
Abstract
Ombrotrophic bogs receive new inputs of elements solely through atmospheric deposition, except for N where inputs are predominantly through N2-fixation, at least in low N deposition environments. At various locations across the globe, including the Athabasca Oil Sands Region (AOSR) of northern Alberta, Canada, element atmospheric deposition has increased as a result of anthropogenic activities. Regional and/or global deposition gradients offer an opportunity to examine questions related to nutrient limitation and element stoichiometry, i.e., the maintenance of relatively constant element ratios in bog lichen/plant tissues despite differing element deposition/availability. Using a dataset of tissue element concentrations in eight lichen/plant species in six AOSR bogs, supplemented with literature data from other sites globally, this synthesis asks: is there evidence of element stoichiometric homeostasis in lichen or plant species in AOSR bogs; if so, do stoichiometric homeostasis relationships extend globally beyond the AOSR, and; do element ratios provide insight into element limitation for the eight species? Mean element ratios and their coefficients of variation, ternary NPK and CaMgK plots, and scaling coefficients revealed widespread evidence of stoichiometric homeostasis. Stoichiometric relationships generally were unaffected by differences in element deposition among the AOSR bogs. Stoichiometric relationships sometimes extended to a species globally, but sometimes did not. Element ratios and ternary diagrams suggested a combination of N-, P-, and K-limitation, both within and beyond the AOSR bogs. Regionally high atmospheric N deposition may have shifted some species from N-limitation prior to the Industrial Revolution to P- or K-limitation today.
Similar content being viewed by others

Introduction
Liebig’s Law of the Minimum, a pillar of soil science and plant nutrition, initially was formulated by Carl Sprengel, who wrote “… it is indisputable that, when a plant needs 12 substances to develop, it will not grow if any one of these is missing, and it will always grow poorly, when one of these is not available in sufficiently large amount as required by the nature of the plant” (van der Ploeg et al. 1999). If plant growth is limited by a low supply of a particular element, growth will increase as the supply of that limiting nutrient increases. The growth response to increasing supply of a limiting element generally may be more pronounced in plants adapted to high nutrient environments than in plants adapted to low nutrient environments (Chapin et al. 1986), like bogs. Further, the growth response is not linear; as the supply of a limiting element increases to the point where plant acquisition exceeds the need for growth, element concentrations in plant tissues may increase, reflecting element storage (cf. Chapin et al. 1990; Lamers et al. 2000). Plant tissue element concentrations do not increase without bound, but rather, as supply increases, ultimately concentrations reach maximum values. An increasing supply of non-limiting elements also can result in increased plant uptake and increased element concentrations in plant tissues, reflecting storage (Chapin et al. 1990). These responses to increasing element availability have implications for assessing the extent to which individual plant species exert stoichiometric control through the examination of element ratios.
If plant acquisition of a particular element in response to variation in availability in soil/peat occurs independently of other elements, plant tissue element ratios would exhibit corresponding variability. However, if a particular plant species exerts stoichiometric control over multi-element uptake and storage, element ratios should be relatively constant, regardless of availability in soil/peat; i.e., there is stoichiometric homeostasis (sensu Sterner and Elser 2002). A literature survey of nutrient concentrations in terrestrial plants (Knecht and Göransson 2004) indicated that there are strong correlations between pairs of elements (N, P, K, Ca, Mg) in plant tissues. Focusing on C, N, P and K, Wang and Moore (2014) concluded that plant functional types (mosses, deciduous trees/shrubs, evergreen shrubs, graminoids, forbs) at Mer Bleue Bog, Ontario, exhibit strong stoichiometric homeostasis. Similarly, Wang et al. (2016) concluded that shrubs, and to a lesser extent mosses, exhibited homeostatic regulation in response to N, P, and K fertilization at Mer Bleue Bog.
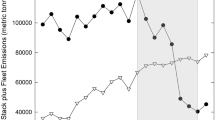
The Athabasca Oil Sands Region (AOSR) in northern Alberta, Canada offers a unique opportunity to assess lichen and plant element tissue concentrations and ratios with respect to stoichiometric homeostasis in bogs. Bogs are ombrotrophic, receiving new inputs of elements solely through atmospheric deposition (Weber 1911; Vitt 2006; Vitt and House 2021), with the notable exception of N where inputs are predominantly through N2-fixation by methanotrophs (Vile et al. 2014), at least in the low N deposition environments of northern Alberta. Increasing N deposition downregulates N2-fixation in bogs (Wieder et al. 2019) and in poor fens (Wieder et al. 2020). In the AOSR, upgrader stack and mine fleet emissions of N and S gases, and dispersion of fugitive dust with high Ca and Mg concentrations, have altered element deposition, with N, S, Ca, and Mg wet and/or dry deposition generally decreasing with increasing distance from the oil sands industrial center (Fenn et al. 2015; Wieder et al. 2016a,b; Hsu et al. 2016; Edgerton et al. 2020; Horb et al. 2021; Wieder et al. 2022; Table S1, Figure S1). Further, P and K deposition appears to vary spatially across the AOSR, although deposition patterns are not clearly associated with oil sands operations (Wieder et al. 2022, Table S1, Figure S1).
Given the ombrotrophic nature of bogs, spatial patterns of element deposition are especially likely to reflect spatial patterns of element availability in peat. Indeed, tissue concentrations of N, S, Ca, Mg, K, and P in several lichen/plant species increase with proximity to the oil sands industrial center and/or are correlated with regional spatial patterns in element deposition (Wieder et al. 2016a, 2021, 2022). If these lichen/plant species exhibit stoichiometric homeostasis, element ratios in lichen thalli, Sphagnum capitula, and leaves of vascular plants should remain relatively constant across bogs in the AOSR, regardless of distance from the oil sands industrial center.
Here, stoichiometric relationships for eight common Alberta bog species, the lichens Cladonia (Cladina) mitis and Evernia mesomorpha, the mosses Sphagnum capillifolium and Sphagnum fuscum, and the vascular plants Vaccinium oxycoccos, Vaccinium vitis-idaea, Rhododendron groenlandicum (Ledum), and Picea mariana, are evaluated using three approaches. First, for each species, element ratios and their coefficients of variation were calculated, providing insight into potential nutrient limitation (ratios) and stoichiometric control (coefficients of variation) (e.g., Koerselman and Meuleman 1996; Güsewell and Koerselman 2002; McGroddy et al. 2004; Jiroušek et al. 2011; Wang and Moore 2014). Second, ternary diagrams were constructed to examine stoichiometric relationships between groups of three elements (N, P, K and Ca, Mg, K), as well as to assess potential nutrient limitation (Güsewell and Koerselman 2002; Olde Venterink et al. 2003; Wang and Moore 2014). Third, stoichiometric relationships between multiple elements were assessed using the niche volume approach of Ågren and Weih (2020). These approaches were applied to tissue chemistry data from eight lichen/plant species in bogs in the AOSR (Wieder et al. 2016a, 2021, 2022). In addition, to achieve a broader geographic perspective, where available, literature tissue chemistry data for these eight species, regardless of the ecosystem in which they were found, were included in analyses.
This analysis focuses on three questions: (1) is there evidence of element stoichiometric homeostasis in the eight lichen/plant species across the six bogs in the AOSR of northern Alberta, Canada; (2) if there is evidence of stoichiometric homeostasis in the AOSR, does it extend to all locations where a species grows globally, and (3) do element ratios provide insight into what element is limiting for each of the eight species?
Methods
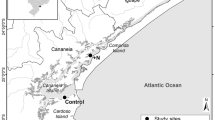
Lichen/plant samples were collected at six bog sites in northern Alberta, Canada, ranging from 12 to 77 km from the oil sands industrial center, operationally defined as the midpoint between the Syncrude and Suncor upgrader stacks (JPH4, 57°6′45″N, 111°25′23″W, 12 km; McKay, 57°13′41″N, 111°42′11″W, 24 km; Kearl, 57°16′21″N, 111°15′38″W, 32 km; McMurray, 56°37′37″N, 111°11′44″W, 49 km; Anzac, 56°28′8″N, 111°2′34″W, 69 km; Horse Creek, 56°19′45″N, 111°35′22″W, 77 km). Vegetation is similar at these sites (Vitt et al. 2020) and is typical of Alberta bogs more generally (Belland and Vitt 1995). Five replicate lichen/plant samples (whole thalli of the lichens Evernia mesomorpha and Cladonia (Cladina) mitis; the capitula (top 1-cm of plant) of the mosses Sphagnum capillifolium and Sphagnum fuscum; aboveground portions of Vaccinium oxycoccos with leaves separated from stems in the laboratory, leaves retained for analysis; topmost 3–5 leaves from individual plants of Vaccinium vitis-idaea and Rhododendron (Ledum) groenlandicum, and apical shoots of current year’s growth of Picea mariana, needles separated from stems and retained for analysis), were collected at each site in June, July, and August of 2018 and 2019 (see Wieder et al. 2022 for details of sample collection, processing, and analysis for Ca, Mg, K, and P; concentrations of N and S were determined on a LECO Truspec CNS Analyzer, cf. Wieder et al. 2021).
Our group has quantified N, S, Ca, Mg, and P deposition using ion exchange resin (IER) collectors deployed in bogs across northern Alberta since 2009 (Wieder et al. 2016a,b, 2021, 2022). Potassium deposition data are from IER collectors deployed at the bog sites by the Wood Buffalo Environmental Association since October 2018 (https://wbea.org/network-and-data/integrated-data-search/). The history of these measurements is summarized in Table S1; element deposition as a function of distance from the oil sands industrial center is provided in Figure S1.
The extant literature was searched for data on N, P, K, Ca, Mg, and S concentrations in the eight lichen/plant species sampled in the six AOSR bogs. Both Cladonia mitis and C. arbuscula were included as these two species cannot be clearly distinguished in the field; the major taxonomic distinguishing characteristic is the presence of rangiformic acid secondary compounds in the former, but not the latter (Ruoss 1987). Further, Vaccinium oxycoccos has several taxonomic synonyms, including Oxycoccos palustris; leaf elemental concentration data for O. palustris have been included. When published papers described a field manipulative experiment, only data from control treatments have been included. Data from greenhouse studies and data for vascular plant seedlings were excluded. Published papers that reported data on only one of the five elements were excluded. Data from different habitat types were included, with data from bog sites being indicated, when possible.
For data on element concentrations/ratios in the eight plant/lichen species in six bogs in the AOSR, stoichiometric homeostasis was examined by calculating coefficients of variation of the lognormal distributions of tissue concentrations. Following Güsewell and Koerselman (2002), all data were log-transformed prior to calculating coefficients of variation as:
where x̄ and SD are the mean and standard deviation, respectively, of the log-transformed data for a given variable. For each species/site combination, \(\overline{x}\) and SD were calculated for the six sampling dates combined (July, July, August of 2018 and 2019; only 3 sampling dates were used for Sphagnum capillifolium as there was insufficient sample for chemical analysis in 2018).
Using our data from the six bogs in the AOSR, and literature data where raw element concentration data on individual samples (as opposed to means or medians) were available, for each species, ternary diagrams were constructed to graphically indicate relationships between N, P and K, and between Ca, Mg, and K. For the N, P, and K ternary diagrams, N:P, N:K, and K:P ratios of 14.5, 2.1 and 3.4, respectively, were used as critical values to identify N, P, and/or K-limitation of species (Olde Venterink et al. 2003). For Ca, Mg, and K ternary diagrams, such critical ratios for bog species have not been developed.
Following Ågren and Weih (2020), stoichiometric volumes were calculated for each of the eight lichen/plant species and for the six AOSR bog sites individually and collectively, assuming that N and P are the two elements that drive stoichiometric relationships. Two volumes, assumed to represent realized niche subvolumes of a species (V) were calculated as the product of element concentrations in lichen/plant tissues:
and
where CN, Cp, and Cn are lichen/plant tissue concentrations of N, P, and n other elements, respectively. Scaling between these two subvolumes was calculated as:
which log-transforms into a linear equation:
where α is the scaling coefficient and β is the intercept. Scaling relationships between \({V}_{NP}\) and \({V}_{Oth}\) were calculated with \({V}_{Oth}\) representing Ca, Mg, K, and S in combination and individually. Regression parameters (α and β) were estimated using reduced major axis regression. A scaling coefficient of 1 indicates that \({V}_{NP}\) and \({V}_{Oth}\) increase proportionally, indicating stoichiometric homeostasis. A scaling coefficient < 1 indicates that concentrations of other elements increase more slowly than concentrations of N and P; a scaling coefficient > 1 indicates that concentrations of other elements increase more rapidly than concentrations of N and P. Either of these situations indicates that element ratios do not remain constant as N and P concentrations change, which would indicate a lack of stoichiometric homeostasis with respect to N and P in combination. Nonsignificant regressions indicate an absence of scaling, and hence no stoichiometric homeostasis.
Literature survey
A search of the literature located 112 published papers or data sets where tissue concentrations of at least two of the elements (N, P, K, Ca, Mg, S) in one or more of the eight lichen/plant species that are the focus of this paper were reported (Table S2). Several of these papers reported means and standard deviations, standard errors, or confidence intervals, but did not provide raw data. Given that the ratio of two means is not mathematically equivalent to the mean of ratios of individual samples, papers that did not include raw data were excluded from analysis unless authors provided their raw data. As such, 41 published papers (Tables S3, S4) were included in analyses of element ratios, coefficients of variation, ternary diagrams, and niche volume approaches to assessing element stoichiometry and nutrient limitation.
Results and discussion
Element ratios and coefficients of variation
In general, across all species and all element ratios, there were differences between at least two of the six bog sites in the AOSR, the exceptions being for Ca:P ratios in Cladonia mitis, for N:K, N:P, and K:P ratios in Sphagnum capillifolium capitula, and for N:Ca and K:Ca ratios in Vaccinium vitis-idaea leaves, which did not differ between sites (Table S3). While statistical analysis provides an objective assessment of stoichiometric homeostasis in element ratios, the extent to which it provides a biologically meaningful assessment is unclear; studies claiming ecologically meaningful element stoichiometries have used a more subjective approach (e.g., Knecht and Göransson 2004; Wang and Moore 2014). For the six bogs in the AOSR, site differences in element ratios, although often statistically significant, often were small in magnitude (Fig. 1, Table S3). Using a subjective rather than statistical approach to assess stoichiometric homeostasis, for 74 of the 120 species/ratio combinations, mean values for each of the six bogs were within 25% of the mean value averaged across all six AOSR bog sites (Fig. 1, Table S3). This 25% criterion was observed in all eight species for K:P ratios and in six or seven of the species for N:K, N:S, N:P, K:S, Ca:S and Ca:Mg ratios, but in only two, three, or four of the species for N:Ca, N:Mg, Ca:P, S:Mg, S:P, or Mg:P ratios. Overall, these patterns suggest a greater degree of stoichiometric homogeneity with respect to N, P, and K, the three elements most commonly considered to be limiting production in bogs (e.g., Aerts et al. 1992; Gunnarsson and Rydin 2000; Hoosbeek et al. 2002; Bragazza et al. 2004; Phuyal et al. 2008; Jiroušek et al. 2011), than for Ca, Mg, or S.

Mean element ratios for eight plant/lichen species (CM Cladonia mitis, EM Evernia mesomorpha, SC Sphagnum capillifolium, SF Sphagnum fuscum, VO Vaccinium oxycoccos, VV Vaccinium vitis-idaea, PG Rhododendron groenlandicum, PM Picea mariana) at the 6 bog sites in the AOSR, averaged across all sampling dates (this study), arranged with increasing distance from the oil sands industrial center from left to right. Also shown are mean element ratios for the 8 plant lichen species at sites other than the 6 bogs in the AOSR (All sites other than this study) and for all sites combined. For a given element ratio/species combination, rectangles show the range of element ratio values that are within 20% ( ±) of the mean for the 6 bog sites in the AOSR. Green-filled rectangles indicate that the individual mean values for each of the 6 bog sites in the AOSR all fall within 20% of the mean value for the 6 bog sites in the AOSR; red-filled rectangles indicate that the individual mean values for each of the 6 bog sites in the AOSR do not all fall within 20% of the mean value for the 6 bog sites in the AOSR. Data, statistical results, and references are in Table S3
If indeed there is greater stoichiometric homogeneity for N, P, and K, than for other elements, coefficients of variation would be expected to be relatively low for N:K, N:P, and K:P ratios as compared to ratios involving Ca, Mg, or S. Such a pattern is evident for element ratio coefficients of variation across the plant/lichen species at the 6 bogs in the AOSR, where median ranks of the within-species N:K, N:P, and K:P coefficients of variation were 3.5, 5, and 3.5 out of 15, respectively (Table 1). This pattern is not as clearly evident in all of the eight species for sites beyond the 6 bogs in the AOSR, but it should be noted that the sample sizes vary considerably between sites and between species for the non-AOSR sites (Table S4). As sample size decreases, in general, the calculated coefficient of variation becomes a less accurate estimate of the underlying population coefficient of variation (cf. Kelley 2007). Thus, interpretations about coefficients of variation in element ratios can be made with a higher degree of confidence for the 6 bogs in the AOSR, where sample sizes are generally comparable and large, than for the other sites globally.
Given their lack of roots and of leaves with waxy cuticles, lichens and Sphagnum species could be expected to exhibit a lesser degree of stoichiometric homeostasis than vascular plants. However, the evidence for this is mixed. For the 15 pairwise element ratios across the six AOSR bog sites (Table S3) the 25% criterion was observed 10 and 4 times in the lichens Cladonia mitis and Evernia mesomorpha, respectively, 6 times each in Sphagnum capillifolium and Sphagnum fuscum, and 15, 15, 6, and 12 times for Vaccinium oxycoccos, Vaccinium vitis-idaea, Rhododendron groenlandicum, and Picea mariana, respectively. In addition to having leaves with waxy cuticles, all of the vascular plant species are mycorrhizal; these characteristics may be important in maintaining element concentrations and/or ratios within relatively narrow ranges. All element ratios at each of the six AOSR bog sites were within 25% of the mean across all of the AOSR bogs for each of the two Vaccinium species, suggesting a relatively high level of stoichiometric homeostasis involving all six elements—N, P, K, Ca, Mg, and S. Compared to the other vascular plant species, the two Vaccinium species also showed relatively low coefficients of variation, especially for N:P, N:K, and K:P ratios (Table S3), providing support for the interpretation that these two species exhibit tight control over element ratios across the six AOSR bogs and beyond, i.e., stoichiometric homeostasis.
If stoichiometric homeostasis is a characteristic of a particular species, mean values for element ratios across the six Alberta bog sites should be comparable to values from other locations across the globe. In general, this was not the case. For 79 of the 100 species-by-element ratio combinations where there were literature data from other sites, mean values for the six Alberta sites were significantly different from the mean values for other sites around the globe (Table S3). For 51 of these 79 instances, however, the means for the Alberta sites and the other sites around the globe were within 25% of the overall mean value across all sites globally.
While these comparisons may suggest some degree of stoichiometric homeostasis for various element ratios in different species across all sites globally, several caveats need to be acknowledged. Most studies that have reported element ratios or have provided raw data from which element ratios could be calculated have rather small sample sizes, often with samples collected on a single sampling date. Element deposition at the time of sampling and/or historical patterns of element deposition often are not reported. If element concentrations vary over the course of a growing season, time of year in which samples were collected could influence element ratios. Climatic conditions differ between sites in the global data set, but often information on climate is not provided in published papers. While some of the eight species examined are restricted to bogs (e.g., Sphagnum fuscum), others occur in both bog and non-bog habitats (Table S2). How these factors may affect element concentrations and/or ratios in lichen/plant tissues within the context of stoichiometric homeostasis is inadequately understood.
With respect to the three questions posed initially, there is evidence for some level of stoichiometric homeostasis in the eight lichen/plant species across the six bogs in the AOSR of northern Alberta, Canada, moreso for N, P, and K than for Ca, Mg, and S. There is less evidence that the patterns of stoichiometric homeostasis for the six bogs in the AOSR prevail across other sites globally.
Regarding the third initial question related to nutrient limitation, element ratios have been used as indicators of nutrient limitation, and as such can be used to assess a hypothesis of N-limitation across each of the eight lichen/plant species in the AOSR bogs. Ideally, critical element ratios are determined from controlled laboratory fertilization studies in which all essential elements are provided in adequate supply for growth except for one element for which availability is experimentally manipulated. Most of the research on nutrient limitation in bogs has focused on N and P, with several studies reporting that as N deposition increases, there is a shift from N-limitation to P-limitation (e.g., Malmer 1988; Aerts et al. 1992; Walbridge and Navaratnam 2006; Phuyal et al. 2008). However, for wetlands, including bogs, in eastern North America, this shift was less apparent (Bedford et al. 1999). Based on fertilization studies, Koerselman and Meuleman (1996) suggested that plant growth is N-limited when N:P < 14 and P-limited when N:P > 16, with co-limitation when 14 ≤ N:P ≤ 16. Applying these criteria, Cladonia mitis, Sphagnum capillifolium, Vaccinium oxycoccos, Vaccinium vitis-idaea and Picea mariana would be regarded as N-limited, while Evernia mesomorpha would be regarded as P-limited at all six of the AOSR bogs (Fig. 1, Tables 2, S3). Element limitation classifications differed between bogs for Sphagnum fuscum, which would be regarded as N- or N + P-limited and for Rhododendron groenlandicum, which would be regarded as N- or P-limited (Table S3). These nutrient limitation interpretations generally hold when the Koerselman and Meuleman (1996) criteria are applied to sites beyond the six AOSR bogs, but with more frequent P-limitation. Across all non-Alberta sites/regions, N-limitation was indicated for S. capillifolium and V. vitis-idaea, but for the other six species, both N- and P-limitation was indicated (Fig. 1, Tables 2, S3).
Critical element ratios for wetland plant species were later modified and expanded to include K, with N:P < 14.5 and N:K < 2.1 indicating N-limitation, N:P > 14.5 and K:P > 3.4 indicating P- or P + N-limitation, and N:K > 2.1 and K:P < 3.4 indicating K- or K + N-limitation (Olde Venterink et al. 2003). Based on these critical ratios, for the six AOSR bogs, Cladonia mitis would be considered to be K- or K + N limited, Evernia mesomorpha would be considered to be P- or P + N-limited, and Picea mariana would be considered to be N-limited (Fig. 1, Tables 2, S3). For a variety of reasons, Sphagnum capillifolium, Sphagnum fuscum, Vaccinium oxycoccos, Vaccinium vitis-idaea, and Rhododendron groenlandicum are less clearly characterized as N, P, or K-limited. For these species, none of the three Olde Venterink et al. (2003) element limitation criteria are unequivocally met, and/or overall mean ratios are close to the critical ratios, and/or differences between sites would lead to different nutrient limitation interpretations (Fig. 1, Tables 2, S3).
These nutrient limitation interpretations are consistent with those obtained from a field N fertilization at Mariana Lake Bog, Alberta (Wieder et al. 2019), where N fertilization had no effect on or inhibited Sphagnum fuscum net primary production, stimulated the growth of Picea mariana (and Rhododendron groenlandicum) and led to increased tissue N concentrations in leaves of Vaccinium oxycoccos, Vaccinium vitis-idaea, Rhododendron groenlandicum, and Picea mariana, suggesting that as N supply increased, N was taken up in excess of the needs for plant growth and stored in plant tissues. Application of the Olde Venterink et al. (2003) critical ratios to species at locations other than the six AOSR bogs generally does not contradict nutrient limitation interpretations for the AOSR bogs, but for several species, which nutrients are limiting is more variable between sites/regions (Table 2). To some extent these differences may be related to site/region differences in N deposition, with high N deposition causing shifts away from N-limitation and toward P- or K-limitation.
Since there is no published evidence for Ca, Mg, or S limitation of lichens/plants in bogs, critical ratios involving these elements have not been developed. However, critical N:S ratios in crop plants have been suggested based on relationships between plant growth parameters in the field and tissue element concentrations. Based on this approach, Rasmussen et al. (1977) suggested a critical N:S ratio of 16 as an indicator of S deficiency in winter wheat; similar values have been suggested for other crop plants (e.g., Camberato and Casteel 2017, 20:1 for corn; Westerman 1975, 17:1–18:1 for alfalfa; Randall et al. 1981, Reussi Calvo et al. 2011, 17:1 to 13:1 for wheat; Randall et al. 2003, 14:1 for rice). These critical ratios for N:S reflect the N:S ratio of plant proteins, such that values higher than the critical ratio suggest S deficiency. Although N:S ratios have not been used as indicators of S deficiency in wetland or bog plants/lichens, N:S ratios for the six Alberta sites and from other sites globally are far lower than 16:1, suggesting that the eight bog lichen/plant species examined here exhibit no evidence of S-deficiency or S-limitation.
N, P, K ternary diagrams
To more closely examine stoichiometric relationships (initial questions 1 and 2) and nutrient limitation (initial question 3) for the eight bog lichen/plant species, ternary NPK diagrams were constructed, graphically delineating regions indicating N-limitation, P- or P + N- co-limitation, or K- or K + N-limitation, along with a central triangular region where N-, P-, or K-limitation cannot be inferred (Olde Venterink et al. 2003). It is important to note that this approach does not consider absolute element concentrations in lichen/plant tissues (in units like mg of an element g−1 dry mass of tissue), but the relative abundance of each of three elements as a proportion of the sum of the three elements.
The ternary diagram approach has not been applied to lichens, a group for which nutrient limitation is not well understood. While in general lichen growth may be limited by water, light, nitrogen, and/or phosphorus (Palmqvist et al. 2008), understanding nutrient limitation in lichens is confounded by potentially different nutrient requirements of the mycobiont and the associated photobiont (cf. Makkonen et al. 2007, Johansson et al. 2011). The use of element ratios to assess nutrient status/limitation in lichens is not common, in part because of the difficulty in growing whole lichens in culture (Nash III 2008), which makes experimental determination of element limitation in controlled nutrient addition experiments a challenge. Acknowledging that the Olde Venterink et al. (2003) element limitation criteria were not developed with lichens in mind, application of this approach in ternary diagrams (Fig. 1a, b) suggests K- or K + N-limitation for C. mitis and P or P + N-limitation for E. mesomorpha at the six AOSR bog sites, with no clear differences between sites (Fig. 2). This interpretation would be the same for the non-Alberta sites, with some suggestion of K- or K + N-limitation for E. mesomorpha. More striking, however, is that across all sites globally, each species maintains a remarkably consistent K:P ratio, across varying relative abundances of N in lichen thalli. For C. mitis (Fig. 2a) and E. mesomorpha (Fig. 2b), the global average (± standard errors) K:P ratios were 2.72 ± 0.02 (n = 275) and 3.93 ± 0.06 (n = 407), respectively (Table S3). While this result points to a remarkable stoichiometric homeostasis in K:P ratio for the two lichen species, neither the physiological basis nor the contributions of the fungal versus algal symbiont in maintaining K:P ratios within such a narrow range is known.

Ternary N, P, K diagrams for each of the eight bog lichen/plant species at the six Alberta bog sites samples in 2018 and 2019 and for other bog and non-bog sites globally. Dashed lines indicate critical ratios from Olde Venterink et al. (2003). Also following Olde Venterink et al. (2003) regions indicating N-limitation, P- or P + K-limitation, and K- or K + N-limitation are identified. Observations in the central triangular region do not clearly indicate N, P or K-limitation, or than a factor other than N, P, or K is limiting. For visual reasons, P concentration is multiplied by 10
In contrast to the case with lichens, element ratios have been widely used as indicators of element limitation and/or biogeochemical niches in bryophytes across a variety of habitats (e.g., Aerts et al. 1992; Bragazza et al. 2004; Jiroušek et al 2011; Waite and Sack 2011; Hájek et al. 2014; Wang and Moore 2014; Wang et al. 2016; Fernández-Martínez et al. 2021). However, only a few studies have focused on Sphagnum species in bogs using ternary diagrams. At Mer Bleue Bog, Ontario, mosses as a plant functional type (S. angustifolium, S. capillifolium, S. cuspidatum, S. fallax, S. magellanicum combined) were determined to be N and P co-limited, with observations lying along the N and P co-limitation line (N:P = 14.5) in a ternary diagram (Wang and Moore 2014). At the same site, after a decade of N, P, and K fertilization at various levels, mosses (S. capillifolium, S. magellanicum, Polytrichum strictum combined) showed consistent K or K + N co-limitation across control and fertilized treatments (Wang and Moore 2016). In neither of these studies were mosses examined at a species-by-species level.
Ternary diagrams do not produce a clear indication of element limitation for Sphagnum capillifolium (Fig. 2c) or Sphagnum fuscum (Fig. 2d). For the six bogs in the AOSR, observations for Sphagnum capillifolium (Fig. 2c) lie close to the N:K = 2:1 line, although a substantial proportion of the observations lie within the central triangular region where element limitation cannot be determined; there are no clear differences between the six bogs. In contrast to the Alberta sites, S. capillifolium from Finland (n = 2) and eastern Canada (n = 14) are displaced to the right, indicating a higher relative abundance of K compared to N and P, with N or P or P + N-limitation. For S. fuscum (Fig. 2d), most of the observations at the six bogs in the AOSR fall within the central triangular region, with no clear differences between these bogs, suggesting limitation by some factor other than N, P, or K. For S. fuscum at other sites, observations are shifted to the right (slightly higher relative abundance of K and slightly lower relative abundance of P; Fig. 2D), suggesting P or P + N co-limitation. These results do not strongly support stoichiometric homeostasis regarding N, P, and K for either Sphagnum species as a global phenomenon.
Previous studies in bogs in the AOSR have shown that N concentrations, but not K concentrations, in the capitula of S. capillifolium and S. fuscum were positively correlated with growing season N or K deposition, respectively; capitulum P concentrations were positively correlated with growing season P deposition for S. capillifolium, but not for S. fuscum (Wieder et al. 2022). These results suggest that capitulum element concentrations differ between elements and between the two Sphagnum species (at least for P). Further, in several studies in bogs across Alberta, no relationship was found between Sphagnum fuscum net primary production and either atmospheric N deposition or experimental N addition; however, in several of these studies, S. fuscum NPP was positively correlated with mean temperature and/or total precipitation in specific months during the growing season (Vile et al. 2014; Wieder et al. 2010, 2016b, 2019). With respect to Sphagnum growth and nutrient limitation, climatic factors may be especially important in AOSR bogs, which lie at the dry end of the climatic regimes under which boreal bogs persist (Gignac and Vitt 1994; Wieder et al. 2006), and where annual potential evapotranspiration to precipitation ratios are close to 1.0 (cf. Winter and Woo 1990).
Further, in Alberta bogs, N inputs to bogs by N2-fixation, predominantly by methanotrophs associated with Sphagnum species at the peat surface, are far greater than N inputs via atmospheric deposition (Vile et al. 2014). Increasing N deposition downregulates N2-fixation in bogs (Wieder et al. 2019) as further indicated at Mer Bleue Bog Ontario, where atmospheric N deposition is about 8 kg N ha−1 year−1 (Turunen et al. 2004) and N2-fixation is a relatively minor source of new N (Živkovič et al. 2022). Within individual sites or restricted geographic regions, such as Mer Bleue Bog or bogs in the AOSR, ternary N, P, K diagrams may show a tight clustering of observations suggesting some degree of stoichiometric homeostasis. Globally, however, where Sphagnum species lie on ternary plots may vary considerably depending on factors including the relative importance of N2-fixation versus atmospheric N deposition, patterns of P and K deposition, and growing season climatic factors. Such variation argues more for plasticity in N, P, and K relative abundances than for N, P, and K stoichiometric homeostasis as a global characteristic of individual Sphagnum species in bogs.
The two Vaccinium species (Vaccinium oxycoccos, Vaccinium vitis-idaea) were remarkably similar in ternary plots (Fig. 2e, f), with no clear differentiation between the six AOSR bogs and with most of the observations globally lying in the central triangular region, suggesting limitation by something other than N, P, or K. Beyond the AOSR bogs, for V. oxycoccos the cluster of observations displaced to the right in the ternary diagram (Fig. 2e) are all from Dwingeloo Bog in the Netherlands where at the time of sample collection, N deposition was extremely high (37 kg N ha−1 year−1; Heijmans et al. 2002; Hoosbeek et al. 2002). This result is consistent with the potential for bog plants to become P-limited (and/or K-limited) in high N deposition environments (Goodman and Perkins 1968; Aerts et al. 1992; Gunnarsson and Rydin 2000; Hoosbeek et al. 2002; Bragazza et al. 2004; Phuyal et al. 2008; Jiroušek et al. 2011). The non-AOSR observations for V. vitis-idaea overlap considerably with the observations from the six AOSR bogs (Fig. 2F). Both V. oxycoccos and V. vitis-idaea are small plants with fine root systems that are very close to the peat surface. As such, Malmer et al. (1994) suggested that V. oxycoccos (and presumably V. vitis-idaea) may rely on the same nutrient sources as Sphagnum mosses. Further shallow-rooted Vaccinium species may rely on the water-conducting capacity of Sphagnum mosses for their water supply (Malmer et al. 1994). Periodic drying of surface peat is deleterious to V. oxycoccos (and presumably V. vitis-idaea) (Rodwell 1991). Therefore, it may be the case that these two Vaccinium species are more limited by water availability than by nutrient availability, at least in Alberta, but possibly globally. In a previous study, we found that V. oxycoccos and V. vitis-idaea leaf N concentrations were positively correlated with growing season N deposition, but V. oxycoccos and V. vitis-idaea leaf P or K concentrations were not positively correlated with growing season P or K deposition, respectively (Wieder et al. 2021, 2022). The sclerophyllous leaves of these two species not only minimize water loss, but also are associated with a high nutrient use efficiency (Small 1972a,b; Chapin III 1980). For all of these reasons, these two Vaccinium species may exhibit stoichiometric homeostasis with respect to N, P, and K, as indicated by the tight clustering of global observations in ternary diagrams, without being clearly limited by any of these three elements.
Like the two Vaccinium species, Rhododendron groenlandicum roots can form associations with ericoid mycorrhizae (Malloch and Malloch 1981; Massicotte et al. 2005; Hébert and Thiffault 2011). However, the roots of R. groenlandicum penetrate deeper into the peat (mean depth of 45 cm at the Acadia Forest Experiment Station, New Brunswick; Flinn and Wein 1977). As with the two Vaccinium species, in a ternary plot, R. groenlandicum observations from the six AOSR bogs mostly cluster in the central triangular region, with a small group of observations from the two bogs most distant from oil sands operations displaced upwardly into the P or P + N-limitation region in 2018, but not in 2019 (Fig. 2g). Across the six AOSR bogs, R. groenlandicum leaf K:P ratios were stoichiometrically maintained at an average value of 4.78 ± 0.14 (mean ± standard error, n = 278) in the face of the relative abundance of N ranging from 30 to 70% of N + 10P + K (Fig. 2g). At Mariana Lake Bog, Alberta, experimental N addition stimulated R. groenlandicum net primary production, but only at N addition rates in excess of 22.6 kg N ha−1 year−1; N addition led to an increase in R. groenlandicum leaf N concentrations in only one of the 5 years of N fertilization (Wieder et al. 2019). Further, there was no clear trend in R. groenlandicum net primary production at the six AOSR bog sites with distance from the oil sands industrial center in either 2018 or 2019 (Vitt et al. 2020). It may be the case that the N deposition gradient across the six AOSR bog sites is not sufficiently steep to affect R. groenlandicum net primary production. However, at five bogs in the AOSR, C:N ratios in R. groenlandicum leaves were inversely correlated with growing season NH4+–N and NO3−–N deposition (Wieder et al. 2021).
Further, across 19 bogs in a 3255 km2 region centered on the oil sands industrial center, N concentrations in R. groenlandicum leaves were positively correlated with interpolated NH4+-N, NO3−–N and DIN deposition (Wieder et al. 2016a). Fertilization of an Alberta bog at a rate of 30 kg N ha−1 year−1 had no significant effect on R. groenlandicum production (Li and Vitt 1997), while fertilization of another Alberta bog at extremely high levels (150 kg ha−1 year−1 of N, 21.5 kg ha−1 year−1 of P) inhibited the growth of R. groenlandicum (Thormann and Bayley 1997). Increases in leaf N concentration with increasing N deposition without a stimulation of net primary production suggests that R. groenlandicum is not N-limited, at least in bogs of the AOSR, and can take up and store N in leaf tissues when N availability increases.
For R. groenlandicum beyond the six AOSR bog sites, some observations lie within the central triangular region in the ternary plot, while some are in the N-limitation region and some are in the P or P + N-limitation region (Fig. 2g). The tight control on the K:P ratio observed for the AOSR bogs does not extend to R. groenlandicum at other sites (Fig. 2g). Ternary NPK plots for R. groenlandicum at Mer Bleue Bog, Ontario, show observations from control (unfertilized) plots in the central triangular region with N fertilization alone displacing observations upward into the P- or P + N-limitation region, and P + K or N + P + K fertilization displacing observations downward into the N-limitation or the K- or K + N-limitation region (Wang et al. 2016). These fertilization experiments led to a conclusion that shrubs at Mer Bleue, including R. groenlandicum are N and P co-limited (Wang et al. 2016). That fertilization can shift the locations of observations in NPK ternary diagrams suggests plasticity in element ratios more than strong stoichiometric homeostasis.
Picea mariana observations fall within the N-limitation sector of the ternary diagram, with no clear differences between the six AOSR bogs, and with observations beyond these bogs also reflecting N-limitation (Fig. 1h). Beyond the AOSR bogs, some observations suggest K- or K + N-limitation. Experimental fertilization at Mariana Lake Bog at rates up to 25 kg N ha−1 year−1 stimulated P. mariana growth and led to small increases in N concentrations in first-year needles (Wieder et al. 2019). Under extremely high rates of N fertilization (up to 672 kg ha−1) P. mariana growth can be stimulated by K and/or P fertilization (Alban and Watt 1981; Wells 1994), reflecting a shift from N- to K- and/or P-limitation. Overall, P. mariana is N-limited, with broad ranges of the relative abundances of N, P, and K and little evidence of strong stoichiometric NPK homeostasis (Fig. 1h).
Ca, Mg, K ternary diagrams
Given that there is no evidence for Ca- or Mg-limitation of any lichen/plant species in bogs, and evidence for potential K-limitation only under very high N deposition, CaMgK ternary diagrams are used there to assess stoichiometric relationships, not nutrient limitation. Across the six AOSR bogs, Ca and Mg deposition, most likely influenced by fugitive dust, decreases with distance from the oil sands industrial center, and K deposition differs between sites, possibly reflecting spatial variability in the effects of wildfires on K deposition (Fig. S1; Wieder et al. 2022). These spatially variable deposition regimes allow for the examination of the extent to which bog lichen/plant species exhibit evidence for stoichiometric homeostasis with respect to Ca, Mg, and K. In interpreting ternary Ca, Mg, K diagrams, the primary focus is the six AOSR bogs, and secondarily, to assess whether stoichiometric patterns, if they exist, can be extended to other sites globally.
In these CaMgK ternary diagrams when observations are arrayed vertically, a relatively constant Mg:K ratio is indicated in the face of variable Ca abundances; when observations are arrayed from upper left to lower right, a relatively constant Ca:Mg ratio is indicated in the face of variable K abundances, and when observations are arrayed from lower left to upper right, a relatively constant Ca:K ratio is indicated in the face of variable Mg abundances. When these patterns appear, lines indicating average element ratios are included in Fig. 3.

Ternary Ca, Mg, K diagrams for each of the eight bog lichen/plant species at the six Alberta bog sites samples in 2018 and 2019 and for other bog and non-bog sites globally. Dashed lines where shown indicate mean Ca:Mg or K:Mg ratios averaged across the Alberta bogs and the two years of sampling. For visual reasons, Ca and Mg concentrations are multiplied by 5
The two lichen species differ with respect to Ca, Mg, K relationships (Fig. 3a, b), which is surprising given that both Cladonia mitis and Evernia mesomorpha are associations between an ascomycete fungus and non-N2-fixing green algal symbiont in the genus Trebouxia (Piercey-Normore 2004, 2006). For C. mitis, across the six AOSR bog sites, observations cluster in the same general region with no clear differences between sites (Fig. 3a), suggesting a degree of stoichiometric homeostasis, especially with respect to Ca:Mg ratios, which have lower coefficients of variation than for K:Ca or K:Mg (Table S4). Beyond the six AOSR bogs, however, observations are more variable (Fig. 3a), suggesting that the degree of stoichiometric homeostasis observed for the AOSR bogs, especially for Ca:Mg, does not prevail globally. In contrast, observations for E. mesomorpha suggest stoichiometric homeostasis with respect to K:Mg ratios, which are relatively constant in the face of quite variable Ca relative abundances (Fig. 3b). This pattern is also reflected in the much lower coefficient of variation for K:Mg than for K:Ca or Ca:Mg for E. mesomorpha across the AOSR bogs (Table S4). Further, the observations for E. mesomorpha are arrayed along a gradient of increasing Ca relative abundance with proximity to the oil sands industrial center, most likely reflecting the influence of Ca- and, to a lesser extent, Mg-rich fugitive dust from oil sands operations (Wieder et al. 2022).
Beyond the six AOSR sites, E. mesomorpha observations also suggest K:Mg homeostasis, but with a greater degree of variability (Fig. 3b). Lichens can accumulate cations abiotically by particulate trapping, retention of soluble cations by cation exchange on cell surfaces, retention of newly formed cations from dissolution of trapped particulates, or biotically through intracellular uptake (Nieboer et al. 1978). Further, the relative role of the fungal versus algal symbiont in Ca, Mg, or K accumulation is unknown. As such, the physicochemical and physiological bases behind apparent Ca, Mg, K stoichiometric homeostatic relationships in lichens remains poorly understood.
Across the six AOSR bogs, the general distribution of observations for the two Sphagnum species was similar, with no clear differences between sites (Fig. 3c, d). There is a suggestion that for S. capillifolium, there is tighter control of Ca:Mg ratios over varying K abundances (Fig. 3c), which is also reflected in the lower coefficient of variation for the Ca:Mg ratio than for K:Ca or K:Mg ratios (Table S4). For S. fuscum, however, coefficients of variation are similar for Ca:Mg, K:Ca, and K:Mg ratios (Table S4). Results from the two Sphagnum species at the 6 AOSR bogs differ considerably from findings at Mer Bleue Bog, Ontario, where ternary Ca, Mg, K diagrams indicated that mosses (Sphagnum capillifolium, Sphagnum magellanicum, Polytrichum strictum in combination) maintained a fairly constant K relative abundance (10%) while Ca relative abundance ranged from 20 to 40% and Mg relative abundance ranged from 50 to 70%; moss Ca, Mg, K stoichiometry was not clearly related to availability in peat as assessed using plant root simulator probes (Wang et al. 2018). Whether the relatively close clustering of observations for S. capillifolium at the six Alberta sites extends more generally to other regions cannot be assessed given the paucity of data for sites beyond the six AOSR bogs. In contrast, observations for S. fuscum beyond the AOSR bogs indicate a greater overall variability (Fig. 2d), suggesting that the stoichiometric homeostasis indicated by the relatively close clustering of observations from the AOSR bogs is not a characteristic of S. fuscum globally.
Among the vascular plant species, Vaccinium oxycoccos had a unique pattern of a very tight clustering of observations in the ternary diagram, with no clear differences between the six AOSR bog sites (Fig. 3e). Coefficients of variation for Ca:Mg, K:Ca, and K:Mg ratios were similar (Table S4) suggesting a tight three-way Ca, Mg, K stoichiometric homeostasis, which appears to prevail at sites beyond Alberta, especially for bogs. Different patterns are manifested for the other three vascular plant species. While many of the observations for V. vitis-idaea from the AOSR bogs also cluster in the upper left regions of the ternary plot with no clear differences between sites, the pattern suggests a maintenance of narrow Ca:Mg ratios across a range of K relative abundance (Fig. 3f). This pattern is supported by lower coefficients of variation for Ca:Mg ratios than for K:Ca or K:Mg ratios (Table S4). The patterns for V. vitis-idaea for the AOSR bogs appear to be consistent with sites beyond Alberta, even though none of these sites are bogs (Fig. 3f, Tables S2, S3). For the six AOSR bogs, observations for R. groenlandicum in the ternary plot (Fig. 3g), along with coefficients of variation (Table S4), indicate a maintenance of narrow Ca:Mg ratios across wide ranges of K relative abundance, similar to the patterns observed for V. vitis-idaea. These patterns exist for R. groenlandicum beyond the AOSR bogs, but with a higher degree of variability (Fig. 3g).
At Mer Bleue Bog, Ontario, Ca, Mg, K ternary diagrams showed that shrubs (Chamaedaphne calyculata, Rhododendron groenlandicum, Vaccinium myrtilloides, Kalmia angustifolia, combined) maintained a relatively constant Mg relative abundance of about 10%, while K and Ca relative abundances varied between 40 and 60% across control and experimentally fertilized plots (Wang et al. 2018). A similar pattern was obtained for Ca, Mg, and K availability assessed using plant root simulator (PRS) probes, suggesting that plant tissue stoichiometry mirrored availability in peat. Overall, Ca:K:Mg ratios in shrub leaves were 4:2:1 and in PRS probes were 4:4:1 (Wang et al. 2018). Across the six AOSR bogs, Ca:K:Mg ratios in the shrub species V. oxycoccos, V. vitis-idaea, and R. groenlandicum were 6:3:1 (n = 180), 3:3:1 (n = 179), and 3:6:1 (n = 179), respectively; collectively for the three shrub species Ca:K:Mg ratios were 4:4:1. Although shrub Ca:K:Mg ratios were similar between Mer Bleue Bog and the AOSR bogs when the three shrub species were combined, the stoichiometric relationships between the three individual shrub species in the AOSR bogs were quite different. Treating bog shrubs collectively as a plant functional group may belie species-specific differences in elemental stoichiometries.
The ternary diagram for Picea mariana suggests two patterns related to stoichiometry for the AOSR bog sites. At relatively high Ca abundance, a tight Ca:Mg ratio is maintained, and at relatively low Ca abundance a tight K:Mg ratio is maintained (Fig. 2h), with no evident differences between sites. Beyond the AOSR bogs, observations tended to have lower relative K relative abundance and higher relative Ca abundance. These differences are reflected in average Ca:K:Mg ratios of 3:10:1 for the AOSR bogs and 4:3:1 for the other sites, suggesting a relatively high K availability, uptake, and storage of K for P. mariana in the AOSR bogs. At the AOSR bog sites, P. mariana needle Ca, Mg, or K concentrations were not correlated with Ca, Mg, or K deposition, respectively, as measured using ion exchange resin collectors (Wieder et al. 2022). Nonetheless, the considerable spread of the observations in Fig. 3h could be the result of within-site and between-site variability in the capture of base cations in dry deposition onto needle surfaces.
Niche volumes and scaling
The niche volume approach of Ågren and Weih (2020) allows for interpretations in lichen/plant regulation of element uptake relative to changing N and P concentrations, thus the niche volume approach is relevant to stoichiometry, but not nutrient limitation questions. When \({V}_{Oth}\) is calculated for a single element (other than N or P), a scaling coefficient (α) of 1 indicates that \({V}_{NP}\) and \({V}_{Oth}\) increase proportionally, which would indicate stoichiometric homeostasis (constant ratio of the element to NxP as NxP increases). A scaling coefficient < 1 indicates that concentrations of other elements in lichen/plant tissue increase more slowly than concentrations of N and P; a scaling coefficient > 1 indicates that concentrations of other elements in lichen/plant tissue increase more rapidly than concentrations of N and P. Scaling coefficients that are significant, but either greater than or less than 1, suggest lichen/plant regulation of element uptake relative to N and P, but in such a way that does not maintain constant element ratios, i.e., scaling exists, but regulation of element uptake does not lead to stoichiometric homeostasis. Nonsignificant regressions indicate an absence of scaling, and a lack of regulation of element uptake relative to N and P, also reflecting an absence of stoichiometric homeostasis. As the number of elements included in \({V}_{Oth}\) increases, scaling coefficients also increase, specifically by 0.703 for each additional element beyond 1 (Ågren and Weih 2020). Ågren and Weih (2020) used 10 published data sets to estimate scaling coefficients between \({V}_{NP}\) and \({V}_{Oth}\), where \({V}_{Oth}\) included Ca, Mg, K, and S. Scaling coefficients ranged from 0.938 and 2.479; when data sets from individual studies included different treatments or other factors, differences between scaling coefficients between data subsets were small.
For the six AOSR bogs, overall there was widespread significance of regressions between \({V}_{NP}\) and \({V}_{Oth}\), with scaling coefficients ranging from − 1.18 to 3.42 (Table 3). Several notable patterns in scaling coefficients were observed. When \({V}_{Oth}\) included Ca, Mg, K, and S, scaling coefficients across all six bog sites were significant except for S. capillifolium and V. vitis-idaea. When these overall scaling coefficients were significant, differences between the six bog sites often were small and/or did not progressively change with increasing distance from the oil sands industrial center. Ågren and Weih (2020) showed that when \({V}_{Oth}\) included four elements, a scaling coefficient of 2.4 was obtained. While scaling coefficients for the two lichens and the two Sphagnum species across the six bog sites ranged from 2.3 to 3.4, coefficients for the vascular plant species were considerably lower, ranging from 0.6 to 1.5 (Table 3). Thus, as N and P concentrations in lichen/plant tissues increase, lichens and Sphagnum can increase their Ca, Mg, K, and S concentrations at a faster rate than the vascular plant species. These differences could be related to the much greater role of cation exchange in retaining base cations in lichens and Sphagnum than in in vascular plants (Wieder et al. 2022), so the differences may be related more to abiotic than biotic factors. In general, scaling coefficients for Ca and Mg, but not K, were higher for the lichens and Sphagnum species than for the four vascular plant species, which would be expected given the higher affinity of cation exchange sites for divalent cations than monovalent cations. For the vascular plant species, several of the scaling coefficients for Ca and Mg were negative, which could be the result of the dilution of leaf Ca and Mg concentrations associated with stimulated growth associated with increasing N and P concentrations (Ågren and Weih 2020). At Mer Bleue Bog, Ontario, eight years of NPK fertilization led to decreases in leaf Ca concentrations in Rhododendron groenlandicum, Chamaedaphne calyculata, and Vaccinium myrtilloides, with a suggestion of a similar response for leaf Mg concentrations (Bubier et al. 2011).
Within-species scaling coefficients for S were generally variable between the six bog sites except for R. groenlandicum and P. mariana for which S scaling coefficients were all significant and positive, but less than 1, indicating a slower rate of S accumulation in leaves/needles than of N and P. The physiological/physicochemical mechanisms that may drive this relationship are not known.
Summary and synthesis
Ecological stoichiometry has been a compelling concept for decades (van der Waal et al. 2018), with Sterner and Elser (2002) arguably providing a catalyst for considerable research across a variety of terrestrial, aquatic, and wetland systems. Only a few studies, however, have focused on element stoichiometric homeostasis in bogs (Wang and Moore 2014; Wang et al. 2016, 2018). There is no objective criterion to determine with certainty that a particular species or plant functional group does or does not exhibit stoichiometric homeostasis with regard to tissue element concentrations. Rather, data are examined for patterns that may suggest stoichiometric homeostasis using pairwise element ratios, coefficients of variation for single elements or element ratios, grouping of observations in ternary diagrams, and niche volume theory, along with other approaches, as well. Subjective interpretation is used to assess any patterns that emerge; sometimes these emergent patterns have been explained mechanistically by our understanding of physiological/physicochemical processes, but sometime not. Stoichiometric homeostasis is related to nutrient limitation in that as the availability of a limiting nutrient increases, the uptake of other non-limiting nutrients would have to increase proportionately in order to maintain stoichiometric homeostasis.
With respect to the question of whether the eight lichen/plant species in the six bogs in the AOSR exhibit stoichiometric homeostasis, the evidence is mixed. Given that atmospheric deposition of N, P, K, S, Ca and Mg differs considerably between the six AOSR bogs (Fig. S1), and assuming that these differences reflect differences in nutrient availability between sites, similar element ratios across all of the bogs would support the concept of stoichiometric homeostasis. However, in general, element ratios differed between at least two of the bogs, although differences did not parallel patterns of element deposition (Fig. S1). When statistically significant differences in element ratios between bogs occurred, however, the magnitude of the differences often was quite small. Using a criterion of mean element ratios at each of the six bogs being within 25% of the mean across all of the six bogs, there was widespread stoichiometric homeostasis, although which element ratios met the 25% criterion differed both within and between species (Fig. 1, Table S3).
Ternary diagrams were especially useful in assessing stoichiometric homeostasis, revealing patterns that were not especially evident through examination of individual element ratios and their coefficients of variation. The NPK ternary diagrams exhibited unexpected relationships. The two lichen species exhibited very tight control over K:P ratios over a broad range of N relative abundances, both for the six AOSR bogs and beyond (Fig. 2). A physiological basis for this is not known. The two Sphagnum species and the two Vaccinium species in the six AOSR bogs displayed fairly tight clusters in their NPK ternary plots, suggesting a high degree of three-element stoichiometric homeostasis, but for three of these species, data from sites beyond Alberta were shifted away from the Alberta cluster, suggesting plasticity in stoichiometry on a global basis.
Ternary CaMgK diagrams also revealed unanticipated results, with six of the lichen/plant species maintaining narrow Ca:Mg variability over a wide range in K abundance and/or narrow K:Mg variability over a wide range in Ca abundance (Fig. 3). These patterns often were evident for sites beyond the AOSR bogs, suggesting a geographically broad stoichiometric homeostasis for Ca:Mg and/or K:Mg, although a mechanistic physiological basis behind these patterns is not known.
Recognizing the cumbersome nature of examining element ratios for several elements (in this case, 6 elements and 15 pairwise ratios; Table 1), Ågren and Weih (2020) proposed a more streamlined approach based on niche volume theory. Overall, the niche volume approach indicates widespread significant scaling of lichen thallus/Sphagnum capitula/leaf/needle Ca, Mg, K, and S concentrations individually and in combination as related to N and P concentrations. Across the eight lichen/plant species in the six AOSR bogs, rarely are the scaling coefficients approximately equal to 1 (Table 3), suggesting that while these eight lichen/plant species may control Ca, Mg, K, and S uptake and storage in relation to N and P, stoichiometric homeostasis, i.e. the maintenance of relatively constant ratios of Ca, Mg, K, and S to N and P, is not strongly indicated.
With respect to nutrient limitation, the Koerselman and Meuleman (1996) approach assumes that plants are N-limited, P-limited, or N + P-colimited. Often using mean N:P ratios for a species within a particular site or region, a species will be categorized as being in one of these three nutrient limitation categories. When Olde Venterink et al. (2003) added the possibility of K-limitation, N:P, N:K, and K:P ratios placed species into one of three categories: N-limitation, P- or P + N-limitation, or K or K + N-limitation. It is possible, however, that the three element ratios would not fall into any of the nutrient limitation categories, as was sometimes the case in this study (Fig. 3, Table 2). Olde Venterink et al. (2003) used NPK ternary diagrams to visualize element ratio means and place species into one of the three nutrient limitation categories, as well as a fourth region of the ternary diagram where neither N-, P-, nor K-limitation is indicated. Plotting raw data in NPK ternary diagrams (individual observations rather than means), as done here (Fig. 2), illustrates within-species variability, and can reveal stoichiometric relationships between these three elements.
While in general, the three approaches to determining nutrient limitation in the eight species examined here produced noncontradictory results (Table 2), the Olde Venterink et al. (2003) approach using mean values for N:P; N:K, and K:P ratios and ternary diagram approach often revealed that a species can be limited in different ways depending on where it grows. Different nutrient limitation in different locations for a given species does not appear to be related to whether the species grows in bog or non-bog habitats, but rather may be due, at least in part to species being N-limited in low N deposition (low N availability) habitats, with shifts to P- or K- limitation in high N deposition habitats (e.g. Goodman and Perkins 1968; Aerts et al. 1992; Gunnarsson and Rydin 2000; Hoosbeek et al. 2002; Bragazza et al. 2004; Phuyal et al. 2008; Jiroušek et al. 2011). Because N deposition and/or N availability often is not given in published papers with lichen/plant tissue chemistry data, the extent to which changing N deposition regimes affects nutrient limitation cannot be rigorously examined using this data set.
Ternary NPK diagrams rather clearly indicated that most of the observations for the two Sphagnum species and the two Vaccinium species in the six AOSR bog sites clustered into the central triangular region where neither N-, P-, of K-limitation is indicated (Fig. 1). For the Sphagnum species, the absence of clear N-limitation is likely, at least in part, the result of methanotrophic N2-fixation in moss capitula, representing a much higher input of N than atmospheric deposition (Vile et al. 2014; Wieder et al. 2019). Given this, a pattern indicative of P- or K-limitation in the NPK ternary diagrams might have been expected, but was not observed. For the Sphagnum and Vaccinium species in the AOSR bogs, water availability may be more of a limiting factor than nutrient availability, especially given the relatively dry climate in which Alberta bogs exist, as compared to boreal bogs globally. For sites beyond the AOSR bogs, higher N deposition and attendantly lower N2-fixation along with wetter climates could shift the Sphagnum and Vaccinium species into regions of either N-limitation or P- or P + N-limitation, as was sometimes seen (Fig. 2).
As a final note, boreal bogs have existed for millennia in environments where atmospheric nutrient deposition was most likely uniformly low, but precipitation/evaporation regimes may have been quite variable. Bogs would have been acidic, nutrient poor ecosystems worldwide. One could speculate that the nutrient limitation indications in Table 2 might have been largely either N-limitation or neither N-, P-, nor K-limitation (water limitation) if data had been collected prior to the Industrial Revolution. Of course, this is a non-testable hypothesis. But if true, the within-species and between-species variability in nutrient limitation seen in Table 2 and Fig. 2 would illustrate the extent to which anthropogenic alteration of nutrient deposition regimes has altered bog ecosystem function, as indicated by which elements are limiting to individual species in bogs, and whether for a given species limiting elements are consistent at local, regional, and global scales.
Data availability
The AOSR bog data are available at the Environmental Data Initiative: https://doi.org/10.6073/pasta/043c6ebb2e2294ab96f0bdcaff10601e
References
Aerts R, Wallén B, Malmer N (1992) Growth-limiting nutrients in Sphagnum-dominated bogs subject to low and high atmospheric nitrogen supply. J Ecol 80:131–140. https://doi.org/10.2307/2261070
Ågren G, Weih M (2020) Multi-dimensional plant element stoichiometry—looking beyond carbon, nitrogen, and phosphorus. Front Plant Sci 11:23. https://doi.org/10.3389/fpls.2020.00023
Alban DH, Watt RF (1981) Fertilization of black spruce of poor site peatland in Minnesota. U.S. Department of Agriculture Forest Service, Research Paper NC-210, 10 p. U.S. Department of Agriculture Forest Service, North Central Forest Experiment Station, St. Paul, Minnesota. https://doi.org/10.2737/NC-RP-210
Bedford BL, Walbridge MR, Aldous A (1999) Patterns in nutrient availability and plant diversity of temperate North American wetlands. Ecology 80:2151–2169. https://doi.org/10.2307/176900
Belland RJ, Vitt DH (1995) Bryophyte vegetation patterns along environmental gradients in continental bogs. Écoscience 2:395–407. https://doi.org/10.1080/11956860.1995.11682308
Bragazza L, Tahvanainen T, Kutnar L, Rydin H, Limpens J, Hájek M, Grosvernier P, Hájek T, Hájková P, Hansen I, Iacumin P, Gerdol R (2004) Nutritional constraints in ombrotrophic Sphagnum plants under increasing atmospheric nitrogen deposition in Europe. New Phytol 163:609e616. https://doi.org/10.1111/j.1469-8137.2004.01154.x
Bubier JL, Smith R, Juutinen S, Moore TR, Minocha R, Long S, Minocha S (2011) Effects of nutrient addition on leaf chemistry, morphology, and photosynthetic capacity of three bog shrubs. Oecologia 167:355–368. https://doi.org/10.1007/s00442-011-1998-9
Camberato J, Casteel S (2017) Sulfur deficiency. Purdue University Department of Agronomy, Soil Fertility Update, July 11, 2017. https://www.agry.purdue.edu/ext/corn/news/timeless/sulfurdeficiency.pdf. Accessed Feb 7, 2022.
Chapin FS III (1980) The mineral nutrition of wild plants. Ann Rev Ecol Syst 11:223–260. https://doi.org/10.1146/annurev.es.11.110180.001313
Chapin FS III, Vitousek PM, van Cleve K (1986) The nature of nutrient limitation in plant communities. Am Nat 127:48–58. https://doi.org/10.1086/284466
Chapin FS III, Schultze E-D, Mooney HA (1990) The ecology and economics of storage in plants. Ann Rev Ecol Syst 21:423–447. https://doi.org/10.1146/annurev.es.21.110190.002231
Edgerton ES, Hsu Y-M, White EM, Fenn ME, Landis MS (2020) Ambient concentrations and total deposition of inorganic sulfur, inorganic nitrogen and base cations in the Athabasca Oil Sands Region. Sci Total Environ 706:134864. https://doi.org/10.1016/j.scitotenv.2019.134864
Fenn ME, Bytnerowicz A, Schilling AL, Ross CS (2015) Atmospheric deposition of nitrogen, sulfur and base cations in jack pine stands in the Athabasca Oil Sands Region, Alberta, Canada. Environ Pollut 196:497–510. https://doi.org/10.1016/j.envpol.2014.08.023
Fernández-Martínez M, Preece C, Corbera J, Cano O, Carcia-Porta J, Sanrans J, Janssens IA, Sabater F, Peñuelas J (2021) Bryophyte C:N: P stoichiometry, biogeochemical niches and elementome plasticity driven by environment and coexistence. Ecol Lett 24:1575–1586. https://doi.org/10.1111/ele.13752
Flinn MA, Wein RW (1977) Depth of underground plant organs and theoretical survival during fire. Can J Bot 55:2550–2554. https://doi.org/10.1139/b77-291
Gignac LD, Vitt DH (1994) Responses of northern peatlands to climate change: effects on bryophytes. J Hattori Bot Lab 75:119–132. https://doi.org/10.18968/jhbl.75.0_119
Goodman GT, Perkins DF (1968) The role of mineral nutrients in Eriophorum communities. IV. Potassium supply as a limiting factor in an E. vaginatum community. J Ecol 56:685–696. https://doi.org/10.2307/2258100
Gunnarsson U, Rydin H (2000) Nitrogen fertilization reduces Sphagnum production in bog communities. New Phytol 147:527–537. https://doi.org/10.1046/j.1469-8137.2000.00717.x
Güsewell S, Koerselman W (2002) Variation in nitrogen and phosphorus concentrations of wetland plants. Perspect Plant Ecol 5:37–61. https://doi.org/10.1078/1433-8319-0000022
Hájek M, Plesková A, Syrovátka V, Peterka T, Laburdová J, Kintrová K, Jiroušek M, Hájek T (2014) Patterns in moss element concentrations in fens across species, habitats, and regions. Perspect Plant Ecol 16:203–218. https://doi.org/10.1016/j.ppees.2014.06.003
Hébert F, Thiffault N (2011) The biology of Canadian weeds. 146. Rhododendron groenlandicum (Oeder) Kron and Judd. Can J Plant Sci 91:725–738. https://doi.org/10.4141/cjps2010-012
Heijmans MMPD, Klees H, de Visser W, Berendse F (2002) Response of a Sphagnum bog plant community to elevated CO2 and N supply. Plant Ecol 162:123–134. https://doi.org/10.1023/A:1020368130679
Hoosbeek MR, van Breemen N, Vasander H, Buttler A, Berendse F (2002) Potassium limits potential growth of bog vegetation under elevated atmospheric CO2 and N deposition. Glob Change Biol 8:1130–1138. https://doi.org/10.1046/j.1365-2486.2002.00535.x
Horb EC, Wentworth GR, Makar PA, Liggio J, Hayden K, Boutzis EI, Beausoleil DL, Hazewinkel RO, Mahaffey AC, Sayanda D, Wyatt F, Dubé M (2021) A decadal synthesis of atmospheric emissions, ambient air quality, and deposition in the oil sands region. Integr Environ Assess Manage 18:333–360. https://doi.org/10.1002/ieam.4539
Hsu Y-M, Bytnerowicz A, Fenn ME, Percy KE (2016) Atmospheric dry deposition of sulfur and nitrogen in the Athabasca Oil Sands Region, Alberta, Canada. Sci Total Environ 568:285–295. https://doi.org/10.1016/j.scitotenv.2016.05.205
Jiroušek M, Hájek M, Bragazza L (2011) Nutrient stoichiometry in Sphagnum along a nitrogen deposition gradient in highly polluted region of Central-East Europe. Environ Pollut 159:585–590. https://doi.org/10.1016/j.envpol.2010.10.004
Johansson O, Olofsson J, Giesler R, Palmqvist K (2011) Lichen responses to nitrogen and phosphorus additions can be explained by the different symbiont responses. New Phytol 191:795–805. https://doi.org/10.1111/j.1469-8137.2011.03739.x
Kelley K (2007) Sample size planning for the coefficient of variation from the accuracy in parameter estimation approach. Behav Res Meth 39:755–766. https://doi.org/10.3758/BF03192966
Knecht MF, Göransson A (2004) Terrestrial plants require nutrients in similar proportions. Tree Physiol 24:447–460. https://doi.org/10.1093/treephys/24.4.447
Koerselman W, Meuleman AFM (1996) The vegetation N: P ratio: a new tool to detect the nature of nutrient limitation. J Appl Ecol 33:1441–1450. https://doi.org/10.2307/2404783
Lamers LPM, Bobbink R, Roelofs JGM (2000) Natural nitrogen filter fails in polluted raised bogs. Glob Change Biol 6:583–586. https://doi.org/10.1046/j.1365-2486.2000.00342.x
Li Y, Vitt DH (1997) Patterns of retention and utilization of aerially deposited nitrogen in boreal peatlands. Écoscience 4:106–116. https://doi.org/10.1080/11956860.1997.11682384
Makkonen S, Hurri R, Hyvärinen M (2007) Differential responses of lichen symbionts to enhanced nitrogen and phosphorus availability: an experiment with Cladina stellaris. Ann Bot 99:877–884. https://doi.org/10.1093/aob/mcm042
Malloch D, Malloch B (1981) The mycorrhizal status of boreal plants: species from northeastern Ontario. Can J Bot 59:2167–2172. https://doi.org/10.1139/b82-131
Malmer N (1988) Patterns in the growth and the accumulation of inorganic constituents in the Sphagnum cover on ombrotrophic bogs in Scandinavia. Oikos 53:105–120. https://doi.org/10.1080/11956860.1997.11682384
Malmer N, Svensson BM, Wallén B (1994) Interactions between Sphagnum mosses and field layer vascular plants in the development of peat-forming systems. Folia Geobot Phytotx 29:483–496. https://doi.org/10.1007/BF02883146
Massicotte HB, Melville LH, Peterson RL (2005) Structural characteristics of root-fungal interactions for five ericaceous species in eastern Canada. Can J Bot 83:1057–1064. https://doi.org/10.1139/b05-046
McGroddy ME, Daufresne T, Hedin LO (2004) Scaling of C:N:P stoichiometry in forests worldwide: implications of terrestrial Redfield-type ratios. Ecology 85:2390–2401. https://doi.org/10.1890/03-0351
Nash III TH (2008) Nutrients, elemental accumulation, and mineral cycling. In: Nash III TH (ed) Lichen biology, 2nd edn. Cambridge University Press, Cambridge, pp 233–251. https://doi.org/10.1017/CBO9780511790478.013
Nieboer E, Richardson DHS, Tomassini FD (1978) Mineral uptake and release by lichens: an overview. Bryologist 81:226–246. https://www.jstor.org/stable/3242185
Olde Venterink H, Wasson WJ, Verkroost AWM, de Ruiter PC (2003) Species richness-productivity patterns differ between N-, P-, and K-limited wetlands. Ecology 84:2919–2999. https://doi.org/10.1890/01-0639
Palmqvist K, Dahlman T, Jonsson A, Nash TH III (2008) The carbon economy of lichens. In: Nash TH (ed) Lichen biology, 2nd edn. Cambridge University Press, Cambridge, pp 182–215
Phuyal M, Artz R, Sheppard L, Leith ID, Johnson D (2008) Long-term nitrogen deposition increases phosphorus limitation of bryophytes in an ombrotrophic bog. Plant Ecol 196:111–121. https://doi.org/10.1007/s11258-007-9338-1
Piercey-Normore M (2004) Selection of algal genotypes by three species of lichen fungi in the genus Cladonia. Botany 82:947–961. https://doi.org/10.1139/b04-084
Piercey-Normore MD (2006) The lichen-forming ascomycete Evernia mesomorpha associates with multiple genotypes of Trebouxia jamesii. New Phytol 169:331–344. https://doi.org/10.1111/j.1469-8137.2005.01576.x
Randall PJ, Spencer K, Freney JR (1981) Sulfur and nitrogen fertilizer effects on wheat. I. Concentrations of sulfur and nitrogen and the nitrogen to sulfur ratio in grain, in relation to the yield response. Aust J Agr Res 32:203–212. https://doi.org/10.1071/AR9810203
Randall PJ, Freney JR, Spencer K (2003) Diagnosing sulfur deficiency in rice by grain analysis. Nutr Cycl Agroecosyst 65:211–219. https://doi.org/10.1023/A:1022631020728
Rasmussen PE, Ramig RE, Ekin LG, Rohde CR (1977) Tissue analysis guidelines for diagnosing sulfur deficiency in white wheat. Plant Soil 46:153–163. https://doi.org/10.1007/BF00693122
Reussi Calvo N, Echeverria H, Sainz Rozas H (2011) Diagnosing sulfur deficiency in spring red wheat: plant analysis. J Plant Nutr 34:573–589. https://doi.org/10.1080/01904167.2011.538118
Rodwell JS (ed) (1991) British plant communities, vol 2. Cambridge University Press, Cambridge, Mires and heaths. https://doi.org/10.1017/9780521391658
Ruoss E (1987) Species differentiation in a group of reindeer lichens (Cladonia subg. Cladina). Bibl Lichenol 25:197–206
Small E (1972a) Ecological significance of four critical elements in plants of raised Sphagnum bogs. Ecology 53:498–503. https://doi.org/10.2307/1934240
Small E (1972b) Photosynthetic rates in relation to nitrogen recycling as an adaptation to nutrient deficiency in peat bog plants. Can J Bot 50:2227–2233. https://doi.org/10.1139/b72-289
Sterner RW, Elser JJ (2002) Ecological stoichiometry: the biology of elements from molecules to the biosphere. Princeton University Press, Princeton. https://doi.org/10.1515/9781400885695
Thormann MM, Bayley SE (1997) Response of aboveground net primary plant production to nitrogen and phosphorus fertilization in peatlands in southern boreal Alberta, Canada. Wetlands 17:502–512. https://doi.org/10.1007/BF03161516
Turunen J, Roulet NT, Moore TR, Richard PJH (2004) Nitrogen deposition and increased carbon accumulation in ombrotrophic peatlands in eastern Canada. Global Biogeochem Cycles 18:GB3002. https://doi.org/10.1029/2003gb002154
van der Ploeg RR, Böhm W, Kirkham MB (1999) On the origin of the theory of mineral nutrition of plants and the Law of the Minimum. Soil Sci Soc Am J 63:1055–1062. https://doi.org/10.2136/sssaj1999.6351055x
van der Waals DB, Elser JJ, Martiny AC, Sterner RW, Cotner JB (2018) Editorial: Progress in ecological stoichiometry. Front Microbiol 9:1957. https://doi.org/10.3389/fmicb.2018.01957
Vile MA, Wieder RK, Živkovič T, Scott KD, Vitt DH, Hartsock JA, Iosue CL, Quinn JC, Petix M, Fillingim H, Popma JMA, Dynarski KA, Jackman TR, Albright CM, Wykoff DD (2014) N2-fixation by methanotrophs sustains carbon and nitrogen accumulation in peatlands. Biogeochemistry 121:317–328. https://doi.org/10.1007/s10533-014-0019-6
Vitt DH, House M, Kitchen S, Wieder RK (2020) A protocol for monitoring plant responses to changing nitrogen deposition regimes in Alberta bogs. Environ Monit Assess 192:743. https://doi.org/10.1007/s10661-020-08645-z
Vitt DH, House M (2021) Bryophytes as key indicators of ecosystem function and structure of northern peatlands. Bryophyte Diversity Evol 43:253–264. https://doi.org/10.11646/BDE.43.1.18
Vitt DH (2006) Functional characteristics and indicators of boreal peatlands. In Wieder RK, Vitt DH (eds) Boreal peatland ecosystems. Ecological Studies 188. Springer, Berlin, pp 9–24. https://doi.org/10.1007/978-3-540-31913-9_2
Waite M, Sack L (2011) Does global stoichiometric theory apply to bryophytes? Tests across an elevation x soil age ecosystem matrix on Mauna Loa. Hawaii J Ecol 99:122–134. https://doi.org/10.1111/j.1365-2745.2010.01746.x
Walbridge MR, Navaratnam JA (2006) Phosphorus in boreal peatlands. In Wieder RK, Vitt DH (eds) Boreal peatland ecosystems. Ecological Studies 188, Springer, Berlin, pp 213–258. https://doi.org/10.1007/978-3-540-31913-9_11
Wang M, Moore TR (2014) Carbon, nitrogen, phosphorus, and calcium stoichiometry in an ombrotrophic peatland reflects plant functional type. Ecosystems 17:673–684. https://doi.org/10.1007/s10021-014-9752-x
Wang M, Larmola T, Murphy MT, Moore TR, Bubier JL (2016) Stoichiometric response of shrubs and mosses to long-term nutrient (N, P and K) addition in an ombrotrophic peatland. Plant Soil 400:403–416. https://doi.org/10.1007/s11104-015-2744-6
Wang M, Talbot J, Moore TR (2018) Drainage and fertilization effects on nutrient availability in an ombrotrophic peatland. Sci Total Environ 621:1255–1263. https://doi.org/10.1016/j.scitotenv.2017.10.103
Weber C (1911) Das Moore. Hannoversche Geschichtsbericht 14:255–270
Wells ED (1994) Effects of planting spacing and refertilization on growth and nutrition of black spruce (Picea mariana (Mill.) B.S.P.) planted on a minerotrophic peatland in Newfoundland, Canada. Can J For Res 24:1302–1311. https://doi.org/10.1139/x94-170
Westerman DT (1975) Indices of sulfur deficiency in alfalfa. II. Plant analyses. Agron J 67:265–268. https://doi.org/10.2134/agronj1975.00021962006700020024x
Wieder RK, Vitt DH, Burke-Scoll M, Scott KD, House M, Vile MA (2010) Nitrogen and sulphur deposition and the growth of Sphagnum fuscum in bogs of the Athabasca Oil Sands Region. Alberta J Limnol 69(Suppl. 1):161–170. https://doi.org/10.3274/JL10-69-S1-16
Wieder RK, Vile MA, Scott KD, Albright CM, McMillen KJ, Vitt DH, Fenn ME (2016a) Differential effects of high atmospheric N and S deposition on bog lichen/plant tissue and porewater chemistry across the Athabasca Oil Sands Region. Environ Sci Technol 50:12630–12640. https://doi.org/10.1021/acs.est.6b03109
Wieder RK, Vile MA, Albright CM, Scott KD, Vitt DH, Quinn JC, Burke-Scoll M (2016b) Effects of altered atmospheric nutrient deposition from Alberta oil sands development on Sphagnum fuscum growth and C, N and S accumulation in peat. Biogeochemistry 129:1–19. https://doi.org/10.1007/s10533-016-0216-6
Wieder RK, Vitt DH, Vile MA, Graham JA, Hartsock JA, Fillingim H, House M, Quinn JC, Scott KD, Petix M, McMillen KJ (2019) Experimental nitrogen addition alters structure and function of a boreal bog: critical load and thresholds revealed. Ecol Monog 89:e11371. https://doi.org/10.1002/ecm.1371
Wieder RK, Vitt DH, Vile MA, Graham JA, Hartsock JA, Popma JMA, Fillingim H, House M, Quinn JC, Scott KD, Petix M, McMillen KJ (2020) Experimental nitrogen addition alters structure and function of a boreal poor fen: Implications for critical loads. Sci Total Environ 733:138619. https://doi.org/10.1016/j.scitotenv.2020.138619
Wieder RK, Vile MA, Scott KD, Albright CM, Quinn JC, Vitt DH (2021) Bog lichen/plant tissue nitrogen and sulfur concentrations as indicators of emissions from oil sands development in Alberta, Canada. Environ Monit Assess 193:208. https://doi.org/10.1007/s10661-021-08929-y
Wieder RK, Scott KD, Vile MA, Herron C (2022) Are bog plant/lichen tissue concentrations of Ca, Mg, K, and P affected by fugitive dust released from oil sands development in the Fort McMurray region of Alberta? Sci Total Environ 849:157684. https://doi.org/10.1016/j.scitotenv.2022.157684
Wieder RK, Vitt DH, Benscoter BW (2006) Peatlands and the boreal forest. In: Wieder RK, Vitt, DH (eds), Boreal peatland ecosystems. Ecological Studies 188, Springer, Berlin, pp 1–8. https://doi.org/10.1007/978-3-540-31913-9_1
Winter TC, Woo MK (1990) Plate 2. Distribution of the difference between precipitation and open-water evapotranspiration in North America. In Wolman MG, Riggs HC (eds) Surface water hydrology. The geology of North America, vol (0–1), Geological Society of America, Boulder. https://doi.org/10.1130/DNAG-GNA-O1
Živkovič T, Helbig M, Moore TR (2022) Seasonal and spatial variability of biological N2 fixation in a cool temperate bog. J Geophys Res-Biogeo 127:e2021JG006481. https://doi.org/10.1029/2021JG006481
Acknowledgements
We acknowledge the traditional, unceded territories of the Cree, Denendeh (Dënësųłinë Nëné), Michif Piyii (Métis), Beaver, Beaver Lake Cree, Bigstone Cree, and Woodland Cree First Nations on which our research sites lie. We acknowledge the systems of oppression that have historically dispossessed indigenous people of their lands and rights. We are ever vigilant that as researchers, we are but transient and humble visitors to these lands. Thanks to Jill Bubier, Andis Karlsons, Monique Heijmans, Marcel Hoosbeek, and Meng Wang for providing raw data on plant tissue chemistry, to Adam Langley, Dale Vitt, and Josh Zalewski for comments on drafts of this manuscript, to Kimberli Scott and Melanie Vile for leadership in field work, lichen/plant tissue analyses, data management, and to Kimberli Scott for archiving the Alberta data set on the Environmental Data Initiative.
Funding
This work was funded under the Oil Sands Monitoring Program, Canada (Grant # 18GRAEM07 and amendments) and is a contribution to the Program, but does not necessarily reflect the position of the Program.
Author information
Authors and Affiliations
Contributions
R.K. Wieder conceptualized, complied literature data, conducted all analyses, prepared all Figures, and wrote the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
R.K. Wieder declares no competing interests.
Additional information
Responsible Editor: Kate Lajtha.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wieder, R.K. Element stoichiometry and nutrient limitation in bog plant and lichen species. Biogeochemistry 160, 355–379 (2022). https://doi.org/10.1007/s10533-022-00968-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-022-00968-y