
Hydroxy-Selenomethionine, an Organic Selenium Source, Increases Selenoprotein Expression and Positively Modulates the Inflammatory Response of LPS-Stimulated Macrophages
 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. THP-1 Cell Culture
2.3. Salmonella Enteritidis Culture
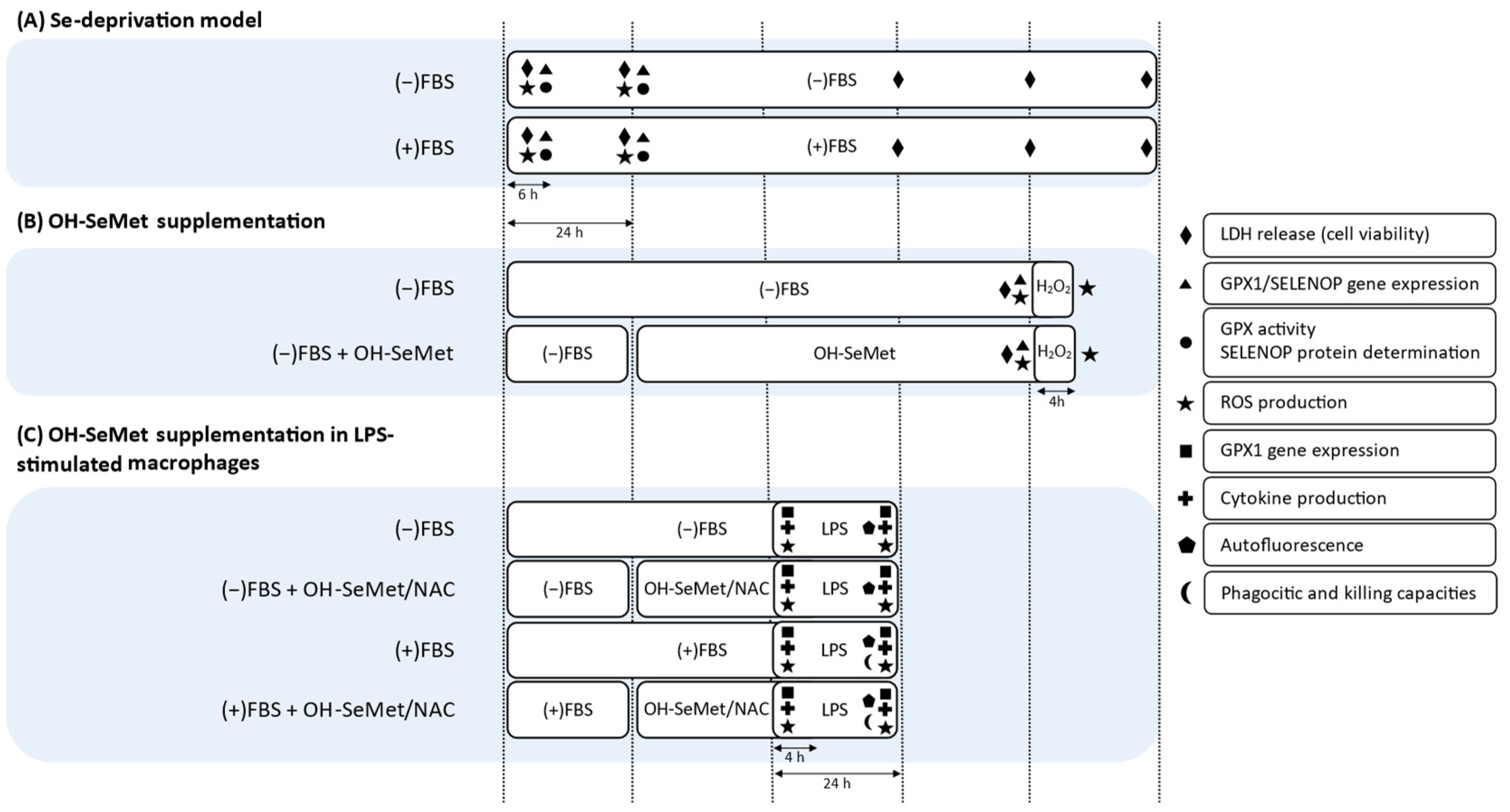
2.4. Se Deprivation and OH-SeMet Supplementation
2.5. OH-SeMet Supplementation in LPS-Stimulated Macrophages
2.6. Statistical Analysis
3. Results
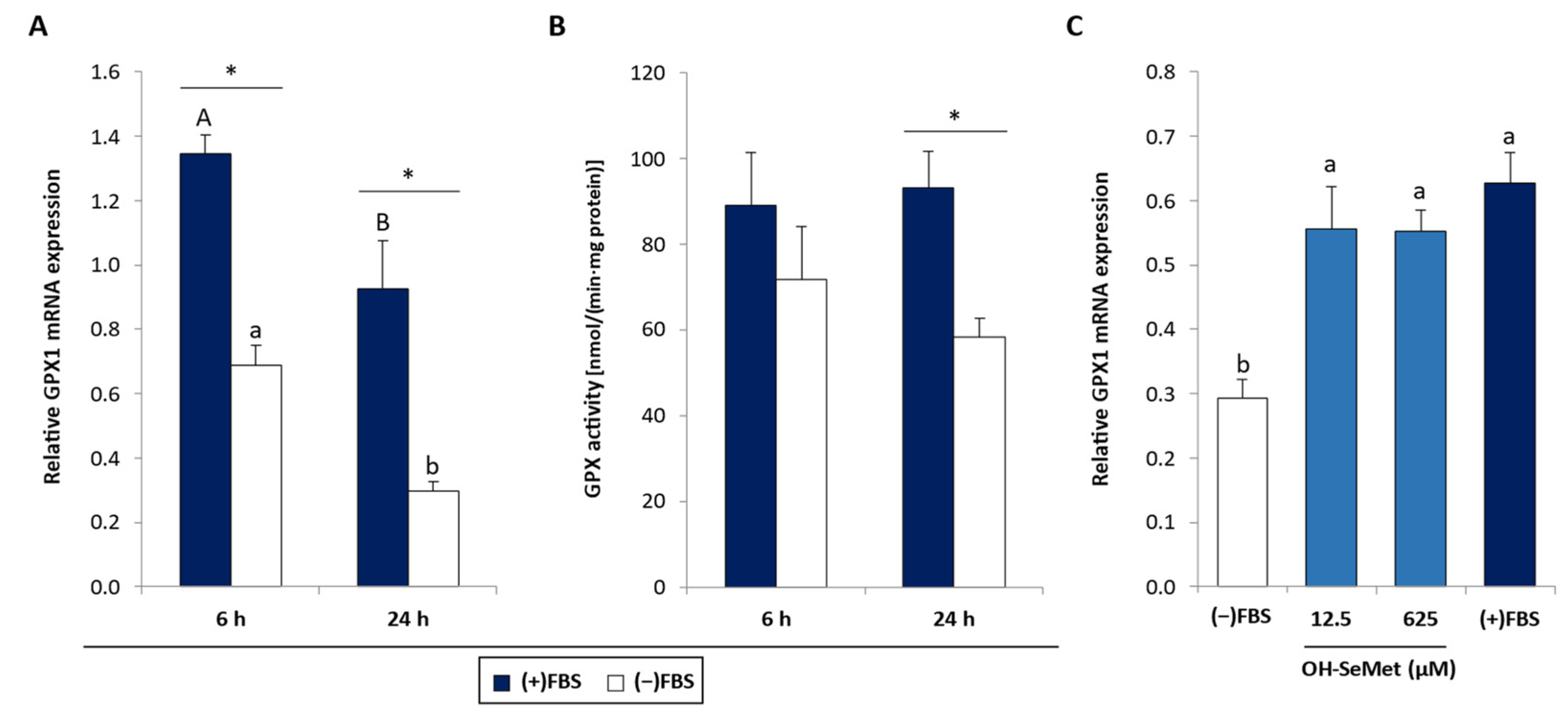
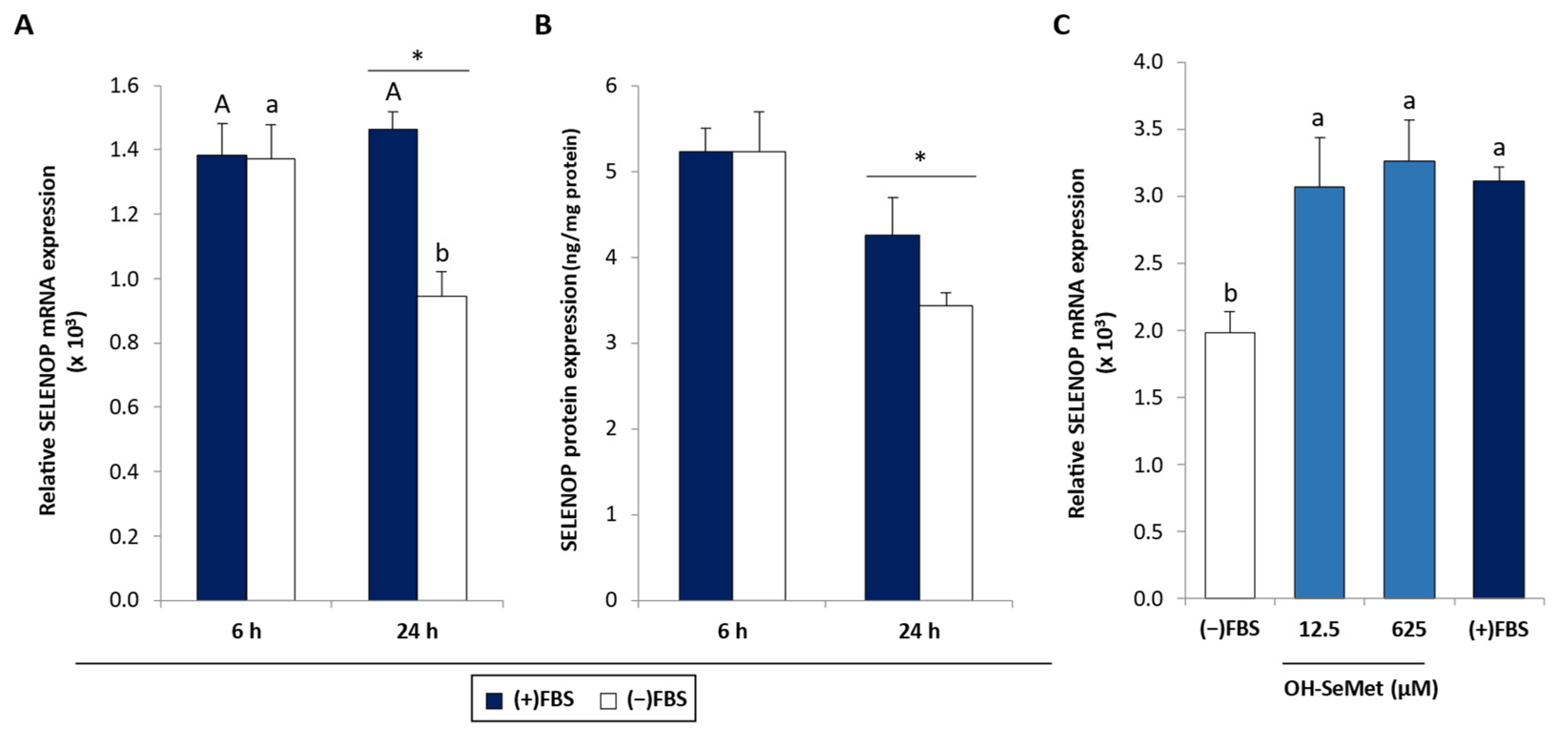
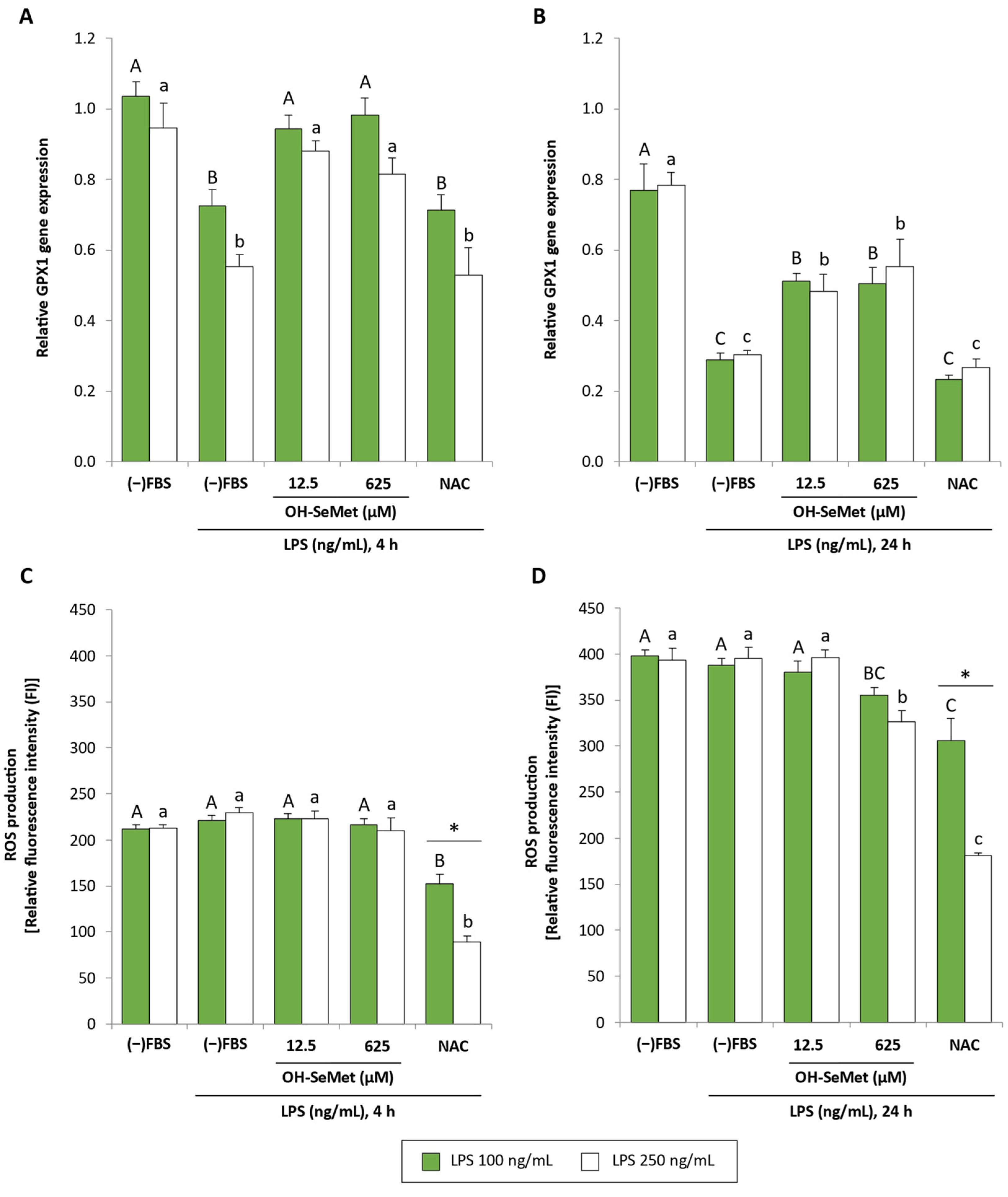
3.1. Se Deprivation and OH-SeMet Supplementation
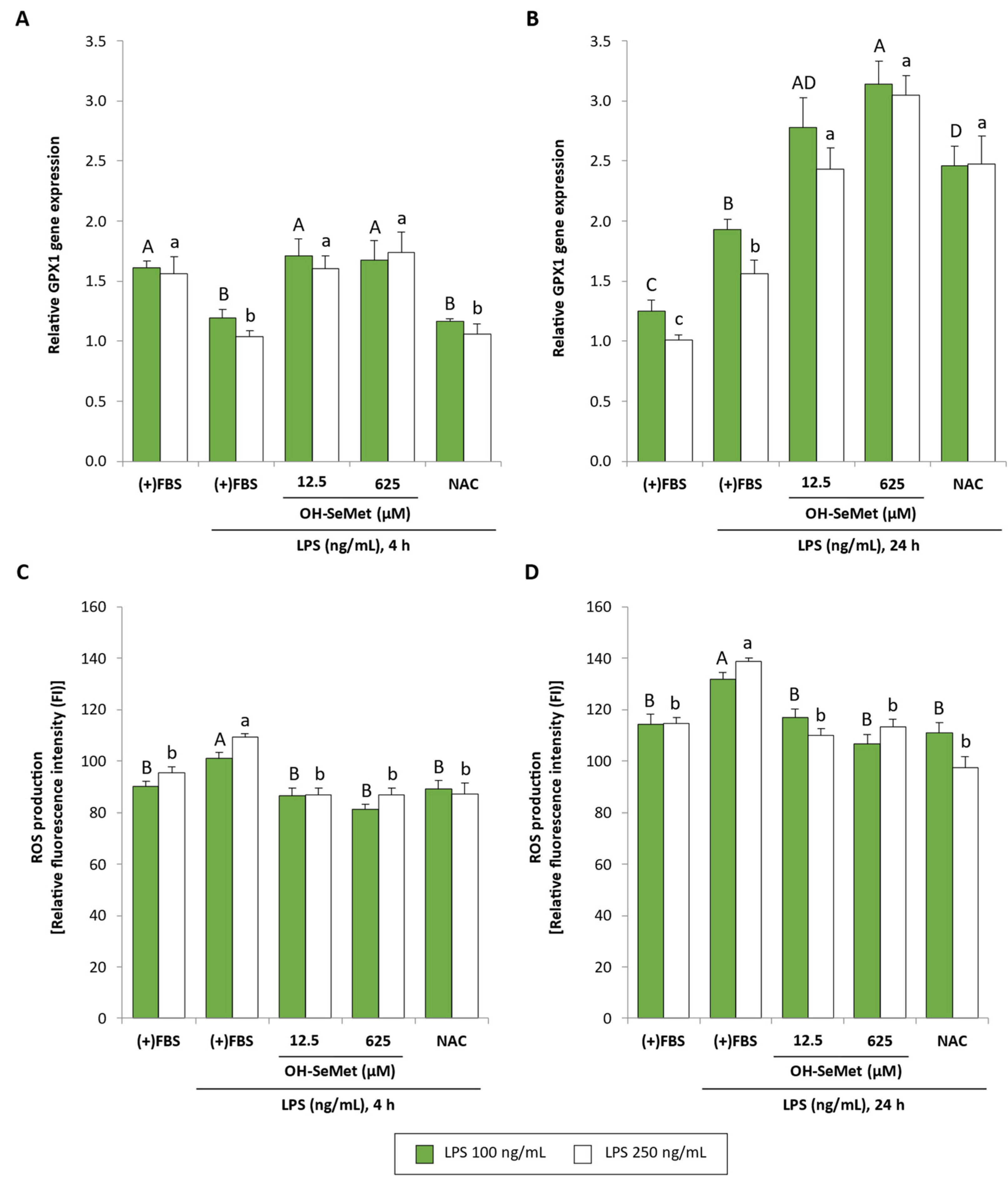
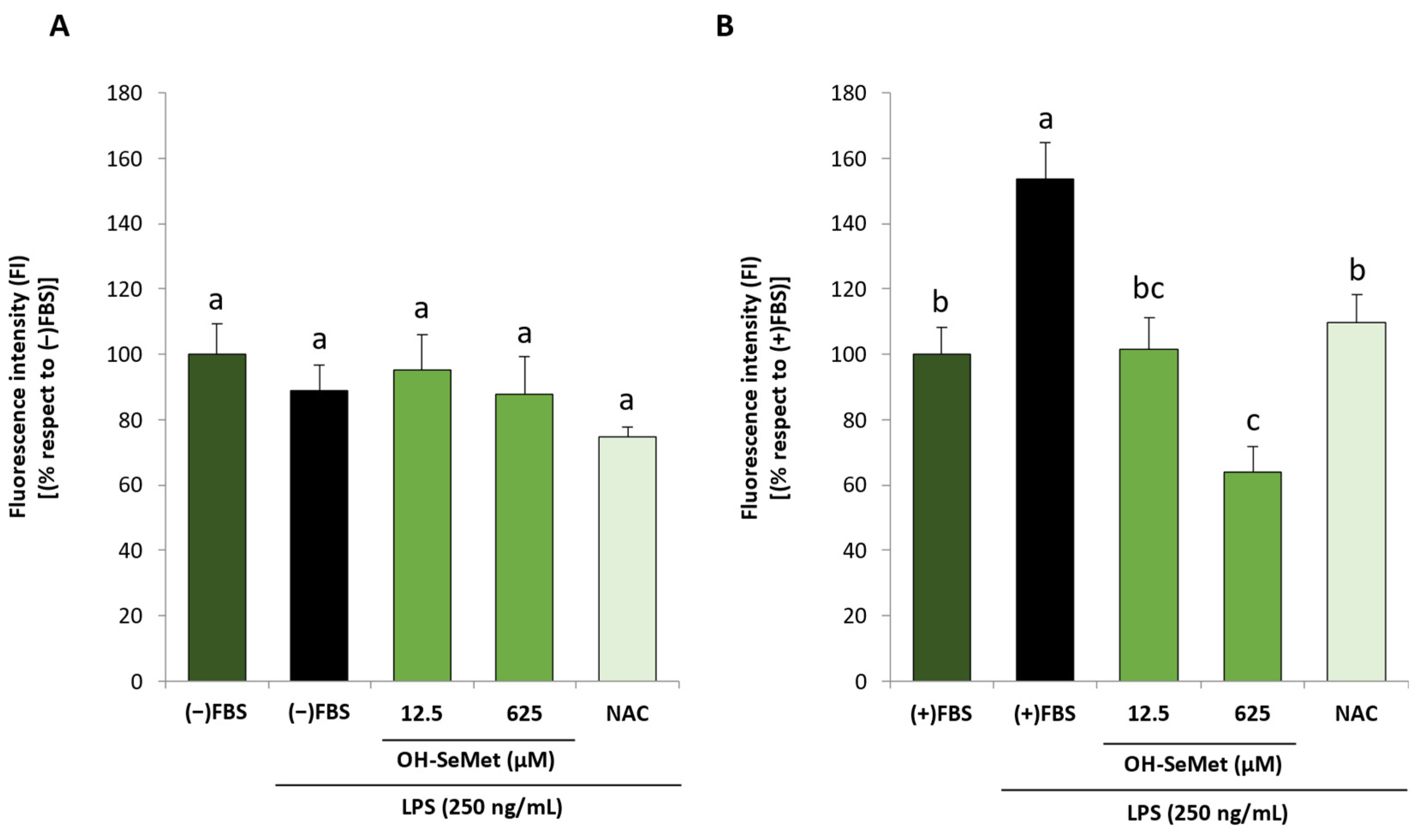
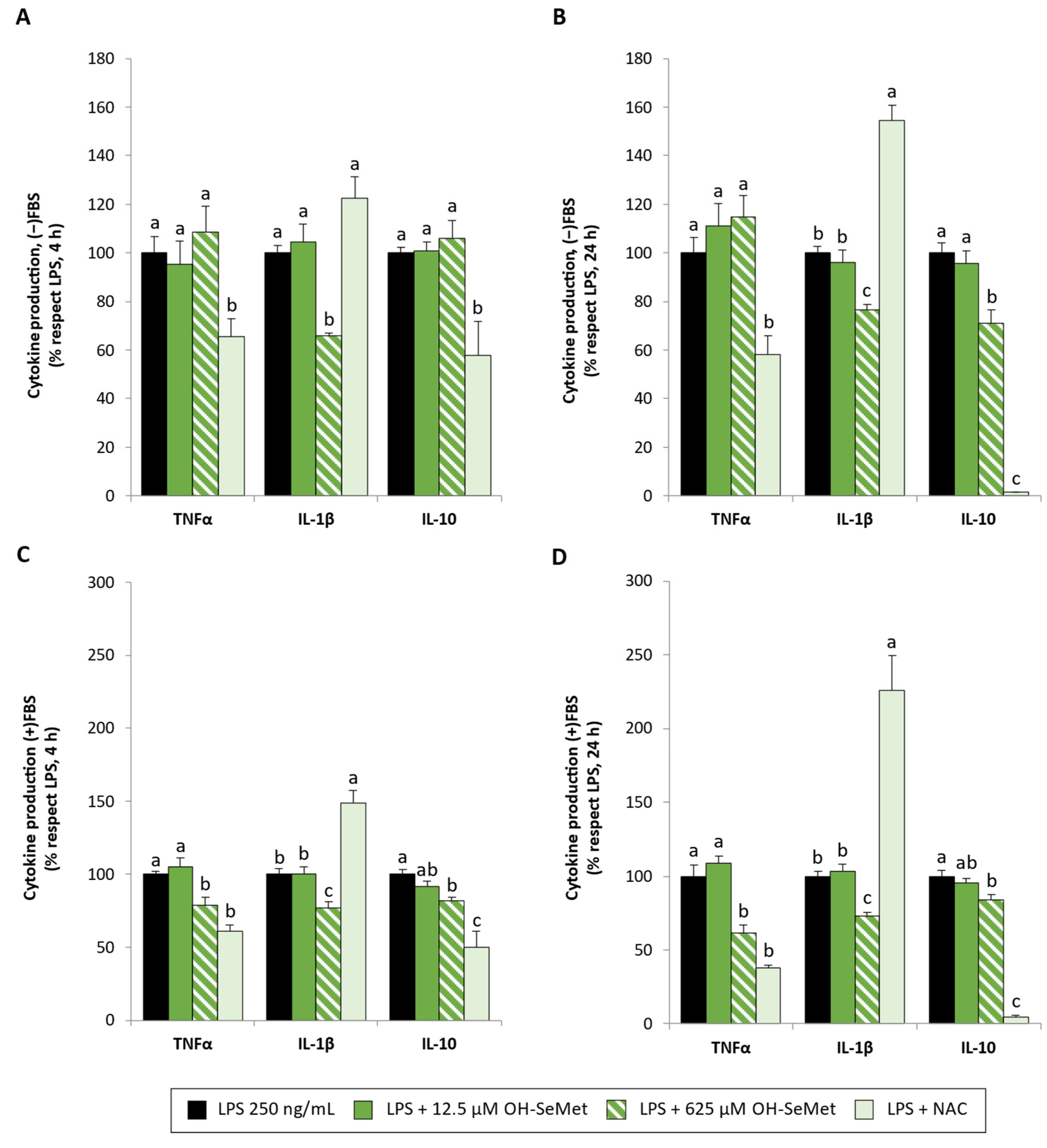
3.2. OH-SeMet Supplementation of LPS-Stimulated Macrophages
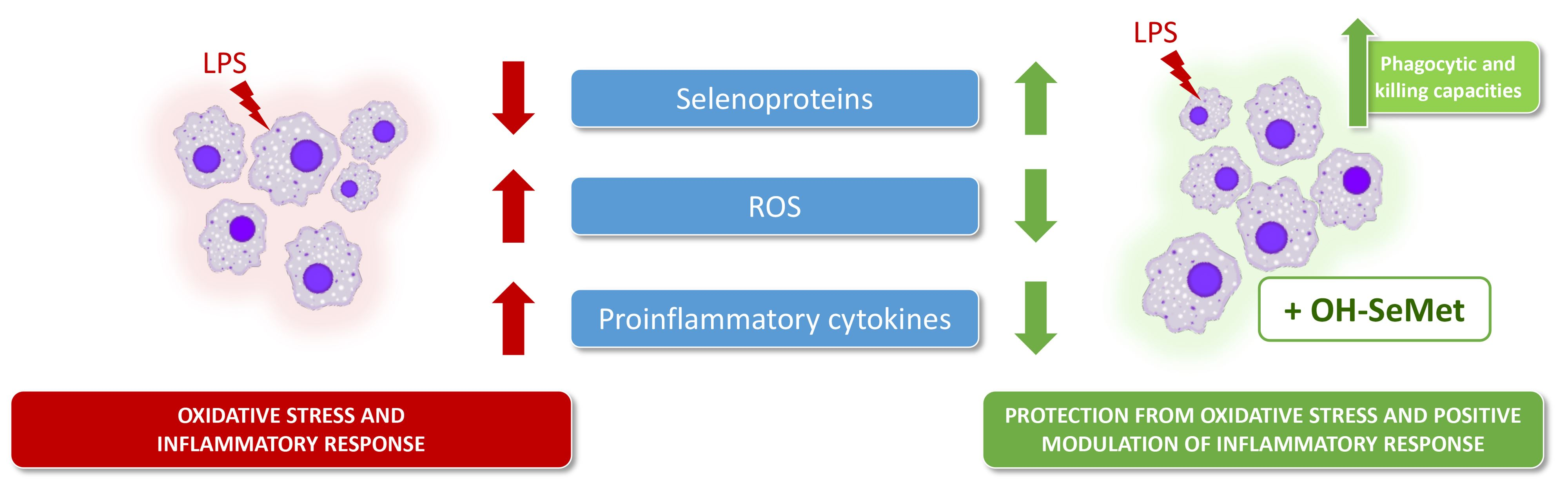
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vunta, H.; Belda, B.J.; Arner, R.J.; Reddy, C.C.; Vanden Heuvel, J.P.; Prabhu, K.S. Selenium attenuates pro-inflammatory gene expression in macrophages. Mol. Nutr. Food Res. 2008, 52, 1316–1323. [Google Scholar] [CrossRef] [PubMed]
- Carlson, B.A.; Yoo, M.H.; Sano, Y.; Sengupta, A.; Kim, J.Y.; Irons, R.; Gladyshev, V.N.; Hatfield, D.L.; Park, J.M. Selenoproteins regulate macrophage invasiveness and extracellular matrix-related gene expression. BMC Immunol. 2009, 10, 57. [Google Scholar] [CrossRef] [PubMed]
- Dhanjal, N.I.K.; Sharma, S.; Prabhu, K.S.; Prakash, N.T. Selenium supplementation through Se-rich dietary matrices can upregulate the anti-inflammatory responses in lipopolysaccharide-stimulated murine macrophages. Food Agric. Immunol. 2017, 28, 1374–1392. [Google Scholar] [CrossRef] [PubMed]
- Vunta, H.; Davis, F.; Palempalli, U.D.; Bhat, D.; Arner, R.J.; Thompson, J.T.; Peterson, D.G.; Reddy, C.C.; Prabhu, K.S. The anti-inflammatory effects of selenium are mediated through 15-deoxy-Δ12,14-prostaglandin J2 in macrophages. J. Biol. Chem. 2007, 2821, 7964–17973. [Google Scholar] [CrossRef]
- Huang, Z.; Rose, A.H.; Hoffmann, P.R. The role of selenium in inflammation and immunity: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2012, 16, 705–743. [Google Scholar] [CrossRef]
- Barrett, C.W.; Short, S.P.; Williams, C.S. Selenoproteins and oxidative stress-induced inflammatory tumorigenesis in the gut. Cell. Mol. Life Sci. 2017, 74, 607–616. [Google Scholar] [CrossRef]
- Avery, J.C.; Hoffmann, P.R. Selenium, Selenoproteins, and Immunity. Nutrients 2018, 10, 1203. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Velichko, O.A. Selenium in Poultry Nutrition: From Sodium Selenite to Organic Selenium Sources. J. Poult. Sci. 2018, 55, 79–93. [Google Scholar] [CrossRef]
- Campo-Sabariz, J.; Moral-Anter, D.; Brufau, M.T.; Briens, M.; Pinloche, E.; Ferrer, R.; Martín-Venegas, R. 2-Hydroxy-(4-methylseleno)butanoic Acid Is Used by Intestinal Caco-2 Cells as a Source of Selenium and Protects against Oxidative Stress. J. Nutr. 2019, 149, 2191–2198. [Google Scholar] [CrossRef]
- Palacio, J.R.; Markert, U.R.; Martínez, P. Anti-inflammatory properties of N-acetylcysteine on lipopolysaccharide-activated macrophages. Inflamm. Res. 2011, 60, 695–704. [Google Scholar] [CrossRef]
- Brufau, M.T.; Campo-Sabariz, J.; Carné, S.; Ferrer, R.; Martín-Venegas, R. Salmosan, a β-galactomannan-rich product, in combination with Lactobacillus plantarum contributes to restore intestinal epithelial barrier function by modulation of cytokine production. J. Nutr. Biochem. 2017, 41, 20–24. [Google Scholar] [CrossRef]
- Brufau, M.T.; Campo-Sabariz, J.; Bou, R.; Carné, S.; Brufau, J.; Vilà, B.; Marqués, A.M.; Guardiola, F.; Ferrer, R.; Martín-Venegas, R. Salmosan, a β-Galactomannan-Rich Product, Protects Epithelial Barrier Function in Caco-2 Cells Infected by Salmonella enterica Serovar Enteritidis. J. Nutr. 2016, 146, 1492–1498. [Google Scholar] [CrossRef]
- Parmentier, M.; Hirani, N.; Rahman, I.; Donaldson, K.; MacNee, W.; Antonicelli, F. Regulation of lipopolysaccharide-mediated interleukin-1β release by N-acetylcysteine in THP-1 cells. Eur. Respir. J. 2000, 16, 933–939. [Google Scholar] [CrossRef]
- Schrauzer, G.N. The nutritional significance, metabolism and toxicology of selenomethionine. Adv. Food Nutr. Res. 2003, 47, 73–112. [Google Scholar] [CrossRef]
- Roman, M.; Jitaru, P.; Barbante, C. Selenium biochemistry and its role for human health. Metallomics 2014, 6, 25–54. [Google Scholar] [CrossRef]
- Varlamova, E.G.; Turovsky, E.A.; Blinova, E.V. Therapeutic Potential and Main Methods of Obtaining Selenium Nanoparticles. Int. J. Mol. Sci. 2021, 22, 10808. [Google Scholar] [CrossRef]
- Heller, R.A.; Sun, Q.; Hackler, J.; Seelig, J.; Seibert, L.; Cherkezov, A.; Minich, W.B.; Seemann, P.; Diegmann, J.; Pilz, M.; et al. Prediction of survival odds in COVID-19 by zinc, age and selenoprotein P as composite biomarker. Redox. Biol. 2021, 38, 101764. [Google Scholar] [CrossRef]
- Mal’tseva, V.N.; Goltyaev, M.V.; Turovsky, E.A.; Varlamova, E.G. Immunomodulatory and Anti-Inflammatory Properties of Selenium-Containing Agents: Their Role in the Regulation of Defense Mechanisms against COVID-19. Int. J. Mol. Sci. 2022, 23, 2360. [Google Scholar] [CrossRef]
- Prabhu, K.S.; Zamamiri-Davis, F.; Stewart, J.B.; Thompson, J.T.; Sordillo, L.M.; Reddy, C.C. Selenium deficiency increases the expression of inducible nitric oxide synthase in RAW 264.7 macrophages: Role of nuclear factor-kappaB in up-regulation. Biochem. J. 2002, 366, 203–209. [Google Scholar] [CrossRef]
- Schoenmakers, E.; Carlson, B.; Agostini, M.; Moran, C.; Rajanayagam, O.; Bochukova, E.; Tobe, R.; Peat, R.; Gevers, E.; Muntoni, F.; et al. Mutation in human selenocysteine transfer RNA selectively disrupts selenoprotein synthesis. J. Clin. Investig. 2016, 126, 992–996. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Gong, Y.; Sun, Y.; Cai, J.; Liu, Q.; Bao, J.; Yang, J.; Zhang, Z. Impact of Selenium Deficiency on Inflammation, Oxidative Stress, and Phagocytosis in Mouse Macrophages. Biol. Trace Elem. Res. 2020, 194, 237–243. [Google Scholar] [CrossRef]
- Briens, M.; Mercier, Y.; Rouffineau, F.; Vacchina, V.; Geraert, P.A. Comparative study of a new organic selenium source v. seleno-yeast and mineral selenium sources on muscle selenium enrichment and selenium digestibility in broiler chickens. Br. J. Nutr. 2013, 110, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Briens, M.; Mercier, Y.; Rouffineau, F.; Mercerand, F.; Geraert, P.A. 2-Hydroxy-4-methylselenobutanoic acid induces additional tissue selenium enrichment in broiler chickens compared with other selenium sources. Poult. Sci. 2014, 93, 85–93. [Google Scholar] [CrossRef]
- Sun, H.; Zhao, L.; Xu, Z.J.; De Marco, M.; Briens, M.; Yan, X.H.; Sun, L.H. Hydroxy-Selenomethionine Improves the Selenium Status and Helps to Maintain Broiler Performances under a High Stocking Density and Heat Stress Conditions through a Better Redox and Immune Response. Antioxidants 2021, 10, 1542. [Google Scholar] [CrossRef]
- Jlali, M.; Briens, M.; Rouffineau, F.; Mercerand, F.; Geraert, P.A.; Mercier, Y. Effect of 2-hydroxy-4-methylselenobutanoic acid as a dietary selenium supplement to improve the selenium concentration of table eggs. J. Anim. Sci. 2013, 91, 1745–1752. [Google Scholar] [CrossRef] [PubMed]
- Jlali, M.; Briens, M.; Rouffineau, F.; Geraert, P.A.; Mercier, Y. Evaluation of the efficacy of 2-hydroxy-4-methylselenobutanoic acid on growth performance and tissue selenium retention in growing pigs. J. Anim. Sci. 2014, 92, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Li, N.Y.; Sun, Z.J.; Ansari, A.R.; Cui, L.; Hu, Y.F.; Li, Z.W.; Briens, M.; Kai, L.; Sun, L.H.; Karrow, N.A.; et al. Impact of Maternal Selenium Supplementation from Late Gestation and Lactation on Piglet Immune Function. Biol. Trace Elem. Res. 2020, 194, 159–167. [Google Scholar] [CrossRef]
- Juniper, D.T.; Rymer, C.; Briens, M. Bioefficacy of hydroxy-selenomethionine as a selenium supplement in pregnant dairy heifers and on the selenium status of their calves. J. Dairy Sci. 2019, 102, 7000–7010. [Google Scholar] [CrossRef]
- Grossi, S.; Rossi, L.; De Marco, M.; Sgoifo Rossi, C.A. The Effect of Different Sources of Selenium Supplementation on the Meat Quality Traits of Young Charolaise Bulls during the Finishing Phase. Antioxidants 2021, 10, 596. [Google Scholar] [CrossRef]
- Sun, P.; Wang, J.; Liu, W.; Bu, D.P.; Liu, S.J.; Zhang, K.Z. Hydroxy-selenomethionine: A novel organic selenium source that improves antioxidant status and selenium concentrations in milk and plasma of mid-lactation dairy cows. J. Dairy Sci. 2017, 100, 9602–9610. [Google Scholar] [CrossRef] [Green Version]
- Maseko, T.; Howell, K.; Dunshea, F.R.; Ng, K. Selenium-enriched Agaricus bisporus increases expression and activity of glutathione peroxidase-1 and expression of glutathione peroxidase-2 in rat colon. Food Chem. 2014, 146, 327–333. [Google Scholar] [CrossRef]
- Song, E.; Su, C.; Fu, J.; Xia, X.; Yang, S.; Xiao, C.; Lu, B.; Chen, H.; Sun, Z.; Wu, S.; et al. Selenium supplementation shows protective effects against patulin-induced brain damage in mice via increases in GSH-related enzyme activity and expression. Life Sci. 2014, 109, 37–43. [Google Scholar] [CrossRef]
- Zhao, L.; Sun, L.H.; Huang, J.Q.; Briens, M.; Qi, D.S.; Xu, S.W.; Lei, X.G. A Novel Organic Selenium Compound Exerts Unique Regulation of Selenium Speciation, Selenogenome, and Selenoproteins in Broiler Chicks. J. Nutr. 2017, 147, 789–797. [Google Scholar] [CrossRef]
- Juniper, D.T.; Kliem, K.E.; Lee, A.; Rymer, C. The effect of stocking rate and supplementary selenium on the fatty acid composition and subsequent peroxidisability of poultry muscle tissues. Animal 2022, 16, 100459. [Google Scholar] [CrossRef]
- Bosschaerts, T.; Guilliams, M.; Noel, W.; Hérin, M.; Burk, R.F.; Hill, K.E.; Brys, L.; Raes, G.; Ghassabeh, G.H.; De Baetselier, P.; et al. Alternatively activated myeloid cells limit pathogenicity associated with African trypanosomiasis through the IL-10 inducible gene selenoprotein P. J. Immunol. 2008, 180, 6168–6175. [Google Scholar] [CrossRef]
- Short, S.P.; Pilat, J.M.; Williams, C.S. Roles for selenium and selenoprotein P in the development, progression, and prevention of intestinal disease. Free Radic. Biol. Med. 2018, 127, 26–35. [Google Scholar] [CrossRef]
- Ding, D.; Mou, D.; Zhao, L.; Jiang, X.; Che, L.; Fang, Z.; Xu, S.; Lin, Y.; Zhuo, Y.; Li, J.; et al. Maternal organic selenium supplementation alleviates LPS induced inflammation, autophagy and ER stress in the thymus and spleen of offspring piglets by improving the expression of selenoproteins. Food Funct. 2021, 12, 11214–11228. [Google Scholar] [CrossRef]
- Bansal, M.P.; Kaur, T. Growth characteristics and selenium status changes of yeast cells with inorganic and organic selenium supplementation: Selenium, a chemopreventive agent. J. Med. Food. 2002, 5, 85–90. [Google Scholar] [CrossRef]
- Sahin, N.; Onderci, M.; Sahin, K.; Kucuk, O. Supplementation with organic or inorganic selenium in heat-distressed quail. Biol. Trace Elem. Res. 2008, 122, 229–237. [Google Scholar] [CrossRef]
- Barrera, L.N.; Cassidy, A.; Wang, W.; Wei, T.; Belshaw, N.J.; Johnson, I.T.; Brigelius-Flohé, R.; Bao, Y. TrxR1 and GPx2 are potently induced by isothiocyanates and selenium, and mutually cooperate to protect Caco-2 cells against free radical-mediated cell death. Biochim. Biophys. Acta 2012, 1823, 1914–1924. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Wang, F.; Wu, Z.; Ma, L.; Baumrucker, C.; Bu, D. Comparison of Selenium Source in Preventing Oxidative Stress in Bovine Mammary Epithelial Cells. Animals 2020, 10, 842. [Google Scholar] [CrossRef] [PubMed]
- Fisinin, V.I.; Papazyan, T.T.; Surai, P.F. Producing selenium-enriched eggs and meat to improve the selenium status of the general population. Crit. Rev. Biotechnol. 2009, 29, 18–28. [Google Scholar] [CrossRef]
- Wang, L.; Jing, J.; Yan, H.; Tang, J.; Jia, G.; Liu, G.; Chen, X.; Tian, G.; Cai, J.; Shang, H.; et al. Selenium Pretreatment Alleviated LPS-Induced Immunological Stress Via Upregulation of Several Selenoprotein Encoding Genes in Murine RAW264.7 Cells. Biol. Trace Elem. Res. 2018, 186, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Krifka, S.; Hiller, K.A.; Spagnuolo, G.; Jewett, A.; Schmalz, G.; Schweikl, H. The influence of glutathione on redox regulation by antioxidant proteins and apoptosis in macrophages exposed to 2-hydroxyethyl methacrylate (HEMA). Biomaterials 2012, 33, 5177–5186. [Google Scholar] [CrossRef] [PubMed]
- Safir, N.; Wendel, A.; Saile, R.; Chabraoui, L. The effect of selenium on immune functions of J774.1 cells. Clin. Chem. Lab. Med. 2003, 41, 1005–1011. [Google Scholar] [CrossRef] [PubMed]
- Gerberick, G.F.; Willoughby, J.B.; Willoughby, W.F. Serum factor requirement for reactive oxygen intermediate release by rabbit alveolar macrophages. J. Exp. Med. 1985, 161, 392–408. [Google Scholar] [CrossRef] [PubMed]
- Jian, Z.J.; Yang, Z.; Mason, G.L.; Slauson, D.O.; Bochsler, P.N. Regulation of superoxide anion generation in bovine alveolar macrophages by bacterial lipopolysaccharide, serum proteins, and modulators of signal transduction. Inflammation 1995, 19, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Guha, M.; Mackman, N. LPS induction of gene expression in human monocytes. Cell. Signal. 2001, 13, 85–94. [Google Scholar] [CrossRef]
- Verstovsek, S.; Zaleskis, G.; Maccubbin, D.L.; Mihich, E.; Ehrke, M.J. Lipopolysaccharide and splenic tumoricidal macrophage activation. J. Leukoc. Biol. 1994, 56, 714–722. [Google Scholar] [CrossRef]
- Sköld, C.M.; Eklund, A.; Halldén, G.; Hed, J. Autofluorescence in human alveolar macrophages from smokers: Relation to cell surface markers and phagocytosis. Exp. Lung Res. 1989, 15, 823–835. [Google Scholar] [CrossRef]
- Pankow, W.; Neumann, K.; Rüschoff, J.; von Wichert, P. Human alveolar macrophages: Comparison of cell size, autofluorescence, and HLA-DR antigen expression in smokers and nonsmokers. Cancer Detect. Prev. 1995, 19, 268–273. [Google Scholar]
- Edelson, J.D.; MacFadden, D.K.; Klein, M.; Rebuck, A.S. Autofluorescence of alveolar macrophages: Problems and potential solutions. Med. Hypotheses. 1985, 17, 403–407. [Google Scholar] [CrossRef]
- Njoroge, J.M.; Mitchell, L.B.; Centola, M.; Kastner, D.; Raffeld, M.; Miller, J.L. Characterization of viable autofluorescent macrophages among cultured peripheral blood mononuclear cells. Cytometry 2001, 44, 38–44. [Google Scholar] [CrossRef]
- Wolfram, T.; Weidenbach, L.M.; Adolf, J.; Schwarz, M.; Schädel, P.; Gollowitzer, A.; Werz, O.; Koeberle, A.; Kipp, A.P.; Koeberle, S.C. The Trace Element Selenium Is Important for Redox Signaling in Phorbol Ester-Differentiated THP-1 Macrophages. Int. J. Mol. Sci. 2021, 22, 11060. [Google Scholar] [CrossRef]
- Kim, S.H.; Johnson, V.J.; Shin, T.Y.; Sharma, R.P. Selenium attenuates lipopolysaccharide-induced oxidative stress responses through modulation of p38 MAPK and NF-kappaB signaling pathways. Exp. Biol. Med. 2004, 229, 203–213. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, R.; Wang, T.; Jiang, H.; Guo, M.; Zhou, E.; Sun, Y.; Yang, Z.; Xu, S.; Cao, Y.; et al. Selenium inhibits LPS-induced pro-inflammatory gene expression by modulating MAPK and NF-κB signaling pathways in mouse mammary epithelial cells in primary culture. Inflammation 2014, 37, 478–485. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I.; Kidd, M.T. Redox Homeostasis in Poultry: Regulatory Roles of NF-κB. Antioxidants 2021, 10, 186. [Google Scholar] [CrossRef]
- Ryan, K.A.; Smith, M.F., Jr.; Sanders, M.K.; Ernst, P.B. Reactive oxygen and nitrogen species differentially regulate Toll-like receptor 4-mediated activation of NF-kappa B and interleukin-8 expression. Infect. Immun. 2004, 72, 2123–2130. [Google Scholar] [CrossRef]
- Al-Shukaili, A.; Al-Abri, S.; Al-Ansari, A.; Monteil, M.A. Effect of N-acetyl-L-cysteine on Cytokine Production by Human Peripheral Blood Mononuclear Cells. Sultan Qaboos Univ. Med. J. 2009, 9, 70–74. [Google Scholar]
- Ipseiz, N.; Pickering, R.J.; Rosas, M.; Tyrrell, V.J.; Davies, L.C.; Orr, S.J.; Czubala, M.A.; Fathalla, D.; Robertson, A.A.; Bryant, C.E.; et al. Tissue-resident macrophages actively suppress IL-1beta release via a reactive prostanoid/IL-10 pathway. EMBO J. 2020, 39, e103454. [Google Scholar] [CrossRef]
- Nelson, S.M.; Lei, X.; Prabhu, K.S. Selenium levels affect the IL-4-induced expression of alternative activation markers in murine macrophages. J. Nutr. 2011, 141, 1754–1761. [Google Scholar] [CrossRef]
- Li, Z.; Dong, Y.; Chen, S.; Jia, X.; Jiang, X.; Che, L.; Lin, Y.; Li, J.; Feng, B.; Fang, Z.; et al. Organic Selenium Increased Gilts Antioxidant Capacity, Immune Function, and Changed Intestinal Microbiota. Front. Microbiol. 2021, 12, 723190. [Google Scholar] [CrossRef]
- Mou, D.; Ding, D.; Yan, H.; Qin, B.; Dong, Y.; Li, Z.; Che, L.; Fang, Z.; Xu, S.; Lin, Y.; et al. Maternal supplementation of organic selenium during gestation improves sows and offspring antioxidant capacity and inflammatory status and promotes embryo survival. Food Funct. 2020, 11, 7748–7761. [Google Scholar] [CrossRef]
- Mou, D.; Ding, D.; Yang, M.; Jiang, X.; Zhao, L.; Che, L.; Fang, Z.; Xu, S.; Lin, Y.; Zhuo, Y.; et al. Maternal organic selenium supplementation during gestation improves the antioxidant capacity and reduces the inflammation level in the intestine of offspring through the NF-κB and ERK/Beclin-1 pathways. Food Funct. 2021, 12, 315–327. [Google Scholar] [CrossRef]
- Joshi, L.; Ponnana, M.; Sivangala, R.; Chelluri, L.K.; Nallari, P.; Penmetsa, S.; Valluri, V.; Gaddam, S. Evaluation of TNF-α, IL-10 and IL-6 Cytokine Production and Their Correlation with Genotype Variants amongst Tuberculosis Patients and Their Household Contacts. PLoS ONE 2015, 10, e0137727. [Google Scholar] [CrossRef]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef]
- Suwanpanya, N.; Wongpratoom, W.; Wanapat, M.; Aiumlamai, S.; Wittayakun, S.; Wachirapakorn, C. The influence of bovine neutrophils on in vitro phagocytosis and killing of Staphylococcus aureus in heifers supplemented with selenium and vitamin E. J. Sci. Technol. 2007, 29, 697–706. [Google Scholar]
- Aribi, M.; Meziane, W.; Habi, S.; Boulatika, Y.; Marchandin, H.; Aymeric, J.L. Macrophage Bactericidal Activities against Staphylococcus aureus Are Enhanced In Vivo by Selenium Supplementation in a Dose-Dependent Manner. PLoS ONE 2015, 10, e0135515. [Google Scholar] [CrossRef] [PubMed]
- Virág, L.; Jaén, R.I.; Regdon, Z.; Boscá, L.; Prieto, P. Self-defense of macrophages against oxidative injury: Fighting for their own survival. Redox Biol. 2019, 26, 101261. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (−)LPS | (+)LPS | |||
|---|---|---|---|---|
| Time | Cytokine (pg/µg) | (+/−)FBS | (−)FBS | (+)FBS |
| 4 h | TNFα | ND | 2.18 ± 0.159 | 15.21 ± 0.838 * |
| IL-1β | ND | 0.27 ± 0.021 | 0.43 ± 0.027 * | |
| IL-10 | ND | 0.02 ± 0.002 | 0.31 ± 0.023 * | |
| 24 h | TNFα | ND | 7.02 ± 0.564 | 13.08 ± 0.742 * |
| IL-1β | ND | 0.99 ± 0.122 | 0.96 ± 0.039 | |
| IL-10 | ND | 0.25 ± 0.014 | 1.07 ± 0.077 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campo-Sabariz, J.; García-Vara, A.; Moral-Anter, D.; Briens, M.; Hachemi, M.A.; Pinloche, E.; Ferrer, R.; Martín-Venegas, R. Hydroxy-Selenomethionine, an Organic Selenium Source, Increases Selenoprotein Expression and Positively Modulates the Inflammatory Response of LPS-Stimulated Macrophages. Antioxidants 2022, 11, 1876. https://doi.org/10.3390/antiox11101876
Campo-Sabariz J, García-Vara A, Moral-Anter D, Briens M, Hachemi MA, Pinloche E, Ferrer R, Martín-Venegas R. Hydroxy-Selenomethionine, an Organic Selenium Source, Increases Selenoprotein Expression and Positively Modulates the Inflammatory Response of LPS-Stimulated Macrophages. Antioxidants. 2022; 11(10):1876. https://doi.org/10.3390/antiox11101876
Chicago/Turabian StyleCampo-Sabariz, Joan, Adriana García-Vara, David Moral-Anter, Mickael Briens, Mohammed A. Hachemi, Eric Pinloche, Ruth Ferrer, and Raquel Martín-Venegas. 2022. "Hydroxy-Selenomethionine, an Organic Selenium Source, Increases Selenoprotein Expression and Positively Modulates the Inflammatory Response of LPS-Stimulated Macrophages" Antioxidants 11, no. 10: 1876. https://doi.org/10.3390/antiox11101876