

Antioxidant, Anti-Inflammatory, and Antibacterial Properties of an Achillea millefolium L. Extract and Its Fractions Obtained by Supercritical Anti-Solvent Fractionation against Helicobacter pylori
 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Material and Ultrasound-Assisted Extraction of Yarrow
2.2. Supercritical Anti-Solvent Fractionation (SAF) of Yarrow Extract
2.3. Chemical Characterization of YE and Its Fractions by HPLC-PAD-ESI-QTOF-MS and GC-MS Analyses
2.4. Helicobacter pylori, Growth Media, and Culture Conditions
2.5. Human Gastric Epithelial Cell Cultures
2.6. Cell Viability
2.7. Antioxidant Activity of YE and Its Fractions against Intracellular Reactive Oxygen Species (ROS) Production on H. pylori-Infected Gastric Cells
2.8. Anti-Inflammatory Activity of YE and Its Fractions on H. pylori-Infected Gastric Cells
2.9. Antibacterial Activity of YE and Its Fractions against H. pylori Strains
2.10. Statistical Analysis
3. Results
3.1. Characterization of YE and Its Fractions
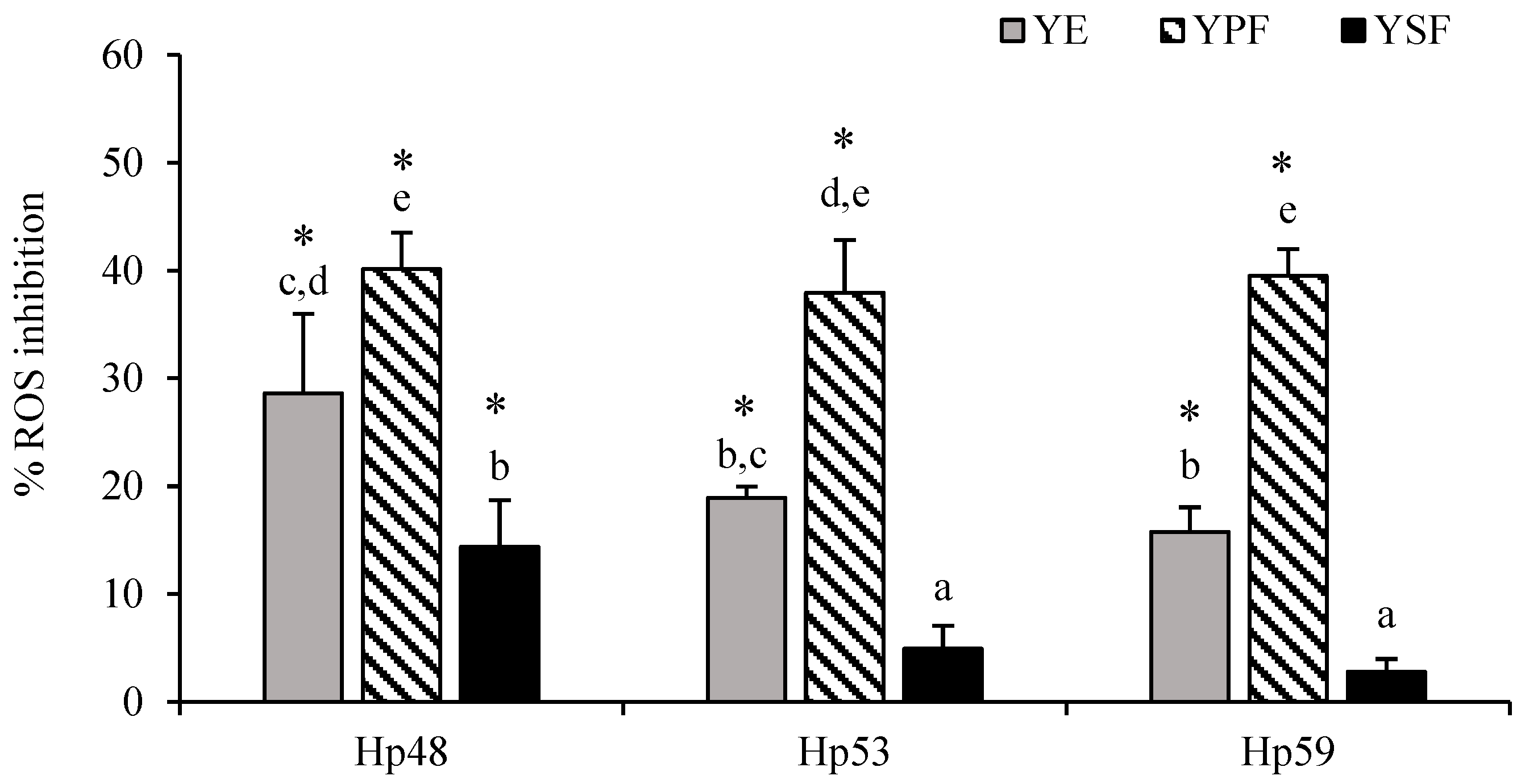
3.2. Antioxidant Activity of YE and Its Fractions against Intracellular ROS Production in H. pylori-Infected AGS Cells
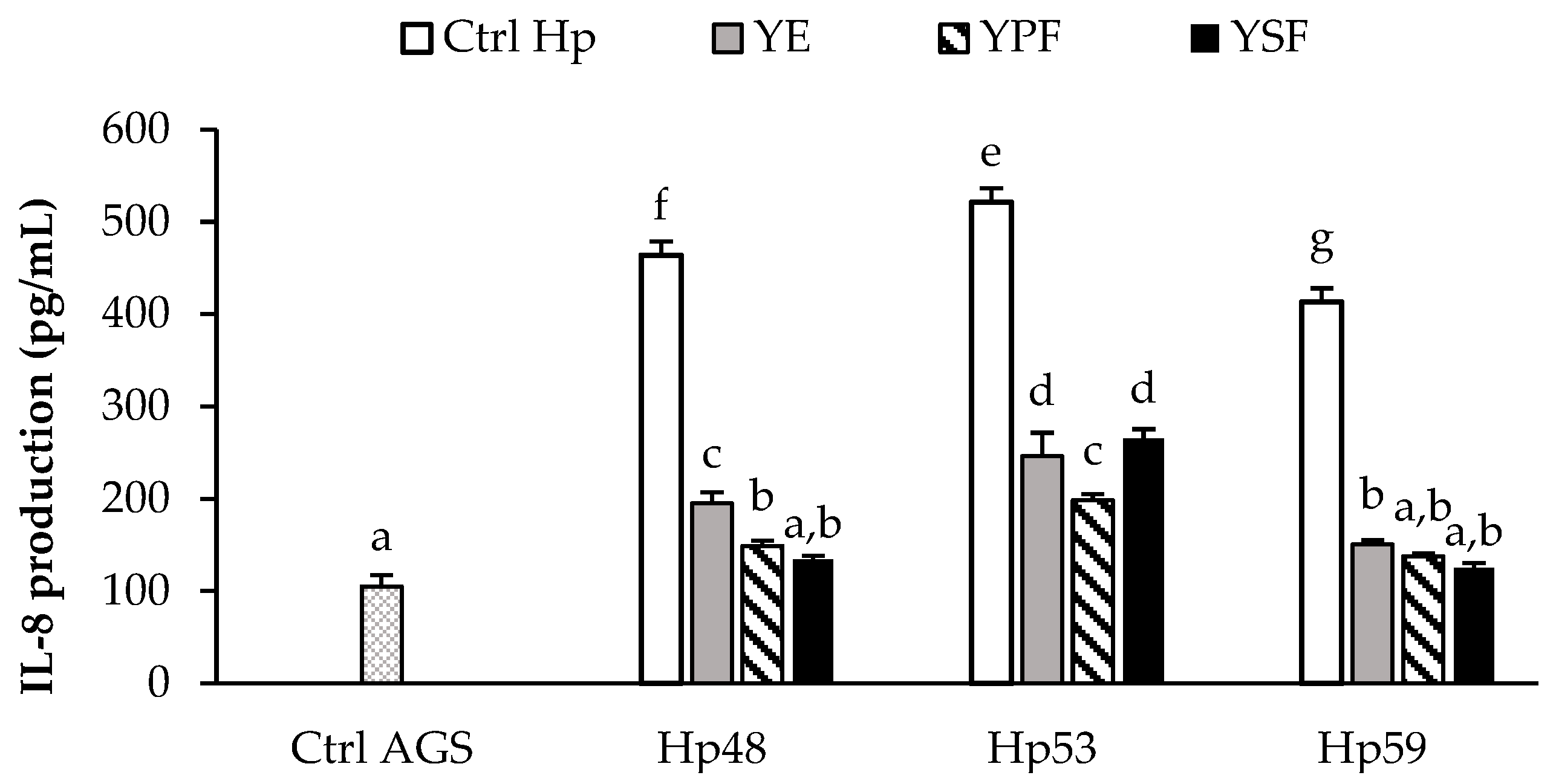
3.3. Effect of the YE and Its Fractions on the Inflammatory Response Induced by H. pylori in AGS Cells
3.4. Antibacterial Activity of YE and Its Fractions against H. pylori Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kotilea, K.; Bontems, P.; Touati, E. Epidemiology, diagnosis and risk factors of Helicobacter pylori infection. In Advances in Experimental Medicine and Biology; Kamiya, S., Backert, S., Eds.; Springer: New York, NY, USA, 2019; Volume 1149, pp. 17–33. [Google Scholar]
- Silvan, J.M.; Martinez-Rodriguez, A.J. Modulation of inflammation and oxidative stress in Helicobacter pylori infection by bioactive compounds from food components. In Current Advances for Development of Functional Foods Modulating Inflammation and Oxidative Stress; Hernandez-Ledesma, B., Martinez-Villaluenga, C., Eds.; Academic Press: London, UK, 2021; pp. 499–516. [Google Scholar]
- Alarcon, T.; Urruzuno, P.; Martinez, M.J.; Domingo, D.; Llorca, L.; Correa, A.; Lopez-Brea, M. Antimicrobial susceptibility of 6 antimicrobial agents in Helicobacter pylori clinical isolates by using EUCAST breakpoints compared with previously used breakpoints. Enferm. Infecc. Microbiol. Clin. 2017, 35, 278–282. [Google Scholar] [CrossRef] [PubMed]
- Alba, C.; Blanco, A.; Alarcón, T. Antibiotic resistance in Helicobacter pylori. Curr. Opin. Infect. Dis. 2017, 35, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Liou, J.M.; Lee, Y.C.; Hong, T.C.; El-Omar, E.M.; Wu, M.S. The interplay between Helicobacter pylori and gastrointestinal microbiota. Gut Microbes. 2021, 13, 1909459. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, Y.; Hu, J.; Wang, X.; Ren, M.; Lu, G.; Lu, X.; Zhang, D.; He, S. The effect of Helicobacter pylori eradication in patients with gastroesophageal reflux disease: A meta-analysis of randomized controlled studies. Dig. Dis. 2020, 38, 261–268. [Google Scholar] [CrossRef]
- Abou Baker, D. Plants against Helicobacter pylori to combat resistance: An ethnopharmacological review. Biotechnol. Rep. 2020, 26, e00470. [Google Scholar] [CrossRef] [PubMed]
- Silvan, J.M.; Gutiérrez-Docio, A.; Moreno-Fernandez, S.; Alarcón-Cavero, T.; Prodanov, M.; Martinez-Rodriguez, A.J. Procyanidin-Rich Extract from Grape Seeds as a Putative Tool against Helicobacter pylori. Foods 2020, 9, 1370. [Google Scholar] [CrossRef] [PubMed]
- Silvan, J.M.; Guerrero-Hurtado, E.; Gutiérrez-Docio, A.; Alarcón-Cavero, T.; Prodanov, M.; Martinez-Rodriguez, A.J. Olive-Leaf Extracts Modulate Inflammation and Oxidative Stress Associated with Human H. pylori Infection. Antioxidants 2021, 10, 2030. [Google Scholar] [CrossRef] [PubMed]
- Silvan, J.M.; Gutierrez-Docio, A.; Guerrero-Hurtado, E.; Domingo-Serrano, L.; Blanco-Suarez, A.; Prodanov, M.; Alarcon-Cavero, T.; Martinez-Rodriguez, A.J. Pre-Treatment with Grape Seed Extract Reduces Inflammatory Response and Oxidative Stress Induced by Helicobacter pylori Infection in Human Gastric Epithelial Cells. Antioxidants 2021, 10, 943. [Google Scholar] [CrossRef]
- Villalva, M.; Silvan, J.M.; Guerrero-Hurtado, E.; Gutierrez-Docio, A.; Navarro del Hierro, J.; Alarcón-Cavero, T.; Prodanov, M.; Martin, D.; Martinez-Rodriguez, A.J. Influence of In Vitro Gastric Digestion of Olive Leaf Extracts on Their Bioactive Properties against H. pylori. Foods 2022, 11, 1832. [Google Scholar] [CrossRef]
- Kusters, J.G.; van Vliet, A.H.; Kuipers, E.J. Pathogenesis of Helicobacter pylori infection. Clin. Microbiol. Rev. 2006, 19, 449–490. [Google Scholar] [CrossRef]
- Garcia-Oliveira, P.; Barral, M.; Carpena, M.; Gullón, P.; Fraga-Corral, M.; Otero, P.; Prieto, M.A.; Simal-Gandara, J. Traditional plants from Asteraceae family as potential candidates for functional food industry. Food Funct. 2021, 12, 2850–2873. [Google Scholar] [CrossRef]
- Gaweł-Bęben, K.; Strzępek-Gomółka, M.; Czop, M.; Sakipova, Z.; Głowniak, K.; Kukula-Koch, W. Achillea millefolium L. and Achillea biebersteinii Afan. hydroglycolic extracts–bioactive ingredients for cosmetic use. Molecules 2020, 25, 3368. [Google Scholar] [CrossRef] [PubMed]
- Rakmai, J.; Cheirsilp, B.; Torrado-Agrasar, A.; Simal-Gándara, J.; Mejuto, J.C. Encapsulation of yarrow essential oil in hydroxypropyl-beta-cyclodextrin: Physiochemical characterization and evaluation of bio-efficacies. CyTA-J. Food 2017, 15, 409–417. [Google Scholar] [CrossRef]
- Vitas, J.S.; Cvetanović, A.D.; Mašković, P.Z.; Švarc-Gajić, J.V.; Malbaša, R.V. Chemical composition and biological activity of novel types of kombucha beverages with yarrow. J. Funct. Foods 2018, 44, 95–102. [Google Scholar] [CrossRef]
- Barda, C.; Grafakou, M.E.; Tomou, E.M.; Skaltsa, H. Phytochemistry and Evidence-Based Traditional Uses of the Genus Achillea L.: An Update (2011–2021). Sci. Pharm. 2021, 89, 50. [Google Scholar] [CrossRef]
- Mainka, M.; Czerwińska, M.E.; Osińska, E.; Ziaja, M.; Bazylko, A. Screening of antioxidative properties and inhibition of inflammation-linked enzymes by aqueous and ethanolic extracts of plants traditionally used in wound healing in Poland. Antioxidants 2021, 10, 698. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.I.; Gopalakrishnan, B.; Venkatesalu, V. Pharmacognosy, phytochemistry and pharmacological properties of Achillea millefolium L.: A review. Phytother. Res. 2017, 31, 1140–1161. [Google Scholar] [CrossRef] [PubMed]
- Mouhid, L.; Gómez de Cedrón, M.; Quijada-Freire, A.; Fernández-Marcos, P.J.; Reglero, G.; Fornari, T.; Ramírez de Molina, A. Yarrow supercritical extract ameliorates the metabolic stress in a model of obesity induced by high-fat diet. Nutrients 2019, 12, 72. [Google Scholar] [CrossRef]
- Pereira, J.M.; Peixoto, V.; Teixeira, A.; Sousa, D.; Barros, L.; Ferreira, I.C.F.R.; Vasconcelos, M.H. Achillea millefolium L. hydroethanolic extract inhibits growth of human tumor cells lines by interfering with cell cycle and inducing apoptosis. Food Chem. Toxicol. 2018, 118, 635–644. [Google Scholar] [CrossRef]
- Vidović, S.; Vasić, A.; Vladić, J.; Jokić, S.; Aladić, K.; Gavarić, A.; Nastić, N. Carbon dioxide supercritical fluid extracts from yarrow and rose hip herbal dust as valuable source of aromatic and lipophilic compounds. Sustain. Chem. Pharm. 2021, 22, 100494. [Google Scholar] [CrossRef]
- Farhadi, N.; Babaei, K.; Farsaraei, S.; Moghaddam, M.; Pirbalouti, A.G. Changes in essential oil compositions, total phenol, flavonoids and antioxidant capacity of Achillea millefolium at different growth stages. Ind. Crops. Prod. 2020, 152, 112570. [Google Scholar] [CrossRef]
- Salehi, B.; Selamoglu, Z.; Sevindik, M.; Fahmy, N.M.; Al-Sayed, E.; El-Shazly, M.; Büsselberg, D. Achillea spp.: A comprehensive review on its ethnobotany, phytochemistry, phytopharmacology and industrial applications. Cell. Mol. Biol. 2020, 66, 78–103. [Google Scholar] [CrossRef] [PubMed]
- Salomon, L.; Lorenz, P.; Bunse, M.; Spring, O.; Stintzing, F.C.; Kammerer, D.R. Comparison of the Phenolic Compound Profile and Antioxidant Potential of Achillea atrata L. and Achillea millefolium L. Molecules 2021, 26, 1530. [Google Scholar] [CrossRef] [PubMed]
- Villalva, M.; Santoyo, S.; Salas-Pérez, L.; Siles-Sánchez, M.D.L.N.; Rodríguez García-Risco, M.; Fornari, T.; Reglero, G.; Jaime, L. Sustainable extraction techniques for obtaining antioxidant and anti-inflammatory compounds from the Lamiaceae and Asteraceae species. Foods 2021, 10, 2067. [Google Scholar] [CrossRef]
- Strzępek-Gomółka, M.; Gaweł-Bęben, K.; Kukula-Koch, W. Achillea species as sources of active phytochemicals for dermatological and cosmetic applications. Oxid. Med. Cell. Long. 2021, 2021, 6643827. [Google Scholar] [CrossRef]
- Abdossi, V.; Kazemi, M. Bioactivities of Achillea millefolium essential oil and its main terpenes from Iran. Int. J. Food Prop. 2016, 19, 1798–1808. [Google Scholar] [CrossRef]
- Verma, R.S.; Joshi, N.; Padalia, R.C.; Goswami, P.; Singh, V.R.; Chauhan, A.; Darokar, M.P. Chemical composition and allelopathic, antibacterial, antifungal and in vitro acetylcholinesterase inhibitory activities of yarrow (Achillea millefolium L.) native to India. Ind. Crops. Prod. 2017, 104, 144–155. [Google Scholar] [CrossRef]
- Mahady, G.B.; Pendland, S.L.; Stoia, A.; Hamill, F.A.; Fabricant, D.; Dietz, B.M.; Chadwick, L.R. In vitro susceptibility of Helicobacter pylori to botanical extracts used traditionally for the treatment of gastrointestinal disorders. Phytother. Res. 2005, 19, 988–991. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, S.F.H.; Yamada, K.; Kadowaki, M.; Usmanghani, K.; Sugiyama, T. Bactericidal activity of medicinal plants, employed for the treatment of gastrointestinal ailments, against Helicobacter pylori. J. Ethnopharmacol. 2009, 121, 286–291. [Google Scholar] [CrossRef]
- Gutierrez-Docio, A.; Almodóvar, P.; Moreno-Fernandez, S.; Silvan, J.M.; Martinez-Rodriguez, A.J.; Alonso, G.L.; Prodanov, M. Evaluation of an integrated ultrafiltration/solid phase extraction process for purification of oligomeric grape seed procyanidins. Membranes 2020, 10, 147. [Google Scholar] [CrossRef] [PubMed]
- Villalva, M.; Jaime, L.; Aguado, E.; Nieto, J.A.; Reglero, G.; Santoyo, S. Anti-inflammatory and antioxidant activities from the basolateral fraction of Caco-2 cells exposed to a rosmarinic acid enriched extract. J. Agric. Food Chem. 2018, 66, 1167–1174. [Google Scholar] [CrossRef] [PubMed]
- Quintana, S.E.; Hernández, D.M.; Villanueva-Bermejo, D.; García-Risco, M.R.; Fornari, T. Fractionation and precipitation of licorice (Glycyrrhiza glabra L.) phytochemicals by supercritical antisolvent (SAS) technique. LWT 2020, 126, 109315. [Google Scholar] [CrossRef]
- Villanueva-Bermejo, D.; Zahran, F.; García-Risco, M.R.; Reglero, G.; Fornari, T. Supercritical fluid extraction of Bulgarian Achillea millefolium. J. Supercrit. Fluids 2017, 119, 283–288. [Google Scholar] [CrossRef]
- Gil-Ramírez, A.; Rodriguez-Meizoso, I. Purification of Natural Products by Selective Precipitation Using Supercritical/Gas Antisolvent Techniques (SAS/GAS). Sep. Purif. Rev. 2019, 50, 32–52. [Google Scholar] [CrossRef]
- Villalva, M.; Jaime, L.; Villanueva-Bermejo, D.; Lara, B.; Fornari, T.; Reglero, G.; Santoyo, S. Supercritical anti-solvent fractionation for improving antioxidant and anti-inflammatory activities of an Achillea millefolium L. extract. Food Res. Int. 2019, 115, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Judzentiene, A.; Mockutë, D. Chemical composition of essential oils produced by pink flower inflorescences of wild Achillea millefolium L. Chemija 2004, 15, 28–32. [Google Scholar]
- Bimbiraitė, K.; Ragažinskienė, O.; Maruška, A.; Kornyšova, O. Comparison of the chemical composition of four yarrow (Achillea millefolium L.) morphotypes. Biologija 2008, 54, 208–212. [Google Scholar] [CrossRef]
- Falconieri, D.; Piras, A.; Porcedda, S.; Marongiu, B.; Gonçalves, M.J.; Cabral, C.; Cavaleiro, C.; Salgueiro, L. Chemical composition and biological activity of the volatile extracts of Achillea millefolium. Nat. Prod. Commun. 2011, 6, 1527–1530. [Google Scholar] [CrossRef] [PubMed]
- Marzouki, H.; Piras, A.; Porcedda, S.; Falconieri, D.; Bagdonaite, E. Influence of extraction methods on the composition of essential oils of Achillea millefolium L. from Lithuania. J. Biodivers. Manag. For. 2014, 3, 1–4. [Google Scholar]
- Silvan, J.M.; Mingo, E.; Martinez-Rodriguez, A.J. Grape seed extract (GSE) modulates Campylobacter pro-inflammatory response in human intestinal epithelial cell lines. Food Agric. Immunol. 2017, 28, 739–753. [Google Scholar] [CrossRef]
- Ivanović, M.; Grujić, D.; Cerar, J.; Islamčević Razboršek, M.; Topalić-Trivunović, L.; Savić, A.; Kočar, D.; Kolar, M. Extraction of Bioactive Metabolites from Achillea millefolium L. with Choline Chloride Based Natural Deep Eutectic Solvents: A Study of the Antioxidant and Antimicrobial Activity. Antioxidants 2022, 11, 724. [Google Scholar] [CrossRef] [PubMed]
- Caporali, S.; De Stefano, A.; Calabrese, C.; Giovannelli, A.; Pieri, M.; Savini, I.; Tesauro, M.; Bernardini, S.; Minieri, M.; Terrinoni, A. Anti-Inflammatory and Active Biological Properties of the Plant-Derived Bioactive Compounds Luteolin and Luteolin 7-Glucoside. Nutrients 2022, 14, 1155. [Google Scholar] [CrossRef] [PubMed]
- Zha, R.P.; Xu, W.; Wang, W.Y.; Dong, L.; Wang, Y.P. Prevention of lipopolysaccharide-induced injury by 3,5-dicaffeoylquinic acid in endothelial cells. Acta Pharmacol. Sin. 2007, 28, 1143–1148. [Google Scholar] [CrossRef]
- Pérez-Torres, I.; Castrejón-Téllez, V.; Soto, M.E.; Rubio-Ruiz, M.E.; Manzano-Pech, L.; Guarner-Lans, V. Oxidative Stress, Plant Natural Antioxidants, and Obesity. Int. J. Mol. Sci. 2021, 22, 1786. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.; Deb, P.K.; Priya, S.; Medina, K.D.; Devi, R.; Walode, S.G.; Rudrapal, M. Dietary Flavonoids: Cardioprotective Potential with Antioxidant Effects and Their Pharmacokinetic, Toxicological and Therapeutic Concerns. Molecules 2021, 26, 4021. [Google Scholar] [CrossRef]
- Speisky, H.; Shahidi, F.; Costa de Camargo, A.; Fuentes, J. Revisiting the Oxidation of Flavonoids: Loss, Conservation or Enhancement of Their Antioxidant Properties. Antioxidants 2022, 11, 133. [Google Scholar] [CrossRef] [PubMed]
- De Stefano, A.; Caporali, S.; Di Daniele, N.; Rovella, V.; Cardillo, C.; Schinzari, F.; Minieri, M.; Pieri, M.; Candi, E.; Bernardini, S. Anti- Inflammatory and Proliferative Properties of Luteolin-7-O-Glucoside. Int. J. Mol. Sci. 2021, 22, 1321. [Google Scholar] [CrossRef] [PubMed]
- Marengo, A.; Fumagalli, M.; Sanna, C.; Maxia, A.; Piazza, S.; Cagliero, C.; Rubiolo, P.; Sangiovanni, E.; Dell’Agli, M. The hydro-alcoholic extracts of Sardinian wild thistles (Onopordum spp.) inhibit TNF alpha-induced IL-8 secretion and NF-kappa B pathway in human gastric epithelial AGS cells. J. Ethnopharmacol. 2018, 210, 469–476. [Google Scholar] [CrossRef]
- El-Sherbiny, G.M.; Elbestawy, M.K. A review–plant essential oils active against Helicobacter pylori. J. Essent. Oil Res. 2022, 34, 203–215. [Google Scholar] [CrossRef]
- Bergonzelli, G.E.; Donnicola, D.; Porta, N.; Corthésy-Theulaz, I.E. Essential oils as components of a diet-based approach to management of Helicobacter infection. Antimicrob. Agents Chemother. 2003, 47, 3240–3246. [Google Scholar] [CrossRef] [PubMed]
- Eftekhar, F.; Nariman, F.; Yousefzadi, M.; Hadian, J.; Ebrahimi, S.N. Anti-Helicobacter pylori activity and Essential Oil Composition of Thymus caramanicus from Iran. Nat. Prod. Commun. 2009, 4, 1139–1142. [Google Scholar] [CrossRef] [PubMed]
- Jeong, M.; Park, J.M.; Han, Y.M.; Kangwan, N.; Kwon, S.O.; Kim, B.N.; Kim, W.H.; Hahm, K.B. Dietary intervention of Artemisia and Green Tea extracts to rejuvenate Helicobacter pylori-associated chronic atrophic gastritis and to prevent tumorigenesis. Helicobacter 2016, 21, 40–59. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Phenolic Compounds | YE | YPF | YSF |
|---|---|---|---|
| Phenolic acids | |||
| Hydroxycinnamic acids | |||
| Caffeic acid 1 | 17.4 ± 0.1 a | - | 18.4 ± 0.1 b |
| Caftaric acid 1 | <L.Q. | 22.5 ± 0.3 | - |
| Chlorogenic acid 1 | 61.7 ± 0.2 a | 190.7 ± 4.1 b | - |
| Cryptochlorogenic acid 1 | 1.1 ± 0.1 a | 4.4 ± 1.4 a | - |
| 1,5- DCQA 1 | 68.7 ± 0.7 a | 179.7 ± 8.7 b | - |
| 3,4- DCQA 1 | 38.3 ± 5.1 a | 69.1 ± 0.5 b | - |
| 3,5- DCQA 1 | 361.7 ± 1.8 b | 1163.4 ± 10.2 c | 10.4 ± 0.1 a |
| 4,5- DCQA 1 | 96.6 ± 0.9 a | 318.7 ± 0.2 b | - |
| Ferulic acid 1 | 7.9 ± 1.7 b | 4.3 ± 0.1 a | - |
| Neochlorogenic acid 1 | 5.8 ± 0.1 a | 13.8 ± 0.14 b | - |
| Rosmarinic acid 1 | 185.0 ± 1.2 | - | - |
| Σ Total Phenolic acids | 844.2 b | 1966.6 c | 28.8 a |
| Flavonoids | |||
| Flavones | |||
| Amentoflavone | 41.9 ± 0.1 a | 41.2 ± 0.1 a | 62.2 ± 0.1 b |
| Apigenin 1 | 195.7 ± 0.4 b | 474.2 ± 0.4 c | 92.5 ± 0.1 a |
| Apigenin-C-hexoside-C-pentoside | 30.3 ± 0.1 a | 84.7 ± 0.2 b | - |
| Apigenin-7-O-glucoside 1 | 179.5 ± 0.8 a | 587.7 ± 1.3 b | - |
| Diosmetin 1 | 50.1 ± 0.1 a | 72.8 ± 0.1 b | 50.2 ± 0.1 a |
| Homoorientin 1 | 2.5 ± 0.7 a | 15.5 ± 0.1 b | - |
| 6-Hydroxyluteolin-7-O-glucoside | 145.2 ± 0.7 a | 466.2 ± 0.4 b | - |
| Luteolin 1 | 447.4 ± 1.2 b | 1304.0 ± 10.0 c | 95.5 ± 1.2 a |
| Luteolin-6,8-di-C-glucoside | 46.9 ± 0.2 a | 151.1 ± 0.1 b | - |
| Luteolin-7-β-glucuronide 1 | 19.9 ± 1.1 a | 59.3 ± 2.8 b | - |
| Luteolin-7-O-glucoside 1 | 768.7 ± 8.0 a | 2385.3 ± 97.5 b | - |
| Schaftoside 1 | 27.2 ± 0.1 a | 88.8 ± 0.4 b | - |
| Schaftoside isomer | 26.2 ± 0.3 a | 89.6 ± 0.2 b | - |
| Vicenin 2 1 | 36.6 ± 0.5 a | 111.7 ± 0.6 b | - |
| Σ Total Flavones | 2018.1 b | 5932.1 c | 300.4 a |
| Flavonols | |||
| Casticin 1 | 28.6 ± 0.1 a | - | 61.8 ± 0.6 b |
| Centaureidin | 391.3 ± 0.4 a | 107.8 ± 0.1 b | 669.6 ± 0.4 c |
| Methoxyquercetin isomer | 376.0 ± 0.8 b | 751.3 ± 0.8 c | 265.0 ± 0.2 a |
| Quercetin 1 | 47.0 ± 0.1 a | 143.6 ± 0.8 b | - |
| Rutin 1 | 50.6 ± 1.0 a | 133.7 ± 2.2 b | - |
| Vitexin 1 | 12.8 ± 0.8 a | 24.8 ± 0.7 b | - |
| Σ Total Flavonols | 906.3 a | 1161.2 c | 996.4 b |
| Σ Total Flavonoids | 2924.4 b | 7093.3 c | 1296.8 a |
| Σ Total phenolic compounds | 3768.6 b | 9060.0 c | 1325.7 a |
| Rt | Compound | YE | YSF |
|---|---|---|---|
| 11.9 | Yomogi alcohol | 2.5 | 2.7 |
| 13.1 | Eucalyptol | 5.2 | 4.2 |
| 13.5 | γ-Vinyl-γ-valerolactone | 2.1 | 1.8 |
| 14.6 | Artemisia ketone | 13.3 | 11.5 |
| 18.4 | Camphor | 16.7 | 15.0 |
| 19.5 | Borneol | 10.5 | 10.5 |
| 20.7 | 3,7-dimethyl-1,5-Octadiene-3,7-diol | 7.1 | 7.5 |
| 22.8 | trans-Chrysanthenyl acetate | 2.9 | 3.0 |
| 24.3 | (5E)-5,9-dimethyl-5,8-decadien-2-one | 1.8 | 1.6 |
| 24.7 | 2,6-dimethyl-1,7-octadiene-3,6-diol | 10.5 | 11.3 |
| 26.0 | N.i. | 8.7 | 10.4 |
| 30.3 | Jasmone | 1.9 | 1.8 |
| 31.2 | β-Caryophyllene | 1.7 | 1.9 |
| 38.2 | β-Caryophyllene oxide | 6.0 | 7.0 |
| 41.7 | β-Eudesmol | 1.5 | 1.9 |
| 51.8 | Saussurea lactone | 4.1 | 3.9 |
| 52.8 | Hexahydrofarnesyl acetone | 3.6 | 4.2 |
| ƩAUC | 23.6 × 106 | 43.8 × 106 |
| YE | YPF | YSF | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Strains | Control Growth | CFU/mL | Log CFU Reduction | MIC (mg/mL) | CFU/mL | Log CFU Reduction | MIC (mg/mL) | CFU/mL | Log CFU Reduction | MIC (mg/mL) |
| Hp48 | 3.6 ± 0.3 × 108 c,A | <1.0 × 102 a,A | >6.3 | 0.14 | 4.0 ± 0.7 × 105 b,A | 2.4 | 0.08 | <1.0 × 102 A,a | >6.3 | 0.08 |
| Hp53 | 3.4 ± 1.6 × 108 d,A | 5.0 ± 1.5 × 103 b,B | 4.8 | 0.14 | 6.5 ± 1.3 × 105 c,A | 2.2 | 0.14 | <1.0 × 102 a,A | >6.8 | 0.08 |
| Hp59 | 9.9 ± 0.9 × 108 d,A | 1.4 ± 0.2 × 102 b,A | 7.1 | 0.14 | 4.8 ± 1.6 × 106 c,B | 2.4 | 0.14 | <1.0 × 102 a,A | >7.1 | 0.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villalva, M.; Silvan, J.M.; Alarcón-Cavero, T.; Villanueva-Bermejo, D.; Jaime, L.; Santoyo, S.; Martinez-Rodriguez, A.J. Antioxidant, Anti-Inflammatory, and Antibacterial Properties of an Achillea millefolium L. Extract and Its Fractions Obtained by Supercritical Anti-Solvent Fractionation against Helicobacter pylori. Antioxidants 2022, 11, 1849. https://doi.org/10.3390/antiox11101849
Villalva M, Silvan JM, Alarcón-Cavero T, Villanueva-Bermejo D, Jaime L, Santoyo S, Martinez-Rodriguez AJ. Antioxidant, Anti-Inflammatory, and Antibacterial Properties of an Achillea millefolium L. Extract and Its Fractions Obtained by Supercritical Anti-Solvent Fractionation against Helicobacter pylori. Antioxidants. 2022; 11(10):1849. https://doi.org/10.3390/antiox11101849
Chicago/Turabian StyleVillalva, Marisol, Jose Manuel Silvan, Teresa Alarcón-Cavero, David Villanueva-Bermejo, Laura Jaime, Susana Santoyo, and Adolfo J. Martinez-Rodriguez. 2022. "Antioxidant, Anti-Inflammatory, and Antibacterial Properties of an Achillea millefolium L. Extract and Its Fractions Obtained by Supercritical Anti-Solvent Fractionation against Helicobacter pylori" Antioxidants 11, no. 10: 1849. https://doi.org/10.3390/antiox11101849