
Heat-Killed Lacticaseibacillus paracasei Ameliorated UVB-Induced Oxidative Damage and Photoaging and Its Underlying Mechanisms


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. PL Preparation
2.2. UVB Irradiation
2.3. Cell Culture
2.4. MTT Assay
2.5. TUNEL Assay
2.6. DCFH-DA Staining Assay
2.7. Oxidative Damage and Antioxidant Assay
2.8. β-Gal Assay
2.9. ELISA Assay
2.10. Melanin Generation and Tyrosinase Activity Assays
2.11. qRT-PCR Assay
2.12. Western Blotting Assay
2.13. Statistical Analyses
3. Results
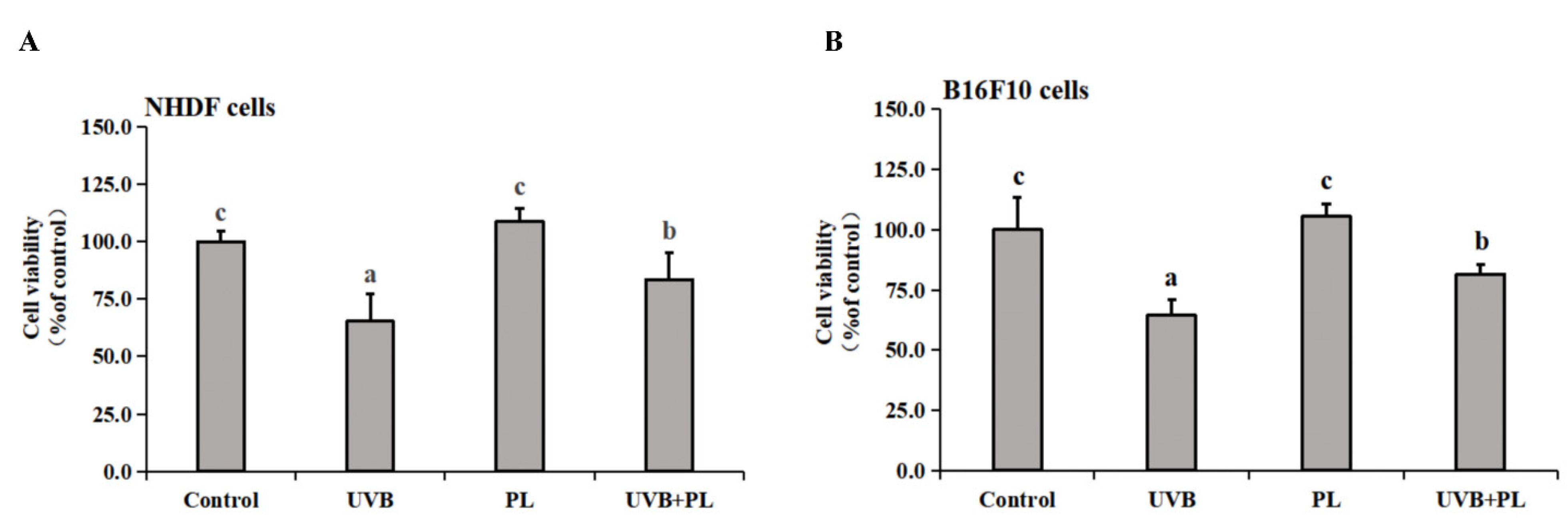
3.1. The Effects of PL on the Viability and DNA Damage of NHDF and B16F10 Cells
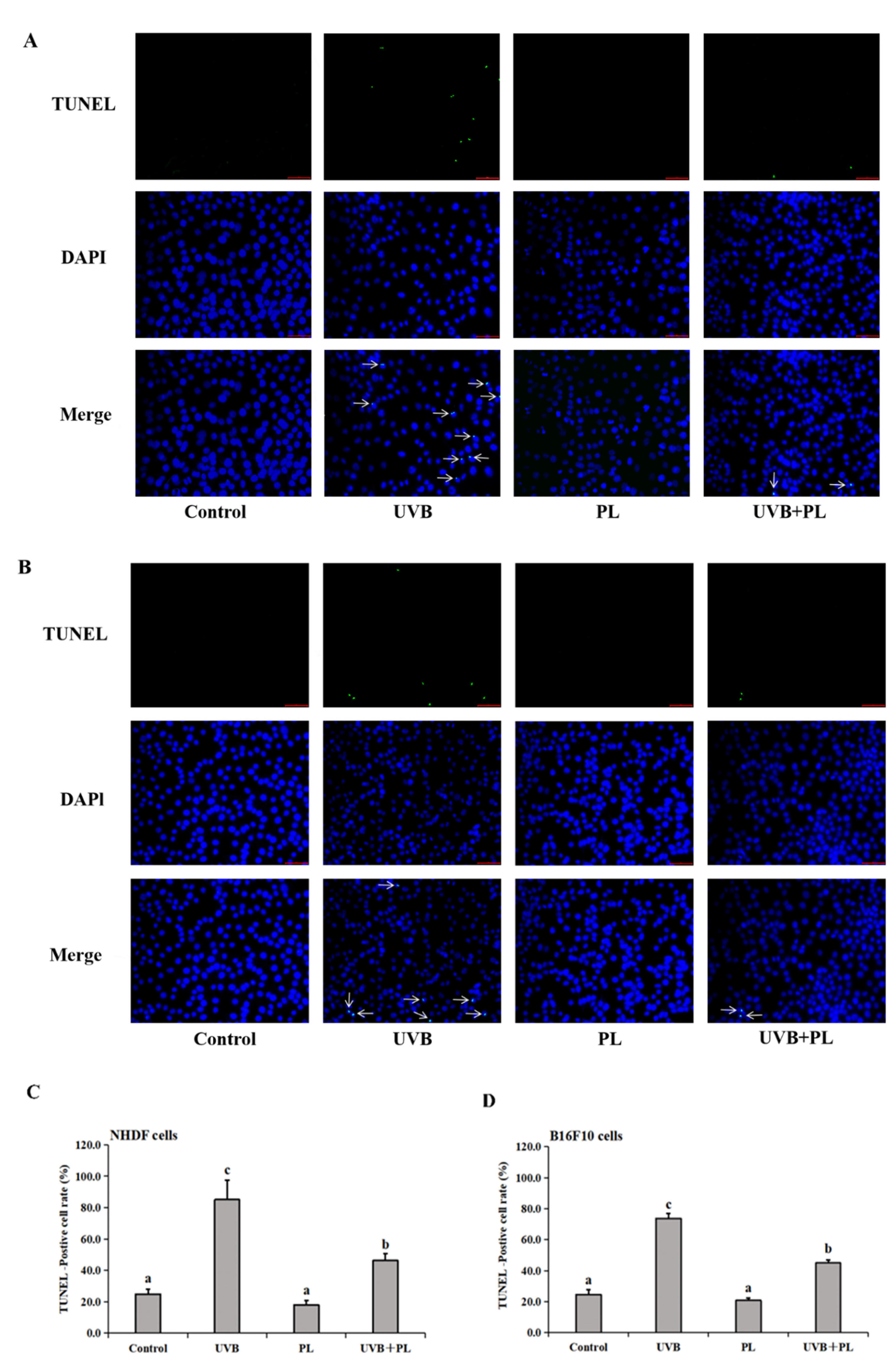
3.2. The Effects of PL on DNA Damage in NHDF and B16F10 Cells
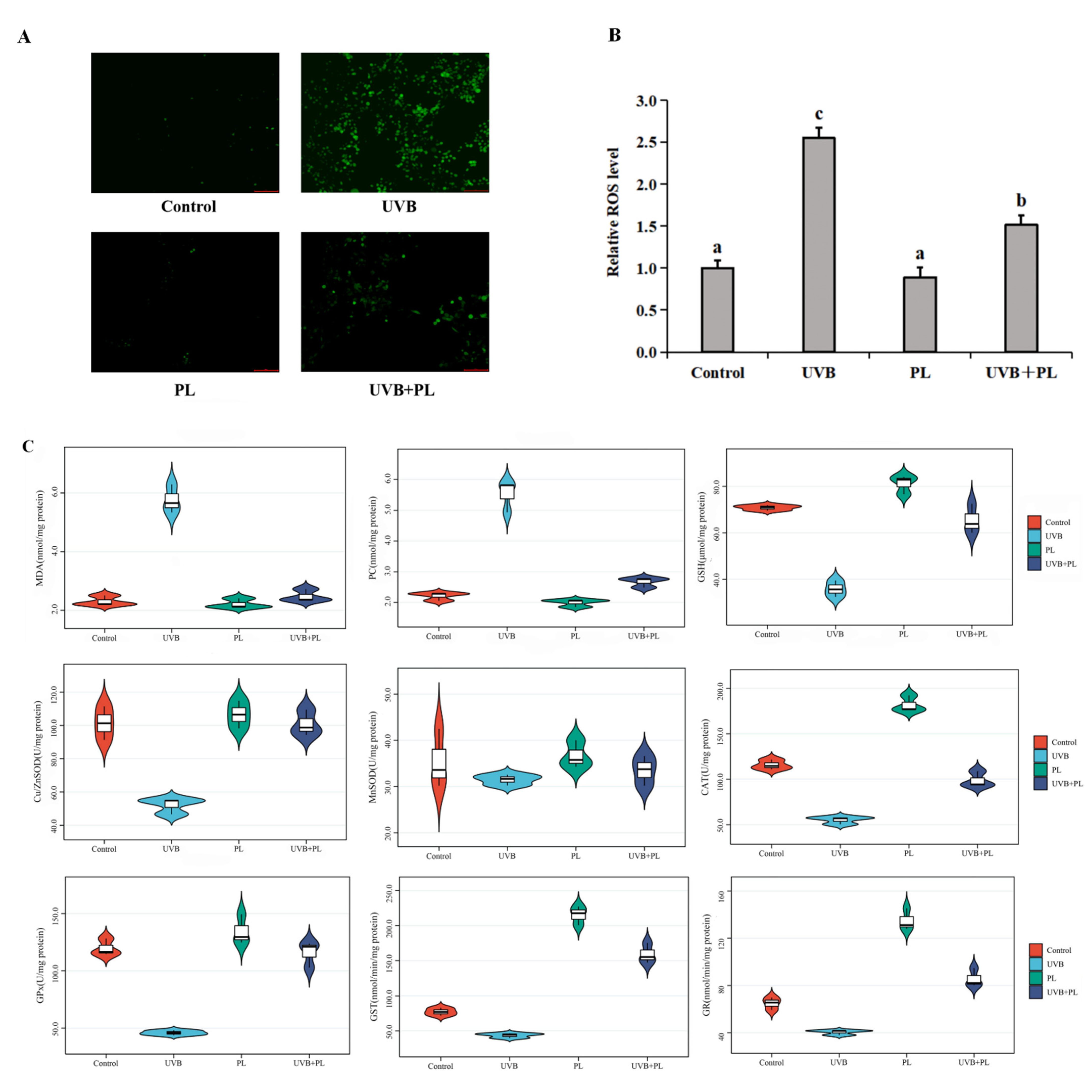
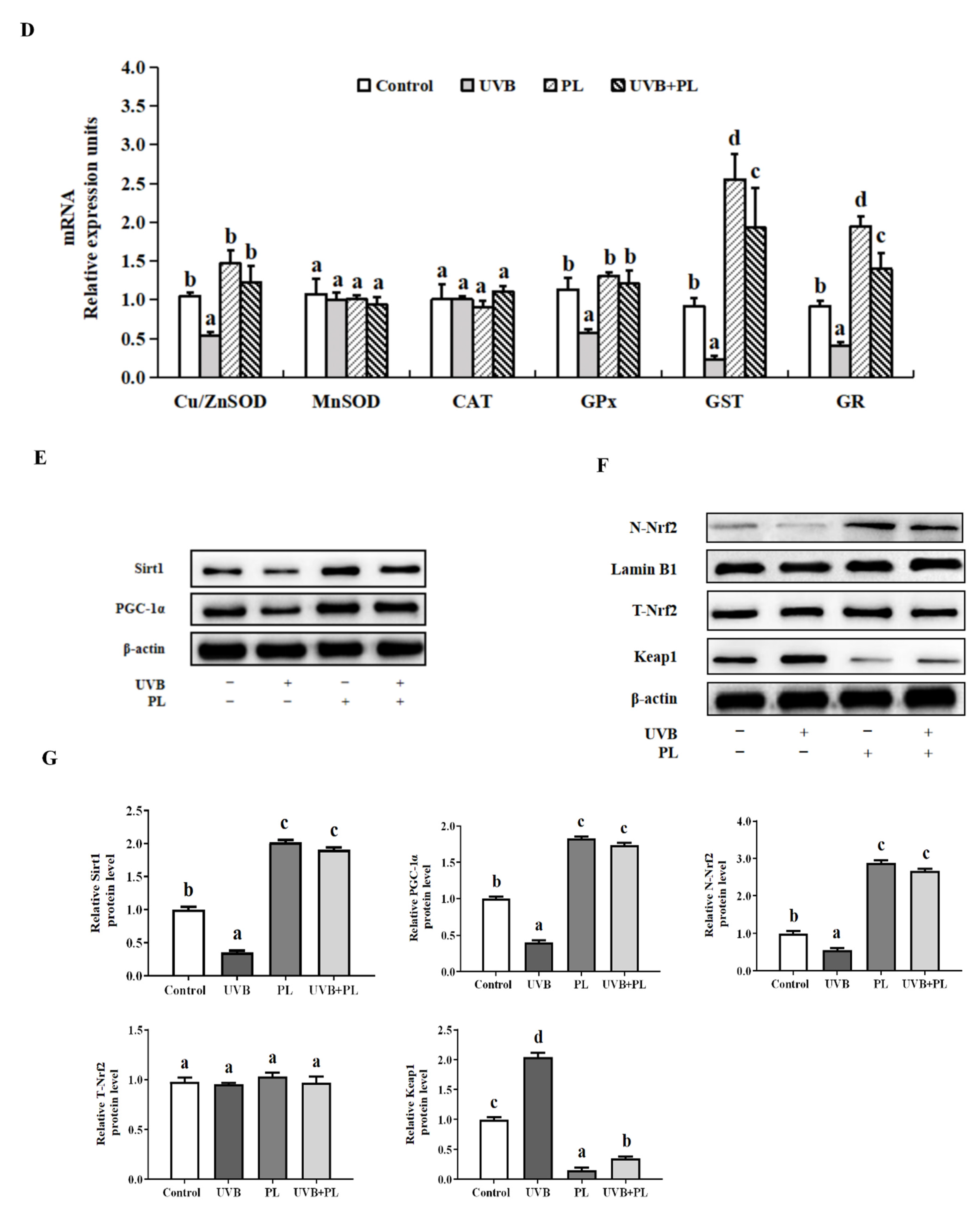
3.3. The Effects of PL on Oxidative Damage and Antioxidant Capacity-Related Parameters in NHDF Cells
3.4. The Effects of PL on the Percentage of SA-β-gal Positive NHDF Cells
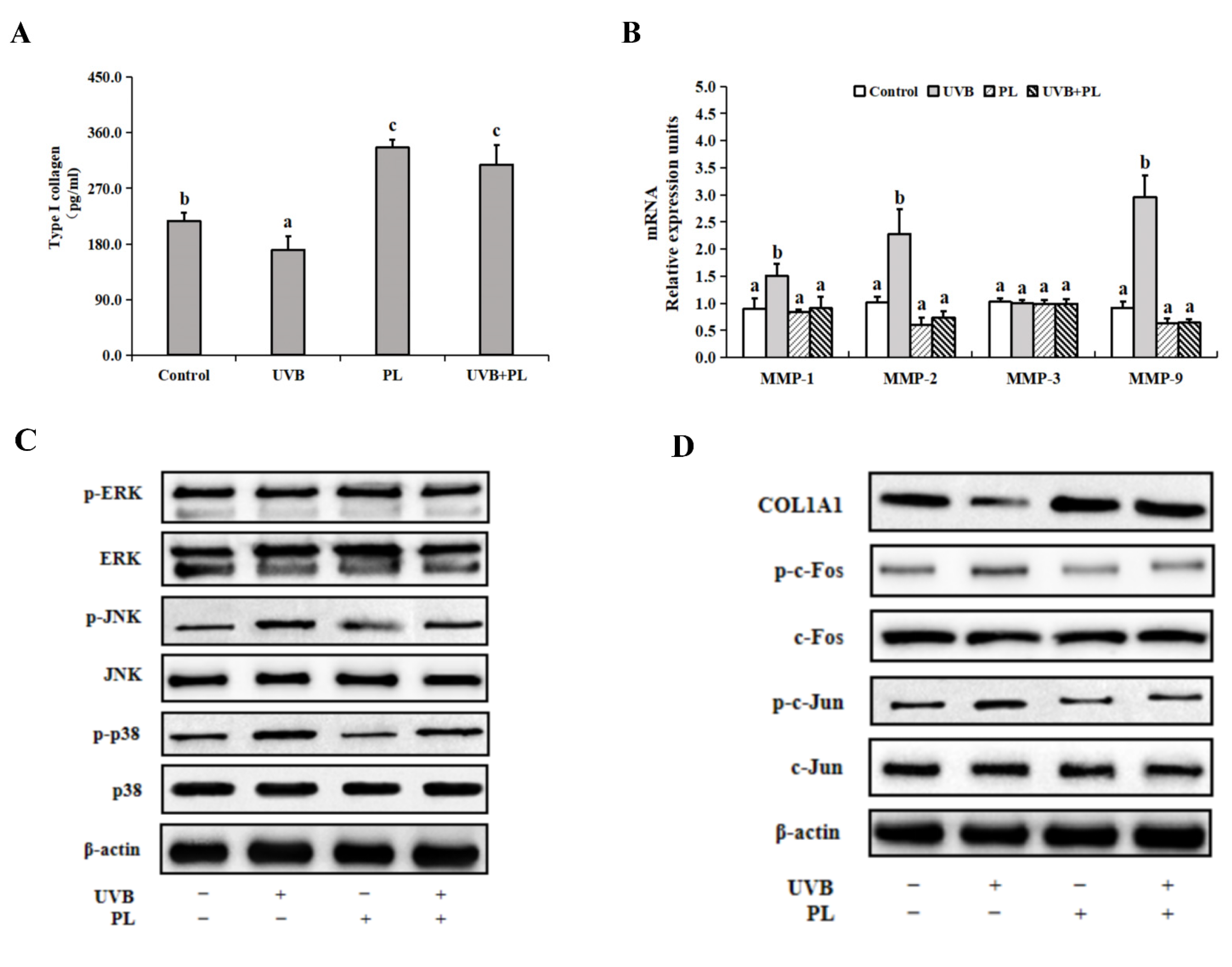
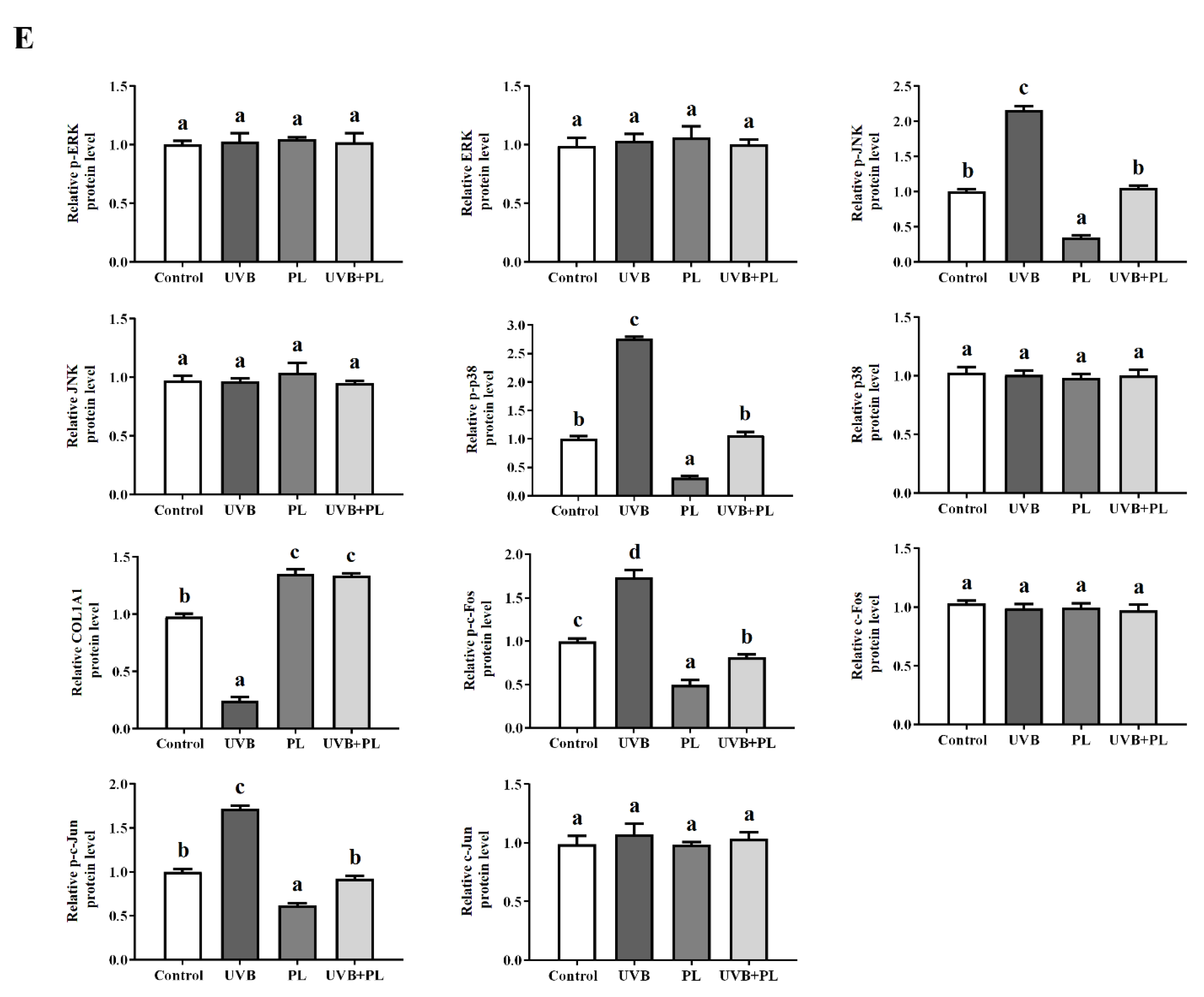
3.5. The Effects of PL on Wrinkle Formation-Related Parameters in NHDF Cells
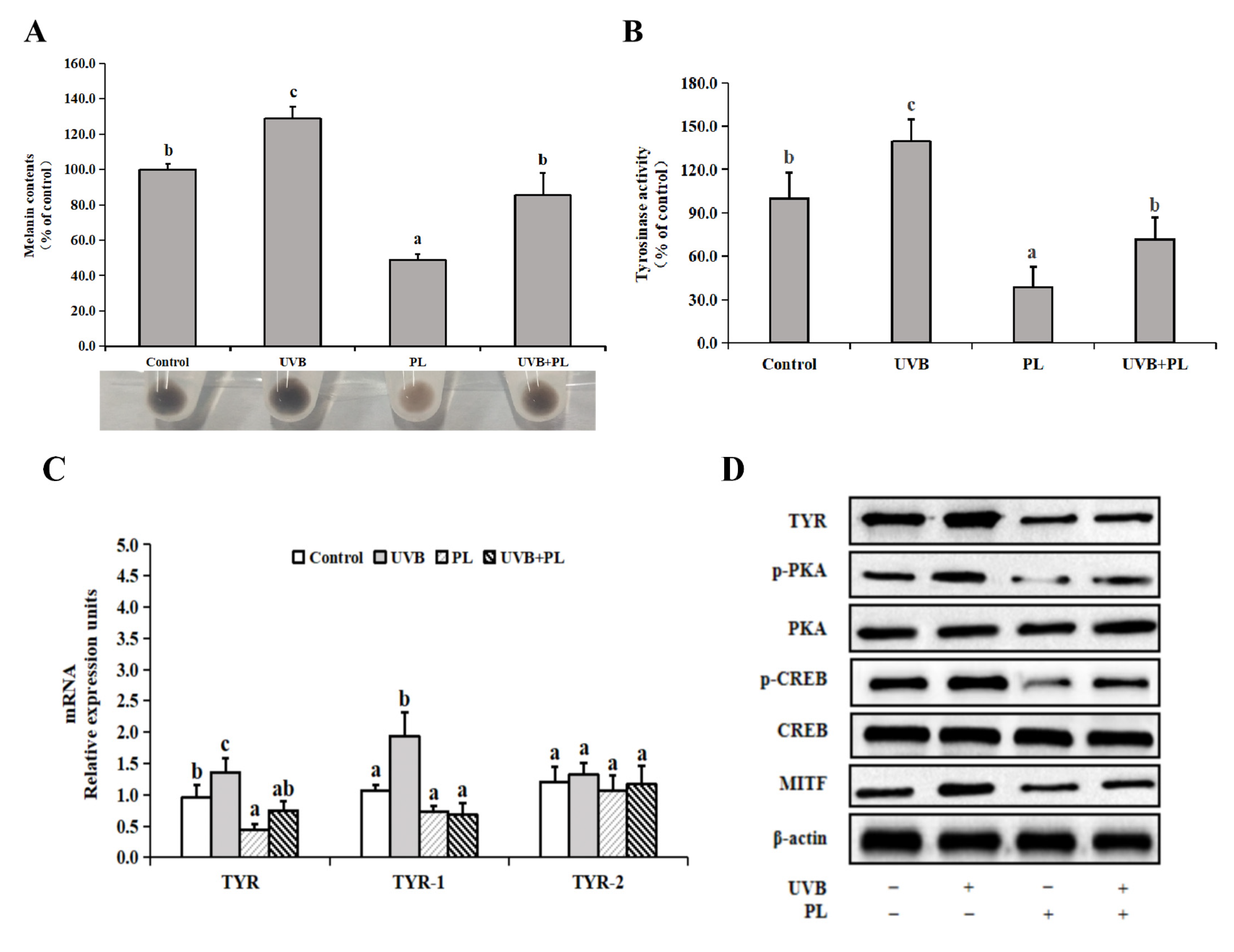
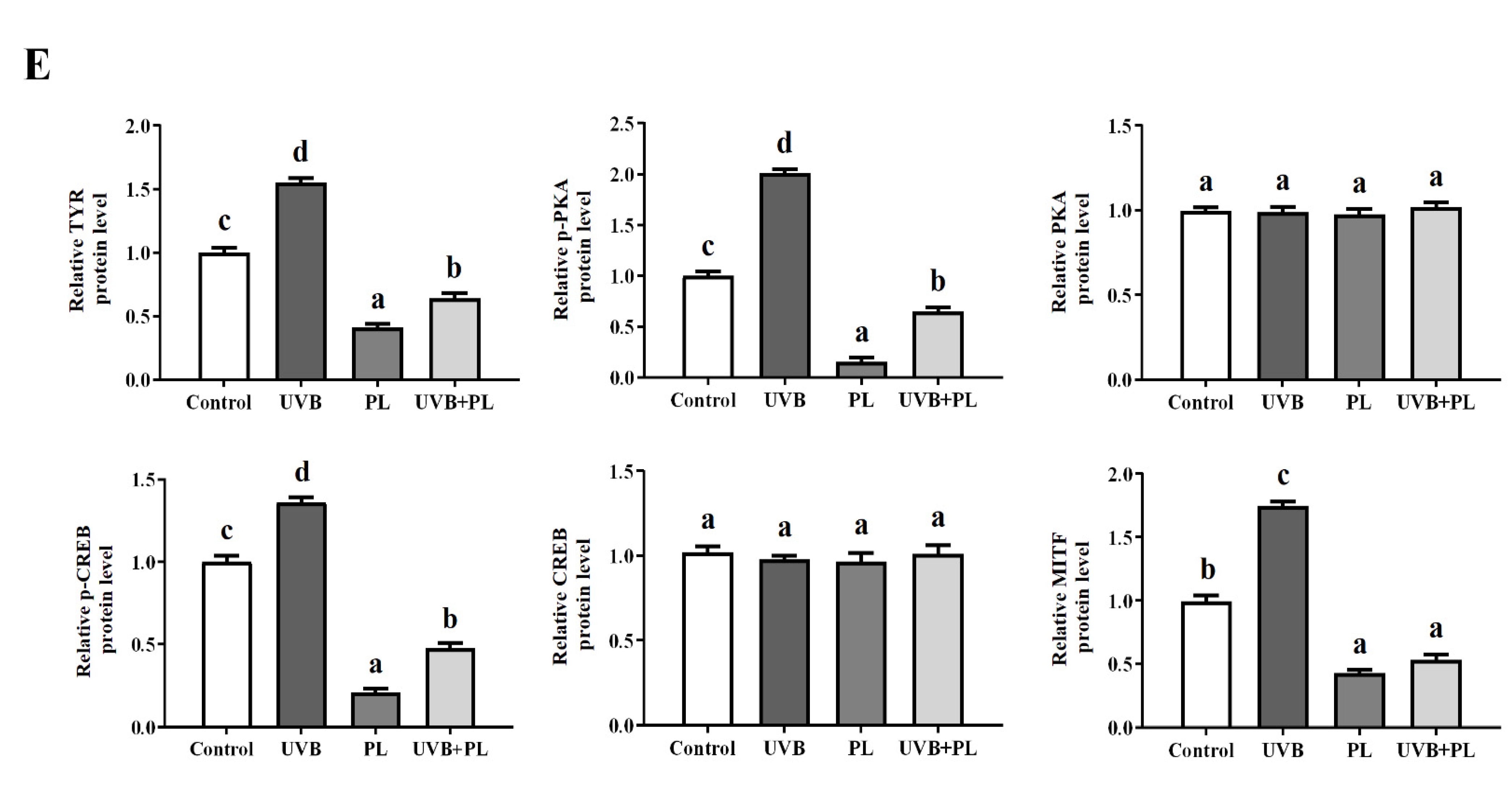
3.6. The Effects of PL on Melanogenesis-Related Parameters in B16F10 Cells
4. Discussion
4.1. PL Prevented UVB-Induced Cytotoxicity in NHDF and B16F10 Cells
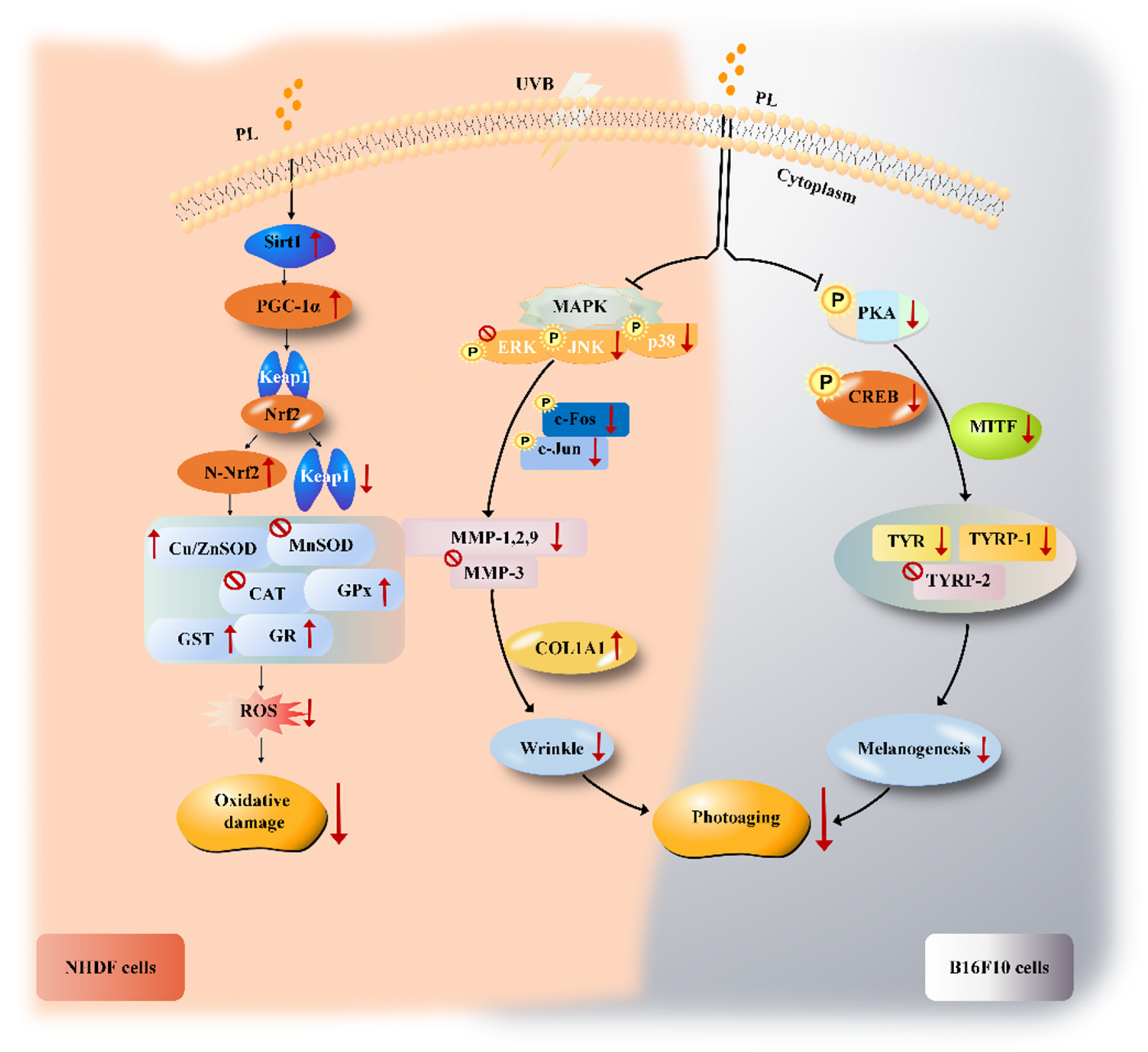
4.2. PL Alleviated UVB-Induced Oxidative Damage Relating to Sirt1/PGC-1α/Nrf2 Signaling Pathway in NHDF Cells
4.2.1. PL Alleviated UVB-Induced Oxidative Damage through Increasing Antioxidant Ability in NHDF Cells
4.2.2. PL Increased Antioxidant Ability Relating to Sirt1/PGC-1α/Nrf2 Signaling Pathway in NHDF Cells
4.3. PL Attenuated UVB-Induced Photoaging through Anti-Wrinkling and Anti-Melanogenic Effects in NHDF and B16F10 Cells
4.3.1. PL Attenuated UVB-Induced Photoaging in NHDF Cells
4.3.2. PL Attenuated UVB-Induced Photoaging through Exerting an Anti-Wrinkling Effect Relating to (JNK, p38)/(c-Fos, c-Jun) Signaling Pathway in NHDF Cells
4.3.3. PL Attenuated UVB-Induced Photoaging through Exerting an Anti-Melanogenesis Effect Relating to PKA/CREB/MITF Signaling Pathway in B16F10 Cells
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hwang, S.; Dong, M.S.; Hong, J.H. Intracellular Ca2+-Mediated AE2 Is Involved in the Vectorial Movement of HaCaT Keratinocyte. Int. J. Mol. Sci. 2020, 21, 8429. [Google Scholar] [CrossRef]
- Su, J.L.; Park, J.W.J. Enhancement of UVB radiation-mediated apoptosis by knockdown of cytosolic NADP+-dependent isocitrate dehydrogenase in HaCaT cells. BMB Rep. 2014, 47, 209. [Google Scholar] [CrossRef]
- Ho, Y.Y.; Sun, D.S.; Chang, H.H. Silver Nanoparticles Protect Skin from Ultraviolet B-Induced Damage in Mice. Int. J. Mol. Sci. 2020, 21, 7082. [Google Scholar] [CrossRef]
- Xiao, X.; Huang, M.; Fan, C.; Zuo, F. DUOX2 participates in skin aging induced by UVB in HSF2 cells by activating NFκB signaling. Exp. Ther. Med. 2020, 21, 157. [Google Scholar] [CrossRef]
- Nguyen, Q.T.; Fang, M.; Do, N.Q.; Jeong, J.; Oh, S.; Zheng, S.; Kim, M.; Choi, J.; Lim, S.; Yi, T.H. Anemopsis californica Attenuates Photoaging by Regulating MAPK, NRF2, and NFATc1 Signaling Pathways. Antioxidants 2021, 10, 1882. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.M.; Ratti, B.A.; Daré, R.G.; Silva, S.O.; da Conceição T Truiti, M.; Ueda-Nakamura, T.; Auzély-Velty, R.; Nakamura, C.V. Dihydrocaffeic acid prevents UVB-induced oxidative stress leading to the inhibition of apoptosis and MMP-1 expression via p38 signaling pathway. Oxidative Med. Cell. Longev. 2019, 2019, 2419096. [Google Scholar] [CrossRef]
- Liu, S.W.; Lu, Y.; Zhao, Y.X.; Chang, X.D. Hawthorn Polyphenol Extract Inhibits UVB-Induced Skin Photoaging by Regulating MMP Expression and Type I Procollagen Production in Mice. J. Agric. Food Chem. 2018, 66, 8537–8546. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, J.E.; Choi, S.S.; Park, T.H. Protective Effects of Silkworm Hemolymph Extract and Its Fractions on UV-induced Photoaging. Biotechnol. Bioprocess Eng. 2017, 22, 37–44. [Google Scholar] [CrossRef]
- Kim, K.; Jeong, H.I.; Yang, I.; Nam, S.J.; Lim, K.M. Acremonidin E produced by Penicillium sp. SNF123, a funga. endophyte of Panax ginseng, has anti-melanogenic activities. J. Ginseng Res. 2019, 45, 98–107. [Google Scholar] [CrossRef]
- Ro, J.; Kim, Y.; Kim, H.; Park, K.; Lee, K.E.; Khadka, P.; Yun, G.; Park, J.; Chang, S.T.; Lee, J.; et al. Pectin Micro- and Nano-capsules of Retinyl Palmitate as Cosmeceutical Carriers for Stabilized Skin Transport. Korean J. Physiol. Pharmacol. 2015, 19, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Notay, M.; Foolad, N.; Vaughn, A.R.; Sivamani, R.K. Probiotics, Prebiotics, and Synbiotics for the Treatment and Prevention of Adult Dermatological Diseases. Am. J. Clin. Dermatol. 2017, 18, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Benyacoub, J.; Bosco, N.; Blanchard, C.; Demont, A.; Philippe, D.; Castielhigounenc, I.; Guéniche, A. Immune modulation property of Lactobacillus paracasei NCC2461 (ST11) strain and impact on skin defences. Benef. Microbes 2014, 5, 129–136. [Google Scholar] [CrossRef]
- Philippe, D.; Stephanie, B.; Benyacoub, J. Oral Lactobacillus paracasei improves skin barrier function recovery and reduces local skin inflammation. Eur. J. Dermatol. 2011, 21, 279–280. [Google Scholar] [CrossRef]
- Chaiyasut, C.; Tirawat, Y.; Sivamaruthi, B.S.; Kesika, P.; Thangaleela, S.; Khongtan, S.; Khampithum, N.; Peerajan, S.; Chaiyasut, K.; Sirilun, S. Effect of Lactobacillus paracasei HII01 supplementation on total cholesterol, and on the parameters of lipid and carbohydrate metabolism, oxidative stress, inflammation and digestion in Thai hypercholesterolemic subjects. Appl. Sci. 2021, 11, 4333. [Google Scholar] [CrossRef]
- Piqué, N.; Berlanga, M.; Miñana-Galbis, D. Health benefits of heat-killed (Tyndallized) probiotics: An overview. Int. J. Mol. Sci. 2019, 20, 2534. [Google Scholar] [CrossRef] [PubMed]
- Akter, S.; Park, J.H.; Jung, H.K. Potential Health-Promoting Benefits of Paraprobiotics, Inactivated Probiotic Cells. J. Microbiol. Biotechnol. 2020, 30, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Hiromi, K.N. New lactic acid bacteria for skin health via oral intake of heat-killed or live cells. Anim. Sci. J. 2018, 89, 835–842. [Google Scholar] [CrossRef]
- Ogawa, M.; Saiki, A.; Matsui, Y.; Tsuchimoto, N.; Nakakita, Y.; Takata, Y.; Nakamura, T. Effects of oral intake of heat-killed Lactobacillus brevis SBC8803 (SBL88™) on dry skin conditions: A randomized, double-blind, placebo-controlled study. Exp. Ther. Med. 2016, 12, 3863–3872. [Google Scholar] [CrossRef] [PubMed]
- Maehata, H.; Arai, S.; Iwabuchi, N.; Abe, F. Immuno-modulation by heat-killed Lacticaseibacillus paracasei MCC1849 and its application to food products. Int. J. Immunopathol. Pharmacol. 2021, 35, 20587384211008291. [Google Scholar] [CrossRef] [PubMed]
- Moroi, M.; Uchi, S.; Nakamura, K.; Sato, S.; Shimizu, N.; Fujii, M.; Kumagai, T.; Saito, M.; Uchiyama, K.; Watanabe, T. Beneficial effect of a diet containing heat-killed Lactobacillus paracasei K71 on adult type atopic dermatitis. J. Dermatol. 2011, 38, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.-H.; Chou, C.-H.; Huang, T.-Y.; Wang, H.-L.; Chien, P.-J.; Chang, W.-W.; Lee, H.-T. Heat-Killed Lactobacilli Preparations Promote Healing in the Experimental Cutaneous Wounds. Cells 2021, 10, 3264. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Toalá, J.; Garcia-Varela, R.; Garcia, H.; Mata-Haro, V.; González-Córdova, A.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Technol. 2018, 75, 105–114. [Google Scholar] [CrossRef]
- Vinderola, G.; Sanders, M.E.; Salminen, S. The Concept of Postbiotics. Foods 2022, 11, 1077. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.R.; Kim, H.; Jung, B.J.; You, G.E.; Chung, D.K. Lipoteichoic Acid Isolated from Lactobacillus plantarum Inhibits Melanogenesis in B16F10 Mouse Melanoma Cells. Mol. Cells 2015, 38, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.F.; Lee, H.Y.; Jung, B.J.; Jang, S.; Chung, D.K.; Kim, H. Lipoteichoic acid isolated from Lactobacillus plantarum down-regulates UV-induced MMP-1 expression and up-regulates type I procollagen through the inhibition of reactive oxygen species generation. Mol. Immunol. 2015, 67, 248–255. [Google Scholar] [CrossRef]
- Yi, Z.J.; Fu, Y.R.; Li, M.; Gao, K.S.; Zhang, X.G. Effect of LTA isolated from bifidobacteria on D-galactose-induced aging. Exp. Gerontol. 2009, 44, 760–765. [Google Scholar] [CrossRef]
- Zhou, Y.; Baker, J.S.; Chen, X.; Wang, Y.; Chen, H.; Davison, G.W.; Yan, X. High-dose astaxanthin supplementation suppresses antioxidant enzyme activity during moderate-intensity swimming training in mice. Nutrients 2019, 11, 1244. [Google Scholar] [CrossRef]
- Lu, Z.; Feng, L.; Jiang, W.-D.; Wu, P.; Liu, Y.; Jiang, J.; Kuang, S.-Y.; Tang, L.; Li, S.-W.; Liu, X.-A. Mannan Oligosaccharides Application: Multipath Restriction from Aeromonas hydrophila Infection in the Skin Barrier of Grass Carp (Ctenopharyngodon idella). Front. Immunol. 2021, 12, 742107. [Google Scholar] [CrossRef]
- Lee, J.-J.; Ng, S.-C.; Hsu, J.-Y.; Liu, H.; Chen, C.-J.; Huang, C.-Y.; Kuo, W.-W.I. Galangin Reverses H2O2-Induced Dermal Fibroblast Senescence via SIRT1-PGC-1α/Nrf2 Signaling. Int. J. Mol. Sci. 2022, 23, 1387. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, J.; Yi, R.; Mu, J.; Zhao, X.; Yang, Z. Hepatoprotective effects of Lactobacillus on carbon tetrachloride-induced acute liver injury in mice. Int. J. Mol. Sci. 2018, 19, 2212. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Lee, J.; Yeom, Z.; Heo, D.; Lim, Y.-H. Neuroprotective effect of Ruminococcus albus on oxidatively stressed SH-SY5Y cells and animals. Sci. Rep. 2017, 7, 14520. [Google Scholar] [CrossRef]
- Luo, D.; Luo, M.; Wang, H.; Liu, X.; Yang, M.; Tian, F.; Qin, S.; Liu, J. Protective Effects of Lactobacillus rhamnosus Peptides against DSS-Induced Inflammatory and Oxidative Damages in Human Colonic Epithelial Cells through NF-κB/Nrf2/HO-1 Signaling Pathway. Int. J. Pept. Res. Ther. 2022, 28, 115. [Google Scholar] [CrossRef]
- Meng, Q.; Qi, X.; Fu, Y.; Chen, Q.; Bian, H. Flavonoids extracted from mulberry (Morus alba L.) leaf improve skeletal muscle mitochondrial function by activating AMPK in type 2 diabetes. J. Ethnopharmacol. 2019, 248, 112326. [Google Scholar] [CrossRef]
- Guo, W.W.; Wang, X.; Chen, X.Q.; Ba, Y.Y.; Wu, X. Flavonones from Penthorum chinense Ameliorate Hepatic Steatosis by Activating the SIRT1/AMPK Pathway in HepG2 Cells. Int. J. Mol. Sci. 2018, 19, 2555. [Google Scholar] [CrossRef]
- Wlaschek, M.; Heinen, G.; Poswig, A.; Schwarz, A.; Krieg, T.; Scharffetter-Kochanek, K. UVA-induced autocrine stimulation of fibroblast-derived collagenase/mmp-1 by interrelated loops ofinterleukin–1 andinterleukin–6. Photochem. Photobiol. 1994, 59, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.G.; Kim, H.H.; Paul, S.; Jang, Y.J.; Cho, Y.H.; Kim, H.J.; Yu, J.M.; Lee, E.S.; An, B.J.; Kang, S.C.; et al. Quercetin-3-O-β-d-glucopyranosyl-(1→6)-β-d-glucopyranoside suppresses melanin synthesis by augmenting p38 MAPK and CREB signaling pathways and subsequent cAMP down-regulation in murine melanoma cells. Saudi J. Biol. Sci. 2015, 22, 706–713. [Google Scholar] [CrossRef]
- Lim, H.Y.; Jeong, D.; Park, S.H.; Shin, K.K.; Hong, Y.H.; Kim, E.; Yu, Y.-G.; Kim, T.-R.; Kim, H.; Lee, J. Antiwrinkle and antimelanogenesis effects of tyndallized Lactobacillus acidophilus KCCM12625P. Int. J. Mol. Sci. 2020, 21, 1620. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Lee, S.; Jang, S.-W.; Shin, H.S.; Park, J.-H.; Park, M.S.; Lee, B.-H. Lysed and disrupted Bifidobacterium bifidum BGN4 cells promote anti-inflammatory activities in lipopolysaccharide-stimulated RAW 264.7 cells. Saudi J. Biol. Sci. 2021, 28, 5115–5118. [Google Scholar] [CrossRef]
- Kim, D.O.; Byun, J.-E.; Seong, H.-A.; Yoon, S.R.; Choi, I.; Jung, H. Thioredoxin-interacting protein-derived peptide (TN13) inhibits LPS-induced inflammation by inhibiting p38 MAPK signaling. Biochem. Biophys. Res. Commun. 2018, 507, 489–495. [Google Scholar] [CrossRef]
- Arai, S.; Iwabuchi, N.; Takahashi, S.; Xiao, J.-Z.; Abe, F.; Hachimura, S.J.P.O. Orally administered heat-killed Lactobacillus paracasei MCC1849 enhances antigen-specific IgA secretion and induces follicular helper T cells in mice. PLoS ONE 2018, 13, e0199018. [Google Scholar] [CrossRef]
- Shukla, G.; Kamboj, S.; Sharma, B. Comparative analysis of antigiardial potential of heat inactivated and probiotic protein of probiotic Lactobacillus rhamnosus GG in murine giardiasis. Probiotics Antimicrob. Proteins 2020, 12, 271–279. [Google Scholar] [CrossRef]
- Ding, Y.; Jiratchayamaethasakul, C.; Lee, S.H. Protocatechuic Aldehyde Attenuates UVA-Induced Photoaging in Human Dermal Fibroblast Cells by Suppressing MAPKs/AP-1 and NF-κB Signaling Pathways. Int. J. Mol. Sci. 2020, 21, 4619. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, S.; Miao, Q.; Jin, K.; Lou, L.; Ye, X.; Xi, Y.; Ye, J. Protective role of hinokitiol against H2O2-induced injury in human corneal epithelium. Curr. Eye Res. 2017, 42, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Huang, J.; Pei, S.; Ouyang, Y.; Ding, Y.; Jiang, L.; Lu, J.; Kang, L.; Huang, L.; Xiang, H. Ganoderma lucidum polysaccharide inhibits UVB-induced melanogenesis by antagonizing cAMP/PKA and ROS/MAPK signaling pathways. J. Cell. Physiol. 2019, 234, 7330–7340. [Google Scholar] [CrossRef] [PubMed]
- Permatasari, F.; Hu, Y.-Y.; Zhang, J.-A.; Zhou, B.-R.; Luo, D. Anti-photoaging potential of Botulinum Toxin Type A in UVB-induced premature senescence of human dermal fibroblasts in vitro through decreasing senescence-related proteins. J. Photochem. Photobiol. B Biol. 2014, 133, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Jiang, R.; Zhou, J.; Xu, X.; Sun, L. Salicylic acid in ginseng root alleviates skin hyperpigmentation disorders by inhibiting melanogenesis and melanosome transport. Eur. J. Pharmacol. 2021, 910, 174458. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Xu, C.; Holck, A.L.; Liu, R. Acrylamide inhibits autophagy, induces apoptosis and alters cellular metabolic profiles. Ecotoxicol. Environ. Saf. 2021, 208, 111543. [Google Scholar] [CrossRef]
- Song, Y.; Yan, L.-C.; Xiao, W.-W.; Feng, L.; Jiang, W.-D.; Wu, P.; Liu, Y.; Kuang, S.-Y.; Tang, L.; Zhou, X.-Q. Enzyme-treated soy protein supplementation in low protein diet enhanced immune function of immune organs in on-growing grass carp. Fish Shellfish. Immunol. 2020, 106, 318–331. [Google Scholar] [CrossRef] [PubMed]
- Sotiropoulou, G.; Zingkou, E.; Pampalakis, G. Redirecting drug repositioning to discover innovative cosmeceuticals. Exp. Dermatol. 2021, 30, 628–644. [Google Scholar] [CrossRef]
- Reisch, M. The Microbiome comes to cosmetics. Chem. Eng. News 2017, 95, 30–34. [Google Scholar] [CrossRef]
- Takano, N.; Takahashi, Y.; Yamamoto, M.; Teranishi, M.; Yamaguchi, H.; Sakamoto, A.N.; Hase, Y.; Fujisawa, H.; Wu, J.; Matsumoto, T. Isolation of a novel UVB-tolerant rice mutant obtained by exposure to carbon-ion beams. J. Radiat. Res. 2013, 54, 637–648. [Google Scholar] [CrossRef]
- Masashi, M.; Masami, K.; Tomoyuki, F.; Taketo, Y.; Masamitsu, I. Exopolysaccharides Isolated from Milk Fermented with Lactic Acid Bacteria Prevent Ultraviolet-Induced Skin Damage in Hairless Mice. Int. J. Mol. Sci. 2017, 18, 146. [Google Scholar] [CrossRef]
- Lim, S.-M.; Lee, N.-K.; Paik, H.-D. Potential neuroprotective effects of heat-killed Lactococcus lactis KC24 using SH-SY5Y cells against oxidative stress induced by hydrogen peroxide. Food Sci. Biotechnol. 2020, 29, 1735–1740. [Google Scholar] [CrossRef] [PubMed]
- Cheon, M.-J.; Lee, N.-K.; Paik, H.-D. Neuroprotective effects of heat-killed Lactobacillus plantarum 200655 isolated from kimchi against oxidative stress. Probiotics Antimicrob. Proteins 2021, 13, 788–795. [Google Scholar] [CrossRef]
- Chuang, C.-H.; Tsai, C.-C.; Lin, E.-S.; Huang, C.-S.; Lin, Y.-Y.; Lan, C.-C.; Huang, C.-C. Heat-killed Lactobacillus salivarius and Lactobacillus johnsonii reduce liver injury induced by alcohol in vitro and in vivo. Molecules 2016, 21, 1456. [Google Scholar] [CrossRef]
- Kai, L.; Xta, B.; Bo, W.; Cong, W.; Lwa, B.; Sza, B.; Yang, L.; Tla, B.; Sda, B. Evaluation of paraprobiotic applicability of Clostridium butyricum CBG01 in improving the growth performance, immune responses and disease resistance in Pacific white shrimp, Penaeus vannamei. Aquaculture 2021, 544, 737041. [Google Scholar] [CrossRef]
- Zhao, B.-B.; Meng, J.; Zhang, Q.-X.; Kang, T.-T.; Lu, R.-R. Protective effect of surface layer proteins isolated from four Lactobacillus strains on hydrogen-peroxide-induced HT-29 cells oxidative stress. Int. J. Biol. Macromol. 2017, 102, 76–83. [Google Scholar] [CrossRef]
- Ryu, J.-S.; Kang, H.-Y.; Lee, J.K. Effect of treadmill exercise and trans-cinnamaldehyde against d-galactose-and aluminum chloride-induced cognitive dysfunction in mice. Brain Sci. 2020, 10, 793. [Google Scholar] [CrossRef]
- Zhao, Z.; Tang, Z.; Zhang, W.; Liu, J.; Li, B.; Ding, S. Inactivated pseudomonas aeruginosa protects against myocardial ischemia reperfusion injury via Nrf2 and HO-1. Exp. Ther. Med. 2020, 19, 3362–3368. [Google Scholar] [CrossRef]
- Li, J.; Li, Q.; Gao, N.; Wang, Z.; Li, F.; Li, J.; Shan, A. Exopolysaccharides produced by Lactobacillus rhamnosus GG alleviate hydrogen peroxide-induced intestinal oxidative damage and apoptosis through the Keap1/Nrf2 and Bax/Bcl-2 pathways in vitro. Food Funct. 2021, 12, 9632–9641. [Google Scholar] [CrossRef]
- Karaca, B.; Yilmaz, M.; Gursoy, U.K. Targeting Nrf2 with Probiotics and Postbiotics in the Treatment of Periodontitis. Biomolecules 2022, 12, 729. [Google Scholar] [CrossRef]
- Ben-Dor, A.; Steiner, M.; Gheber, L.; Danilenko, M.; Dubi, N.; Linnewiel, K.; Zick, A.; Sharoni, Y.; Levy, J. Carotenoids activate the antioxidant response element transcription system. Mol. Cancer Ther. 2005, 4, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Jia, Y.; Yang, S.; Zhao, N.; Hu, Y.; Hong, J.; Gao, S.; Zhao, R. Sodium butyrate protects against high-fat diet-induced oxidative stress in rat liver by promoting expression of nuclear factor E2-related factor 2. Br. J. Nutr. 2019, 122, 400–410. [Google Scholar] [CrossRef]
- Nam, Y.; Kim, J.; Baek, J.; Kim, W. Improvement of Cutaneous Wound Healing via Topical Application of Heat-Killed Lactococcus chungangensis CAU 1447 on Diabetic Mice. Nutrients 2021, 13, 2666. [Google Scholar] [CrossRef]
- Doi, M.; Yamaoka, I.; Fukunaga, T.; Nakayama, M. Isoleucine, a potent plasma glucose-lowering amino acid, stimulates glucose uptake in C2C12 myotubes. Biochem. Biophys. Res. Commun. 2003, 312, 1111–1117. [Google Scholar] [CrossRef] [PubMed]
- La Sala, L.; Cattaneo, M.; De Nigris, V.; Pujadas, G.; Testa, R.; Bonfigli, A.R.; Genovese, S.; Ceriello, A. Oscillating glucose induces microRNA-185 and impairs an efficient antioxidant response in human endothelial cells. Cardiovasc. Diabetol. 2016, 15, 71. [Google Scholar] [CrossRef]
- Liang, Y.; Simaiti, A.; Xu, M.; Lv, S.; Jiang, H.; He, X.; Fan, Y.; Zhu, S.; Du, B.; Yang, W.J.N. Antagonistic Skin Toxicity of Co-Exposure to Physical Sunscreen Ingredients Zinc Oxide and Titanium Dioxide Nanoparticles. Nanomaterials 2022, 12, 2769. [Google Scholar] [CrossRef]
- Sanches, P.L.; Geaquinto, L.R.d.O.; Cruz, R.; Schuck, D.C.; Lorencini, M.; Granjeiro, J.M.; Ribeiro, A.R.L. Toxicity evaluation of TiO2 nanoparticles on the 3D skin model: A systematic review. Front. Bioeng. Biotechnol. 2020, 8, 575. [Google Scholar] [CrossRef]
- Gollavilli, H.; Hegde, A.R.; Managuli, R.S.; Bhaskar, K.V.; Dengale, S.J.; Reddy, M.S.; Kalthur, G.; Mutalik, S. Naringin nano-ethosomal novel sunscreen creams: Development and performance evaluation. Colloids Surf. B Biointerfaces 2020, 193, 111122. [Google Scholar] [CrossRef]
- Wen, W.J.; Chen, J.W.; Ding, L.G.; Luo, X.; Zheng, X.P.; Dai, Q.; Gu, Q.Q.; Cui, L.; Liang, M.; Guo, X.L.; et al. Astragaloside exerts anti-photoaging effects in UVB-induced premature senescence of rat dermal fibroblasts through enhanced autophagy. Arch. Biochem. Biophys. 2018, 657, 31–40. [Google Scholar] [CrossRef]
- Song, F.; Wang, L.; Mu, J.; Ma, H. Protective Effects and Molecular Mechanism of Total Flavonoids from Lycium Barbarum Leaves on Photoaged Human Dermal Fibroblasts. Evid.-Based Complement. Altern. Med. 2022, 1, 12. [Google Scholar] [CrossRef]
- Im, A.; Lee, B.; Kang, D.J.; Chae, S. Protective effects of tyndallized Lactobacillus acidophilus IDCC 3302 against UVB-induced photodamage to epidermal keratinocytes cells. Int. J. Mol. Med. 2019, 43, 2499–2506. [Google Scholar] [CrossRef]
- İncili, G.K.; Karatepe, P.; Akgöl, M.; Kaya, B.; Kanmaz, H.; Hayaloğlu, A.A. Characterization of Pediococcus acidilactici postbiotic and impact of postbiotic-fortified chitosan coating on the microbial and chemical quality of chicken breast fillets. Int. J. Biol. Macromol. 2021, 184, 429–437. [Google Scholar] [CrossRef]
- Kwon, K.-R.; Alam, M.B.; Park, J.-H.; Kim, T.-H.; Lee, S.-H. Attenuation of UVB-induced photo-aging by polyphenolic-rich Spatholobus suberectus stem extract via modulation of MAPK/AP-1/MMPs signaling in human keratinocytes. Nutrients 2019, 11, 1341. [Google Scholar] [CrossRef]
- Pittayapruek, P.; Meephansan, J.; Prapapan, O.; Komine, M.; Ohtsuki, M. Role of matrix metalloproteinases in photoaging and photocarcinogenesis. Int. J. Mol. Sci. 2016, 17, 868. [Google Scholar] [CrossRef]
- Nam, Y.; Yoon, S.; Baek, J.; Kim, J.-H.; Park, M.; Hwang, K.; Kim, W. Heat-killed Lactiplantibacillus plantarum LRCC5314 mitigates the effects of stress-related type 2 diabetes in mice via gut microbiome modulation. J. Microbiol. Biotechnol. 2022, 32, 324–332. [Google Scholar] [CrossRef]
- Shen, J.; Sakaida, I.; Uchida, K.; Terai, S.; Okita, K. Leptin enhances TNF-α production via p38 and JNK MAPK in LPS-stimulated Kupffer cells. Life Sci. 2005, 77, 1502–1515. [Google Scholar] [CrossRef]
- Meng, Z.; Oh, S. Antioxidant and Antimelanogenic Activities of Kimchi-Derived Limosilactobacillus fermentum JNU532 in B16F10 Melanoma Cells. J. Microbiol. Biotechnol. 2021, 31, 990–998. [Google Scholar] [CrossRef]
- Lee, S.; Park, H.-O.; Yoo, W. Anti-melanogenic and antioxidant effects of cell-free supernatant from Lactobacillus gasseri BNR17. Microorganisms 2022, 10, 788. [Google Scholar] [CrossRef]
- Malka, O.; Malishev, R.; Bersudsky, M.; Rajendran, M.; Krishnamohan, M.; Shaik, J.; Tikhonov, E.; Sultan, E.; Koren, O.; Apte, R.N. Tryptophol acetate and tyrosol acetate, metabolites secreted by a probiotic yeast, hlock cytokine storm. bioRxiv 2021, 12, 472991. [Google Scholar] [CrossRef]
- Wen, K.-C.; Chang, C.-S.; Chien, Y.-C.; Wang, H.-W.; Wu, W.-C.; Wu, C.-S.; Chiang, H.-M. Tyrosol and its analogues inhibit alpha-melanocyte-stimulating hormone induced melanogenesis. Int. J. Mol. Sci. 2013, 14, 23420–23440. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Sakamoto, K. Pyruvic acid/ethyl pyruvate inhibits melanogenesis in B16F10 melanoma cells through PI3K/AKT, GSK3β, and ROS-ERK signaling pathways. Genes Cells 2019, 24, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.-C.; Lee, S.-H. Protocatechuic aldehyde inhibits α-MSH-induced melanogenesis in B16F10 melanoma cells via PKA/CREB-associated MITF downregulation. Int. J. Mol. Sci. 2021, 22, 3861. [Google Scholar] [CrossRef]
- Cho, B.-R.; Jun, H.-j.; Thach, T.T.; Wu, C.; Lee, S.-J. Betaine reduces cellular melanin content via suppression of microphthalmia-associated transcription factor in B16-F1 murine melanocytes. Food Sci. Biotechnol. 2017, 26, 1391–1397. [Google Scholar] [CrossRef]
- Sanna, C.; Maxia, A.; Fenu, G.; Loi, M.C. So uncommon and so singular, but underexplored: An updated overview on ethnobotanical uses, biological properties and phytoconstituents of sardinian endemic plants. Plants 2020, 9, 958. [Google Scholar] [CrossRef]
- Park, H.-J.; Cho, J.-H.; Hong, S.-H.; Kim, D.-H.; Jung, H.-Y.; Kang, I.-K.; Cho, Y.-J. Whitening and anti-wrinkle activities of ferulic acid isolated from Tetragonia tetragonioides in B16F10 melanoma and CCD-986sk fibroblast cells. J. Nat. Med. 2018, 72, 127–135. [Google Scholar] [CrossRef]
- Dahal, R.H.; Nguyen, T.M.; Shim, D.S.; Kim, J.Y.; Lee, J.; Kim, J. Development of multifunctional cosmetic cream using bioactive materials from Streptomyces sp. T65 with synthesized mesoporous silica particles SBA-15. Antioxidants 2020, 9, 278. [Google Scholar] [CrossRef] [Green Version]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Zhang, X.; Song, Y.; Zheng, B.; Wen, Z.; Gong, M.; Meng, L. Heat-Killed Lacticaseibacillus paracasei Ameliorated UVB-Induced Oxidative Damage and Photoaging and Its Underlying Mechanisms. Antioxidants 2022, 11, 1875. https://doi.org/10.3390/antiox11101875
Xu J, Zhang X, Song Y, Zheng B, Wen Z, Gong M, Meng L. Heat-Killed Lacticaseibacillus paracasei Ameliorated UVB-Induced Oxidative Damage and Photoaging and Its Underlying Mechanisms. Antioxidants. 2022; 11(10):1875. https://doi.org/10.3390/antiox11101875
Chicago/Turabian StyleXu, Jing, Xiaofang Zhang, Yan Song, Bin Zheng, Zhengshun Wen, Miao Gong, and Lingting Meng. 2022. "Heat-Killed Lacticaseibacillus paracasei Ameliorated UVB-Induced Oxidative Damage and Photoaging and Its Underlying Mechanisms" Antioxidants 11, no. 10: 1875. https://doi.org/10.3390/antiox11101875