
Factors and Mechanisms Influencing Conjugation In Vivo in the Gastrointestinal Tract Environment: A Review

Abstract
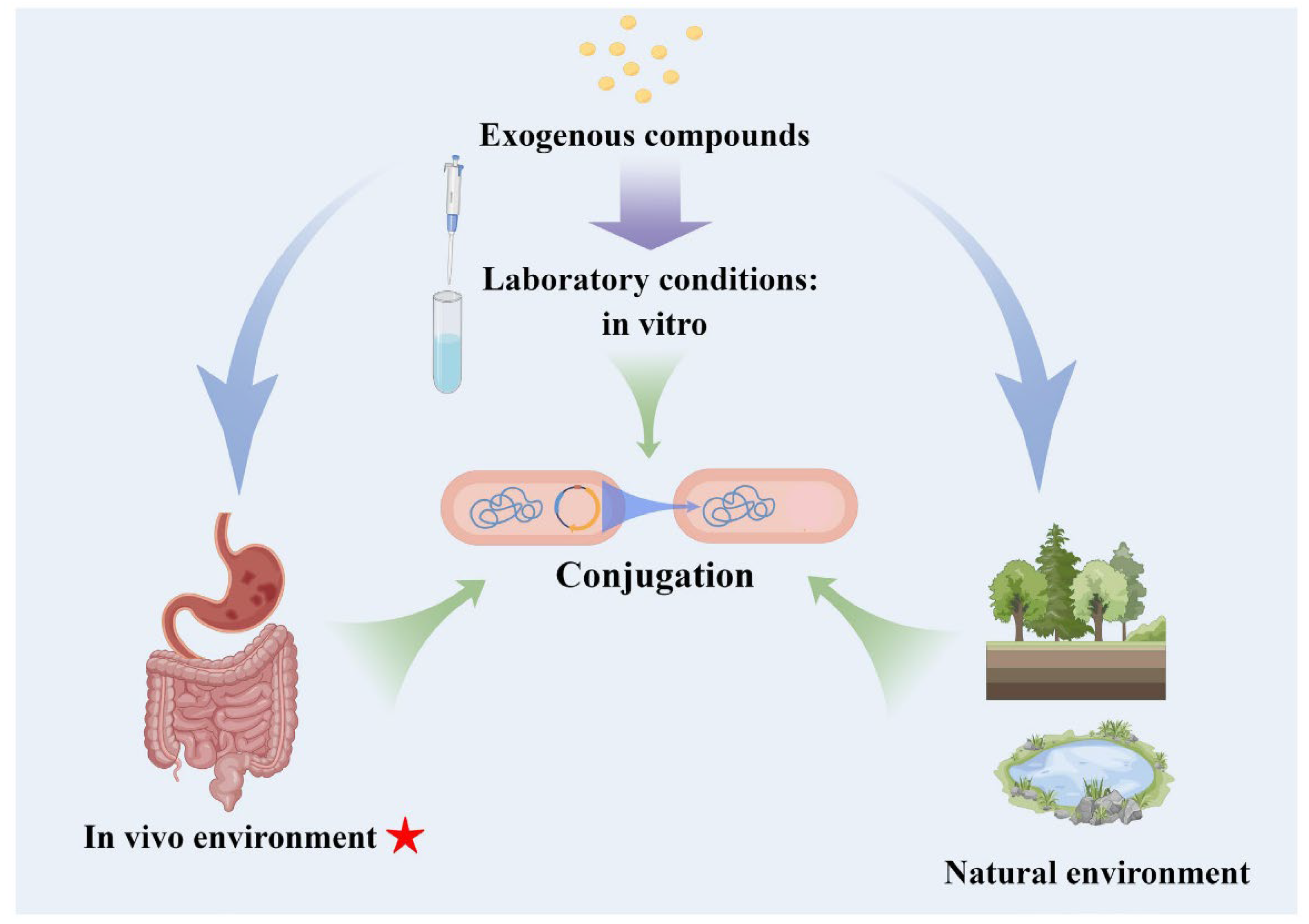
:1. Introduction
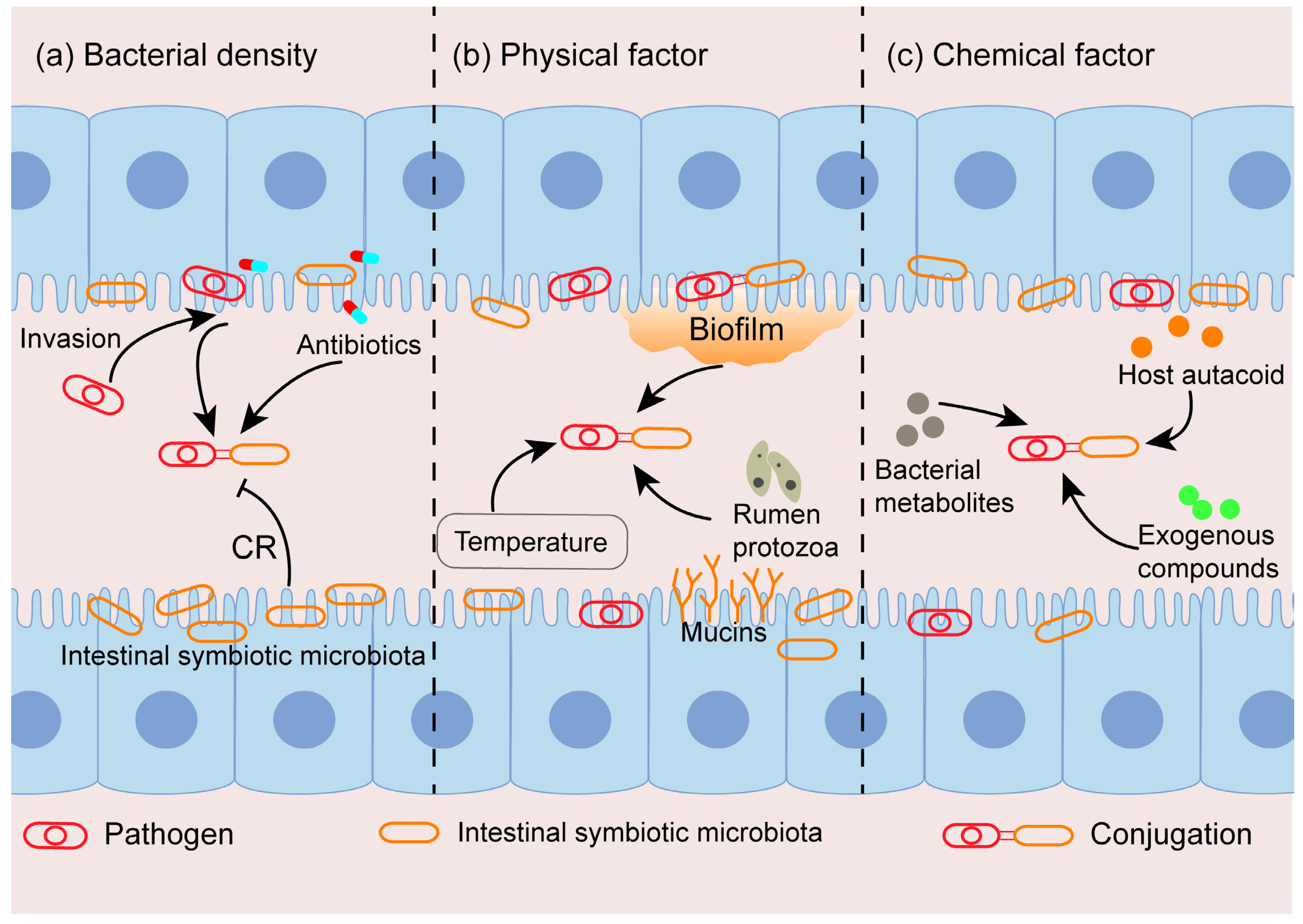
2. Factors Influencing Conjugation In Vivo
2.1. The Density of Donor and Recipient Bacteria
2.2. Physical Factor
2.2.1. Temperature
2.2.2. Biofilm Formation
2.2.3. Others
2.3. Chemical Factor
2.3.1. Host Autacoid
2.3.2. Bacterial Metabolites
2.3.3. Exogenous Compounds

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | In Vivo Model | Strain | Plasmid | Effects | Reference |
|---|---|---|---|---|---|
| Factors affecting the density of donor and recipient bacteria: | |||||
| Ampicillin | Murine gut | E. coli O80:H26 to S. Heidelberg | IncI2 | Promote | [39] |
| Enrofloxacin | Chicken gut | E. coli E2 to Salmonella SE211 | IncY | Promote | [34] |
| Penicillin | Rat gut | K. pneumoniae HN-1 to gastrointestinal tract E. coli | blaNDM-1-bearing plasmid | Promote | [38] |
| Physical factor: | |||||
| Temperature | Chicken meat | mcr-1-positive donors to mcr-1-negative recipients (designated as R7 and E. coli C600) | mcr-1-positive plasmid | 37 °C > 25 °C | [43] |
| Biofilm | Simulation in vitro | / | / | Promotion | [22] |
| Rumen protozoa | Rumen protozoa | K. pneumoniae TCR2003 to S. Typhimurium SL1344 | pGFP | Promotion | [50] |
| Mucin | Simulation in vitro | K. pneumoniae A2312NM to K. pneumoniae MP13 | pHNSHP45 | Inhibition | [33] |
| Chemical factor: | |||||
| pH | Simulation in vitro | E. hormaechei to E. coli BL21(DE3) | pET28a-EGFP | / | [52] |
| Bile salts | Simulation in vitro | K. pneumoniae A2312NM to K. pneumoniae MP13 | pHNSHP45 | Promotion | [33] |
| Norepinephrine (NE) | Gut (simulation in vitro) | S. Typhimurium 5678 to E. coli C600N | Promotion | [54] | |
| cCF10 | Murine gut | / | pCF10 | Promotion | [59] |
| Metabolites of lactic acid bacteria | Chicken gut (simulation in vitro) | E. coli MG1655 to E. coli MG1655 | IncI1 | Inhibition | [60] |
| Indole | Simulation in vitro | E. coli SM10λπ-PAO1 and SM10λπ-EC600 | pUCP24T | Inhibition | [15] |
| 2-hexadecynoic acid (2-HDA) | Murine gut | E. coli MDS42 to E. coli MDS52 | pOX-38 | Inhibition | [62] |
| Melatonin | Mouse infection model | E. coli DH5α to E. coli EC600 | RP4–7 | Inhibition | [58] |
| Artificial sweeteners | Murine gut (simulation in vitro) | E. coli K-12 MG1655 to mice feces | pKJK5 plasmid | Promotion | [32] |
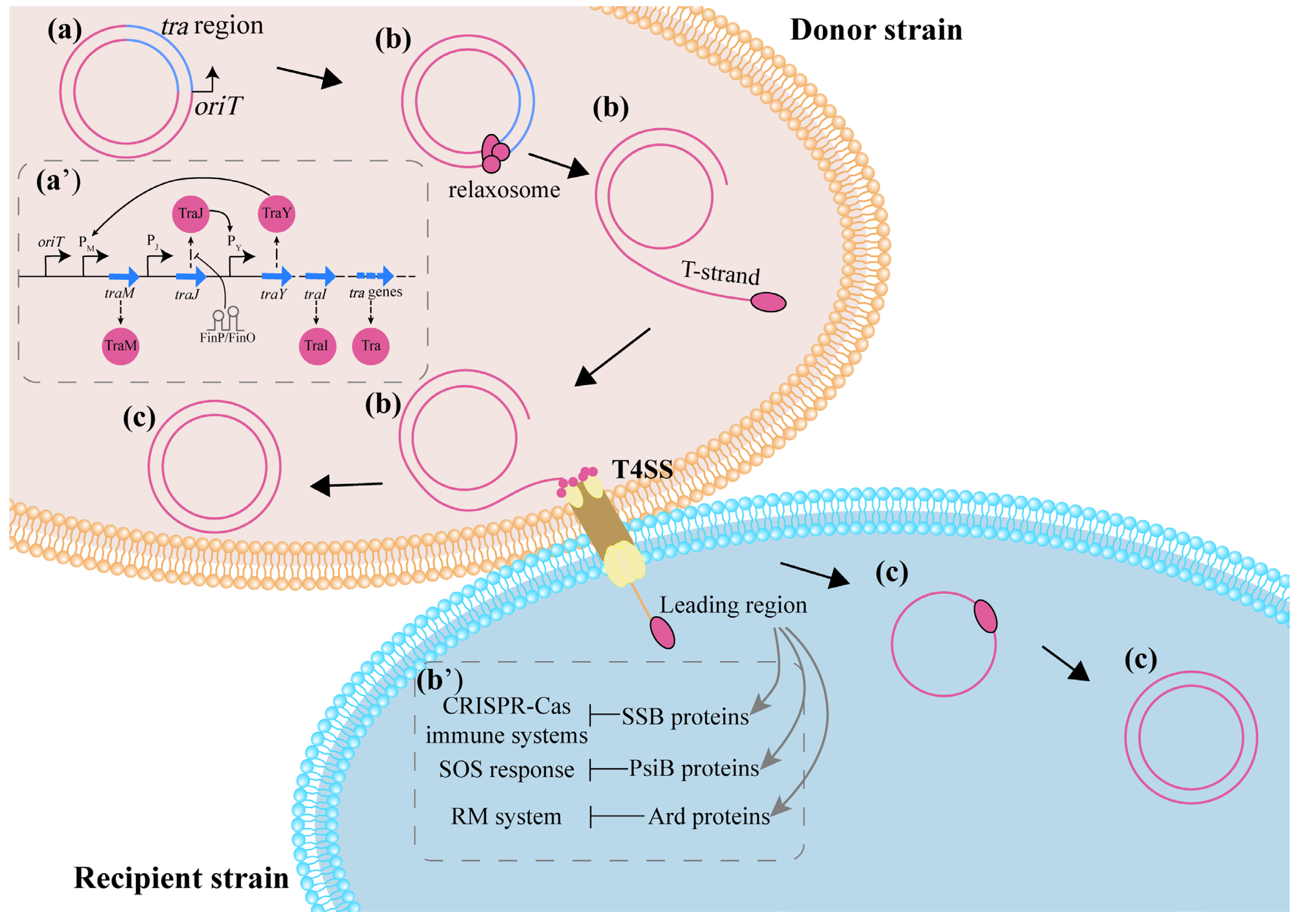
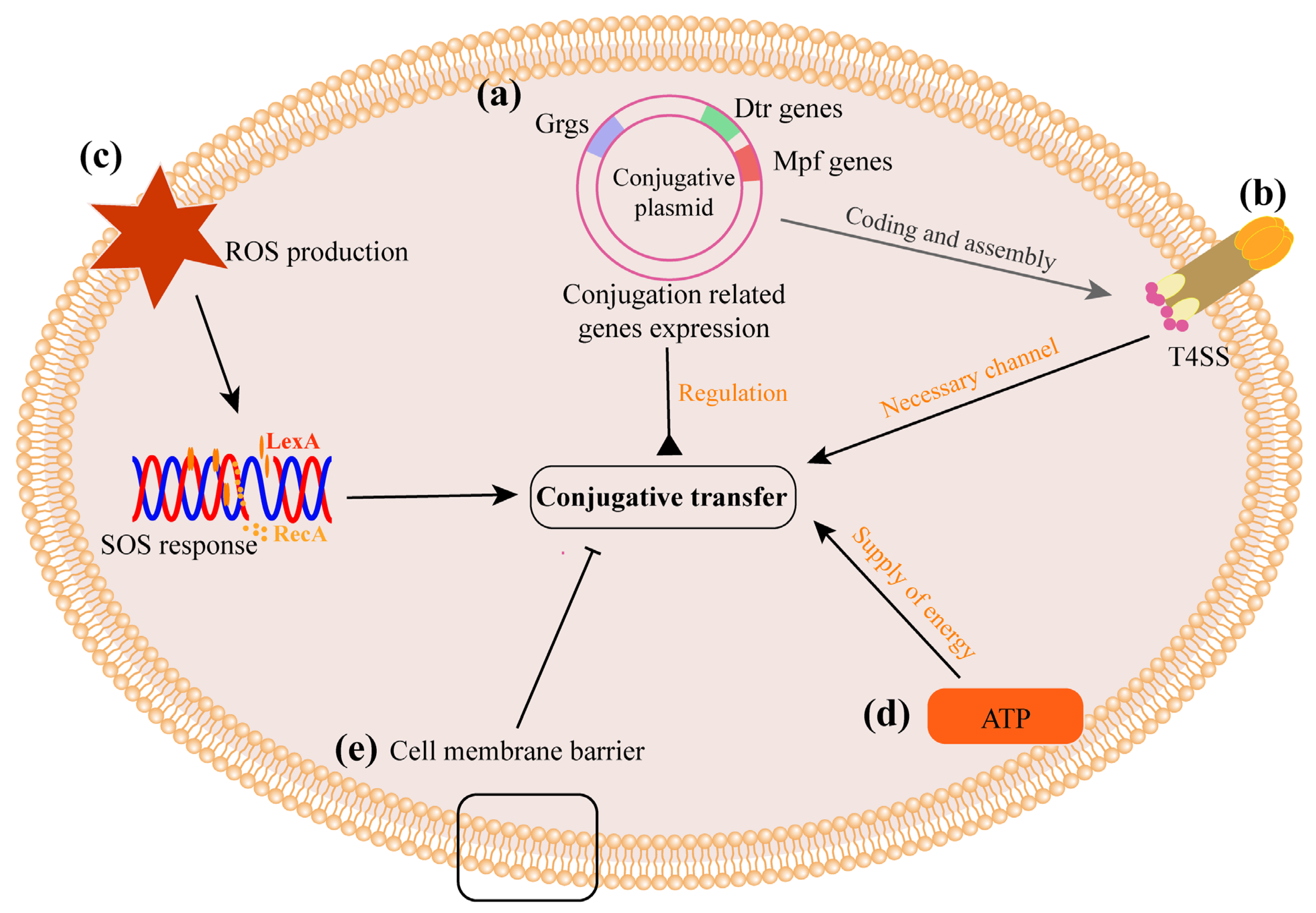
3. Potential Mechanisms Influencing CF In Vivo
3.1. Bacterial Colonization
3.2. The Process of Conjugation
3.2.1. Interference in the Expression of Conjugation-Related Genes Directly Affects CF
3.2.2. ROS Production and SOS Responses Increase CF
3.2.3. Energy Supply Is Necessary for Conjugation
3.2.4. Cell Membrane Is a Barrier to Plasmid Transfer
4. Conclusions and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumarasamy, K.K.; Toleman, M.A.; Walsh, T.R.; Bagaria, J.; Butt, F.; Balakrishnan, R.; Chaudhary, U.; Doumith, M.; Giske, C.G.; Irfan, S.; et al. Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: A molecular, biological, and epidemiological study. Lancet Infect. Dis. 2010, 10, 597–602. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Sun, J.; Chen, C.; Cui, C.Y.; Zhang, Y.; Liu, X.; Cui, Z.H.; Ma, X.Y.; Feng, Y.; Fang, L.X.; Lian, X.L.; et al. Plasmid-encoded tet(X) genes that confer high-level tigecycline resistance in Escherichia coli. Nat. Microbiol. 2019, 4, 1457–1464. [Google Scholar] [CrossRef]
- He, T.; Wang, R.; Liu, D.; Walsh, T.R.; Zhang, R.; Lv, Y.; Ke, Y.; Ji, Q.; Wei, R.; Liu, Z.; et al. Emergence of plasmid-mediated high-level tigecycline resistance genes in animals and humans. Nat. Microbiol. 2019, 4, 1450–1456. [Google Scholar] [CrossRef]
- Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Stevenson, C.; Hall, J.P.; Harrison, E.; Wood, A.; Brockhurst, M.A. Gene mobility promotes the spread of resistance in bacterial populations. ISME J. 2017, 11, 1930–1932. [Google Scholar] [CrossRef] [Green Version]
- Holmes, A.H.; Moore, L.S.P.; Sundsfjord, A.; Steinbakk, M.; Regmi, S.; Karkey, A.; Guerin, P.J.; Piddock, L.J.V. Understanding the mechanisms and drivers of antimicrobial resistance. Lancet 2016, 387, 176–187. [Google Scholar] [CrossRef]
- Soler, N.; Forterre, P. Vesiduction: The fourth way of HGT. Environ. Microbiol. 2020, 22, 2457–2460. [Google Scholar] [CrossRef]
- Frost, L.S.; Leplae, R.; Summers, A.O.; Toussaint, A. Mobile genetic elements: The agents of open source evolution. Nat. Rev. Microbiol. 2005, 3, 722–732. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, J.; Mao, L.; Li, J.; Yuan, Z.; Bond, P.L.; Guo, J. Antiepileptic drug carbamazepine promotes horizontal transfer of plasmid-borne multi-antibiotic resistance genes within and across bacterial genera. ISME J. 2019, 13, 509–522. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Z.; Yu, Y.; Chen, Z.; Jin, M.; Yang, D.; Zhao, Z.; Wang, J.; Shen, Z.; Wang, X.; Qian, D.; et al. Nanoalumina promotes the horizontal transfer of multiresistance genes mediated by plasmids across genera. Proc. Natl. Acad. Sci. USA 2012, 109, 4944–4949. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Lopez, R.; Machon, C.; Longshaw, C.M.; Martin, S.; Molin, S.; Zechner, E.L.; Espinosa, M.; Lanka, E.; de la Cruz, F. Unsaturated fatty acids are inhibitors of bacterial conjugation. Microbiology (Reading) 2005, 151, 3517–3526. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Wang, Y.; Song, H.; Lu, J.; Yuan, Z.; Guo, J. Copper nanoparticles and copper ions promote horizontal transfer of plasmid-mediated multi-antibiotic resistance genes across bacterial genera. Environ. Int. 2019, 129, 478–487. [Google Scholar] [CrossRef]
- Pu, Q.; Fan, X.T.; Sun, A.Q.; Pan, T.; Li, H.; Bo Lassen, S.; An, X.L.; Su, J.Q. Co-effect of cadmium and iron oxide nanoparticles on plasmid-mediated conjugative transfer of antibiotic resistance genes. Environ. Int. 2021, 152, 106453. [Google Scholar] [CrossRef]
- Xiong, R.; Liu, Y.; Pu, J.; Liu, J.; Zheng, D.; Zeng, J.; Chen, C.; Lu, Y.; Huang, B. Indole Inhibits IncP-1 Conjugation System Mainly Through Promoting korA and korB Expression. Front. Microbiol. 2021, 12, 628133. [Google Scholar] [CrossRef]
- Shun-Mei, E.; Zeng, J.M.; Yuan, H.; Lu, Y.; Cai, R.X.; Chen, C. Sub-inhibitory concentrations of fluoroquinolones increase conjugation frequency. Microb. Pathog. 2018, 114, 57–62. [Google Scholar] [CrossRef]
- Ding, P.; Lu, J.; Wang, Y.; Schembri, M.A.; Guo, J. Antidepressants promote the spread of antibiotic resistance via horizontally conjugative gene transfer. Environ. Microbiol. 2022, 24, 5261–5276. [Google Scholar] [CrossRef]
- Song, Z.; Zuo, L.; Li, C.; Tian, Y.; Wang, H. Copper Ions Facilitate the Conjugative Transfer of SXT/R391 Integrative and Conjugative Element Across Bacterial Genera. Front. Microbiol 2020, 11, 616792. [Google Scholar] [CrossRef]
- He, K.; Xue, B.; Yang, X.; Wang, S.; Li, C.; Zhang, X.; Zhao, C.; Wang, X.; Qiu, Z.; Shen, Z.; et al. Low-concentration of trichloromethane and dichloroacetonitrile promote the plasmid-mediated horizontal transfer of antibiotic resistance genes. J. Hazard Mater. 2022, 425, 128030. [Google Scholar] [CrossRef]
- Cen, T.; Zhang, X.; Xie, S.; Li, D. Preservatives accelerate the horizontal transfer of plasmid-mediated antimicrobial resistance genes via differential mechanisms. Environ. Int. 2020, 138, 105544. [Google Scholar] [CrossRef]
- Li, X.; Wen, C.; Liu, C.; Lu, S.; Xu, Z.; Yang, Q.; Chen, Z.; Liao, H.; Zhou, S. Herbicide promotes the conjugative transfer of multi-resistance genes by facilitating cellular contact and plasmid transfer. J. Environ. Sci. 2022, 115, 363–373. [Google Scholar] [CrossRef]
- Hausner, M.; Wuertz, S. High rates of conjugation in bacterial biofilms as determined by quantitative in situ analysis. Appl. Environ. Microbiol. 1999, 65, 3710–3713. [Google Scholar] [CrossRef] [Green Version]
- Madsen, J.S.; Burmolle, M.; Hansen, L.H.; Sorensen, S.J. The interconnection between biofilm formation and horizontal gene transfer. FEMS Immunol. Med. Microbiol. 2012, 65, 183–195. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Tong, Z.; Shi, J.; Jia, Y.; Yang, K.; Wang, Z. Correlation between Exogenous Compounds and the Horizontal Transfer of Plasmid-Borne Antibiotic Resistance Genes. Microorganisms 2020, 8, 1211. [Google Scholar] [CrossRef]
- Li, W.; Zhang, G. Detection and various environmental factors of antibiotic resistance gene horizontal transfer. Environ. Res. 2022, 212, 113267. [Google Scholar] [CrossRef]
- Feng, G.; Huang, H.; Chen, Y. Effects of emerging pollutants on the occurrence and transfer of antibiotic resistance genes: A review. J. Hazard Mater. 2021, 420, 126602. [Google Scholar] [CrossRef]
- Berg, R.D. The indigenous gastrointestinal microflora. Trends Microbiol. 1996, 4, 430–435. [Google Scholar] [CrossRef]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef]
- Stecher, B.; Denzler, R.; Maier, L.; Bernet, F.; Sanders, M.J.; Pickard, D.J.; Barthel, M.; Westendorf, A.M.; Krogfelt, K.A.; Walker, A.W.; et al. Gut inflammation can boost horizontal gene transfer between pathogenic and commensal Enterobacteriaceae. Proc. Natl. Acad. Sci. USA 2012, 109, 1269–1274. [Google Scholar] [CrossRef] [Green Version]
- Sassone-Corsi, M.; Raffatellu, M. No vacancy: How beneficial microbes cooperate with immunity to provide colonization resistance to pathogens. J. Immunol. 2015, 194, 4081–4087. [Google Scholar] [CrossRef] [Green Version]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Henderson, I.R.; Guo, J. Non-caloric artificial sweeteners modulate conjugative transfer of multi-drug resistance plasmid in the gut microbiota. Gut. Microbes 2023, 15, 2157698. [Google Scholar] [CrossRef]
- Yang, X.; Shu, R.; Hou, L.; Ren, P.; Lu, X.; Huang, Z.; Zhong, Z.; Wang, H. mcr-1-Mediated In Vitro Inhibition of Plasmid Transfer Is Reversed by the Intestinal Environment. Antibiotics 2022, 11, 875. [Google Scholar] [CrossRef]
- Zhao, Y.; Cao, Z.; Cui, L.; Hu, T.; Guo, K.; Zhang, F.; Wang, X.; Peng, Z.; Liu, Q.; Dai, M. Enrofloxacin Promotes Plasmid-Mediated Conjugation Transfer of Fluoroquinolone-Resistance Gene qnrS. Front. Microbiol. 2021, 12, 773664. [Google Scholar] [CrossRef]
- Ott, L.C.; Mellata, M. Models for Gut-Mediated Horizontal Gene Transfer by Bacterial Plasmid Conjugation. Front. Microbiol. 2022, 13, 891548. [Google Scholar] [CrossRef]
- Kessler, C.; Hou, J.; Neo, O.; Buckner, M.M.C. In situ, in vivo and in vitro approaches for studying AMR plasmid conjugation in the gut microbiome. FEMS Microbiol. Rev. 2022, 47, fuac044. [Google Scholar] [CrossRef]
- Crits-Christoph, A.; Hallowell, H.A.; Koutouvalis, K.; Suez, J. Good microbes, bad genes? The dissemination of antimicrobial resistance in the human microbiome. Gut Microbes 2022, 14, 2055944. [Google Scholar] [CrossRef]
- Ye, L.; Chan, E.W.C.; Chen, S. Selective and suppressive effects of antibiotics on donor and recipient bacterial strains in gut microbiota determine transmission efficiency of blaNDM-1-bearing plasmids. J. Antimicrob. Chemother. 2019, 74, 1867–1875. [Google Scholar] [CrossRef] [Green Version]
- Laskey, A.; Devenish, J.; Kang, M.; Savic, M.; Chmara, J.; Dan, H.; Lin, M.; Robertson, J.; Bessonov, K.; Gurnik, S.; et al. Mobility of β-lactam resistance under ampicillin treatment in gut microbiota suffering from pre-disturbance. Microb. Genom. 2021, 7, 713. [Google Scholar] [CrossRef]
- Zhang, P.; Mao, D.; Gao, H.; Zheng, L.; Chen, Z.; Gao, Y.; Duan, Y.; Guo, J.; Luo, Y.; Ren, H. Colonization of gut microbiota by plasmid-carrying bacteria is facilitated by evolutionary adaptation to antibiotic treatment. ISME J. 2022, 16, 1284–1293. [Google Scholar] [CrossRef]
- Xiao, X.; Zeng, F.; Li, R.; Liu, Y.; Wang, Z. Subinhibitory Concentration of Colistin Promotes the Conjugation Frequencies of Mcr-1- and bla(NDM-5)-Positive Plasmids. Microbiol. Spectr. 2022, 10, e0216021. [Google Scholar] [CrossRef]
- Stecher, B.; Maier, L.; Hardt, W.D. ‘Blooming’ in the gut: How dysbiosis might contribute to pathogen evolution. Nat. Rev. Microbiol. 2013, 11, 277–284. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Bai, X.; Sheng, H.; Chen, J.; Wang, Z.; Wang, T.; Sun, R.; Feng, Z.; Wang, Y.; Peng, K.; et al. Conjugative transfer of mcr-1-bearing plasmid from Salmonella to Escherichia coli in vitro on chicken meat and in mouse gut. Food Res. Int. 2022, 157, 111263. [Google Scholar] [CrossRef]
- Yang, B.; Wang, Z.; Jia, Y.; Fang, D.; Li, R.; Liu, Y. Paclitaxel and its derivative facilitate the transmission of plasmid-mediated antibiotic resistance genes through conjugative transfer. Sci. Total Environ. 2022, 10, 152245. [Google Scholar] [CrossRef]
- Wang, H.; Qi, H.; Zhu, M.; Gong, S.; Huang, Z.; Zhang, Y.; Chen, X.; Jiao, X. MoS2 decorated nanocomposite: Fe2O3@MoS2 inhibits the conjugative transfer of antibiotic resistance genes. Ecotoxicol. Environ. Saf. 2019, 10, 109781. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef]
- Høiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef] [Green Version]
- Molin, S.; Tolker-Nielsen, T. Gene transfer occurs with enhanced efficiency in biofilms and induces enhanced stabilisation of the biofilm structure. Curr. Opin. Biotechnol. 2003, 14, 255–261. [Google Scholar] [CrossRef]
- Paone, P.; Cani, P.D. Mucus barrier, mucins and gut microbiota: The expected slimy partners? Gut 2020, 69, 2232–2243. [Google Scholar] [CrossRef]
- McCuddin, Z.P.; Carlson, S.A.; Rasmussen, M.A.; Franklin, S.K. Klebsiella to Salmonella gene transfer within rumen protozoa: Implications for antibiotic resistance and rumen defaunation. Vet. Microbiol. 2006, 114, 275–284. [Google Scholar] [CrossRef]
- Camilleri, M. Bile acid detergency: Permeability, inflammation, and effects of sulfation. Am. J. Physiol. Gastrointest. Liver Physiol. 2022, 322, G480–G488. [Google Scholar] [CrossRef]
- Zhou, M.; Cai, Q.; Zhang, C.; Ouyang, P.; Yu, L.; Xu, Y. Antibiotic resistance bacteria and antibiotic resistance genes survived from the extremely acidity posing a risk on intestinal bacteria in an in vitro digestion model by horizontal gene transfer. Ecotoxicol. Environ. Saf. 2022, 247, 114247. [Google Scholar] [CrossRef]
- Alverdy, J.; Holbrook, C.; Rocha, F.; Seiden, L.; Wu, R.L.; Musch, M.; Chang, E.; Ohman, D.; Suh, S. Gut-derived sepsis occurs when the right pathogen with the right virulence genes meets the right host: Evidence for in vivo virulence expression in Pseudomonas aeruginosa. Ann. Surg. 2000, 232, 480–489. [Google Scholar] [CrossRef]
- Peterson, G.; Kumar, A.; Gart, E.; Narayanan, S. Catecholamines increase conjugative gene transfer between enteric bacteria. Microb. Pathog. 2011, 51, 1–8. [Google Scholar] [CrossRef]
- Cipolla-Neto, J.; Amaral, F.G.D. Melatonin as a Hormone: New Physiological and Clinical Insights. Endocr. Rev. 2018, 39, 990–1028. [Google Scholar] [CrossRef] [Green Version]
- Morera-Fumero, A.L.; Abreu-Gonzalez, P. Role of melatonin in schizophrenia. Int. J. Mol. Sci. 2013, 14, 9037–9050. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, S.; Patel, K.K.; Dehari, D.; Agrawal, A.K.; Singh, S. Melatonin and its ubiquitous anticancer effects. Mol. Cell. Biochem. 2019, 462, 133–155. [Google Scholar] [CrossRef]
- Jia, Y.; Yang, B.; Shi, J.; Fang, D.; Wang, Z.; Liu, Y. Melatonin prevents conjugative transfer of plasmid-mediated antibiotic resistance genes by disrupting proton motive force. Pharmacol. Res. 2022, 175, 105978. [Google Scholar] [CrossRef]
- Hirt, H.; Greenwood-Quaintance, K.E.; Karau, M.J.; Till, L.M.; Kashyap, P.C.; Patel, R.; Dunny, G.M. Enterococcus faecalis Sex Pheromone cCF10 Enhances Conjugative Plasmid Transfer In Vivo. mBio 2018, 9, e00037-18. [Google Scholar] [CrossRef] [Green Version]
- Duxbury, S.J.N.; Alderliesten, J.B.; Zwart, M.P.; Stegeman, A.; Fischer, E.A.J.; de Visser, J. Chicken gut microbiome members limit the spread of an antimicrobial resistance plasmid in Escherichia coli. Proc. Biol. Sci. 2021, 288, 20212027. [Google Scholar] [CrossRef]
- Yanofsky, C.; Horn, V.; Gollnick, P. Physiological studies of tryptophan transport and tryptophanase operon induction in Escherichia coli. J. Bacteriol. 1991, 173, 6009–6017. [Google Scholar] [CrossRef] [Green Version]
- Palencia-Gándara, C.; Getino, M.; Moyano, G.; Redondo, S.; Fernández-López, R.; González-Zorn, B.; de la Cruz, F. Conjugation Inhibitors Effectively Prevent Plasmid Transmission in Natural Environments. mBio 2021, 12, e0127721. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, Z.; Ding, P.; Lu, J.; Klumper, U.; Murray, A.K.; Gaze, W.H.; Guo, J. Non-antibiotic pharmaceuticals promote conjugative plasmid transfer at a community-wide level. Microbiome 2022, 10, 124. [Google Scholar] [CrossRef]
- Jia, Y.; Wang, Z.; Fang, D.; Yang, B.; Li, R.; Liu, Y. Acetaminophen promotes horizontal transfer of plasmid-borne multiple antibiotic resistance genes. Sci. Total Environ. 2021, 782, 146916. [Google Scholar] [CrossRef]
- van der Waaij, D.; Berghuis-de Vries, J.M.; Lekkerkerk, L.-v. Colonization resistance of the digestive tract in conventional and antibiotic-treated mice. J. Hyg. 1971, 69, 405–411. [Google Scholar] [CrossRef] [Green Version]
- Gorbach, S.L.; Barza, M.; Giuliano, M.; Jacobus, N.V. Colonization resistance of the human intestinal microflora: Testing the hypothesis in normal volunteers. Eur. J. Clin. Microbiol. Infect. Dis. 1988, 7, 98–102. [Google Scholar] [CrossRef]
- Bakkeren, E.; Herter, J.A.; Huisman, J.S.; Steiger, Y.; Gül, E.; Newson, J.P.M.; Brachmann, A.O.; Piel, J.; Regoes, R.; Bonhoeffer, S.; et al. Pathogen invasion-dependent tissue reservoirs and plasmid-encoded antibiotic degradation boost plasmid spread in the gut. eLife 2021, 10, e69744. [Google Scholar] [CrossRef]
- Licht, T.R.; Christensen, B.B.; Krogfelt, K.A.; Molin, S. Plasmid transfer in the animal intestine and other dynamic bacterial populations: The role of community structure and environment. Microbiology 1999, 145, 2615–2622. [Google Scholar] [CrossRef] [Green Version]
- Mastroeni, P.; Grant, A.J. Spread of Salmonella enterica in the body during systemic infection: Unravelling host and pathogen determinants. Expert Rev. Mol. Med. 2011, 13, e12. [Google Scholar] [CrossRef]
- Monack, D.M.; Mueller, A.; Falkow, S. Persistent bacterial infections: The interface of the pathogen and the host immune system. Nat. Rev. Microbiol. 2004, 2, 747–765. [Google Scholar] [CrossRef]
- Broz, P.; Ohlson, M.B.; Monack, D.M. Innate immune response to Salmonella typhimurium, a model enteric pathogen. Gut Microbes 2012, 3, 62–70. [Google Scholar] [CrossRef] [Green Version]
- Bakkeren, E.; Huisman, J.S.; Fattinger, S.A.; Hausmann, A.; Furter, M.; Egli, A.; Slack, E.; Sellin, M.E.; Bonhoeffer, S.; Regoes, R.R.; et al. Salmonella persisters promote the spread of antibiotic resistance plasmids in the gut. Nature 2019, 573, 276–280. [Google Scholar] [CrossRef]
- Lupp, C.; Robertson, M.L.; Wickham, M.E.; Sekirov, I.; Champion, O.L.; Gaynor, E.C.; Finlay, B.B. Host-mediated inflammation disrupts the intestinal microbiota and promotes the overgrowth of Enterobacteriaceae. Cell Host. Microbe 2007, 2, 204. [Google Scholar] [CrossRef] [Green Version]
- Virolle, C.; Goldlust, K.; Djermoun, S.; Bigot, S.; Lesterlin, C. Plasmid Transfer by Conjugation in Gram-Negative Bacteria: From the Cellular to the Community Level. Genes 2020, 11, 1239. [Google Scholar] [CrossRef]
- Shen, Z.; Tang, C.M.; Liu, G.Y. Towards a better understanding of antimicrobial resistance dissemination: What can be learnt from studying model conjugative plasmids? Mil. Med. Res. 2022, 9, 3. [Google Scholar] [CrossRef]
- Delcour, A.H. Outer membrane permeability and antibiotic resistance. Biochim. Biophys. Acta 2009, 1794, 808–816. [Google Scholar] [CrossRef] [Green Version]
- Samuels, A.L.; Lanka, E.; Davies, J.E. Conjugative junctions in RP4-mediated mating of Escherichia coli. J. Bacteriol. 2000, 182, 2709–2715. [Google Scholar] [CrossRef] [Green Version]
- Zatyka, M.; Bingle, L.; Jones, A.C.; Thomas, C.M. Cooperativity between KorB and TrbA repressors of broad-host-range plasmid RK2. J. Bacteriol. 2001, 183, 1022–1031. [Google Scholar] [CrossRef] [Green Version]
- Pansegrau, W.; Balzer, D.; Kruft, V.; Lurz, R.; Lanka, E. In vitro assembly of relaxosomes at the transfer origin of plasmid RP4. Proc. Natl. Acad. Sci. USA 1990, 87, 6555–6559. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Wang, Y.; Lu, J.; Bond, P.L.; Guo, J. Nonnutritive sweeteners can promote the dissemination of antibiotic resistance through conjugative gene transfer. ISME J. 2021, 15, 2117–2130. [Google Scholar] [CrossRef]
- Schröder, G.; Lanka, E. The mating pair formation system of conjugative plasmids-A versatile secretion machinery for transfer of proteins and DNA. Plasmid 2005, 54, 1–25. [Google Scholar] [CrossRef]
- Wathugala, N.D.; Hemananda, K.M.; Yip, C.B.; Hynes, M.F. Defining the requirements for the conjugative transfer of Rhizobium leguminosarum plasmid pRleVF39b. Microbiology 2020, 166, 318–331. [Google Scholar] [CrossRef]
- Cabezon, E.; Ripoll-Rozada, J.; Pena, A.; de la Cruz, F.; Arechaga, I. Towards an integrated model of bacterial conjugation. FEMS Microbiol. Rev. 2015, 39, 81–95. [Google Scholar] [CrossRef] [Green Version]
- Ilangovan, A.; Connery, S.; Waksman, G. Structural biology of the Gram-negative bacterial conjugation systems. Trends Microbiol. 2015, 23, 301–310. [Google Scholar] [CrossRef]
- Huttener, M.; Prieto, A.; Aznar, S.; Bernabeu, M.; Glaria, E.; Valledor, A.F.; Paytubi, S.; Merino, S.; Tomas, J.; Juarez, A. Expression of a novel class of bacterial Ig-like proteins is required for IncHI plasmid conjugation. PLoS Genet. 2019, 15, e1008399. [Google Scholar] [CrossRef] [Green Version]
- Hüttener, M.; Hergueta, J.; Bernabeu, M.; Prieto, A.; Aznar, S.; Merino, S.; Tomás, J.; Juárez, A. Roles of Proteins Containing Immunoglobulin-Like Domains in the Conjugation of Bacterial Plasmids. mSphere 2022, 7, e0097821. [Google Scholar] [CrossRef]
- Yi, L.; Durand, R.; Grenier, F.; Yang, J.; Yu, K.; Burrus, V.; Liu, J.H. PixR, a Novel Activator of Conjugative Transfer of IncX4 Resistance Plasmids, Mitigates the Fitness Cost of mcr-1 Carriage in Escherichia coli. mBio 2022, 13, e0320921. [Google Scholar] [CrossRef]
- Emerit, J.; Michelson, A.M. La semaine des hopitaux: Organe fonde par l’Association d’enseignement medical des hopitaux de Paris. Free. Radic. Med. Biol. 1982, 58, 2670–2675. [Google Scholar]
- Bayir, H. Reactive oxygen species. Crit. Care Med. 2005, 33, S498–S501. [Google Scholar] [CrossRef]
- Lu, J.; Wang, Y.; Jin, M.; Yuan, Z.; Bond, P.; Guo, J. Both silver ions and silver nanoparticles facilitate the horizontal transfer of plasmid-mediated antibiotic resistance genes. Water Res. 2020, 169, 115229. [Google Scholar] [CrossRef]
- Marrakchi, M.; Liu, X.; Andreescu, S. Oxidative stress and antibiotic resistance in bacterial pathogens: State of the art, methodologies, and future trends. Adv. Exp. Med. Biol. 2014, 806, 483–498. [Google Scholar] [CrossRef]
- Chen, X.; Yin, H.; Li, G.; Wang, W.; Wong, P.K.; Zhao, H.; An, T. Antibiotic-resistance gene transfer in antibiotic-resistance bacteria under different light irradiation: Implications from oxidative stress and gene expression. Water Res. 2019, 149, 282–291. [Google Scholar] [CrossRef]
- Dwyer, D.J.; Kohanski, M.A.; Collins, J.J. Role of reactive oxygen species in antibiotic action and resistance. Curr. Opin. Microbiol. 2009, 12, 482–489. [Google Scholar] [CrossRef] [Green Version]
- Žgur-Bertok, D. DNA damage repair and bacterial pathogens. PLoS Pathog. 2013, 9, e1003711. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.K.; Alhhazmi, A.; DeCoteau, J.F.; Luo, Y.; Geyer, C.R. RecA Inhibitors Potentiate Antibiotic Activity and Block Evolution of Antibiotic Resistance. Cell Chem. Biol. 2016, 23, 381–391. [Google Scholar] [CrossRef]
- Memar, M.Y.; Yekani, M.; Celenza, G.; Poortahmasebi, V.; Naghili, B.; Bellio, P.; Baghi, H.B. The central role of the SOS DNA repair system in antibiotics resistance: A new target for a new infectious treatment strategy. Life Sci. 2020, 262, 118562. [Google Scholar] [CrossRef]
- Beaber, J.W.; Hochhut, B.; Waldor, M.K. SOS response promotes horizontal dissemination of antibiotic resistance genes. Nature 2004, 427, 72–74. [Google Scholar] [CrossRef]
- Baharoglu, Z.; Bikard, D.; Mazel, D. Conjugative DNA transfer induces the bacterial SOS response and promotes antibiotic resistance development through integron activation. PLoS Genet. 2010, 6, e1001165. [Google Scholar] [CrossRef] [Green Version]
- Chen, I.; Christie, P.J.; Dubnau, D. The ins and outs of DNA transfer in bacteria. Science 2005, 310, 1456–1460. [Google Scholar] [CrossRef] [Green Version]
- Borisov, V.B.; Murali, R.; Verkhovskaya, M.L.; Bloch, D.A.; Han, H.; Gennis, R.B.; Verkhovsky, M.I. Aerobic respiratory chain of Escherichia coli is not allowed to work in fully uncoupled mode. Proc. Natl. Acad. Sci. USA 2011, 108, 17320–17324. [Google Scholar] [CrossRef] [Green Version]
- Jewett, M.C.; Miller, M.L.; Chen, Y.; Swartz, J.R. Continued protein synthesis at low [ATP] and [GTP] enables cell adaptation during energy limitation. J. Bacteriol. 2009, 191, 1083–1091. [Google Scholar] [CrossRef] [Green Version]
- Maier, B.; Chen, I.; Dubnau, D.; Sheetz, M.P. DNA transport into Bacillus subtilis requires proton motive force to generate large molecular forces. Nat. Struct. Mol. Biol. 2004, 11, 643–649. [Google Scholar] [CrossRef] [Green Version]
- Sohlenkamp, C.; Geiger, O. Bacterial membrane lipids: Diversity in structures and pathways. FEMS Microbiol. Rev. 2016, 40, 133–159. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.M.; Nielsen, K.M. Mechanisms of, and barriers to, horizontal gene transfer between bacteria. Nat. Rev. Microbiol. 2005, 3, 711–721. [Google Scholar] [CrossRef]
- Lu, J.; Wang, Y.; Li, J.; Mao, L.; Nguyen, S.H.; Duarte, T.; Coin, L.; Bond, P.; Yuan, Z.; Guo, J. Triclosan at environmentally relevant concentrations promotes horizontal transfer of multidrug resistance genes within and across bacterial genera. Environ. Int. 2018, 121, 1217–1226. [Google Scholar] [CrossRef]
- Bishop, R.E.; Leskiw, B.K.; Hodges, R.S.; Kay, C.M.; Weiner, J.H. The entericidin locus of Escherichia coli and its implications for programmed bacterial cell death. J. Mol. Biol. 1998, 280, 583–596. [Google Scholar] [CrossRef]
- Koebnik, R.; Locher, K.P.; Van Gelder, P. Structure and function of bacterial outer membrane proteins: Barrels in a nutshell. Mol. Microbiol. 2000, 37, 239–253. [Google Scholar] [CrossRef]
- Spector, J.; Zakharov, S.; Lill, Y.; Sharma, O.; Cramer, W.A.; Ritchie, K. Mobility of BtuB and OmpF in the Escherichia coli outer membrane: Implications for dynamic formation of a translocon complex. Biophys. J. 2010, 99, 3880–3886. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.; Li, S.; Zhang, S.; Yu, X. Reduction in horizontal transfer of conjugative plasmid by UV irradiation and low-level chlorination. Water Res. 2016, 91, 331–338. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Huang, Y.; Zhang, H.; Liu, Z.; Huan, Q.; Xiao, X.; Wang, Z. Factors and Mechanisms Influencing Conjugation In Vivo in the Gastrointestinal Tract Environment: A Review. Int. J. Mol. Sci. 2023, 24, 5919. https://doi.org/10.3390/ijms24065919
Liu W, Huang Y, Zhang H, Liu Z, Huan Q, Xiao X, Wang Z. Factors and Mechanisms Influencing Conjugation In Vivo in the Gastrointestinal Tract Environment: A Review. International Journal of Molecular Sciences. 2023; 24(6):5919. https://doi.org/10.3390/ijms24065919
Chicago/Turabian StyleLiu, Wei, Yanhu Huang, Han Zhang, Ziyi Liu, Quanmin Huan, Xia Xiao, and Zhiqiang Wang. 2023. "Factors and Mechanisms Influencing Conjugation In Vivo in the Gastrointestinal Tract Environment: A Review" International Journal of Molecular Sciences 24, no. 6: 5919. https://doi.org/10.3390/ijms24065919