
Bioinspired Lipase Immobilized Membrane for Improving Hesperidin Lipophilization
 , , , , , , and
, , , , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
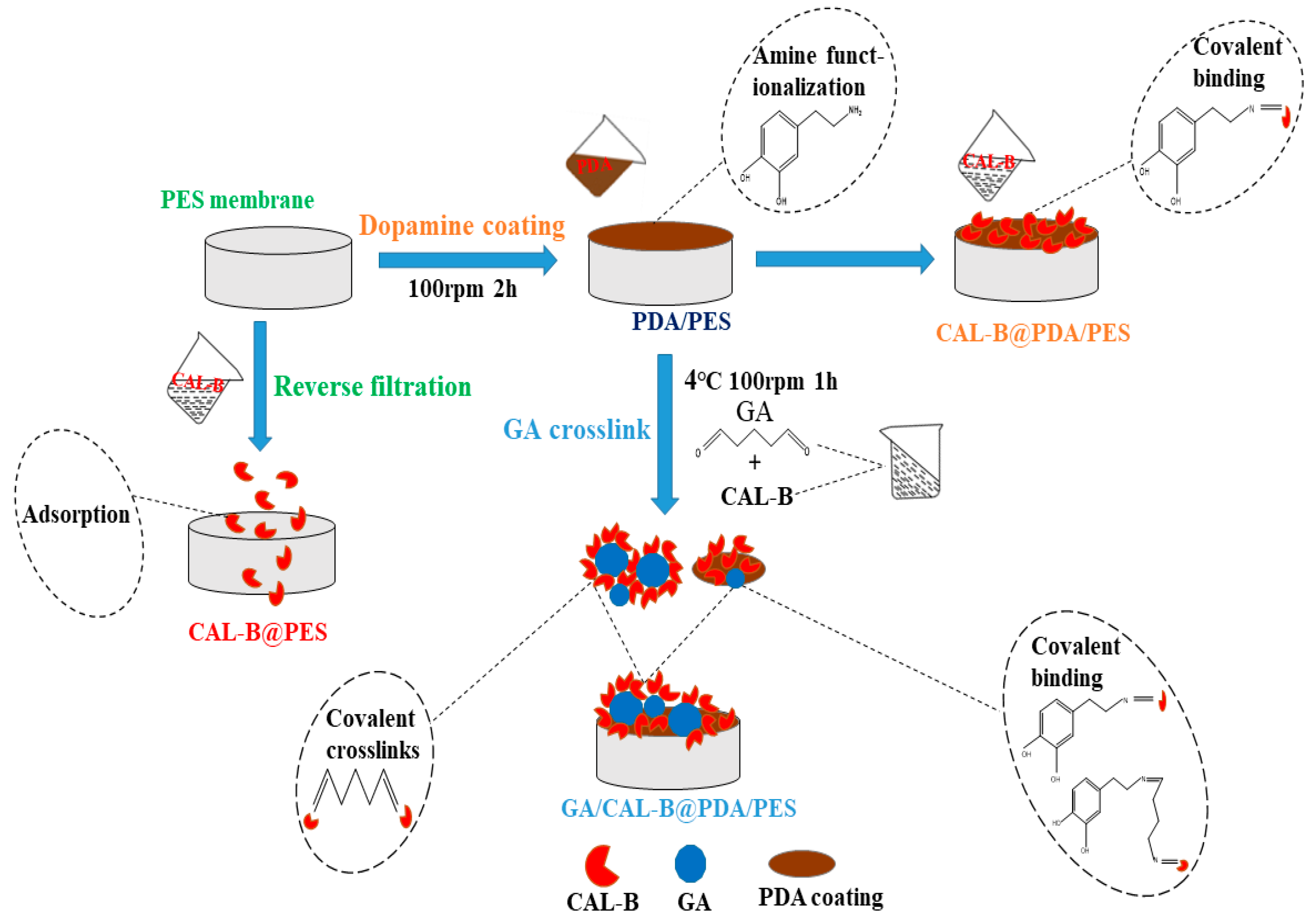
2.2. Fabrication of Lipase Immobilized Membranes
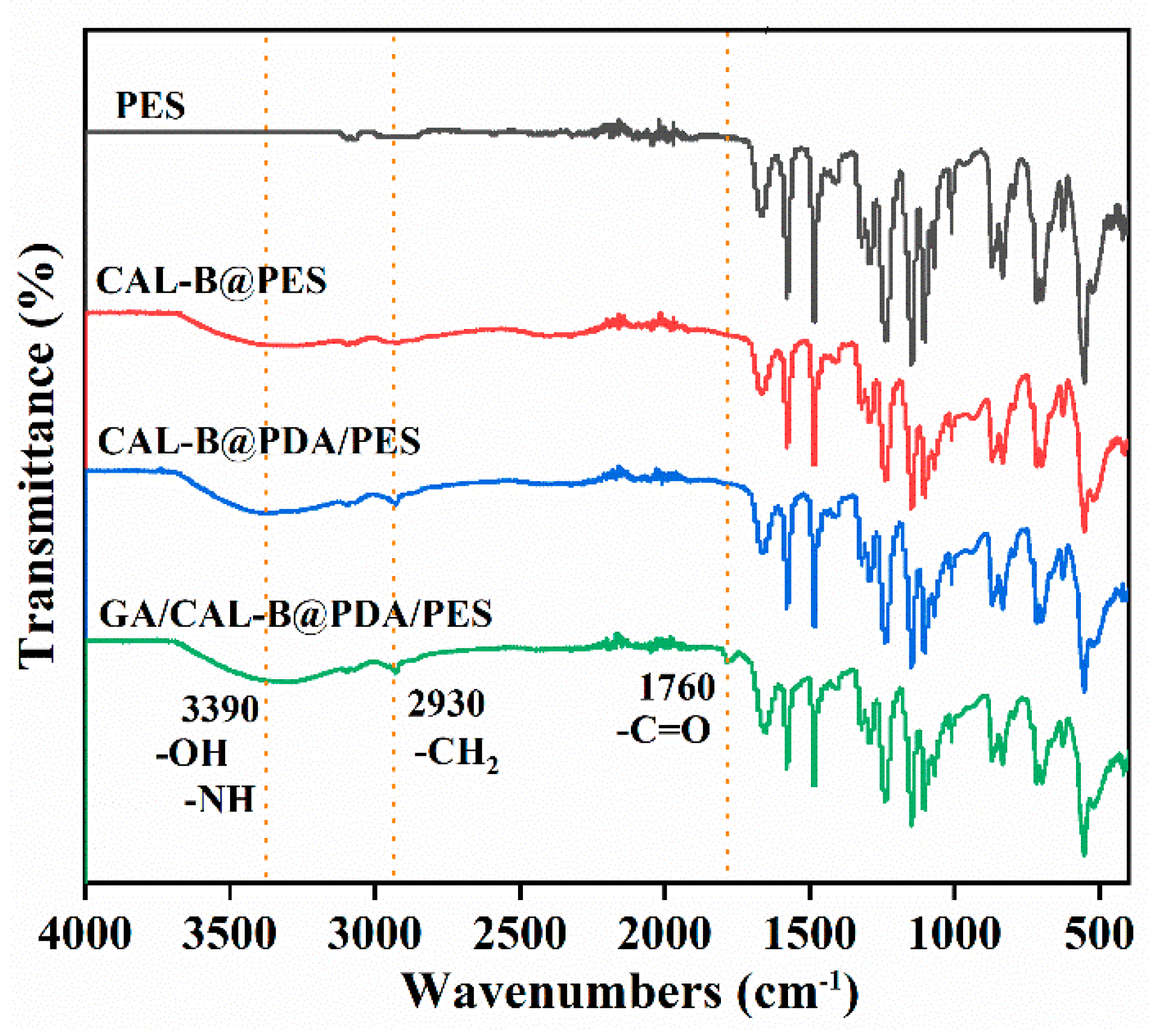
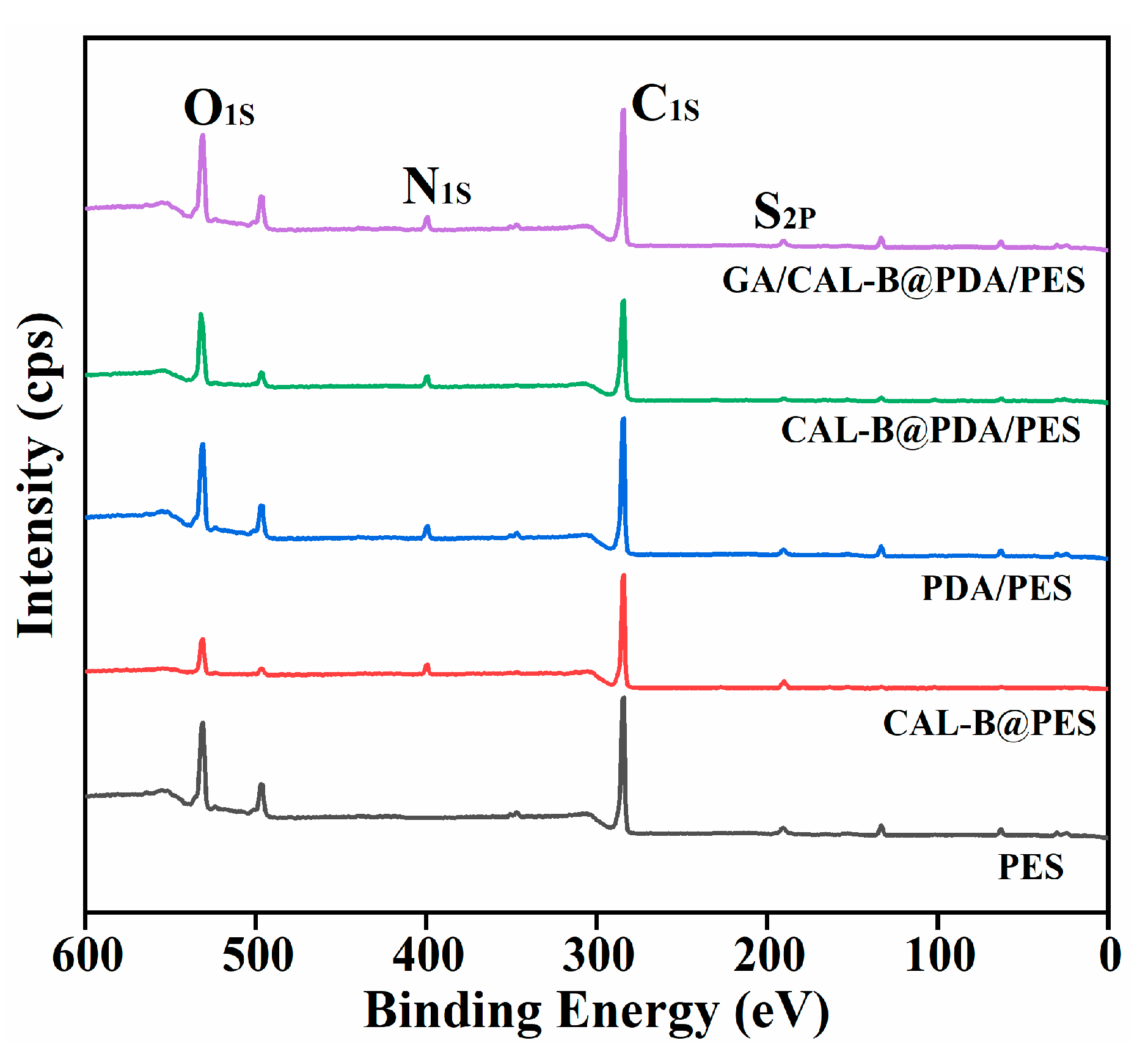
2.3. Characterization and Analysis
2.4. Measurement of CAL-B Esterification Activity
2.5. Stability Test of BLIMs
2.6. Enzymatic Esterification of Hesperidin with BLIMs
2.7. Data Analysis
3. Results
3.1. The Characterization of BLIMs
3.2. The BLIMs Fabrication
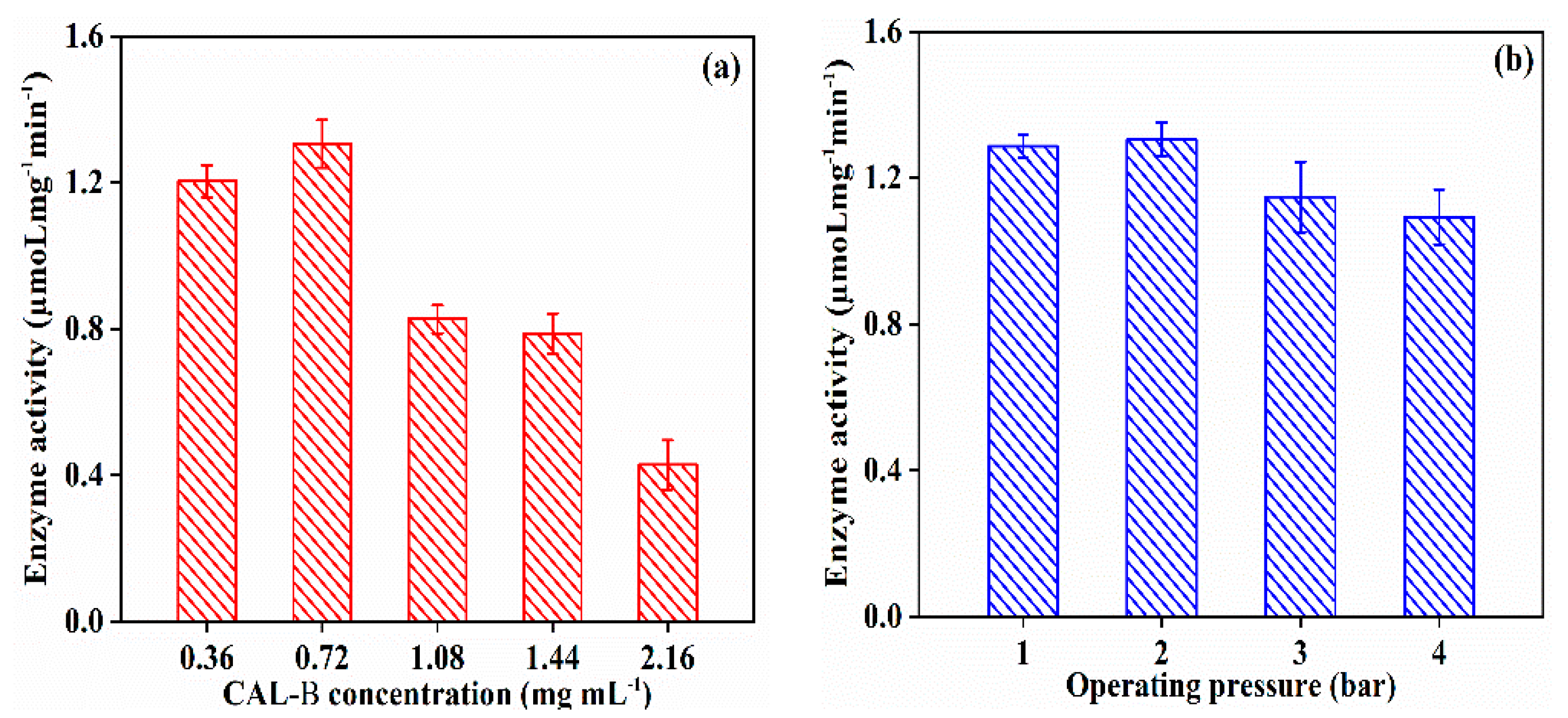
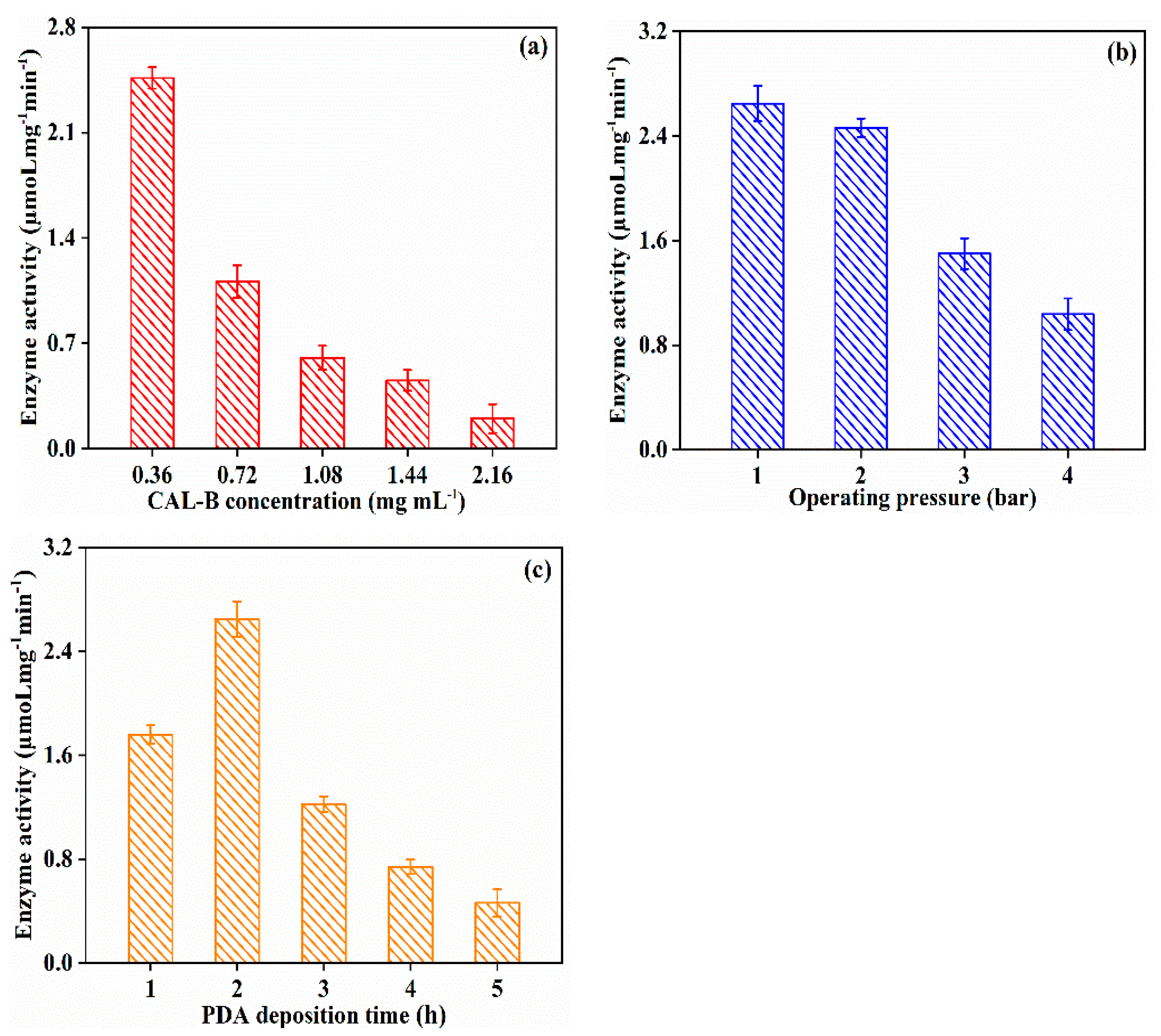
3.2.1. Effect of Fabrication Operating Condition on CAL-B@PES Performance
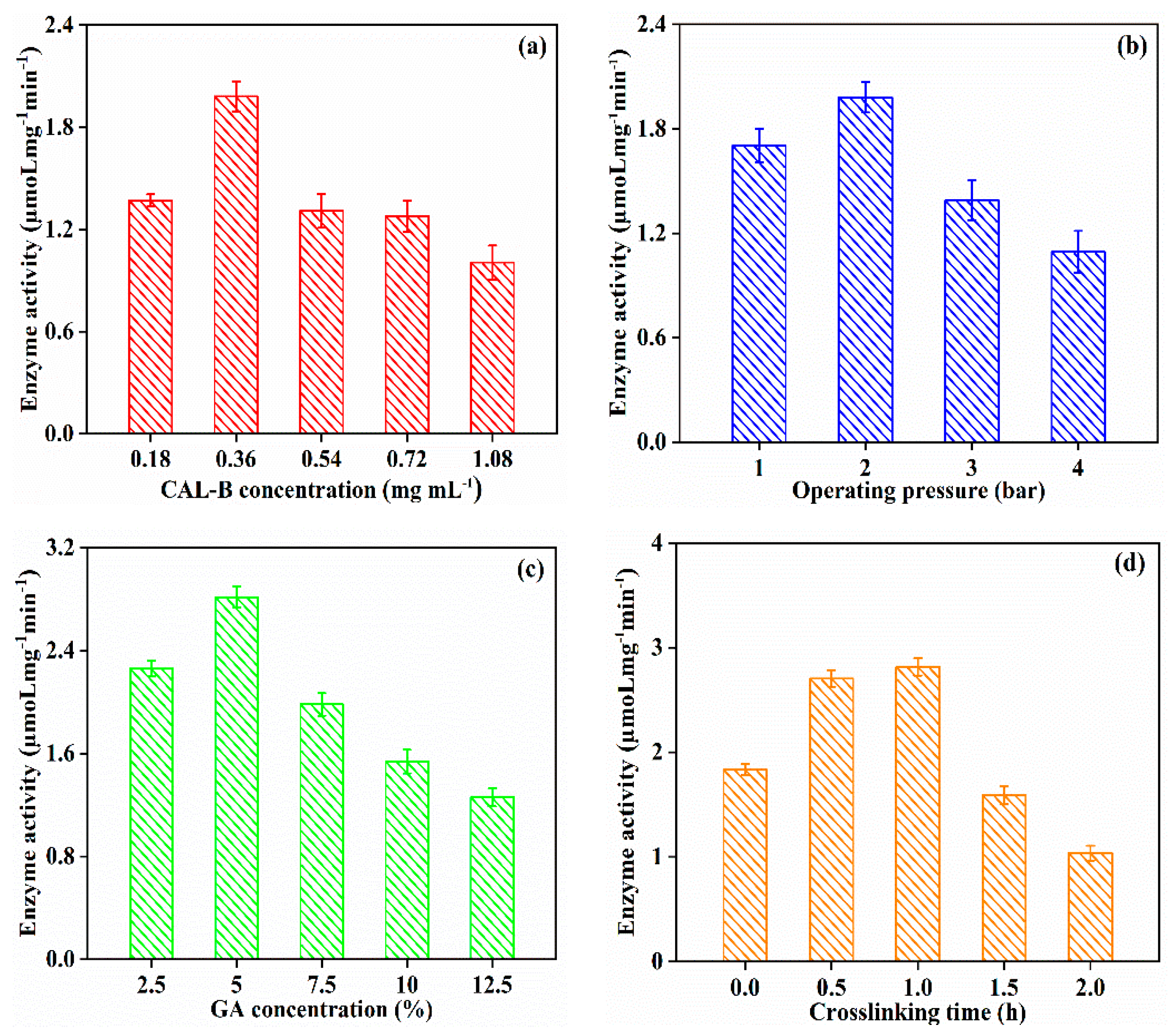
3.2.2. Effect of Fabrication Operating Condition on CAL-B@PDA/PES Performance
3.2.3. Effect of Fabrication Operating Condition on GA/CAL-B@PDA/PES Performance
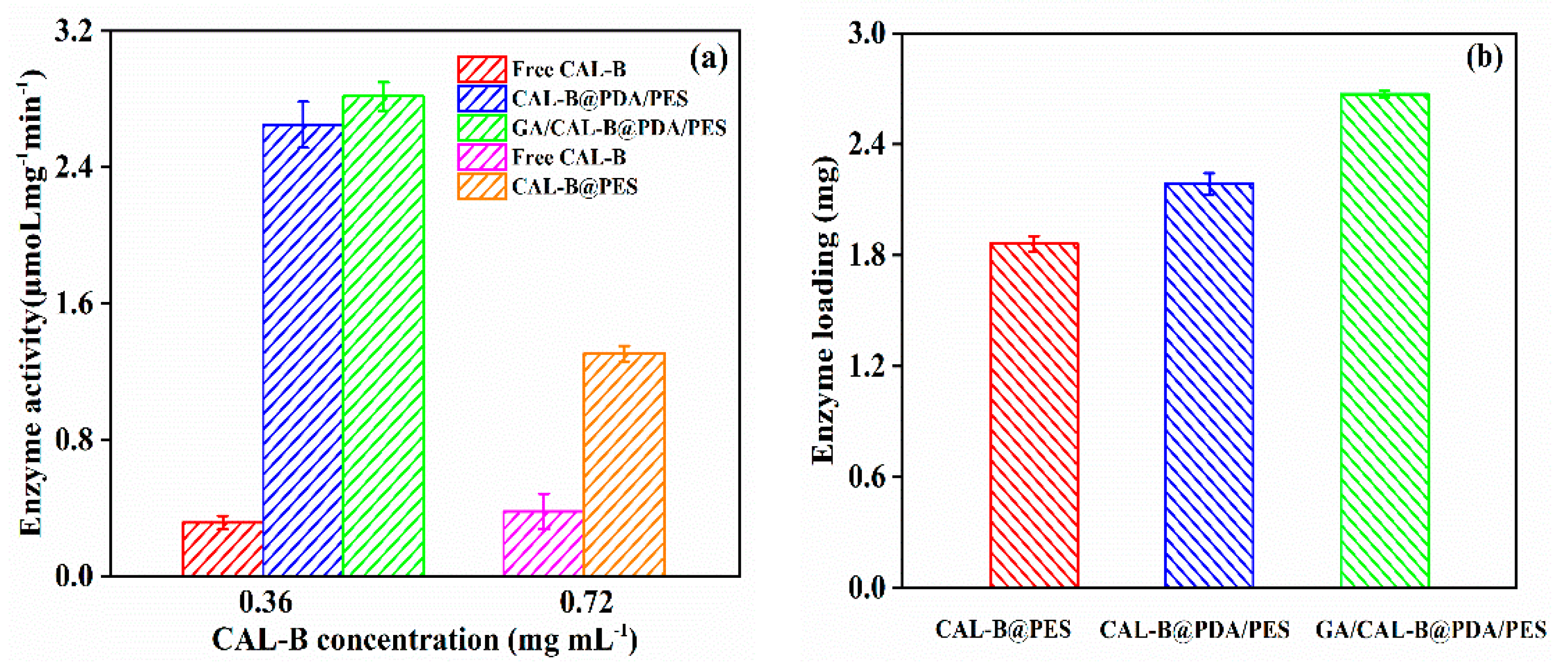
3.3. The Enzyme Activity and Enzyme Loading for Free and Immobilized CAL-B
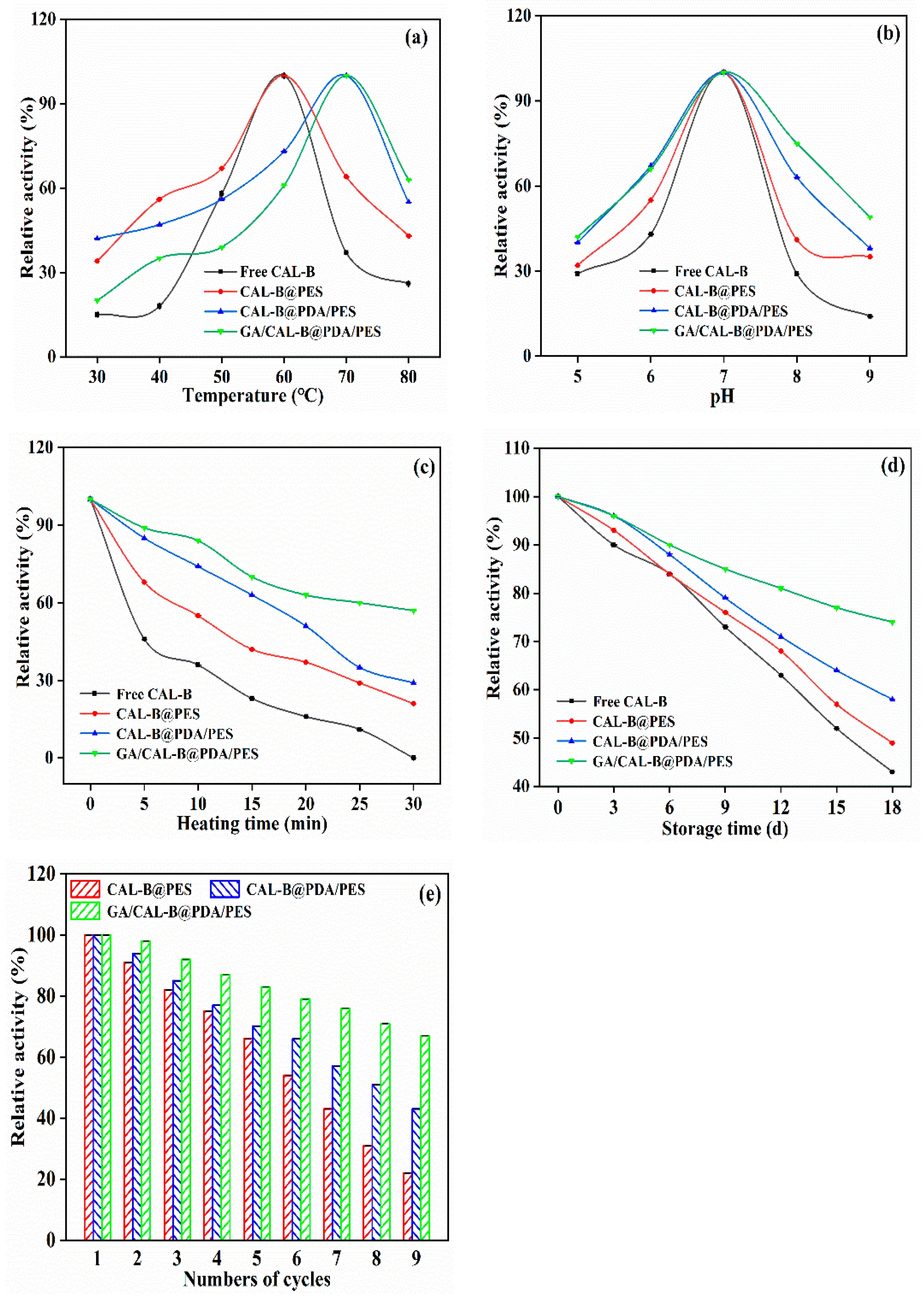
3.4. Stability Analysis of BLIMs
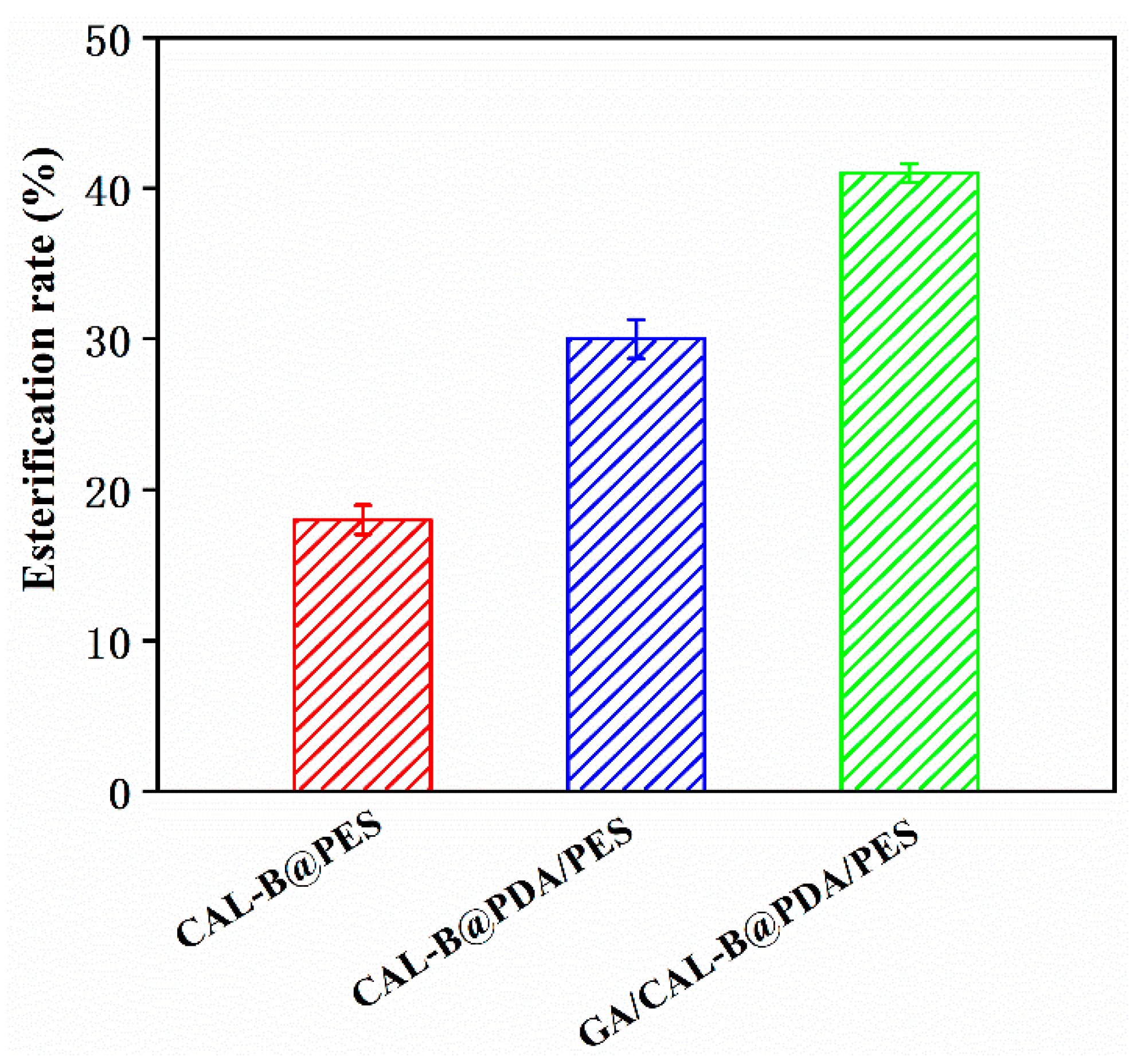
3.5. Hesperidin Lipophilization by BLIMs
3.6. BLIMs Performance
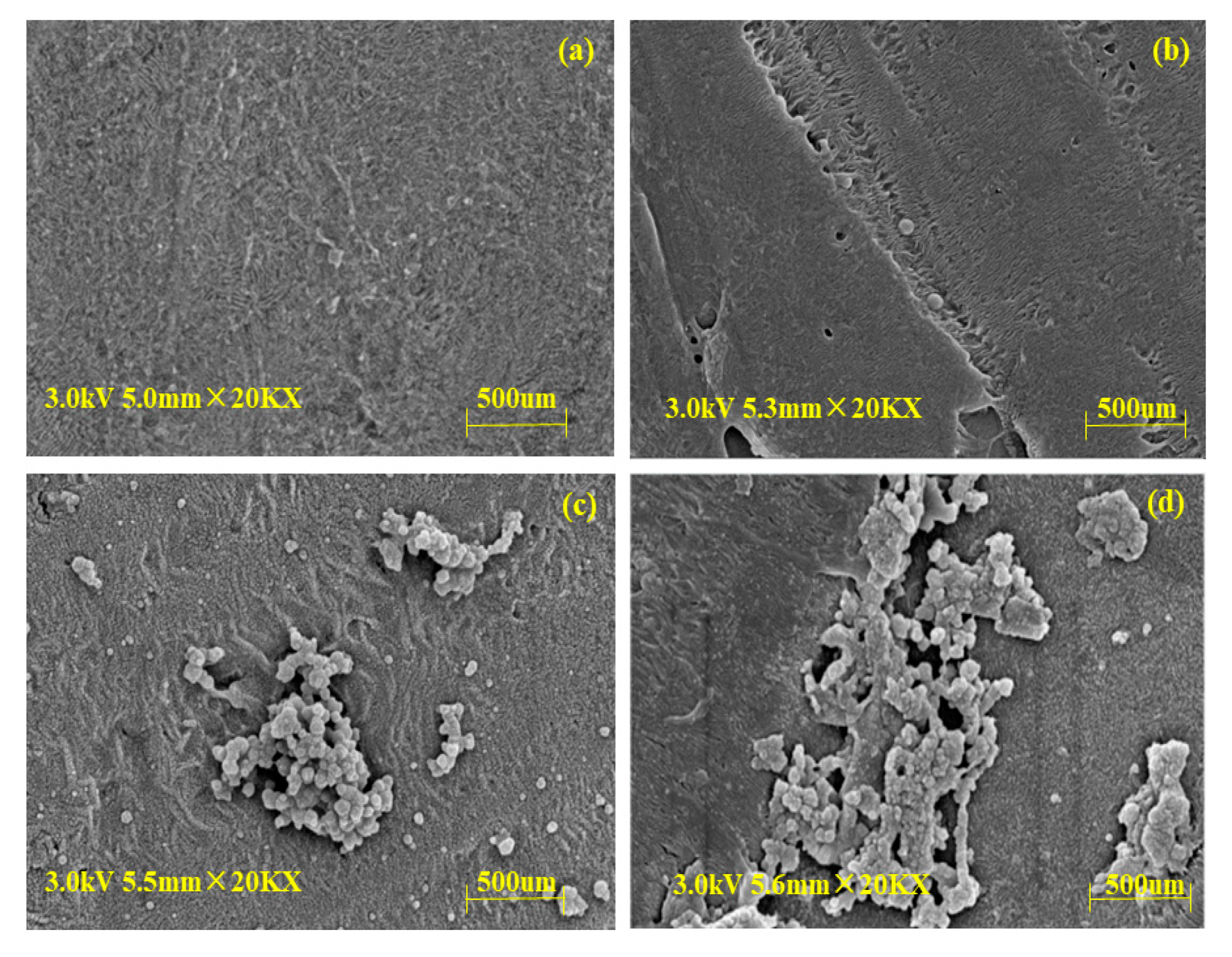
3.6.1. Morphologies of BLIMs after Esterification
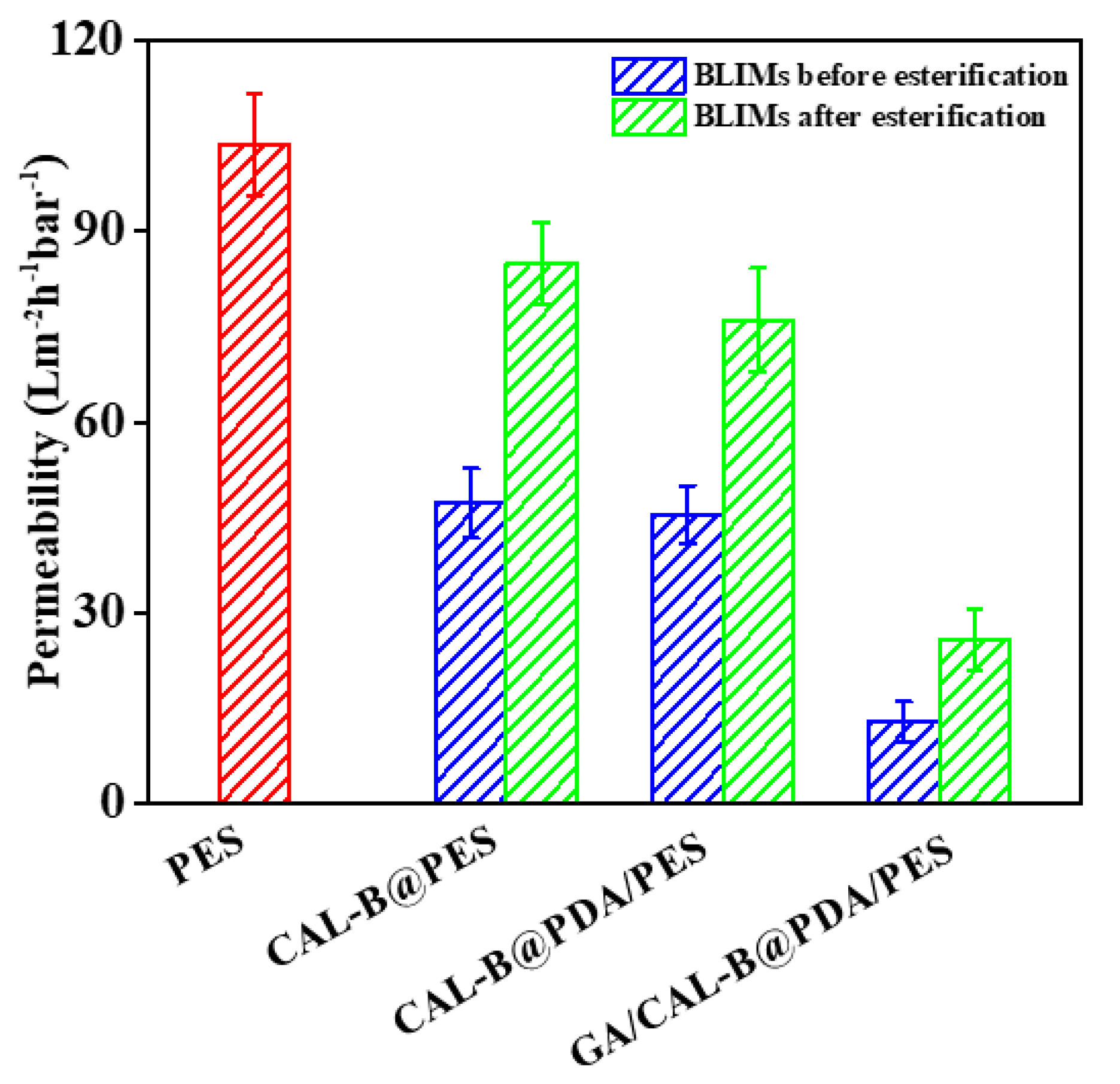
3.6.2. BLIMs Permeability
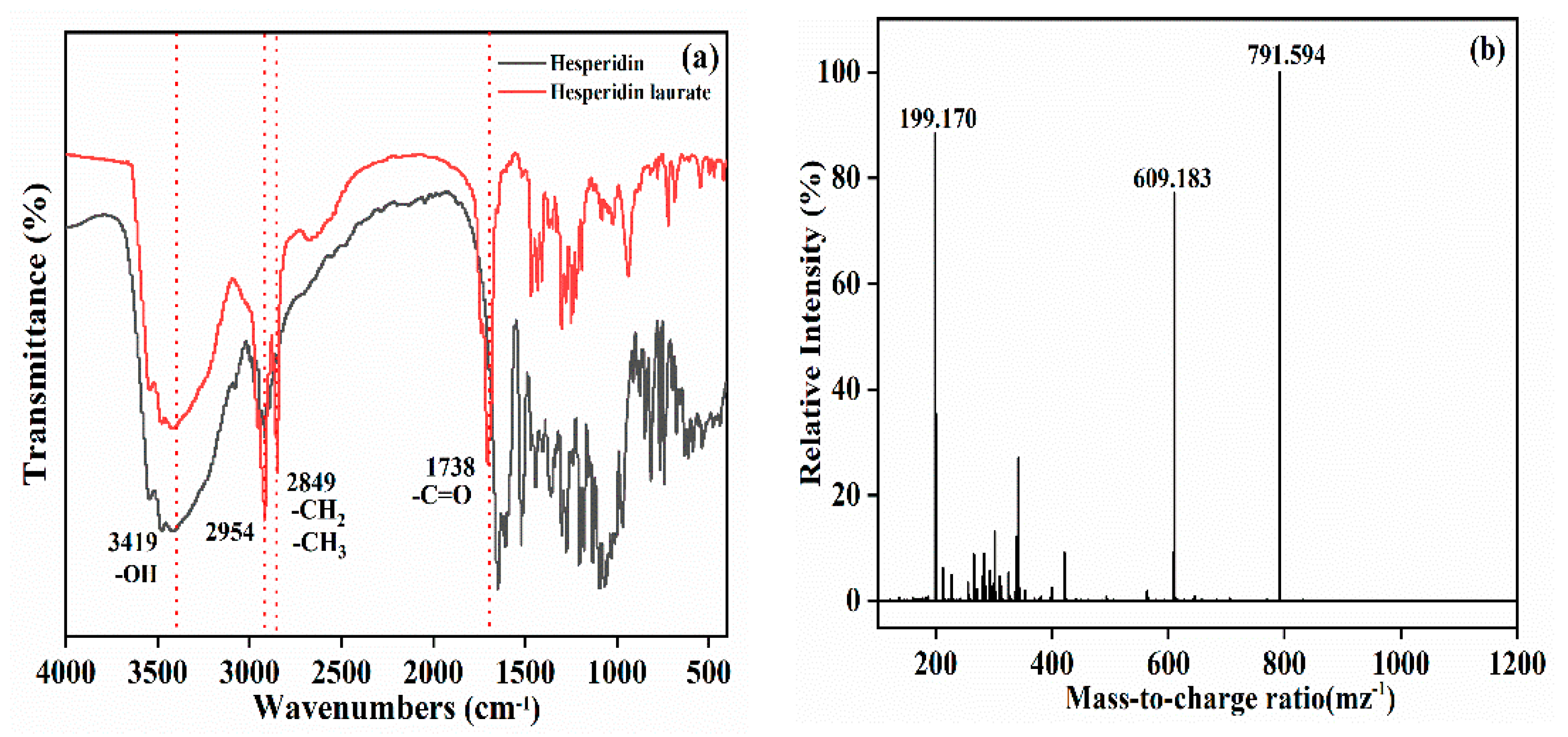
3.6.3. The Esterified Products
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lei, D.; Li, J.; Zhang, C.; Li, S.; Zhu, Z.; Wang, F.; Deng, Q.; Grimi, N. Complexation of soybean protein isolate with β-glucan and myricetin: Different affinity on 7S and 11S globulin by QCM-D and molecular simulation analysis. Food Chem. X 2022, 15, 100426. [Google Scholar] [CrossRef]
- Li, S.; Xu, H.; Sui, Y.; Mei, X.; Shi, J.; Cai, S.; Xiong, T.; Carrillo, C.; Castagnini, J.M.; Zhu, Z.; et al. Comparing the LC-MS phenolic acids profiles of seven different varieties of brown rice (Oryza sativa L.). Foods 2022, 11, 1552. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Huang, Y.; Huang, J.; Li, S.; Zhu, Z.; Deng, Q.; Cheng, S. Formation of protein-anthocyanin complex induced by grape skin extracts interacting with wheat gliadins: Multi-spectroscopy and molecular docking analysis. Food Chem. 2022, 385, 132702. [Google Scholar] [CrossRef]
- Gao, W.; Zhang, N.; Li, S.; Li, S.; Zhu, S.; Cong, X.; Cheng, S.; Barba, F.J.; Zhu, Z. Polysaccharides in selenium-enriched tea: Extraction performance under innovative technologies and antioxidant activities. Foods 2022, 11, 2545. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhu, S.; Yang, W.; Huang, Q.; Ho, C.-T. The biological fate and bioefficacy of citrus flavonoids: Bioavailability, biotransformation, and delivery systems. Food Funct. 2021, 12, 3307–3323. [Google Scholar] [CrossRef] [PubMed]
- Cao, R.; Zhao, Y.; Zhou, Z.; Zhao, X. Enhancement of the water solubility and antioxidant activity of hesperidin by chitooligosaccharide. J. Sci. Food Agric. 2017, 98, 2422–2427. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, H.J. Antioxidant activities of premature and mature mandarin (Citrus unshiu) peel and juice extracts. Food Sci. Biotechnol. 2022, 31, 627–633. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Sicari, V.; Tundis, R.; Leporini, M.; Falco, T.; Calabrò, V. The influence of ultrafiltration of Citrus limon L. burm. cv femminello comune juice on its chemical composition and antioxidant and hypoglycemic properties. Antioxidants 2019, 8, 23. [Google Scholar] [CrossRef]
- Araújo, M.E.M.B.; Contesini, F.J.; Franco, Y.E.M.; Sawaya, A.C.H.F.; Alberto, T.G.; Dalfré, N.; Carvalho, P.D.O. Optimized enzymatic synthesis of hesperidin fatty acid esters in a two-phase system containing lonic liquid. Molecules 2011, 16, 7171. [Google Scholar] [CrossRef]
- Sun, T.; Dong, Z.; Wang, J.; Huang, F.; Zheng, M. Ultrasound-assisted interfacial immobilization of lipase on hollow mesoporous silica spheres in a pickering emulsion system: A hyperactive and sustainable biocatalyst. ACS Sustain. Chem. Eng. 2020, 8, 17280–17290. [Google Scholar] [CrossRef]
- Jafarian, F.; Bordbar, A.-K.; Razmjou, A.; Zare, A. The fabrication of a high performance enzymatic hybrid membrane reactor (EHMR) containing immobilized candida rugosa lipase (CRL) onto graphene oxide nanosheets-blended polyethersulfone membrane. J. Membr. Sci. 2020, 613, 118435. [Google Scholar] [CrossRef]
- Thangaraj, B.; Solomon, P.R. Immobilization of lipases—A review. Part I: Enzyme immobilization. Chembioeng. Rev. 2019, 6, 157–166. [Google Scholar] [CrossRef]
- Widhyahrini, K.; Handayani, N.; Wahyuningrum, D.; Nurbaiti, S.; Radiman, C.L. The microwave-assisted synthesis of polyethersulfone (PES) as a matrix in immobilization of candida antarctica lipase B (Cal-B). Bull. Chem. React. Eng. 2017, 12, 343. [Google Scholar] [CrossRef]
- T.sriwong, K.; Matsuda, T. Recent advances in enzyme immobilization utilizing nanotechnology for biocatalysis. Org. Process Res. Dev. 2022, 26, 1857–1877. [Google Scholar] [CrossRef]
- Luo, J.; Song, S.; Zhang, H.; Zhang, H.; Zhang, J.; Wan, Y. Biocatalytic membrane: Go far beyond enzyme immobilization. Eng. Life Sci. 2020, 20, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Cao, T.; Pázmándi, M.; Galambos, I.; Kovács, Z. Continuous production of galacto-oligosaccharides by an enzyme membrane reactor utilizing free enzymes. Membranes 2020, 10, 203. [Google Scholar] [CrossRef]
- Sun, H.; Tang, B.; Wu, P. Development of hybrid ultrafiltration membranes with improved water separation properties using modified superhydrophilic metal-organic framework nanoparticles. ACS Appl. Mater. Interfaces 2017, 9, 21473–21484. [Google Scholar] [CrossRef]
- Fan, R.; Burghardt, J.P.; Prell, F.; Zorn, H.; Czermak, P. Production and purification of fructo-oligosaccharides using an enzyme membrane bioreactor and subsequent fermentation with probiotic bacillus coagulans. Sep. Purif. Technol. 2020, 251, 117291. [Google Scholar] [CrossRef]
- Guimarães, M.; Pérez-Gregorio, M.; Mateus, N.; Freitas, V.d.; Galinha, C.F.; Crespo, J.G.; Portugal, C.A.M.; Cruz, L. An efficient method for anthocyanins lipophilization based on enzyme retention in membrane systems. Food Chem. 2019, 300, 125167. [Google Scholar] [CrossRef]
- Thuanthong, M.; Gobba, C.D.; Sirinupong, N.; Youravong, W.; Otte, J. Purification and characterization of angiotensin-converting enzyme inhibitory peptides from nile tilapia (Oreochromis niloticus) skin gelatine produced by an enzymatic membrane reactor. J. Funct. Foods 2017, 36, 243–254. [Google Scholar] [CrossRef]
- Chen, Z.; Sun, Z.; Ming, S.; Li, S.; Zhu, Z.; Zhang, W. Bioinspired proteolytic membrane (BPM) with bilayer pepsin structure for protein hydrolysis. Sep. Purif. Technol. 2020, 259, 118214. [Google Scholar] [CrossRef]
- Wang, J.; Tian, J.; Gao, S.; Shi, W. Dopamine triggered one step polymerization and codeposition of reactive surfactant on PES membrane surface for antifouling modification. Sep. Purif. Technol. 2020, 249, 117148. [Google Scholar] [CrossRef]
- Muñiz-Mouro, A.; Gullón, B.; Lu-Chau, T.A.; Eibes, G. Green and sustainable synthesis of oligorutin using an enzymatic membrane reactor: Process optimization. Food Bioprod. Process 2020, 124, 434–444. [Google Scholar] [CrossRef]
- Rasouli, H.; Iliuta, I.; Bougie, F.; Garnier, A.; Iliuta, M.C. Enzyme-immobilized flat-sheet membrane contactor for green carbon capture. Chem. Eng. J. 2021, 421, 129587. [Google Scholar] [CrossRef]
- Zhang, H.; Luo, J.; Li, S.; Wei, Y.; Wan, Y. Biocatalytic membrane based on polydopamine coating: A platform for studying immobilization mechanisms. Langmuir 2018, 34, 2585–2594. [Google Scholar] [CrossRef]
- Su, Z.; Luo, J.; Sigurdardóttir, S.B.; Manferrari, T.; Jankowska, K.; Pinelo, M. An enzymatic membrane reactor for oligodextran production: Effects of enzyme immobilization strategies on dextranase activity. Carbohyd. Polym. 2021, 271, 118430. [Google Scholar] [CrossRef]
- Zhou, F.; Luo, J.; Song, S.; Wan, Y. Nanostructured polyphenol-mediated coating: A versatile platform for enzyme immobilization and micropollutant removal. Ind. Eng. Chem. Res. 2020, 59, 2708–2717. [Google Scholar] [CrossRef]
- Touqeer, T.; Mumtaz, M.W.; Mukhtar, H.; Irfan, A.; Akram, S.; Shabbir, A.; Rashid, U.; Nehdi, I.A.; Choong, T.S.Y. Fe3O4-PDA-lipase as surface functionalized nano biocatalyst for the production of biodiesel using waste cooking oil as feedstock: Characterization and process optimization. Energies 2019, 13, 177. [Google Scholar] [CrossRef]
- Li, Y.; Shi, S.; Cao, H.; Zhao, Z.; Su, C.; Wen, H. Improvement of the antifouling performance and stability of an anion exchange membrane by surface modification with graphene oxide (GO) and polydopamine (PDA). J. Membr. Sci. 2018, 566, 44–53. [Google Scholar] [CrossRef]
- Zarghami, S.; Mohammadi, T.; Sadrzadeh, M. Preparation, characterization and fouling analysis of in-air hydrophilic/underwater oleophobic bio-inspired polydopamine coated PES membranes for oily wastewater treatment. J. Membr. Sci. 2019, 582, 402–413. [Google Scholar] [CrossRef]
- Barbosa, O.; Ortiz, C.; Berenguer-Murcia, A.; Torres, R.; Rodrigues, R.C.; Fernandez-Lafuente, R. Glutaraldehyde in bio-catalysts design: A useful crosslinker and a versatile tool in enzyme immobilization. RSC Adv. 2013, 4, 1583–1600. [Google Scholar] [CrossRef]
- Neelam, V.; Lovely, S.; Anjum, G.; Vinita, H.; Vikas, H. Novel approach using activated cellulose film for efficient immobilization of purified diamine oxidase to enhance enzyme performance and stability. Prep. Biochem. Biotech. 2020, 50, 468–476. [Google Scholar]
- Xu, J.; Du, P.; Bi, W.; Yao, G.; Li, S.; Liu, H. Graphene oxide aerogels co-functionalized with polydopamine and polyethylenimine for the adsorption of anionic dyes and organic solvents. Chem. Eng. Res. Des 2019, 154, 192–202. [Google Scholar] [CrossRef]
- Kondabagil, K.; Karanth, N.G.; Sajja, H.K.; Divakar, S. An esterification method for determination of lipase activity. Biotechnol. Lett. 2000, 22, 1511–1514. [Google Scholar]
- Wang, F.; Luo, X.; Guo, J.; Zhang, W. Treatment of soy sauce wastewater with biomimetic dynamic membrane for colority removal and chemical oxygen demand lowering. An. Acad. Bras. Cienc. 2021, 93, e20210425. [Google Scholar] [CrossRef] [PubMed]
- Ardhaoui, M.; Falcimaigne, A.; Engasser, J.-M.; Moussou, P.; Pauly, G.; Ghoul, M. Acylation of natural flavonoids using lipase of candida antarctica as biocatalyst. J. Mol. Catal B Enzym. 2004, 29, 63–67. [Google Scholar] [CrossRef]
- Geng, X.; Wang, J.; Ye, J.; Yang, S.; Han, Q.; Lin, H.; Liu, F. Electrosprayed polydopamine membrane: Surface morphology, chemical stability and separation performance study. Sep. Purif. Technol. 2020, 244, 116857. [Google Scholar] [CrossRef]
- Zhu, L.; Jiang, J.; Zhu, B.; Xu, Y. Immobilization of bovine serum albumin onto porous polyethylene membranes using strongly attached polydopamine as a spacer. Colloids Surf. B Biointerfaces 2011, 86, 111–118. [Google Scholar] [CrossRef]
- Elias, N.; Wahab, R.A.; Jye, L.W.; Mahat, N.A.; Chandren, S.; Jamalis, J. Taguchi orthogonal design assisted immobilization of candida rugosa lipase onto nanocellulose-silica reinforced polyethersulfone membrane: Physicochemical characterization and operational stability. Cellulose 2021, 28, 5669–5691. [Google Scholar] [CrossRef]
- Huang, X.; Ge, D.; Xu, Z. Preparation and characterization of stable chitosan nanofibrous membrane for lipase immobilization. Eur. Polym. J. 2007, 43, 3710–3718. [Google Scholar] [CrossRef]
- Ye, P.; Jiang, J.; Xu, Z. Adsorption and activity of lipase from candida rugosa on the chitosan-modified poly(acrylonitrile-co-maleic acid) membrane surface. Colloids Surf. B Biointerfaces 2007, 60, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Asmat, S.; Anwer, A.H.; Husain, Q. Immobilization of lipase onto novel constructed polydopamine grafted multiwalled carbon nanotube impregnated with magnetic cobalt and its application in synthesis of fruit flavours. Int. J. Biol. Macromol. 2019, 140, 484–495. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.; Liu, J.; Wang, J.; Zhang, Y.; Zhang, B.; Zhang, Y.; Xiang, X.; Chen, R. Surface modification of halloysite nanotubes with dopamine for enzyme immobilization. ACS Appl. Mater. Interfaces 2013, 5, 10559–10564. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; He, W.; Shen, H.; Zhou, Z.; Li, M.; Su, P.; Yang, Y. Construction of multiple enzyme metal–organic frameworks biocatalyst via DNA scaffold: A promising strategy for enzyme encapsulation. Chem. Eng. J. 2019, 363, 174–182. [Google Scholar] [CrossRef]
- Luo, J.; Meyer, A.S.; Mateiu, R.V.; Kalyani, D.; Pinelo, M. Functionalization of a membrane sublayer using reverse filtration of enzymes and dopamine coating. ACS Appl. Mater. Interfaces 2014, 6, 22894–22904. [Google Scholar] [CrossRef] [PubMed]
- Emese, B.; Daniel, B.; Anamaria, T.; Péter, F.; Szilvia, K.; Tivadar, F. Recyclable solid-phase biocatalyst with improved stability by sol-gel entrapment of beta-D-galactosidase. J. Mol. Catal B. Enzym. 2016, 123, 81–90. [Google Scholar]
- Temkov, M.; Petrovski, A.; Gjorgieva, E.; Popovski, E.; Lazarova, M.; Boev, I.; Paunovic, P. Inulinase immobilization on polyethylene glycol/polypyrrole multiwall carbon nanotubes producing a catalyst with enhanced thermal and operational stability. Eng. Life Sci. 2019, 19, 617–630. [Google Scholar] [CrossRef]
- Luo, Q.; Huang, Q.; Chen, Z.; Yao, L. Temperature dependence of the pore structure in polyvinylidene fluoride (PVDF)/graphene composite membrane probed by electrochemical impedance spectroscopy. Polymers 2018, 10, 1123. [Google Scholar] [CrossRef]
- Wang, H.; Jia, C.; Xia, X.; Karangwa, E.; Zhang, X. Enzymatic synthesis of phytosteryl lipoate and its antioxidant properties. Food Chem. 2017, 240, 736–742. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elements (%) | PES | CAL-B@ PES | PDA/PES | CAL-B@PD A/PES | GA/CAL-B@ PDA/PES |
|---|---|---|---|---|---|
| C | 95.71 | 86.13 | 87.07 | 74.91 | 77.14 |
| N | 0 | 5.12 | 3.36 | 5.51 | 4.85 |
| O | 4.2 | 8.3 | 9.33 | 19.27 | 17.75 |
| S | 0.8 | 0.44 | 0.23 | 0.32 | 0.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ming, S.; Li, S.; Chen, Z.; Chen, X.; Wang, F.; Deng, S.; Marszałek, K.; Zhu, Z.; Zhang, W.; Barba, F.J. Bioinspired Lipase Immobilized Membrane for Improving Hesperidin Lipophilization. Antioxidants 2022, 11, 1906. https://doi.org/10.3390/antiox11101906
Ming S, Li S, Chen Z, Chen X, Wang F, Deng S, Marszałek K, Zhu Z, Zhang W, Barba FJ. Bioinspired Lipase Immobilized Membrane for Improving Hesperidin Lipophilization. Antioxidants. 2022; 11(10):1906. https://doi.org/10.3390/antiox11101906
Chicago/Turabian StyleMing, Shanxiu, Shuyi Li, Zhe Chen, Xujun Chen, Feifei Wang, Shaonan Deng, Krystian Marszałek, Zhenzhou Zhu, Wenxiang Zhang, and Francisco J. Barba. 2022. "Bioinspired Lipase Immobilized Membrane for Improving Hesperidin Lipophilization" Antioxidants 11, no. 10: 1906. https://doi.org/10.3390/antiox11101906