
Prevalence and Molecular Mechanisms of Carbapenem Resistance among Gram-Negative Bacilli in Three Hospitals of Northern Lebanon
 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Results
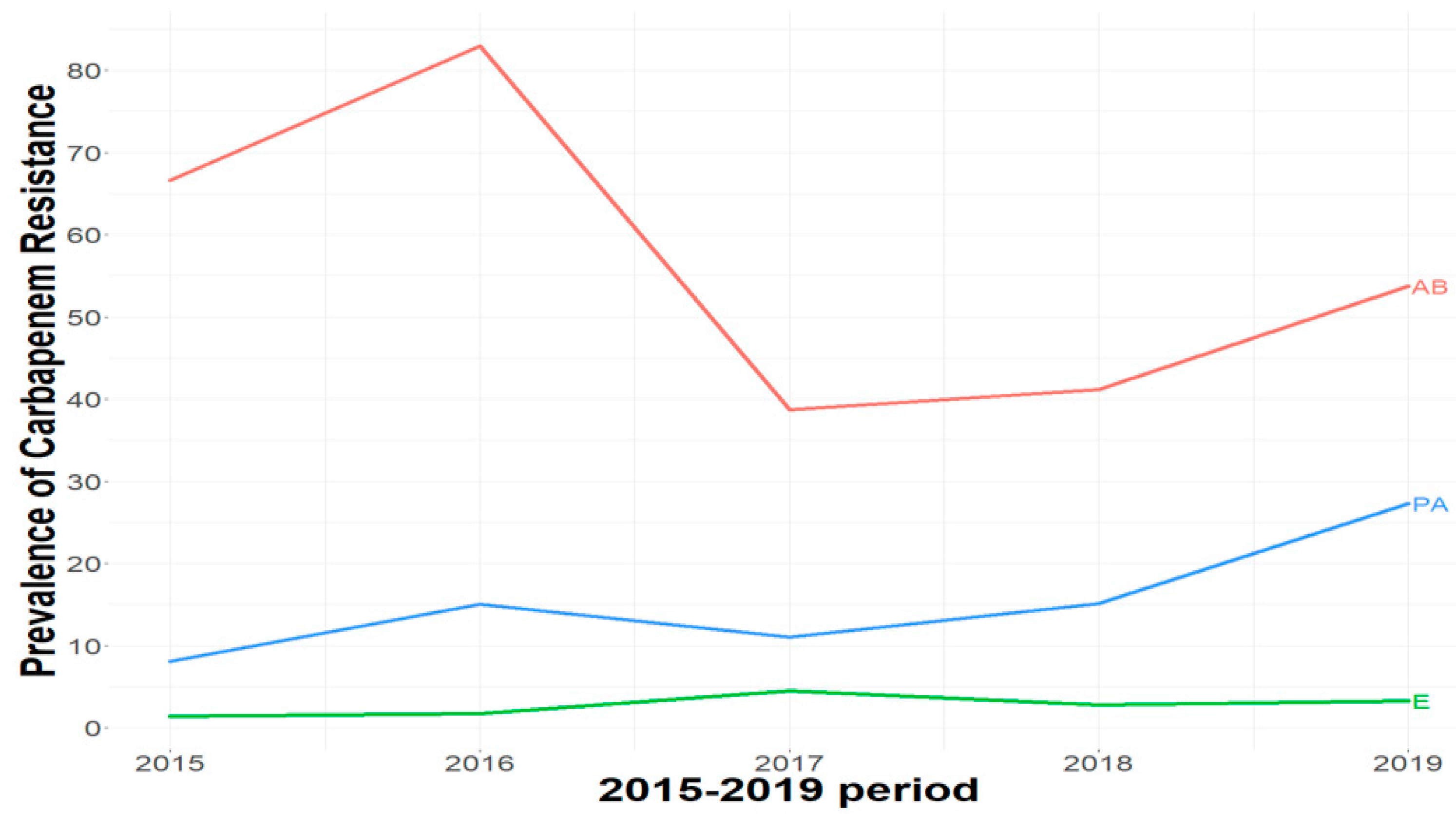
2.1. Carbapenem Resistance Levels among Gram-Negative Bacteria
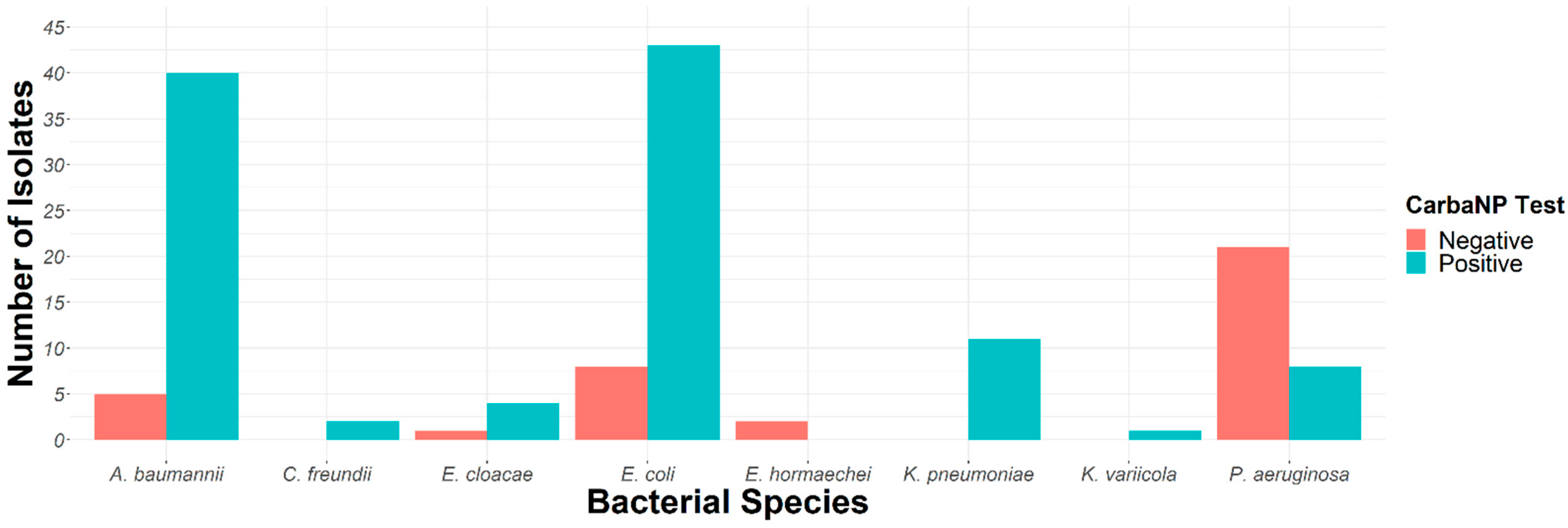
2.2. Carbapenemase Characterization
2.3. Whole-Genome Sequencing of Discrepant Results
3. Discussion
4. Materials and Methods
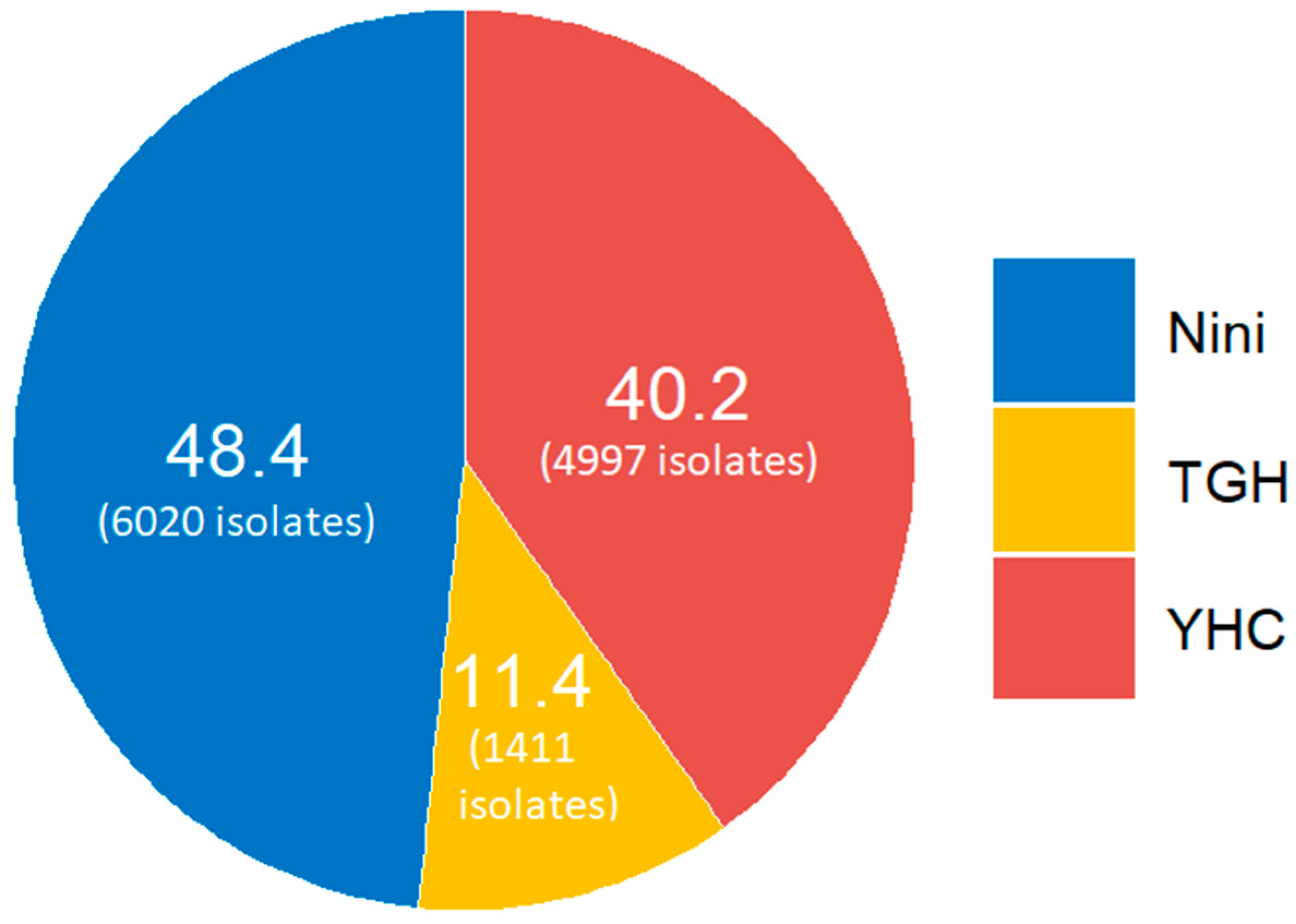
4.1. The Bacterial Isolates and Prevalence
4.2. Biochemical and Immunoenzymatic Assays for Carbapenemase Detection
4.3. The Lateral Flow Immunoassays (LFIA)
4.4. Molecular Detection of Carbapenemase Genes and Sanger Sequencing
4.5. Whole-Genome Sequencing
4.6. Nucleotide Sequence Accession Number
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morrill, H.J.; Pogue, J.M.; Kaye, K.S.; La Plante, K.L. Treatment Options for Carbapenem-Resistant Enterobacteriaceae Infections. Open Forum Infect. Dis. 2015, 2, ofv050. [Google Scholar] [CrossRef]
- Rostami, S.; Farajzadeh Sheikh, A.; Shoja, S.; Farahani, A.; Tabatabaiefar, M.A.; Jolodar, A.; Sheikhi, R. Investigating of four main carbapenem-resistance mechanisms in high-level carbapenem resistant Pseudomonas aeruginosa isolated from burn patients. J. Chin. Med. Assoc. JCMA 2018, 81, 127–132. [Google Scholar] [CrossRef]
- Ambler, R.P. The structure of beta-lactamases. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1980, 289, 321–331. [Google Scholar] [CrossRef]
- Djahmi, N.; Dunyach-Remy, C.; Pantel, A.; Dekhil, M.; Sotto, A.; Lavigne, J.P. Epidemiology of carbapenemase-producing Enterobacteriaceae and Acinetobacter baumannii in Mediterranean countries. BioMed. Res. Int. 2014, 2014, 305784. [Google Scholar] [CrossRef]
- Moghnieh, R.; Araj, G.F.; Awad, L.; Daoud, Z.; Mokhbat, J.E.; Jisr, T.; Abdallah, D.; Azar, N.; Irani-Hakimeh, N.; Balkis, M.M.; et al. A compilation of antimicrobial susceptibility data from a network of 13 Lebanese hospitals reflecting the national situation during 2015–2016. Antimicrob. Resist. Infect Control. 2019, 8, 41. [Google Scholar] [CrossRef]
- Beyrouthy, R.; Robin, F.; Dabboussi, F.; Mallat, H.; Hamzé, M.; Bonnet, R. Carbapenemase and virulence factors of Enterobacteriaceae in North Lebanon between 2008 and 2012: Evolution via endemic spread of OXA-48. J. Antimicrob. Chemother. 2014, 69, 2699–2705. [Google Scholar] [CrossRef]
- Chamoun, K.; Farah, M.; Araj, G.; Daoud, Z.; Moghnieh, R.; Salameh, P.; Saade, D.; Mokhbat, J.; Abboud, E.; Hamze, M.; et al. Surveillance of antimicrobial resistance in Lebanese hospitals: Retrospective nationwide compiled data. Int. J. Infect. Dis. 2016, 46, 64–70. [Google Scholar] [CrossRef]
- Yaghi, J.; Fattouh, N.; Akkawi, C.; Chamy, L.E.; Maroun, R.G.; Khalil, G. Unusually High Prevalence of Cosecretion of Ambler Class A and B Carbapenemases and Nonenzymatic Mechanisms in Multidrug-Resistant Clinical Isolates of Pseudomonas aeruginosa in Lebanon. Microb. Drug Resist. 2019, 26, 150–159. [Google Scholar] [CrossRef]
- Osman, M.; Halimeh, F.B.; Rafei, R.; Mallat, H.; Tom, J.E.; Raad, E.B.; Hamze, M. Investigation of an XDR-Acinetobacter baumannii ST2 outbreak in an intensive care unit of a Lebanese tertiary care hospital. Future Microbiol. 2020, 15, 1535–1542. [Google Scholar] [CrossRef]
- Dahdouh, E.; Hajjar, M.; Suarez, M.; Daoud, Z. Acinetobacter baumannii Isolated from Lebanese Patients: Phenotypes and Genotypes of Resistance, Clonality, and Determinants of Pathogenicity. Front. Cell. Infect. Microbiol. 2016, 6, 163. [Google Scholar] [CrossRef] [Green Version]
- Osman, M.; Al Mir, H.; Rafei, R.; Dabboussi, F.; Madec, J.Y.; Haenni, M.; Hamze, M. Epidemiology of antimicrobial resistance in Lebanese extra-hospital settings: An overview. J. Glob. Antimicrob. Resist. 2019, 17, 123–129. [Google Scholar] [CrossRef]
- Hamze, M. Epidemiology and Antibiotic Susceptibility Patterns of Carbapenem Resistant Gram Negative Bacteria Isolated from Two Tertiary Care Hospitals in North Lebanon. Int. Arab. J. Antimicrob. Agents 2018, 8. [Google Scholar] [CrossRef]
- Vedel, G. Simple method to determine β-lactam resistance phenotypes in Pseudomonas aeruginosa using the disc agar diffusion test. J. Antimicrob. Chemother. 2005, 56, 657–664. [Google Scholar] [CrossRef]
- Boutal, H.; Vogel, A.; Bernabeu, S.; Devilliers, K.; Creton, E.; Cotellon, G.; Plaisance, M.; Oueslati, S.; Dortet, L.; Jousset, A.; et al. A multiplex lateral flow immunoassay for the rapid identification of NDM-, KPC-, IMP- and VIM-type and OXA-48-like carbapenemase-producing Enterobacteriaceae. J. Antimicrob. Chemother. 2018, 73, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Hammoudi, D.; Ayoub Moubareck, C.; Aires, J.; Adaime, A.; Barakat, A.; Fayad, N.; Hakime, N.; Houmani, M.; Itani, T.; Najjar, Z.; et al. Countrywide spread of OXA-48 carbapenemase in Lebanon: Surveillance and genetic characterization of carbapenem-non-susceptible Enterobacteriaceae in 10 hospitals over a one-year period. Int. J. Infect. Dis. 2014, 29, 139–144. [Google Scholar] [CrossRef]
- Nawfal Dagher, T.; Azar, E.; Al-Bayssari, C.; Chamieh, A.S.; Rolain, J.M. First Detection of Colistin-Resistant Klebsiella pneumoniae in Association with NDM-5 Carbapenemase Isolated from Clinical Lebanese Patients. Microb. Drug Resist. 2019, 25, 925–930. [Google Scholar] [CrossRef]
- El-Herte, R.I.; Kanj, S.S.; Matar, G.M.; Araj, G.F. The threat of carbapenem-resistant Enterobacteriaceae in Lebanon: An update on the regional and local epidemiology. J. Infect. Public Health 2012, 5, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Diab, M.; Hamze, M.; Bonnet, R.; Saras, E.; Madec, J.Y.; Haenni, M. OXA-48 and CTX-M-15 extended-spectrum beta-lactamases in raw milk in Lebanon: Epidemic spread of dominant Klebsiella pneumoniae clones. J. Med. Microbiol. 2017, 66, 1688–1691. [Google Scholar] [CrossRef]
- Arabaghian, H.; Salloum, T.; Alousi, S.; Panossian, B.; Araj, G.F.; Tokajian, S. Molecular Characterization of Carbapenem Resistant Klebsiella pneumoniae and Klebsiella quasipneumoniae Isolated from Lebanon. Sci. Rep. 2019, 9, 531. [Google Scholar] [CrossRef]
- Voulgari, E.; Poulou, A.; Dimitroulia, E.; Politi, L.; Ranellou, K.; Gennimata, V.; Markou, F.; Pournaras, S.; Tsakris, A. Emergence of OXA-162 Carbapenemase- and DHA-1 AmpC Cephalosporinase-Producing Sequence Type 11 Klebsiella pneumoniae Causing Community-Onset Infection in Greece. Antimicrob. Agents Chemother. 2015, 60, 1862–1864. [Google Scholar] [CrossRef] [Green Version]
- Al Atrouni, A.; Hamze, M.; Jisr, T.; Lemarié, C.; Eveillard, M.; Joly-Guillou, M.-L.; Kempf, M. Wide spread of OXA-23-producing carbapenem-resistant Acinetobacter baumannii belonging to clonal complex II in different hospitals in Lebanon. Int. J. Infect. Dis. 2016, 52, 29–36. [Google Scholar] [CrossRef]
- Jousset, A.B.; Oueslati, S.; Bernabeu, S.; Takissian, J.; Creton, E.; Vogel, A.; Sauvadet, A.; Cotellon, G.; Gauthier, L.; Bonnin, R.A.; et al. False-Positive Carbapenem-Hydrolyzing Confirmatory Tests Due to ACT-28, a Chromosomally Encoded AmpC with Weak Carbapenemase Activity from Enterobacter kobei. Antimicrob. Agents Chemother. 2019, 63, e02388-18. [Google Scholar] [CrossRef]
- Girlich, D.; Naas, T.; Nordmann, P. Biochemical characterization of the naturally occurring oxacillinase OXA-50 of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2004, 48, 2043–2048. [Google Scholar] [CrossRef]
- Evans, B.A.; Amyes, S.G. OXA beta-lactamases. Clin. Microbiol. Rev. 2014, 27, 241–263. [Google Scholar] [CrossRef]
- Al-Mir, H.; Osman, M.; Drapeau, A.; Hamze, M.; Madec, J.-Y.; Haenni, M. WGS Analysis of Clonal and Plasmidic Epidemiology of Colistin-Resistance Mediated by mcr Genes in the Poultry Sector in Lebanon. Front. Microbiol. 2021, 12, 624194. [Google Scholar] [CrossRef]
- Al-Mir, H.; Osman, M. Spread of ESC-, carbapenem- and colistin-resistant Escherichia coli clones and plasmids within and between food workers in Lebanon. J. Antimicrob. Chemother. 2021, 76, 3135–3143. [Google Scholar] [CrossRef]
- Dabbousi, A.A.; Dabboussi, F.; Hamze, M.; Osman, M. The Emergence and Dissemination of Multidrug Resistant Pseudomonas aeruginosa in Lebanon: Current Status and Challenges during the Economic Crisis. Antibiotics 2022, 11, 687. [Google Scholar] [CrossRef]
- Dagher, L.A.; Hassan, J.; Kharroubi, S. Nationwide Assessment of Water Quality in Rivers across Lebanon by Quantifying Fecal Indicators Densities and Profiling Antibiotic Resistance of Escherichia coli. Antibiotics 2021, 10, 883. [Google Scholar] [CrossRef]
- Osman, M.; Cummings, K.J.; El Omari, K.; Kassem, I.I. Catch-22: War, Refugees, COVID-19, and the Scourge of Antimicrobial Resistance. Front. Med. 2022, 9, 921921. [Google Scholar] [CrossRef]
- Kassem, I.I.; Osman, M. A brewing storm: The impact of economic collapse on the access to antimicrobials in Lebanon. J. Glob. Antimicrob. Resist. 2022, 29, 313–315. [Google Scholar] [CrossRef]
- Osman, M.; Kasir, D.; Kassem, I.I.; Hamze, M. Shortage of appropriate diagnostics for antimicrobial resistance in Lebanese clinical settings: A crisis amplified by COVID-19 and economic collapse. J. Glob. Antimicrob. Resist. 2021, 27, 72–74. [Google Scholar] [CrossRef]
- Osman, M.; Rafei, R. Antimicrobial resistance in the protracted Syrian conflict: Halting a war in the war. Future Microbiol. 2021, 16, 825–845. [Google Scholar] [CrossRef]
- Al-Mir, H.; Osman, M.; Azar, N.; Madec, J.Y.; Hamze, M.; Haenni, M. Emergence of clinical mcr-1-positive Escherichia coli in Lebanon. J. Glob. Antimicrob. Resist. 2019, 19, 83–84. [Google Scholar] [CrossRef]
- Hassan, J.; Mann, D.; Li, S.; Deng, X. Emergence of the Mobile Colistin Resistance Gene, mcr-1, in Multidrug-Resistant E. coli Isolated from the Fecal Matter of Toddlers in a Community. Antimicrob. Agents Chemother. 2021, 65, e00243-21. [Google Scholar] [CrossRef] [PubMed]
- Sourenian, T.; Mann, D.; Li, S.; Deng, X.; Jaafar, H.; Kassem, I.I. Dissemination of multidrug-resistant Escherichia coli harboring the mobile colistin resistance gene mcr-1.1 on transmissible plasmids in the Mediterranean Sea. J. Glob. Antimicrob. Resist. 2020, 22, 84–86. [Google Scholar] [CrossRef] [PubMed]
- Hmede, Z.; Sulaiman, A.A.A.; Jaafar, H.; Kassem, I.I. Emergence of plasmid-borne colistin resistance gene mcr-1 in multidrug-resistant Escherichia coli isolated from irrigation water in Lebanon. Int. J. Antimicrob. Agents 2019, 54, 102–104. [Google Scholar] [CrossRef]
- Alhaj Sulaiman, A.A.; Kassem, I.I. First report of the plasmid-borne colistin resistance gene (mcr-1) in Proteus mirabilis isolated from domestic and sewer waters in Syrian refugee camps. Travel Med. Infect. Dis. 2020, 33, 101482. [Google Scholar] [CrossRef]
- CASFM, E. European Society of Clinical Microbiology and Infectious Disease Guidelines. 2019. Available online: https://www.sfm-microbiologie.org/wp-content/uploads/2022/05/CASFM2022_V1.0.pdf? (accessed on 28 July 2022).
- Nordmann, P.; Poirel, L.; Dortet, L. Rapid detection of carbapenemase-producing Enterobacteriaceae. Emerg. Infect. Dis. 2012, 18, 1503–1507. [Google Scholar] [CrossRef]
- Dortet, L.; Poirel, L.; Errera, C.; Nordmann, P. CarbAcineto NP test for rapid detection of carbapenemase-producing Acinetobacter spp. J. Clin. Microbiol. 2014, 52, 2359–2364. [Google Scholar] [CrossRef]
- Çekin, Z.K.; Dabos, L.; Malkoçoğlu, G.; Fortineau, N.; Bayraktar, B.; Iorga, B.I.; Naas, T.; Aktaş, E. Carbapenemase -producing Pseudomonas aeruginosa isolates from Turkey: First report of P. aeruginosa high-risk clones with VIM-5- and IMP-7-type carbapenemases in a tertiary hospital. Diagn. Microbiol. Infect. Dis. 2021, 99, 115174. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Year | Enterobacterales | P. aeruginosa | A. baumannii | |||
|---|---|---|---|---|---|---|
| N | Resistant isolates (%) a | N | Resistant isolates (%) a | N | Resistant isolates (%) a | |
| 2015 | 2081 | 31 (1.4%) | 244 | 20 (8.1%) | 18 | 12 (66.7%) |
| 2016 | 2157 | 37 (1.7%) | 193 | 29 (15%) | 47 | 39 (83%) |
| 2017 | 1696 | 78 (4.5%) | 172 | 19 (11%) | 31 | 12 (38.7%) |
| 2018 | 2125 | 60 (2.8%) | 238 | 36 (15.1%) | 34 | 14 (41.2%) |
| 2019 | 3151 | 105 (3.3%) | 187 | 51 (27.3%) | 54 | 29 (53.7%) |
| Total | 11,210 | 311 (2.7%) | 1034 | 155 (15%) | 184 | 106 (58%) |
| Bacteria | Number of Isolates | CarbaNP-Positive Isolates | Carbapenemases (Sanger Sequencing) |
|---|---|---|---|
| K. pneumoniae | n = 11 | 11 (100%) | OXA-48 (6) OXA-181 (1) NDM-5 (1) (3) a |
| K. variicola | n = 1 | 1 (100%) | (1) a |
| E. cloacae | n = 5 | 4 (80%) | OXA-48 (1) VIM-4 (1) VIM-1 (1) NDM-1 (1) |
| E. hormaechei | n = 2 | 0 | - |
| C. freundii | n = 2 | 2 (100%) | OXA-48 (2) |
| E. coli | n = 51 | 43 (84%) | OXA-48 (11) OXA-162 (1) OXA-181 (12) OXA-244 (6) NDM-5 (4) NDM-19 (9) |
| P. aeruginosa | n = 29 | 8 (27%) | VIM-62 (2) VIM-2 (2) IMP-15 (1) (3) a |
| A. baumannii | n = 45 | 33 (73%) | OXA-23-LIKE (31) (2) a |
| Species | MICs (μg/mL) | Phenotypic Tests | Resfinder | MLST | Accession Numbers | |||
|---|---|---|---|---|---|---|---|---|
| IMP | MER | Carba NP | NG-Test Carba5 | ß-Lactamases Genes | Other Resistant Genes | |||
| K. pneumoniae 1 | 0.19 | 0.032 | + | - | blaCTX-M-15, blaSHV-1, blaOXA-1 | aac(6′)-Ib-cr, aac(3)-Iia, OqxA, OqxB, fosA, tet(A) | ST-48 | JALHBC000000000 |
| K. pneumoniae 2 | 0.38 | 1.5 | + | - | blaCTX-M-15, blaSHV-81, blaTEM-1-B | sul1, sul2, oqxA, dfrA17, OqxA, OqxB, qacE, aadA5, tet(A), fosA | ST-37 | JALHBA000000000 |
| K. pneumoniae 3 | 6 | 6 | + | Faint NDM band | blaNDM-1, blaCTX-M-15, blaSHV-106 | aac(6′)-Ib-cr, msr(E), sul1, OqxA, dfrA12, OqxB, fosA, aadA2, mph(E) | ST-15 | JALHBB000000000 |
| K. variicola | 1.5 | 2 | + | Faint NDM band | blaNDM-1, blaSHV-12, blaLEN-9, blaOXA-10, blaOXA-1 | aac(6′)-Ib-cr, rmtC, rmtH, erm(B), aph(3′)-Ia, sul1, sul2, OqxB, OqxA, dfrA14, floR, qacE, fosA, qacE, tet(B), aadA1, ARR-2, catB3, cmlA1, catA2 | ST-3195 | JALHAY000000000 |
| P. aeruginosa 1 | >32 | >32 | + | - | blaOXA-50-like (T16A, K112E), blaPDC-11 | sul1, crpP, fosA, catB7, aph(6)-Id, aph(3″)-Ib | ST-357 | JALHAX000000000 |
| P. aeruginosa 2 | >32 | >32 | + | - | blaOXA-50-like (R49C, A133G, A181T), blaPDC-45 | aph(3′)-IIb, fosA, catB7 | ST-893 | JALHAW000000000 |
| P. aeruginosa 3 | >32 | >32 | + | - | blaOXA-50-like (R167H, D109E), blaPDC-1-like | Sul1, fosA, aph(3′)-IIb, crpP, catB7 | ST-277 | Pending |
| A. baumannii 1 | >32 | >32 | + | - | blaOXA-72, blaOXA-64, blaADC-26 | aac(6′)-lan, aph(6)-Id, aph(3″)-lb, aac(3)-Iia, sul2, aac(3)-Iia, tet(B) | ST-229 * | JALHAZ000000000 |
| A. baumannii 2 | >32 | >32 | + | - | blaOXA-23, blaOXA-66, blaTEM-1D, blaADC-73 | armA, msr(E), aph(3′)-Ia, sul2, tet(B), aph(6)-Id | ST-1841-like * | Pending |
| Species | N | Hospital a | Type of Infection | Year |
|---|---|---|---|---|
| Klebsiella pneumoniae | 11 | Nini (7) YHC (4) |
| 2015–2018 |
| Klebsiella variicola | 1 | YHC (1) |
| 2018 |
| Enterobacter cloacae | 5 | Nini (3) TGH (2) |
| 2017–2018 |
| Enterobacter hormaechei | 2 | Nini (1) TGH (1) |
| 2017 |
| Citrobacter freundii | 2 | YHC (2) |
| 2017 |
| Escherichia coli | 51 | Nini (22) YHC (25) TGH (4) |
| 2016–2018 |
| Pseudomonas aeruginosa | 29 | YHC (25) TGH (4) |
| 2016–2018 |
| Acinetobacter baumannii | 45 | Nini (8) YHC (22) TGH (15) |
| 2016–2019 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rima, M.; Oueslati, S.; Dabos, L.; Daaboul, D.; Mallat, H.; Bou Raad, E.; Achkar, M.; Mawlawi, O.; Bernabeu, S.; Bonnin, R.A.; et al. Prevalence and Molecular Mechanisms of Carbapenem Resistance among Gram-Negative Bacilli in Three Hospitals of Northern Lebanon. Antibiotics 2022, 11, 1295. https://doi.org/10.3390/antibiotics11101295
Rima M, Oueslati S, Dabos L, Daaboul D, Mallat H, Bou Raad E, Achkar M, Mawlawi O, Bernabeu S, Bonnin RA, et al. Prevalence and Molecular Mechanisms of Carbapenem Resistance among Gram-Negative Bacilli in Three Hospitals of Northern Lebanon. Antibiotics. 2022; 11(10):1295. https://doi.org/10.3390/antibiotics11101295
Chicago/Turabian StyleRima, Mariam, Saoussen Oueslati, Laura Dabos, Dina Daaboul, Hassan Mallat, Elie Bou Raad, Marcel Achkar, Osman Mawlawi, Sandrine Bernabeu, Rémy A. Bonnin, and et al. 2022. "Prevalence and Molecular Mechanisms of Carbapenem Resistance among Gram-Negative Bacilli in Three Hospitals of Northern Lebanon" Antibiotics 11, no. 10: 1295. https://doi.org/10.3390/antibiotics11101295