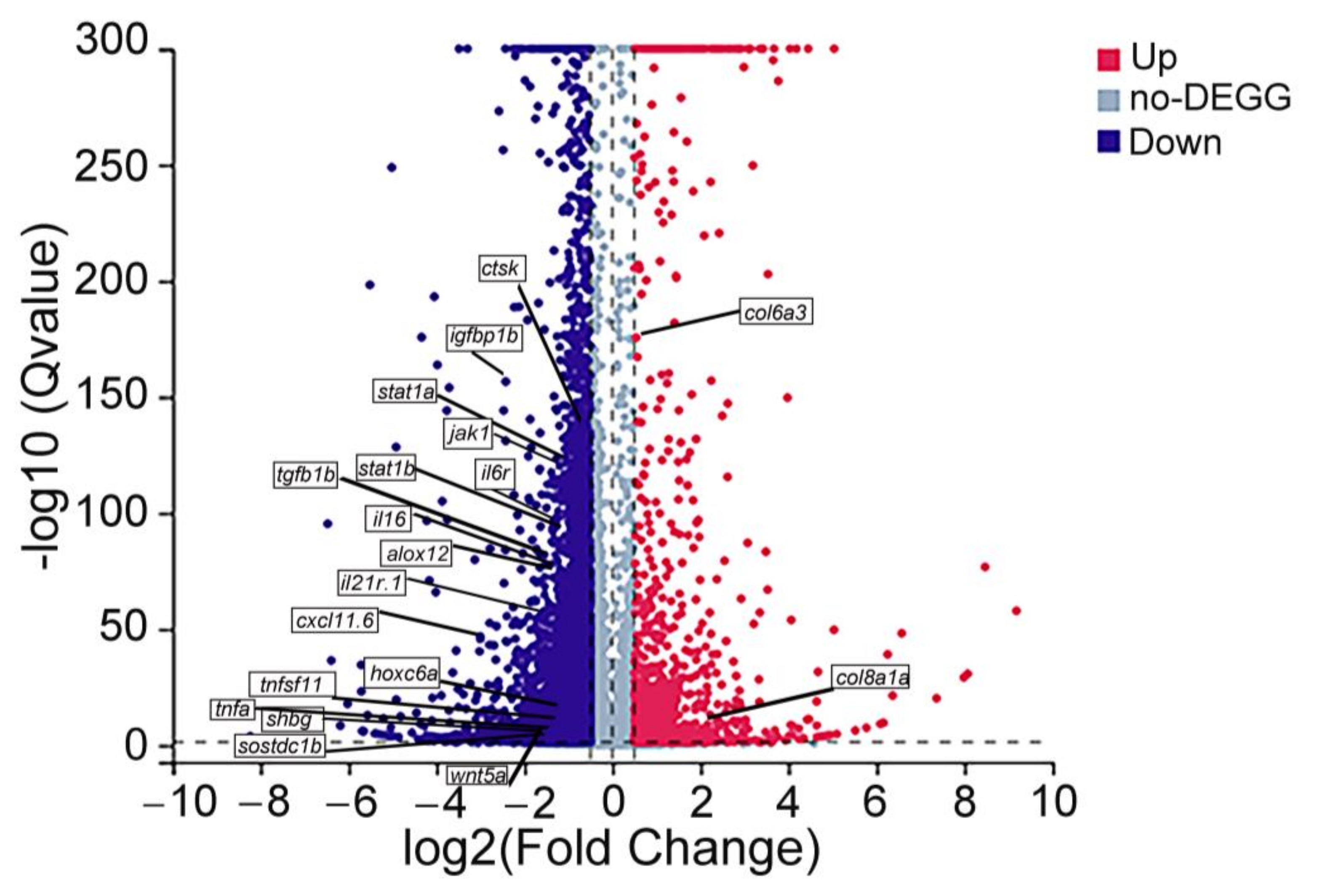
3.7. Analysis of Transcriptomic Data
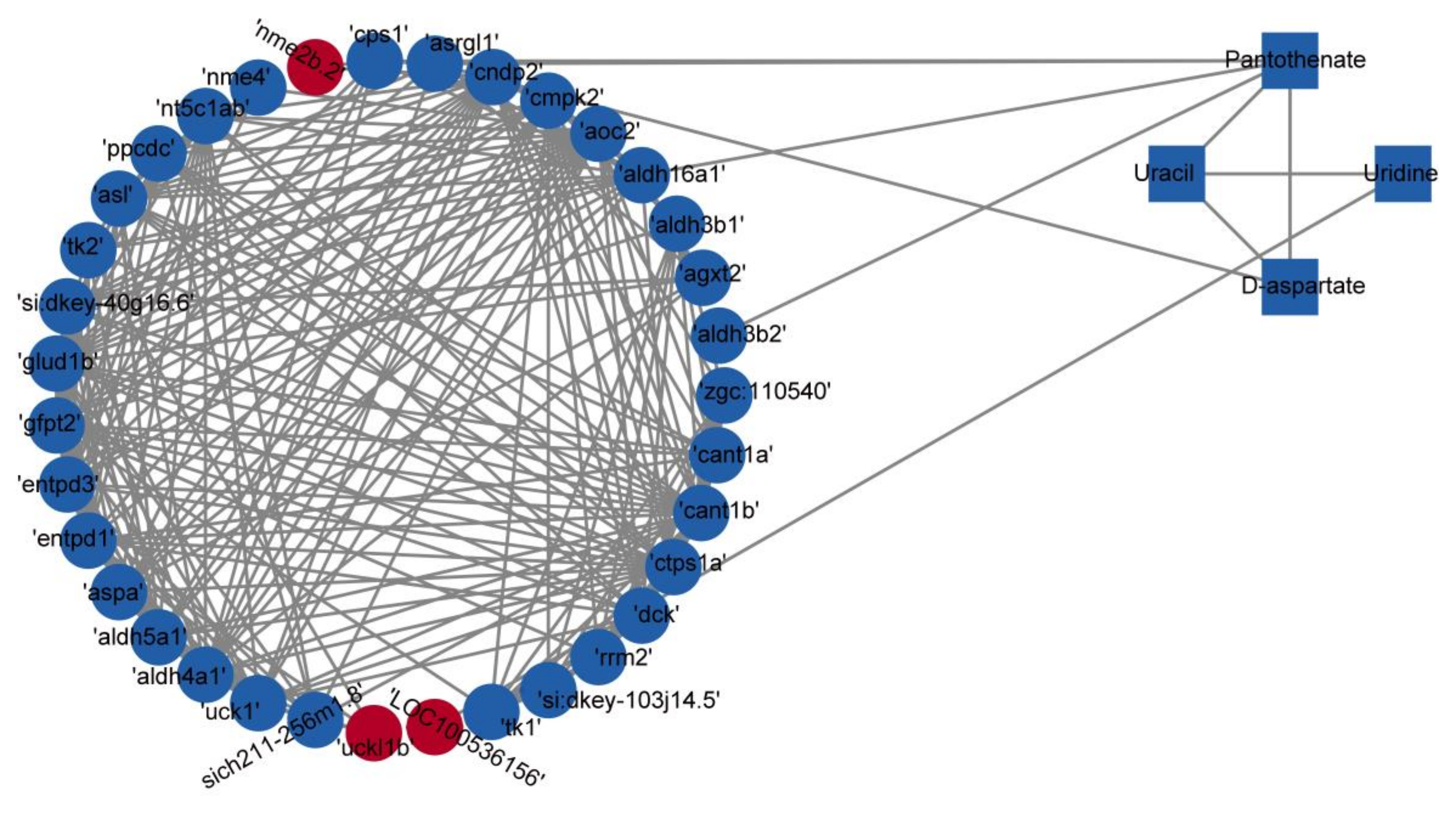
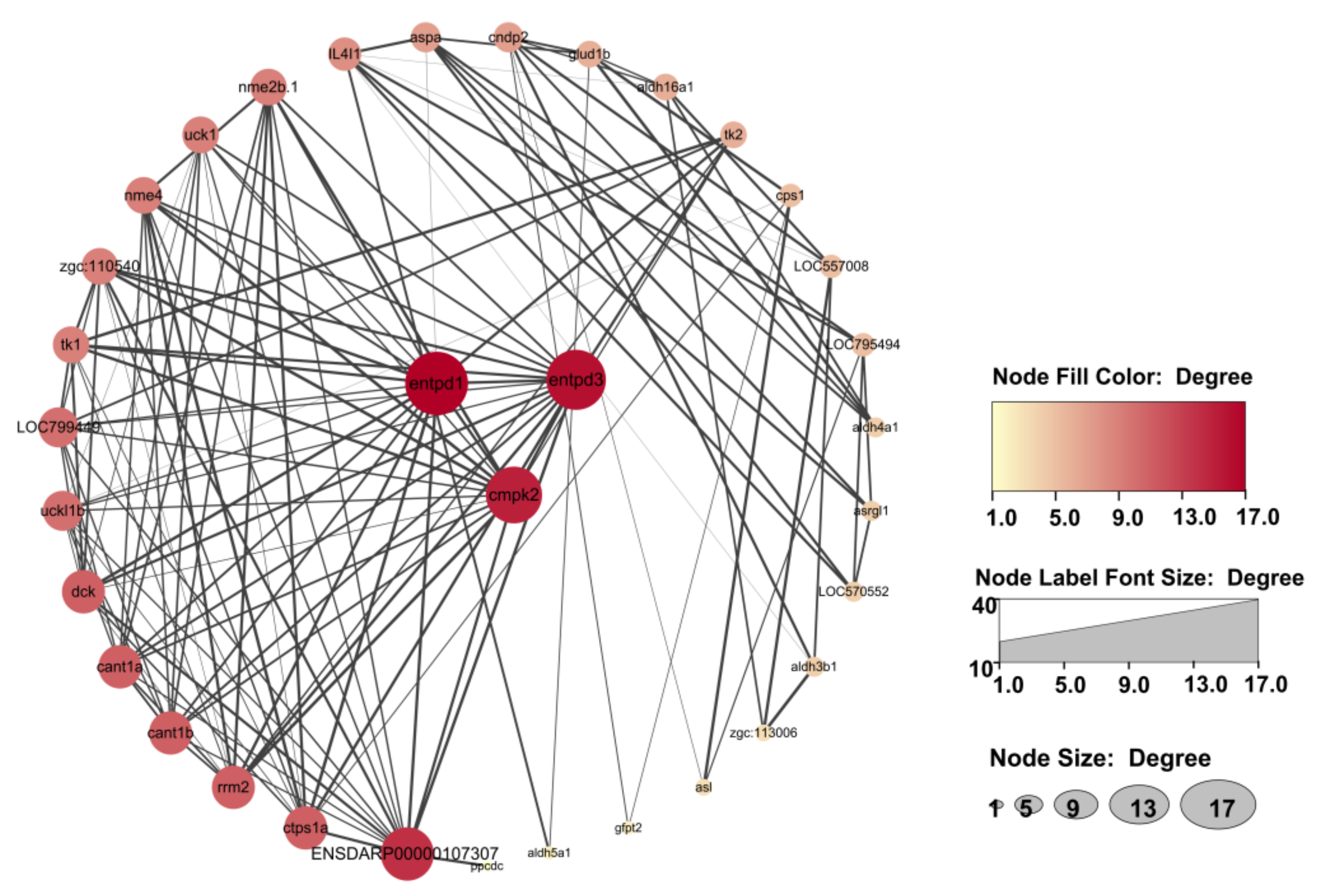
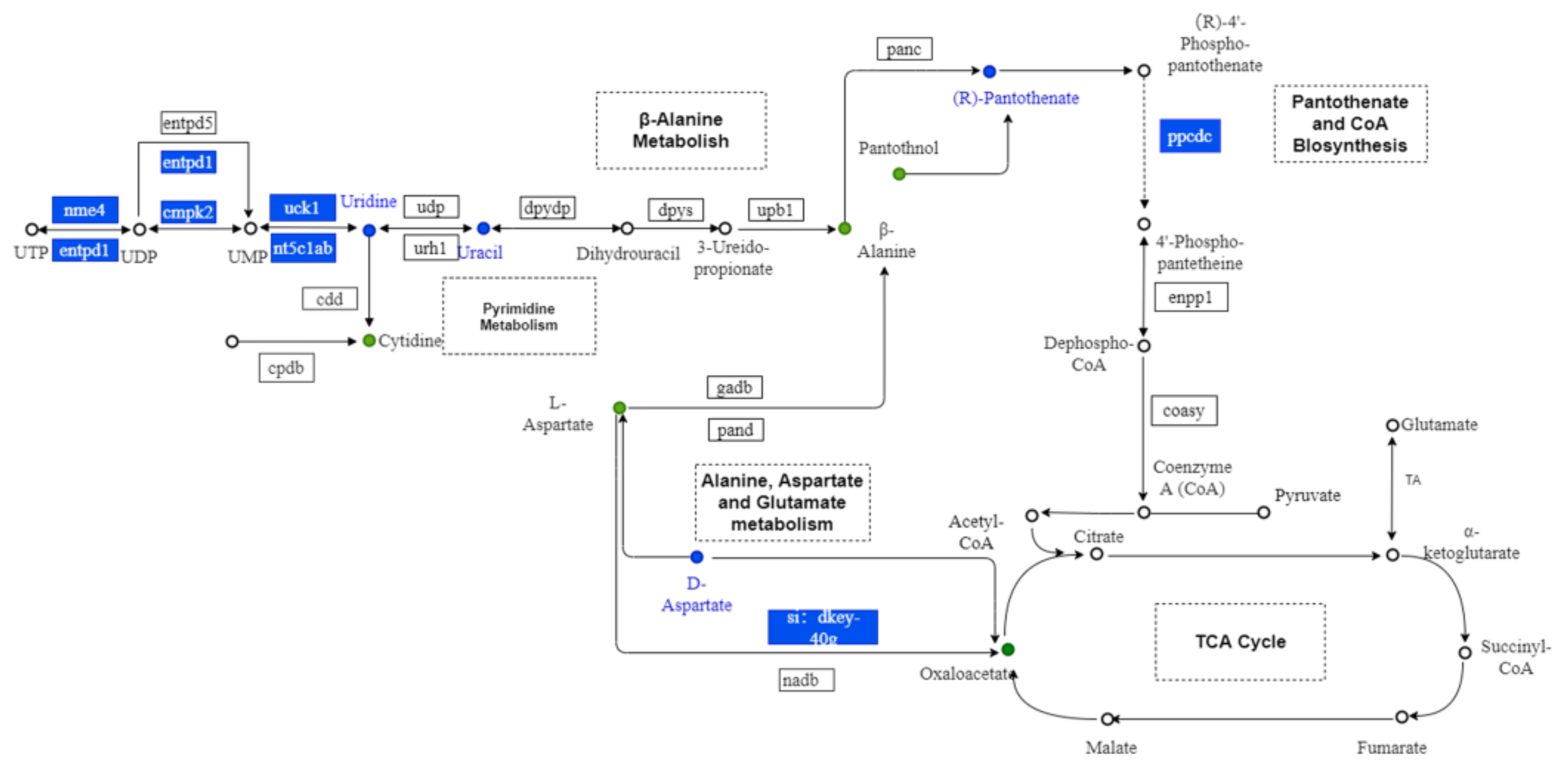
3.8. Integrated Analysis of Metabolomic and Transcriptomic Data
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Buck, D.W., 2nd; Dumanian, G.A. Bone biology and physiology: Part I. The fundamentals. Plast. Reconstr. Surg. 2012, 129, 1314–1320. [Google Scholar] [CrossRef] [PubMed]
- Mosekilde, L. Age-related changes in bone mass, structure, and strength—Effects of loading. Z. Rheumatol. 2000, 59 (Suppl. 1), 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, C.J.; Beaupré, G.S.; Carter, D.R. A theoretical analysis of the relative influences of peak BMD, age-related bone loss and menopause on the development of osteoporosis. Osteoporos. Int. 2003, 14, 843–847. [Google Scholar] [CrossRef] [PubMed]
- Welsh, L.; Rutherford, O.M. Hip bone mineral density is improved by high-impact aerobic exercise in postmenopausal women and men over 50 years. Eur. J. Appl. Physiol. Occup. Physiol. 1996, 74, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Bruton, A.; Matute-Llorente, Á.; González-Agüero, A.; Casajús, J.A.; Vicente-Rodríguez, G. Plyometric exercise and bone health in children and adolescents: A systematic review. World J. Pediatr. 2017, 13, 112–121. [Google Scholar] [CrossRef]
- Mages, M.; Shojaa, M.; Kohl, M.; von Stengel, S.; Becker, C.; Gosch, M.; Jakob, F.; Kerschan-Schindl, K.; Kladny, B.; Klöckner, N.; et al. Exercise effects on bone mineral density in men. Nutrients 2021, 13, 4244. [Google Scholar] [CrossRef]
- Ponzano, M.; Rodrigues, I.B.; Hosseini, Z.; Ashe, M.C.; Butt, D.A.; Chilibeck, P.D.; Stapleton, J.; Thabane, L.; Wark, J.D.; Giangregorio, L.M. Progressive resistance training for improving health-related outcomes in people at risk of fracture: A systematic review and meta-analysis of randomized controlled trials. Phys. Ther. 2021, 101, pzaa221. [Google Scholar] [CrossRef]
- Ravnholt, T.; Tybirk, J.; Jørgensen, N.R.; Bangsbo, J. High-intensity intermittent “5-10-15” running reduces body fat, and increases lean body mass, bone mineral density, and performance in untrained subjects. Eur. J. Appl. Physiol. 2018, 118, 1221–1230. [Google Scholar] [CrossRef]
- Giannotti, S.; Bottai, V.; Dell’osso, G.; De Paola, G.; Bugelli, G.; Pini, E.; Guido, G. Disuse osteoporosis of the upper limb: Assessment of thirty patients. Clin. Cases Miner. Bone Metab. 2013, 10, 129–132. [Google Scholar]
- Alexandre, C.; Vico, L. Pathophysiology of bone loss in disuse osteoporosis. Jt. Bone Spine 2011, 78, 572–576. [Google Scholar] [CrossRef]
- Starnes, J.W.; Parry, T.L.; O’Neal, S.K.; Bain, J.R.; Muehlbauer, M.J.; Honcoop, A.; Ilaiwy, A.; Christopher, P.M.; Patterson, C.; Willis, M.S. Exercise-induced alterations in skeletal muscle, heart, liver, and serum metabolome identified by non-targeted metabolomics analysis. Metabolites 2017, 7, 40. [Google Scholar] [CrossRef]
- Fiorenza, M.; Hostrup, M.; Gunnarsson, T.P.; Shirai, Y.; Schena, F.; Iaia, F.M.; Bangsbo, J. Neuromuscular fatigue and metabolism during high-intensity intermittent exercise. Med. Sci. Sports Exerc. 2019, 51, 1642–1652. [Google Scholar] [CrossRef]
- Gaitán, J.M.; Moon, H.Y.; Stremlau, M.; Dubal, D.B.; Cook, D.B.; Okonkwo, O.C.; Van Praag, H. Effects of aerobic exercise training on systemic biomarkers and cognition in late middle-aged adults at risk for Alzheimer’s disease. Front. Endocrinol. 2021, 12, 660181. [Google Scholar] [CrossRef]
- Isung, J.; Granqvist, M.; Trepci, A.; Huang, J.; Schwieler, L.; Kierkegaard, M.; Erhardt, S.; Jokinen, J.; Piehl, F. Differential effects on blood and cerebrospinal fluid immune protein markers and kynurenine pathway metabolites from aerobic physical exercise in healthy subjects. Sci. Rep. 2021, 11, 1669. [Google Scholar] [CrossRef]
- Zhou, W.; Zeng, G.; Lyu, C.; Kou, F.; Zhang, S.; Wei, H. The effect of exhaustive exercise on plasma metabolic profiles of male and female rats. J. Sports Sci. Med. 2019, 18, 253–263. [Google Scholar]
- Sabatine, M.S.; Liu, E.; Morrow, D.A.; Heller, E.; McCarroll, R.; Wiegand, R.; Berriz, G.F.; Roth, F.P.; Gerszten, R.E. Metabolomic identification of novel biomarkers of myocardial ischemia. Circulation 2005, 112, 3868–3875. [Google Scholar] [CrossRef]
- Lv, H.; Jiang, F.; Guan, D.; Lu, C.; Guo, B.; Chan, C.; Peng, S.; Liu, B.; Guo, W.; Zhu, H.; et al. Metabolomics and its application in the development of discovering biomarkers for osteoporosis research. Int. J. Mol. Sci. 2016, 17, 2018. [Google Scholar] [CrossRef]
- Zhao, Q.; Shen, H.; Su, K.J.; Zhang, J.G.; Tian, Q.; Zhao, L.J.; Qiu, C.; Zhang, Q.; Garrett, T.J.; Liu, J.; et al. Metabolomic profiles associated with bone mineral density in US Caucasian women. Nutr. Metab. 2018, 15, 57. [Google Scholar] [CrossRef]
- Busse, B.; Galloway, J.L.; Gray, R.S.; Harris, M.P.; Kwon, R.Y. Zebrafish: An emerging model for orthopedic research. J. Orthop. Res. 2020, 38, 925–936. [Google Scholar] [CrossRef]
- Fiaz, A.W.; Léon-Kloosterziel, K.M.; Gort, G.; Schulte-Merker, S.; van Leeuwen, J.L.; Kranenbarg, S. Swim-training changes the spatio-temporal dynamics of skeletogenesis in zebrafish larvae (Danio rerio). PLoS ONE 2012, 7, e34072. [Google Scholar] [CrossRef]
- Suniaga, S.; Rolvien, T.; Vom Scheidt, A.; Fiedler, I.A.K.; Bale, H.A.; Huysseune, A.; Witten, P.E.; Amling, M.; Busse, B. Increased mechanical loading through controlled swimming exercise induces bone formation and mineralization in adult zebrafish. Sci. Rep. 2018, 8, 3646. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, K.; Fiedler, I.A.; Kurzyukova, A.; López-Delgado, A.C.; McGowan, L.M.; Geurtzen, K.; Hammond, C.L.; Busse, B.; Knopf, F. Skeletal biology and disease modeling in zebrafish. J. Bone Miner. Res. 2021, 36, 436–458. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Gut, P.; Reischauer, S.; Stainier, D.Y.R.; Arnaout, R. Little fish, big data: Zebrafish as a model for cardiovascular and metabolic disease. Physiol. Rev. 2017, 97, 889–938. [Google Scholar] [CrossRef]
- Gonzales, J.M., Jr. Preliminary evaluation on the effects of feeds on the growth and early reproductive performance of zebrafish (Danio rerio). J. Am. Assoc. Lab. Anim. Sci. JAALAS 2012, 51, 412–417. [Google Scholar]
- Westerfield, M. The Zebrafish Book. A Guide for the Laboratory Use of Zebrafish (Danio rerio), 4th ed.; University of Oregon Press: Eugene, OR, USA, 2000; p. 1. [Google Scholar]
- Wise, T.N.; Schwalbe, M.A.B.; Tytell, E.D. Hydrodynamics of linear acceleration in bluegill sunfish, Lepomis macrochirus. J. Exp. Biol. 2018, 221 Pt 23, jeb190892. [Google Scholar] [CrossRef]
- Luo, S.Y.; Chen, J.F.; Zhong, Z.G.; Lv, X.H.; Yang, Y.J.; Zhang, J.J.; Cui, L. Salvianolic acid B stimulates osteogenesis in dexamethasone-treated zebrafish larvae. Acta Pharmacol. Sin. 2016, 37, 1370–1380. [Google Scholar] [CrossRef]
- Fleming, A.; Sato, M.; Goldsmith, P. High-throughput in vivo screening for bone anabolic compounds with zebrafish. J. Biomol. Screen. 2005, 10, 823–831. [Google Scholar] [CrossRef]
- Pasqualetti, S.; Congiu, T.; Banfi, G.; Mariotti, M. Alendronate rescued osteoporotic phenotype in a model of glucocorticoid-induced osteoporosis in adult zebrafish scale. Int. J. Exp. Pathol. 2015, 96, 11–20. [Google Scholar] [CrossRef]
- Yin, H.; Wang, J.; Wu, M.; Ma, Y.; Wang, S.; Su, Q. Preventive effects of evodiamine on dexamethasone-induced osteoporosis in zebrafish. Biomed. Res. Int. 2019, 2019, 5859641. [Google Scholar] [CrossRef]
- Carnovali, M.; Banfi, G.; Mariotti, M. Age-dependent modulation of bone metabolism in zebrafish scales as new model of male osteoporosis in lower vertebrates. Geroscience 2021, 43, 927–940. [Google Scholar] [CrossRef]
- Barker, M.L.; Rayens, W. Partial least squares for discrimination. J. Chemom. A J. Chemom. Soc. 2003, 17, 166–173. [Google Scholar] [CrossRef]
- Westerhuis, J.A.; Hoefsloot, H.C.J.; Smit, S.; Vis, D.J.; Smilde, A.K.; van Velzen, E.J.J.; van Duijnhoven, J.P.; van Dorsten, F.A. Assessment of PLSDA cross validation. Metabolomics 2008, 4, 81–89. [Google Scholar] [CrossRef]
- Li, R.; Li, Y.; Kristiansen, K.; Wang, J. SOAP: Short oligonucleotide alignment program. Bioinformatics 2008, 24, 713–714. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Kolde, R. Package ‘Pheatmap’; R Package: Vienna, Austria, 2019. [Google Scholar]
- Vimalraj, S. Alkaline phosphatase: Structure, expression and its function in bone mineralization. Gene 2020, 754, 144855. [Google Scholar] [CrossRef]
- Nakamura, M.; Aoyama, N.; Yamaguchi, S.; Sasano, Y. Expression of tartrate-resistant acid phosphatase and cathepsin K during osteoclast differentiation in developing mouse mandibles. Biomed. Res. 2021, 42, 13–21. [Google Scholar] [CrossRef]
- Gao, L.; Li, Y.; Yang, Y.J.; Zhang, D.Y. The effect of moderate-intensity treadmill exercise on bone mass and the transcription of peripheral blood mononuclear cells in ovariectomized rats. Front. Physiol. 2021, 12, 729910. [Google Scholar] [CrossRef]
- Dror, A.D.; Virk, K.; Lee, K.; Gerston, A.; Prakash, A.; Abbott, M.J.; Jaque, S.V.; Sumida, K.D. Resistance training threshold for elevating bone mineral density in growing female rats. Int. J. Sports Med. 2018, 39, 382–389. [Google Scholar] [CrossRef]
- Shojaa, M.; von Stengel, S.; Kohl, M.; Schoene, D.; Kemmler, W. Effects of dynamic resistance exercise on bone mineral density in postmenopausal women: A systematic review and meta-analysis with special emphasis on exercise parameters. Osteoporos. Int. 2020, 31, 1427–1444. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, K.; Suzuki, N.; Sato, Y.; Nemoto, T.; Ikegame, M.; Shimizu, N.; Kondo, T.; Furusawa, Y.; Wada, S.; Hattori, A. Osteoblast activity in the goldfish scale responds sensitively to mechanical stress. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2010, 156, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Totland, G.K.; Fjelldal, P.G.; Kryvi, H.; Løkka, G.; Wargelius, A.; Sagstad, A.; Hansen, T.; Grotmol, S. Sustained swimming increases the mineral content and osteocyte density of salmon vertebral bone. J. Anat. 2011, 219, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Sakakura, Y.; Shide, N.; Tsuruga, E.; Irie, K.; Yajima, T. Effects of running exercise on the mandible and tibia of ovariectomized rats. J. Bone Miner. Metab. 2001, 19, 159–167. [Google Scholar] [CrossRef]
- Hagihara, Y.; Nakajima, A.; Fukuda, S.; Goto, S.; Iida, H.; Yamazaki, M. Running exercise for short duration increases bone mineral density of loaded long bones in young growing rats. Tohoku J. Exp. Med. 2009, 219, 139–143. [Google Scholar] [CrossRef]
- Yuan, Y.; Chen, X.; Zhang, L.; Wu, J.; Guo, J.; Zou, D.; Chen, B.; Sun, Z.; Shen, C.; Zou, J. The roles of exercise in bone remodeling and in prevention and treatment of osteoporosis. Prog. Biophys. Mol. Biol. 2016, 122, 122–130. [Google Scholar] [CrossRef]
- Tong, X.; Chen, X.; Zhang, S.; Huang, M.; Shen, X.; Xu, J.; Zou, J. The effect of exercise on the prevention of osteoporosis and bone angiogenesis. Biomed. Res. Int. 2019, 2019, 8171897. [Google Scholar] [CrossRef]
- Panahi, N.; Arjmand, B.; Ostovar, A.; Kouhestani, E.; Heshmat, R.; Soltani, A.; Larijani, B. Metabolomic biomarkers of low BMD: A systematic review. Osteoporos. Int. 2021, 32, 2407–2431. [Google Scholar] [CrossRef]
- You, Y.S.; Lin, C.Y.; Liang, H.J.; Lee, S.H.; Tsai, K.S.; Chiou, J.M.; Chen, Y.C.; Tsao, C.K.; Chen, J.H. Association between the metabolome and low bone mineral density in Taiwanese women determined by (1)H NMR spectroscopy. J. Bone Miner. Res. 2014, 29, 212–222. [Google Scholar] [CrossRef]
- Mason, D.J. The role of glutamate transporters in bone cell signalling. Musculoskelet. Neuronal. Interact. 2004, 4, 128–131. [Google Scholar]
- Lorenzo, M.P.; Valiente, L.; Buendia, I.; Gortázar, A.R.; Garcia, A. Optimization and validation of a chiral CE-LIF method for quantitation of aspartate, glutamate and serine in murine osteocytic and osteoblastic cells. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2020, 1152, 122259. [Google Scholar] [CrossRef]
- Hohnholt, M.C.; Andersen, V.H.; Andersen, J.V.; Christensen, S.K.; Karaca, M.; Maechler, P.; Waagepetersen, H.S. Glutamate dehydrogenase is essential to sustain neuronal oxidative energy metabolism during stimulation. J. Cereb. Blood Flow. Metab. 2018, 38, 1754–1768. [Google Scholar] [CrossRef]
- Gong, R.; Xiao, H.M.; Zhang, Y.H.; Zhao, Q.; Su, K.J.; Lin, X.; Mo, C.L.; Zhang, Q.; Du, Y.T.; Lyu, F.Y.; et al. Identification and functional characterization of metabolites for bone mass in peri- and postmenopausal Chinese women. J. Clin. Endocrinol. Metab. 2021, 106, e3159–e3177. [Google Scholar] [CrossRef]
- Yu, L.; Qi, H.; An, G.; Bao, J.; Ma, B.; Zhu, J.; Ouyang, G.; Zhang, P.; Fan, H.; Zhang, Q. Association between metabolic profiles in urine and bone mineral density of pre- and postmenopausal Chinese women. Menopause 2019, 26, 94–102. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, S.; Mei, F.; Zhao, M.; Xia, G.; Shen, X. Tilapia nilotica head lipids improved bone loss by regulating inflammation and serum metabolism through gut microbiota in ovariectomized rats. Front. Nutr. 2021, 8, 792793. [Google Scholar] [CrossRef]
- Huang, C.C.; Lin, W.T.; Hsu, F.L.; Tsai, P.W.; Hou, C.C. Metabolomics investigation of exercise-modulated changes in metabolism in rat liver after exhaustive and endurance exercises. Eur. J. Appl. Physiol. 2010, 108, 557–566. [Google Scholar] [CrossRef]
- Wang, W.; Wang, Y.; Hu, J.; Duan, H.; Wang, Z.; Yin, L.; He, F. Untargeted metabolomics reveal the protective effect of bone marrow mesenchymal stem cell transplantation against ovariectomy-induced osteoporosis in mice. Cell Transplant. 2022, 31, 9636897221079745. [Google Scholar] [CrossRef]
- Misra, B.B.; Jayapalan, S.; Richards, A.K.; Helderman, R.C.M.; Rendina-Ruedy, E. Untargeted metabolomics in primary murine bone marrow stromal cells reveals distinct profile throughout osteoblast differentiation. Metabolomics 2021, 17, 86. [Google Scholar] [CrossRef]
- Rokitzki, L.; Sagredos, A.; Reuss, F.; Petersen, G.; Keul, J. Pantothenic acid levels in blood of athletes at rest and after aerobic exercise. Z. Ernahr. 1993, 32, 282–288. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, H.; Li, G.H.; Long, M.T.; Cheung, C.L.; Vasan, R.S.; Hsu, Y.H.; Kiel, D.P.; Liu, C.T. Metabolomics insights into osteoporosis through association with bone mineral density. J. Bone Miner. Res. 2021, 36, 729–738. [Google Scholar] [CrossRef]
- Saoi, M.; Britz-McKibbin, P. New advances in tissue metabolomics: A review. Metabolites 2021, 11, 672. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway | Name | KEGG ID | Mode | Expression Level | VIP | FC | p. Value | Regulation | |
|---|---|---|---|---|---|---|---|---|---|
| Control Group | Exercise Group | ||||||||
| Alanine, aspartate, and glutamate metabolism | D-aspartate | C00402 | ESI+ | 5.34 × 107 ± 2.52 × 107 | 3.04 × 107 ± 1.67 × 107 | 1.46 | 0.57 | 0.045 | down |
| Pantothenate and CoA biosynthesis | Pantothenate | C00864 | ESI+ | 1.27 × 108 ± 2.27 × 107 | 8.96 × 107 ± 2.33 × 107 | 1.13 | 0.71 | 0.022 | down |
| Pantothenate and CoA biosynthesis; pyrimidine metabolism; β-alanine metabolism | Uracil | C00106 | ESI− | 4.44 × 107 ± 9.00 × 106 | 3.32 × 107 ± 6.69 × 106 | 1.00 | 0.75 | 0.048 | down |
| Pyrimidine metabolism | Uridine | C00299 | ESI− | 2.59 × 107 ± 4.91 × 106 | 1.82 × 107 ± 4.17 × 106 | 1.13 | 0.70 | 0.015 | down |
| Name | Gene ID | Average FPKM | Log2FC | q. Value | Regulation | |
|---|---|---|---|---|---|---|
| Control Group | Exercise Group | |||||
| Alanine, aspartate, and glutamate metabolism | ||||||
| si:dkey-40g16.6 | 337166 | 2.34 ± 1.66 | 1.13 ± 1.56 | −1.07 | 1.51 × 10−15 | down |
| glud1b | 373092 | 159.80 ± 56.49 | 112.42 ± 29.39 | −0.54 | <1.00 × 10−300 | down |
| asl | 393423 | 12.97 ± 3.68 | 7.90 ± 3.175 | −0.76 | 8.17 × 10−37 | down |
| aldh4a1 | 394133 | 23.90 ± 17.11 | 15.72 ± 12.01 | −0.64 | 5.39 × 10−57 | down |
| asrgl1 | 541402 | 9.65 ± 1.95 | 6.57 ± 3.10 | −0.58 | 2.19 × 10−15 | down |
| cps1 | 555623 | 0.53 ± 0.38 | 0.32 ± 0.19 | −0.66 | 4.65 × 10− 5 | down |
| aspa | 557232 | 4.67 ± 6.14 | 2.63 ± 3.30 | −0.87 | 2.21 × 10−12 | down |
| aldh5a1 | 565235 | 6.80 ± 3.46 | 4.68 ± 3.26 | −0.56 | 8.02 × 10−17 | down |
| gfpt2 | 569945 | 1.84 ± 1.66 | 1.15 ± 1.28 | −0.69 | 9.76 × 10−7 | down |
| si:dkey-103j14.5 | 570552 | 0.46 ± 0.62 | 0.26 ± 0.26 | −0.90 | 5.62 × 10−6 | down |
| agxt2 | 619269 | 36.68 ± 22.26 | 25.92 ± 29.65 | −0.51 | 2.48 × 10−56 | down |
| si:ch211-56m1.8 | 795494 | 1.25 ± 0.72 | 0.86 ± 0.52 | −0.58 | 1.68 × 10−8 | down |
| Pantothenate and CoA biosynthesis | ||||||
| LOC100536156 | 100536156 | 0.61 ± 0.77 | 1.53 ± 2.23 | 1.26 | 6.81 × 10−8 | up |
| ppcdc | 393753 | 7.05 ± 1.83 | 4.76 ± 1.10 | −0.61 | 4.21 × 10−13 | down |
| Pyrimidine metabolism | ||||||
| LOC100536156 | 100536156 | 0.61 ± 0.77 | 1.53 ± 2.23 | 1.26 | 6.81 × 10−8 | up |
| nme2b.2 | 30084 | 17,455.26 ± 18 | 32,248.48 ± 21 | 0.98 | <1.00 × 10−300 | up |
| uckl1b | 558466 | 11.70 ± 3.13 | 21.01 ± 14.84 | 0.80 | 4.08 × 10−100 | up |
| entpd3 | 100005551 | 40.96 ± 29.61 | 13.65 ± 9.90 | −1.63 | <1.00 × 10−300 | down |
| rrm2 | 30733 | 9.68 ± 10.39 | 4.45 ± 4.27 | −1.13 | 5.09 × 10−65 | down |
| ctps1a | 322089 | 10.92 ± 6.16 | 7.30 ± 2.21 | −0.61 | 1.00 × 10−27 | down |
| tk1 | 327590 | 0.95 ± 0.81 | 0.48 ± 0.36 | −1.00 | 3.54 × 10−8 | down |
| nme4 | 394170 | 1.17 ± 0.88 | 0.67 ± 0.31 | −0.82 | 1.93 × 10−3 | down |
| cant1a | 406558 | 12.62 ± 6.37 | 8.66 ± 3.85 | −0.57 | 7.55 × 10−24 | down |
| tk2 | 437016 | 6.14 ± 1.63 | 4.35 ± 1.54 | −0.51 | 4.89 × 10−8 | down |
| entpd1 | 445151 | 19.69 ± 7.60 | 13.17 ± 5.73 | −0.61 | 3.22 × 10−54 | down |
| cant1b | 445201 | 16.76 ± 7.31 | 8.55 ± 4.27 | −1.00 | 5.64 × 10−87 | down |
| uck1 | 447928 | 4.76 ± 2.91 | 3.15 ± 2.51 | −0.62 | 4.26 × 10−6 | down |
| dck | 474325 | 3.45 ± 1.97 | 1.85 ± 0.98 | −0.89 | 1.20 × 10−7 | down |
| zgc:110540 | 541538 | 2.72 ± 2.44 | 1.74 ± 0.71 | −0.67 | 2.08 × 10−6 | down |
| cmpk2 | 570478 | 2.00 ± 1.34 | 0.86 ± 0.26 | −1.26 | 3.90 × 10−16 | down |
| nt5c1ab | 799449 | 1.46 ± 1.04 | 0.86 ± 0.84 | −0.89 | 6.11 × 10−8 | down |
| β-Alanine metabolism | ||||||
| aldh3b1 | 282559 | 3.51 ± 3.03 | 2.39 ± 1.62 | −0.57 | 9.81 × 10−8 | down |
| cndp2 | 327288 | 21.54 ± 12.27 | 13.88 ± 6.10 | −0.64 | 3.11 × 10−56 | down |
| aldh16a1 | 492710 | 14.81 ± 5.35 | 9.87 ± 4.32 | −0.62 | 2.47 × 10−50 | down |
| aoc2 | 541429 | 84.09 ± 53.64 | 36.07 ± 14.10 | −1.28 | <1.00 × 10−300 | down |
| aldh3b2 | 557008 | 1.76 ± 0.37 | 1.18 ± 0.64 | −0.61 | 1.48 × 10−8 | down |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, J.-L.; Yang, W.-Y.; Zhang, Q.; Feng, H.; Wang, X.-B.; Li, H.; Zhou, S.; Xiao, S.-M. Integration of Metabolomics and Transcriptomics to Reveal the Metabolic Characteristics of Exercise-Improved Bone Mass. Nutrients 2023, 15, 1694. https://doi.org/10.3390/nu15071694
Hou J-L, Yang W-Y, Zhang Q, Feng H, Wang X-B, Li H, Zhou S, Xiao S-M. Integration of Metabolomics and Transcriptomics to Reveal the Metabolic Characteristics of Exercise-Improved Bone Mass. Nutrients. 2023; 15(7):1694. https://doi.org/10.3390/nu15071694
Chicago/Turabian StyleHou, Jin-Li, Wan-Yu Yang, Qiong Zhang, Hao Feng, Xiao-Bao Wang, Hui Li, Sheng Zhou, and Su-Mei Xiao. 2023. "Integration of Metabolomics and Transcriptomics to Reveal the Metabolic Characteristics of Exercise-Improved Bone Mass" Nutrients 15, no. 7: 1694. https://doi.org/10.3390/nu15071694