
Tolerance and Growth of Adult Schizothorax prenanti Exposed and Re-Exposed to Supersaturated Gas Downstream of a Dam


1
School of Energy and Power Engineering, Xihua University, Chengdu 610039, China
2
State Key Laboratory of Hydraulics and Mountain River Engineering, Sichuan University, Chengdu 610065, China
*
Author to whom correspondence should be addressed.
Water 2022, 14(16), 2501; https://doi.org/10.3390/w14162501
Submission received: 19 July 2022
/
Revised: 11 August 2022
/
Accepted: 12 August 2022
/
Published: 14 August 2022
(This article belongs to the Special Issue Response Behavior of Fish to Total Dissolved Gas)
Abstract
:Total dissolved gas (TDG) is suggested to be one possible explanation for the extensive mortality of endemic fish suffered from gas bubble disease (GBD). We investigated the effects of water depth on adult Schitzothorax prenanti susceptibility to GBD in TDG supersaturated water 11.5 km downstream of Dagangshan dam in a two-year (2016–2017) live cage study. The probability of survival significantly increased at greater depth. The mortality of fish at 1–2 m was reduced to 50% of that at 0–1 m and the mortality rate for fish at 2–3 m was reduced to 25% of that at 0–1 m. Fish that survived in-situ TDG exposure in 2016 were relocated to equilibrated water and observed for 274 d to investigate sublethal effects of GBD. The surviving fish showed a substantial reduction in growth compared to the control group. During the flood discharge period in 2017, fish were re-exposed to TDG supersaturated water after a period of recovery (274 d) in equilibrated water. The mortality rate of re-exposed fish decreased to 35% compared to newly introduced fish. Our results contribute to the protection of aquatic organisms and the improvement of eco-friendly hydroelectric dam operations in the Yangtze River.
1. Introduction
Total dissolved gas (TDG) supersaturation generated by dam spillover travels downstream for hundreds of kilometers and causes gas bubble disease (GBD) in fish [1,2]. The consequences of GBD are dependent on TDG pressure. GBD rapidly increases fish mortality in the short term at high TDG pressures and more slowly over a longer term at lower TDG pressures. Gas bubbles in the lateral line, gills, fins, eyes, or skin, as well as mortality, are symptoms of acute GBD at supersaturation levels > 110% [3,4]. TDG levels of 100–110% cause chronic GBD that results in stunted growth, secondary pathogenic infections, abnormal behavior, and some deaths [4,5]. The overwhelming majority of studies of the effects of supersaturation have been undertaken in the laboratory under controlled conditions [6]. However, these studies do not provide sufficient information for the establishment of dissolved gas management guidelines because fish in rivers downstream of the TDG source experience varying TDG levels due to factors that are difficult to simulate in the laboratory, such as dam operation, flow turbulence, and water depth [1,2]. The effects of TDG supersaturation are largely determined by both TDG level and water depth. For a given depth, the risk of mortality induced by TDG may be species-specific [7]. Knowledge of mortality rates and other effects of TDG on fish that are related to specific water depths allows us to predict the effects of dam spilling.
The degree of fish tolerance to TDG supersaturation depends on the developmental stage of the fish and its species [3,8]. Some studies have suggested that fish larvae are more sensitive to gas supersaturation than juveniles, whereas others have suggested that resistance to gas supersaturation decreases as fish develop from hatchlings to juveniles [8,9,10,11]. Adulthood is generally presumed to be the most tolerant life stage, but studies of adults in natural habitats are poorly documented due to most research being undertaken in laboratory conditions [12]. Moreover, most research has been conducted on salmonid species, and the lack of research into non-salmonid species limits our understanding of diverse sublethal outcomes of GBD because there is high variation between species in tolerance to supersaturation and the etiology of GBD [6].
The upper Yangtze River basin is an important hydroelectric resource in China; over 1000 hydropower stations have been constructed [13]. TDG supersaturation commonly occurs as a byproduct of dam operation, and it causes mortality associated with GBD at high levels in wild fish. Few measures have been taken to protect endemic fish from harm due to TDG supersaturation [6]. There is growing interest in the survival of endemic species, and we know that endemic fish are exposed to TDG supersaturation. It is therefore critical to gain more understanding of the effects of TDG supersaturation on endemic species.
Schizothorax prenanti is a valuable commercial fish inhabiting the upper reaches and tributaries of the Yangtze River [14]. The S. prenanti population has plummeted as a result of overfishing, water pollution, and the construction of hydropower stations [15], and its survival is directly threatened by TDG during the flood season. The purpose of this study was to determine the incidence of GBD in S. prenanti and the influence of TDG on adult S. prenanti survival. The subsequent growth of survivors and their future tolerance of TDG were also investigated after S. prenanti were re-exposed to TDG supersaturation. The results of this study increase our understanding of the negative effects of TDG supersaturation on S. prenanti and are a basis for the development of environmentally sustainable river management policies.
2. Materials and Methods
2.1. Study Site and Fish Acclimation
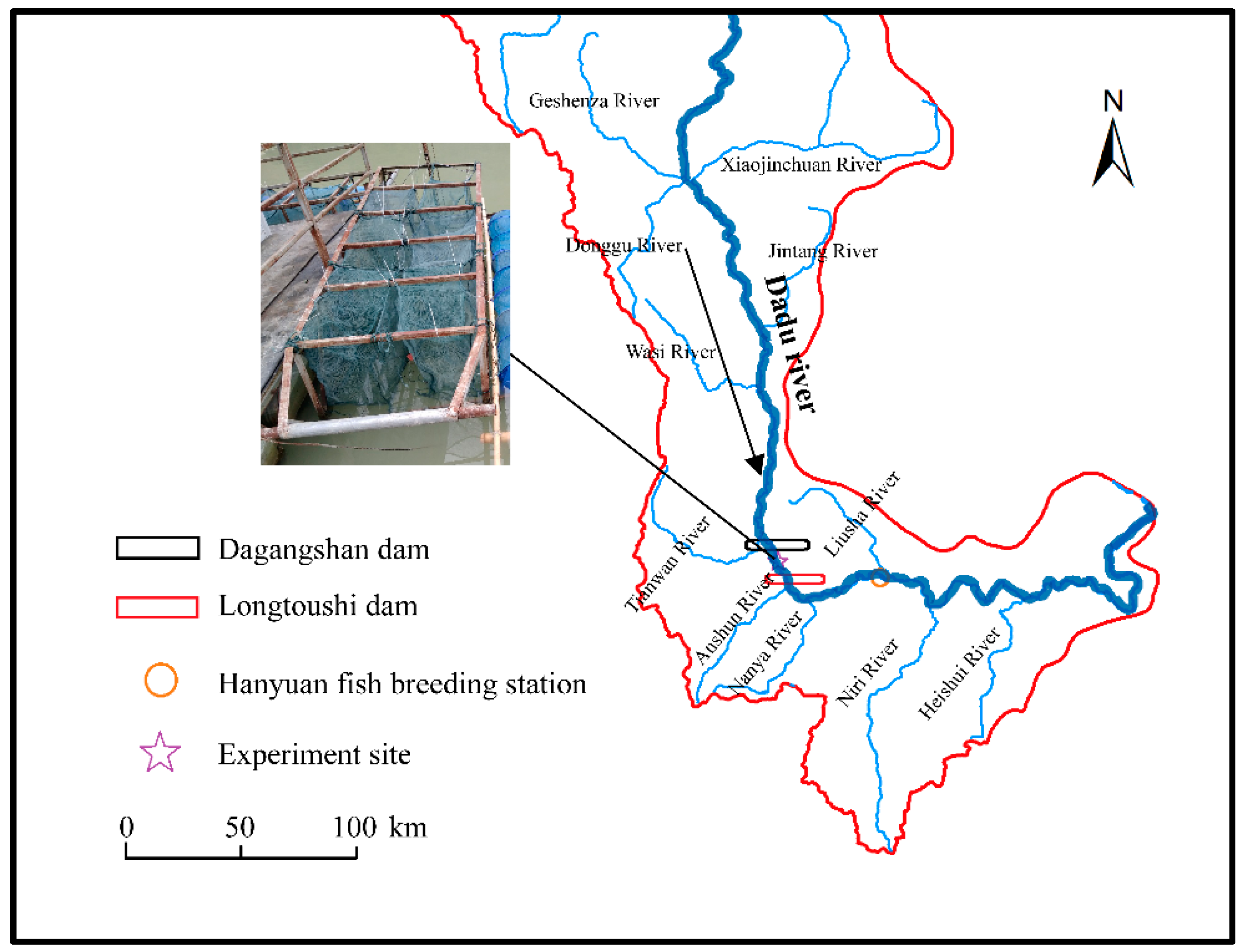
Dadu River is the longest tributary of the Minjiang River in the Yangtze River basin and is a major habitat for wild S. prenanti. The main course of the Dadu River contains a series of reservoirs created by hydroelectric dams. The experimental research of this study was conducted in the middle reach of the Dadu river, 11.6 km downstream of the Dagangshan dam, where TDG supersaturation occurs frequently from June to September as a result of the flood discharges of the Dagangshan dam. The experimental site was 57 km from the nearest fish breeding station in Hanyuan county, and the experimental fish used in this study were artificially propagated and cultured by this breeding station (Figure 1).
The experimental platform floated on the river and could be moved by pulling on rigid ropes fixed on the riverbank. A rigid frame (length 0.5 m, width 0.6 m, height 3 m) was covered with netting to form a cage and was fixed to one side of the experimental platform. The interior of the cage was subdivided into three cages by horizontal netting to confine fish to water depths of 0–1 m, 1–2 m, and 2–3 m during the period of the experiment. The entire cage could be rotated vertically by 90° so that its vertical axis was parallel to the water surface to enable us to check the mortality of the experimental fish.
The fish were transported from Hanyuan fish breeding station before the experiment and were kept temporarily in several tanks (length 1.2 m, width 0.8 m, height 0.8 m) filled with aerated river water for 24 h to relieve stress induced by transportation. Total dissolved gas was 100 ± 2% and dissolved oxygen (DO) was 7 ± 0.2 mg/L during the acclimation period.
2.2. Experiment Design
Two in situ experiments were carried out over two years during separate periods of planned flood discharge from the Dagangshan hydropower station. The TDG level and duration were typical of the downstream TDG risk to fish. First year experiment was conducted from 26 September 2016 10:00:00 to 1 October 2016 10:00:00. Prior to the experiment, 120 adult fish with average length 39.0 ± 2.0 cm and average weight 524.0 ± 54.0 g were divided equally into three groups and then placed in the cage fixed to the platform and were submerged to depths 0–1 m, 1–2 m, and 2–3 m. During the period of exposure, the entire cage was rotated by 90°, and adult fish were checked for mortality twice a day at 10:00 h and 16:00 h. Dead fish were removed, recorded, and examined for obvious external signs of GBD such as emphysema, bleeding, gill congestion, and cutaneous lesions. Typical symptoms of GBD were inspected under a photomicroscope (Guangzhou Micro-Shot Optical Technology Co., Ltd., Guangzhou, China). TDG and DO of the surface water layer at the experimental site were manually monitored during the experiment using a Point Four tracker (Point Four Systems Inc., Coquitlam, BC, Canada) and a YSI 6600 recirculating system monitor (YSI Inc., Yellow Springs, OH, USA).
On 1 October 2016 10:00:00, when the flood discharge was over, the survivors in the three cage sections were collected with scooping nets and placed into three tanks (length 1.2 m, width 0.8 m, height 0.8 m) that were then immediately transported to the Hanyuan breeding station to allow the fish to recover. Three culture tanks were used in the Hanyuan breeding station to contain the survivors of the three treatment groups. The culture tank consisted of a pool with radius 2 m and height 1.5 m, with a plastic pipe radius 15 cm and height 1 m in the center of the pool to allow river water from the Dadu river to be continuously introduced into the pool to overflowing and so control the water level to remain at 1 m. Twenty healthy live adults from the breeding station were selected as a control group and kept in another identical culture tank. After 65 d, fish survival was checked, and dead fish were removed from the culture tanks. Thereafter, each treatment group was fed a commercial diet (Wudi Yideyuan Biological Technology Co. Ltd., Shandong, China) containing 33–35% protein twice a day. TDG levels were 100.0 ± 1.2%, and dissolved oxygen (DO) was in the range 8.4–9.5 mg/L in the culture tanks. On day 65, all fish from the three treatment and control groups were individually weighed to measure their initial weight, and they were then weighed on days 113, 184, 228, and 274 to monitor change in weight.
On day 280 (3 July 2017), a sample of twenty fish from each experimental tank (0–1 m, 1–2 m, 2–3 m depth initial TDG exposure) together with 90 newly-purchased fish from Hanyuan fish breeding station with average length 35.0 ± 6.0 cm and average weight 578.0 ± 25.8 g was quickly transferred to the experimental cage to be temporarily cultivated to investigate re-exposure to TDG. Temporary cultivation was identical to that of the 2016 exposure experiment. Fish that survived in the 2016 experiment were placed in the same water layer that they had been raised in 2016. The newly acquired fish, which had not participated in the 2016 experiment, were also distributed in 0–1 m, 1–2 m, and 2–3 m water depths as a control. The 2017 exposure experiment ran from 4 July 2017 10:00:00 to 10 July 2017 10:00:00, and the experimental protocol was the same as for the 2016 exposure experiment.
2.3. Statistical Analysis
The Cox proportional hazards model was used as the hazard function h(t). It represents the likelihood of an incident occurring at time t, given that the subject has survived until that time, as:
where H0 is the baseline hazard probability when all coefficients b is zero; the covariates x are all the factors that affect the time until an event; and b is the hazard ratio for the covariates, indicating the size of the effect of the covariates on the dependent variable. The Cox proportional hazards model was run using the R coxph function with random intercepts specified as offsets to calculate coefficient standard errors and confidence intervals.
Experimental fish were exposed to TDG supersaturation during the flood periods in 2016 and 2017. The data collected from these exposures were used to fit a Cox proportional hazards model to create a function to represent the risk of TDG-induced mortality associated with water depth. The model was also used to analyze the influence of re-exposure on fish mortality in TDG supersaturated water in 2017.
Fish with GBD were placed in saturated water for observation on day 6. No more deaths had occurred by day 42. The average weights of fish from the treatment and control groups were calculated from bulk weighing on day 65 and were taken to be the initial weights. All individuals in each group were weighed (W, g) and recorded on days 113, 184, 228, and 272. Relative weight growth rate (RGR, g/d) was calculated using the equation:
where W1 and Wi are, respectively, the individual and mean weights (g) of the fish at days t1 (65 d) and ti (113, 184, 228, and 272 d).
RGR = (Wi − W1)/(ti − t1)
3. Results
3.1. Survival Characteristics of Fish Exposed to TDG Supersaturation
During the flood release at Dagangshan hydropower in September 2016, the average TDG of the surface water was 121% with a range 115–127% and DO vary between 111.2% and 133.3%. In July 2017, TDG at the study site ranged from 111% to 121% and DO vary between 107.0% and 126.5%.
Signs of GBD were found on most of the dead fish, indicating that GBD was a major cause of death (Table 1), and typical symptoms of GBD were shown in Figure 2. The most common GBD symptoms in dead fish from each treatment group were analyzed. In 2016, 21 S. prenanti were observed with emphysema on the dorsal fin in the 24 dead fish in the 0–1 m group; 7 S. prenanti were observed with bleeding in the pectoral fins and anal fins in the 1–2 m group; and 5 S. prenanti were observed with fin bleeding in the 2–3 m group. The prevalence of GBD symptoms in the 0–1 m was greater than in the 1–2 m and 2–3 m groups. Dorsal fin emphysema was observed in 17 S. prenanti in the 0–1 m group and in 12 S. prenanti in the 1–2 m group. Pelvic fin emphysema was observed in 3 S. prenanti in the 2–3 m group. There were individual variations in the external symptoms of GBD. Some dead fish were found without GBD symptoms; death could have been due to transportation or extrusions in the cage.
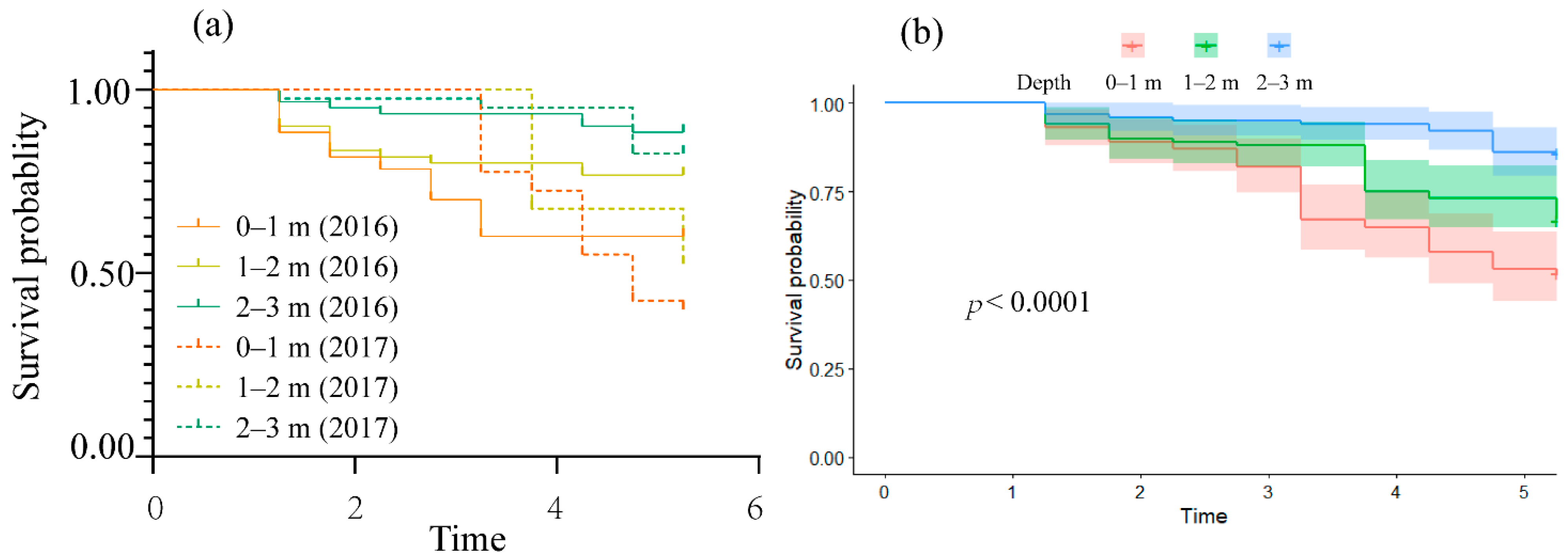
The relationship between exposure time and mortality is shown in Figure 3a. The final mortality of adult S. prenanti were 40% (0–1 m), 23% (1–2 m), and 12% (2–3 m) in 2016 and 60% (0–1 m), 47% (1–2 m), and 17% (2–3 m) in 2017. These results indicate that increase in hydrostatic pressure at greater depths caused the increase in survival probabilities.
All individual fish exposed to supersaturated water were analyzed using the Cox proportional hazards model. The absolute value of a calculation was used to indicate the effect of TDG supersaturated water depth on adult S. prenanti (Figure 3b, n = 150). The coefficients calculated from the Cox proportional hazards fitting model indicated that water depth significantly reduced the risk of death (coeff = −0.7, p < 0.01). The mortality rate of fish in the 1–2 m group decreased by 50% compared to the 0–1 m group and further decreased in the 2–3 m treatment group by 50% compared to the 1–2 m group (hazard ratio, HR, =0.5, 95% confidence interval, CI, =0.38–0.65).
3.2. Growth Characteristics of Fish after Exposed to TDG Supersaturation
The final mortality of fish in 0–1 m, 1–2 m, and 2–3 m were, respectively, 58% (27 d), 38% (42 d), and 28% (13 d) in the recovery period. No deaths occurred after 42 d of recovery, suggesting that any indirect effects of TDG supersaturation that affected mortality disappeared over time, but the possibility of other chronic effects was not eliminated (Figure 4a). Significantly lower rates of increase in weight were observed in adult fish exposed to TDG than in the controls on day 113 of recovery (p < 0.05; Figure 4b). The average rate of increase in weight of adult fish on day 184 of recovery decreased from 1.79 g/d in the control group to 1.08 g/d (1–2 m) and 0.60 g/d (0–1 m) in adult fish (p < 0.05), but no significant difference was found in the 2–3 m group compared with the control group. There were no significant differences between groups on days 228 and 272 of recovery.
3.3. Survival Characteristics of Fish Re-Exposed to TDG Supersaturation
Figure 5 shows the mortality in 2017 of the re-introduced S. prenanti that survived in the 2016 experiment and newly introduced fish that did not participate in the 2016 experiment. The survival rate of re-introduced fish after experiencing flood discharge from the Dagangshan dam in 2017 decreased by 40% in the 0–1 m group, and the survival rate of the newly introduced fish decreased by 60% in the 0–1 m group. The survival rate of re-introduced S. prenanti in the 1–2 m group decreased by 15%, and the survival rate of newly introduced fish in the 1–2 m group decreased by 47%. Re-introduced fish in the 2–3 m group had a 100% survival rate (Figure 5a).
The results of the Cox proportional hazards model (Figure 5b) suggested that exposure significantly increased adult fish tolerance to supersaturation (Coeff = −1.06, p < 0.01). The mortality rate of re-exposure for adult fish was 35% of that of initial exposure (HR = 0.35, 95% CI = 0.18–0.68).
4. Discussion
4.1. Effect of Water Depth on Fish Survival
Several studies have found that clinical symptoms of GBD presented at elevated levels of TDG supersaturation [3,4]. We found that both GBD and death occurred, which suggests that TDG levels (about 121% in 2016, 117% in 2017) exceeded some critical level for S. prenanti. Studies have shown that the risk of mortality was associated with both water depth and the severity of GBD [6,7,16]. We found that increased water depth significantly decreased mortality. Increase in water depth increases hydrostatic pressure, thus inhibiting embolism formation in the blood, preventing death and promoting the re-dissolution of gas embolisms by re-establishing blood flow to ischemic tissue [16,17]. The prevalence and severity of GBD in S. prenanti exposed to TDG also varies greatly among individual fish in a river setting. An explanation for this is that the formation and development of externally observable GBD symptoms may be related to breathing frequency, epidermal tension, internal cardiovascular pressure, and tissue oxygen consumption [12,17].
Results for fish in the surface layer (0–1 m) represent the worst-case scenario in which there is limited available depth for fish. Actual TDG exposure is usually lower than exposure to TDG at the surface because fish often swim at some depth due to environmental and physiological influences [18]. Wang et al. (2015) found that S. prenanti migrated to depths > 3 m after 1 h of exposure to TDG-supersaturated water but they did not determine if S. prenanti stayed in deep TDG-supersaturated water for extended periods of time [19]. Depth migration of individual fish appears to be strongly influenced by diel, which influences exposure to TDG supersaturation [20]. This pattern of depth distribution may also be related to the presence of predators in TDG supersaturated water, implying that wild S. prenanti stay at the surface for half of the day and migrate to deeper waters for the other half [21,22]. Johnson et al. (2007) obtained data on depth distribution of fish by telemetry and calculated the spatial variability of saturated dissolved gases and found that adult fish are at a depth < 3 m for more than half the time [23]. Fish in cages do not have the same opportunity to choose the most favorable water depths as wild fish, and they thus have greater mortality rates than wild fish. To determine if the limited monitoring data we collected on TDG levels during spilling in 2016 and 2017 is representative of gas levels over the long term, we would require a longer observational data time series.
Some fish are likely to encounter TDG supersaturation on their migratory path because spawning migration and the flood season coincide [24]. S. prenanti is a potamodromous fish species that tends to move close to shore where the water is shallower because lower flow rates mean lower energy costs [25,26]. Suitable water depth for S. prenanti breeding is 0.15–1.5 m, which is not deep enough to prevent the onset of GBD [27]. Physical damage from GBD also changes swimming behavior, predation vulnerability, and positive buoyancy, and thus decreases fish survival rates [28,29]. These adverse effects of TDG supersaturation on S. prenanti during spawning periods may be important reasons for the decline of this and other wild species.
4.2. Growth of Fish after Exposure to TDG Supersaturation
TDG supersaturation may or may not decrease the growth rate of fish depending on the species, TDG level, and exposure time [30,31,32]. This study showed that the adverse effects on growth of S. prenanti were mainly restricted to 0–1 m and 1–2 m in supersaturated water. One explanation for this is that gas bubbles on the epidermis and other organs and tissues of the fish acted as a trigger, thus consuming more energy to relieve oxidative stress [32,33]. We found that the growth of surviving fish was significantly inhibited after a succession of deaths due at least partially to GBD. One explanation for this phenomenon is that physical damage from GBD (exophthalmos, visceral injuries, and gill gas embolism) makes fish more vulnerable and susceptible to bacterial infection; thus, GBD can be indirectly responsible for fish mortality [34]. Additionally, stress from transportation and handling can also explain fish mortality absent signs of GBD. Fish may be also affected by bubbles that inhibit blood flow, which increases fish sensitivity to environmental or physiological changes. The high follow-up mortality rate we found implies that S. prenanti suffered serious TDG damage, which explains the significant reduction in growth.
Growth was not significantly inhibited by 184 d in the fish that experienced TDG supersaturation stress in 2–3 m depth, which implies that 2–3 m depth greatly mitigates the potential effects of GBD. Moreover, surviving S. prenanti showed no difference in growth rate after remaining in saturated water for 228 d, an observation that suggests that adult S. prenanti have an inherent ability to recover from GBD. However, we note that the fish growth experiment was performed at the fish breeding station where environmental factors (such as flow velocity and water temperature) differed from those found in the river, so there may be differences in the effects of GBD on fish growth between fish kept in artificial surroundings and free-living wild fish.
4.3. Survival of Fish Re-Exposed to TDG Supersaturation
Equilibrated water gives fish exposed to TDG supersaturation a chance to recover from GBD [16]. Cao et al. (2020) found that the survival time of yellow catfish (Pelteobagrus fulvidraco) increased as recovery time rose from 3 h to 9 h at relatively high TDG levels (130%, 135%, and 140%) [3]. This result supports our finding that individuals with stronger tolerance were screened after TDG exposure and long-term recovery. However, re-exposure did not reveal any change in resistance in other studies. Wang et al. (2015) found that half lethal time for juvenile S. prenanti intermittently exposed to TDG supersaturation for periods of 2–6 h was not significantly different from that of fish continuously exposed to TDG supersaturation, which is inconsistent with the findings of our study [19]. However, the recovery time between exposures allowed in the Wang et al. study was much less than we allowed in this study, and the degree of TDG supersaturation was greater than in our study. We hypothesize that the extended survival time of fish re-exposed to TDG supersaturation is influenced by TDG supersaturation level, exposure time, and recovery time.
Our finding suggests that TDG supersaturation is an adaptive pressure on fish that selects for genes that are expressed in ways that result in a higher tolerance to supersaturation. Gray et al. (1982) hypothesized that higher tolerance of TDG supersaturation in fish in the Columbia River, compared to controls from Italy, was due to the long history of supersaturated TDG exposure in Columbia River water, and that this suggested that species tolerance may be influenced by exposure acting as a form of genetic screening [35]. Our study was conducted for only two years, and we have no long-term data to support this hypothesis. But these examples of adaptation provide some support for our claim that does not depend on long term data. The classic work of Dobzhansky showed that in wild fruit flies, the genetic composition of a population varied widely over a season [36], and Grants et al. (2009) have shown that finches on the Galapagos can certainly adapt rapidly to environmental change and may even speciate over a few generations [37]. These examples show that organisms must be able to adapt quickly to environmental change in order to avoid extinction.
4.4. Future Study
A succession of high dams exceeding 200 m have been constructed in the upper reaches of the Yangtze River in China. The effects of TDG supersaturation caused by the operation of these dams on downstream fish have become increasingly noticeable [38,39,40]. Most research into in situ TDG effects has been conducted on salmonids, and research into non-salmonid species is slight in comparison [6]. This imbalance suggests that, with the exception of salmonids, existing data are not sufficient to provide a sound theoretical basis for mitigating the adverse ecological effects of TDG supersaturation on other fish species. This study shows that the Cox hazard proportional model is suitable for predicting TDG-induced mortality in S. prenanti and other species. We found that, critically, the latent effects of TDG supersaturation on growth and survival are likely to be nonnegligible for adults that survive exposure to TDG supersaturation. More data are needed from cage experiments to determine the tolerance to TDG supersaturation of specific endemic fish at current dam discharge schedules, and an extended observation period is needed to provide more details on the subsequent survival and growth of fish with GBD. Future studies could also explore the molecular mechanisms of fish damage due to TDG supersaturation and whether they are heritable to determine the extent to which the local environment of endemic fish affects their tolerance to TDG supersaturation.
Hydrostatic pressure is known to protect fish from damage by total dissolved gas (TDG) supersaturation, and dam construction has resulted in shallower spawning depths than in the past, thus spawning fish are more vulnerable to the effects of TDG supersaturation [27]. We recommend that more in-situ experiments on TDG tolerance should be carried out on migratory fish during the period of flood discharge. Moreover, acoustic telemetry can be used to investigate changes in depth-use of migratory fish in response to ambient TDG supersaturation in order to accurately assess the threat of TDG to migratory fish.
Although this study focused on TDG risk associated with operation of artificial dams, TDG supersaturation widely exists in the natural world. For instance, Niagara Falls was observed to generate TDG levels between 120 and 130% [41]. Nitrogen supersaturation occurred in four shallow lakes in southwestern Manitoba because of the large drop in lake volume brought on by ice formation, which resulted in a high trout mortality [42]. The heavy algal bloom was also found to cause exceptionally high supersaturation of DO, and finally resulted in occurrence of GBD in various wild fish species [43]. Future studies should pay more attention to the survival of fish under gas supersaturation caused by natural stressors.
Author Contributions
Conceptualization, Y.W.; methodology, Q.Y.; software, R.L..; validation, Q.Y.; formal analysis, Q.J.; investigation, Q.Y. and Q.J.; resources, K.L.; data curation, Y.W.; writing—original draft preparation, Q.Y.; writing—review and editing, Y.W.; visualization, R.L.; supervision, Y.W.; project administration, K.L.; funding acquisition, K.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Yangtze River Water Science Research Joint Fund (U2240212) and the National Natural Science Foundation of China (51809186, 52179075).
Institutional Review Board Statement
The animal study proposal was approved by the Ethics Committee for Animal Experiments of Sichuan University. All experimental procedures were performed in accordance with the Regulations for the Administration of Affairs Concerning Experimental Animals approved by the State Council of the People’s Republic of China.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets used and analyzed during the current study are available from the corresponding authors upon reasonable request.
Acknowledgments
We are also grateful for the thoughtful comments provided by two anonymous reviewers.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yuan, Y.; Wei, Q.; Yuan, Q.; Wang, Y.; Liang, R.; Li, K.; Zhu, D.Z. Impact of TDG supersaturation on native fish species under different hydropower flood discharge programs. Aquat. Toxicol. 2021, 237, 105898. [Google Scholar] [CrossRef]
- Xue, S.; Li, K.; Liang, R.; Cao, L.; Wang, Y.; Li, Y.; Feng, J. In situ study on the impact of total dissolved gas supersaturation on endemic fish in the Upper Yangtze River. River Res. Appl. 2019, 35, 1511–1519. [Google Scholar] [CrossRef]
- Cao, C.; Deng, Y.; Yin, Q.; Li, N.; Liu, X.; Shi, H.; Yang, Y.; Xu, L. Effects of continuous acute and intermittent exposure on the tolerance of juvenile yellow catfish (Pelteobagrus fulvidraco) in total dissolved gas supersaturated water. Ecotoxicol. Environ. Saf. 2020, 201, 110855. [Google Scholar] [CrossRef] [PubMed]
- Mesa, M.G.; Weiland, L.K.; Maule, A.G. Progression and severity of gas bubble trauma in juvenile salmonids. Am. Fish. Soc. 2000, 129, 174–185. [Google Scholar] [CrossRef]
- Tsai, J.Y.; Felt, S.A.; Bouley, D.M.; Green, S.L. Acute and chronic outcomes of gas-bubble disease in a colony of African Clawed Frogs (Xenopus laevis). Comp. Med. 2017, 67, 4–10. [Google Scholar] [PubMed]
- Pleizier, N.K.; Algera, D.; Cooke, S.J.; Brauner, C.J. A meta-analysis of gas bubble trauma in fish. Fish Fish. 2020, 21, 1175–1194. [Google Scholar] [CrossRef]
- Yuan, Q.; Wang, Y.; Liang, R.; Feng, J.; Li, Y.; An, R.; Li, K. Field observations of the lethality characteristics of endangered and endemic fish under the stress of total dissolved gas supersaturation. River Res. Appl. 2021, 37, 1156–1167. [Google Scholar] [CrossRef]
- Geist, D.R.; Linley, T.J.; Cullinan, V.; Deng, Z. The effects of total dissolved gas on chum salmon fry survival, growth, gas bubble disease, and seawater tolerance. N. Am. J. Fish. Manag. 2013, 33, 200–215. [Google Scholar] [CrossRef]
- Meekin, T.K.; Turner, B.K. Tolerance of salmonid eggs, juveniles and squawfish to supersaturated nitrogen. In Nitrogen Supersaturation Investigations in the Mid-Columbia River; Washington Department of Fisheries Technical Report: Olympia, WA, USA, 1974; Volume 12, pp. 75–126. [Google Scholar]
- Harvey, H.H.; Cooper, A.C. Origin and treatment of a supersaturated river water. Prog. Rep. 1962, 9, 7. [Google Scholar]
- Nebeker, A.V.; Andros, J.D.; McCrady, J.K.; Stevens, D.G. Survival of steelhead trout (Salmo gairdneri) eggs, embryos, and fry in air-supersaturated water. J. Fish. Board Can. 1978, 35, 261–264. [Google Scholar] [CrossRef]
- Weitkamp, D.E.; Katz, M. A review of dissolved gas supersaturation literature. Trans. Am. Fish. Soc. 1980, 109, 659–702. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, Y.; Zhang, R.; Ouyang, S.; Wang, X.; Liao, X. Integrated optimization of hydroelectric energy in the upper and middle Yangtze River. Renew. Sust. Energ. Rev. 2015, 45, 481–512. [Google Scholar] [CrossRef]
- Ding, R. (Ed.) The Fishes of Sichuan, China; Sichuan Publishing House of Science and Technology: Chengdu, China, 1194; p. 641. (In Chinese) [Google Scholar]
- Liu, J. A quantitative analysis on threat and priority of conservation order of the endemic fishes in upper reaches of the Yangtze River. Environ. Sci. Sin. 2004, 24, 395–399. [Google Scholar]
- Pleizier, N.K.; Nelson, C.; Cooke, S.J.; Brauner, C.J. Understanding gas bubble trauma in an era of hydropower expansion: How do fish compensate at depth? Can. J. Fish. Aquat. Sci. 2020, 77, 556–563. [Google Scholar] [CrossRef]
- Elston, R.; Colt, J.; Abernethy, S.; Maslen, W. Gas bubble reabsorption in Chinook salmon: Pressurization effects. J. Aquat. Anim. Health 1997, 9, 317–321. [Google Scholar] [CrossRef]
- Algera, D.A.; Kamal, R.; Ward, T.D.; Pleizier, N.K.; Brauner, C.J.; Crossman, J.A.; Leake, A.; Zhu, D.Z.; Power, M.; Cooke, S.J. Exposure risk of fish downstream of a hydropower facility to supersaturated total dissolved gas. Water Resour. Res. 2021, 58, e2021WR031887. [Google Scholar] [CrossRef]
- Wang, Y.; Li, K.; Li, J.; Li, R.; Deng, Y. Tolerance and avoidance characteristics of Prenant’s schizothoracin Schizothorax prenanti to total dissolved gas supersaturated water. N. Am. J. Fish. Manag. 2015, 35, 827–834. [Google Scholar] [CrossRef]
- Brege, D.A.; Absolon, R.F.; Graves, R.J. Seasonal and diel passage of juvenile salmonids at John Day Dam on the Columbia River. N. Am. J. Fish. Manage. 1996, 16, 659–665. [Google Scholar] [CrossRef]
- Reddin, D.G.; Downton, P.; Friedland, K.D. Diurnal and nocturnal temperatures for Atlantic salmon postsmolts (Salmo salar L.) during their early marine life. Fish. Bull. 2006, 104, 415–428. [Google Scholar]
- Davidsen, J.G.; Manel-La, N.P.; Økland, F.; Diserud, O.H.; Thorstad, E.B.; Finstad, B.; Sivertsgård, R.; McKinley, R.S.; Rikardsen, A.H. Changes in swimming depths of Atlantic salmon Salmo salar post-smolts relative to light intensity. J. Fish Biol. 2008, 73, 1065–1074. [Google Scholar] [CrossRef]
- Johnson, E.L.; Clabough, T.S.; Peery, C.A.; Bennett, D.H.; Bjornn, T.C.; Caudill, C.C.; Richmond, M.C. Estimating adult Chinook salmon exposure to dissolved gas supersaturation downstream of hydroelectric dams using telemetry and hydrodynamic models. River Res. Appl. 2007, 23, 963–978. [Google Scholar] [CrossRef]
- Beeman, J.W.; Maule, A.G. Migration depths of juvenile Chinook salmon and steelhead relative to total dissolved gas supersaturation in a Columbia River reservoir. Trans. Am. Fish. Soc. 2006, 135, 584–594. [Google Scholar] [CrossRef]
- Keefer, M.L.; Peery, C.A.; Ringe, R.R.; Bjornn, T.C. Regurgitation rates of intragastric radio transmitters by adult Chinook salmon and steelhead during upstream migration in the Columbia and Snake rivers. N. Am. J. Fish. Manag. 2004, 24, 47–54. [Google Scholar] [CrossRef]
- Keefer, M.L.; Caudill, C.C.; Peery, C.A.; Bjornn, T.C. Route selection in a large river during the homing migration of Chinook salmon (Oncorhynchus tshawytscha). Can. J. Fish. Aquat. Sci. 2006, 63, 1752–1762. [Google Scholar] [CrossRef]
- Yu, Z.; Zhang, J.; Zhao, J.; Peng, W.; Fu, Y.; Wang, Q.; Zhang, Y. A new method for calculating the downstream ecological flow of diversion-type small hydropower stations. Ecol. Indic. 2021, 125, 107530. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; An, R.; Li, K. Effects of total dissolved gas supersaturation on the swimming performance of two endemic fish species in the upper Yangtze River. Sci. Rep. 2018, 8, 10063. [Google Scholar] [CrossRef]
- Stephenson, J.R.; Gingerich, A.J.; Brown, R.S.; Pflugrath, B.D.; Deng, Z.; Carlson, T.J.; Langeslay, M.J.; Ahmann, M.L.; Johnson, R.L.; Seaburg, A.G. Assessing barotrauma in neutrally and negatively buoyant juvenile salmonids exposed to simulated hydro-turbine passage using a mobile aquatic barotrauma laboratory. Fish. Res. 2010, 106, 271–278. [Google Scholar] [CrossRef]
- Dawley, E.M.; Ebel, W.J. Effects of various concentrations of dissolved atmospheric gas on juvenile chinook salmon and steelhead trout. Fish. Bull. 1975, 73, 777–796. [Google Scholar]
- Gunnarsli, K.S.; Toften, H.; Mortensen, A. Effects of nitrogen gas supersaturation on growth and survival in juvenile Atlantic cod (Gadus morhua L.). Aquaculture 2008, 283, 175–179. [Google Scholar] [CrossRef]
- Liu, X.Q.; Li, K.F.; Du, J.; Li, J.; Li, R. Growth rate, catalase and superoxide dismutase activities in rock carp (Procypris rabaudi Tchang) exposed to supersaturated total dissolved gas. J. Zhejiang Univ. Sci. B 2011, 12, 909–914. [Google Scholar] [CrossRef]
- Jiang, W.; Wu, S. Biochemical responses and survival of rock carp (Procypris rabaudi) to total dissolved gas supersaturated water. Ichthyol. Res. 2015, 62, 171–176. [Google Scholar]
- Huchzermeyer, K.D.A. Clinical and pathological observations on Streptococcus sp. infection on South African trout farms with gas supersaturated water supplies. Onderstepoort J. Vet. Res. 2003, 70, 95–105. [Google Scholar] [PubMed]
- Gray, R.H.; Page, T.L.; Saroglia, M.G.; Bronzi, P. Comparative tolerance to gas supersaturated water of carp, Cyprinus carpio, and black bullhead, Ictalurus melas, from the USA and Italy. J. Fish Biol. 1982, 20, 223–227. [Google Scholar] [CrossRef]
- Dobzhansky, T. Genetics and the Origin of Species; Columbia University Press: New York, NY, USA, 1982. [Google Scholar]
- Grant, P.R.; Grant, B.R. The secondary contact phase of allopatric speciation in Darwin’s finches. Proc. Natl. Acad. Sci. USA 2009, 106, 20141–20148. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, Q.; Wang, Y.; Zhu, D.Z.; Liang, R.; Qin, L.; Li, R.; Ji, Q.; Li, K. River habitat assessment and restoration in high dam flood discharge systems with total dissolved gas supersaturation. Water Res. 2022, 221, 118833. [Google Scholar] [CrossRef] [PubMed]
- Ji, Q.; Li, K.; Wang, Y.; Liang, R.; Feng, J.; Yuan, Q.; Zhang, P.; Zhu, D.Z. Total dissolved gases induced tolerance and avoidance behaviors in pelagic fish in the Yangtze River, China. Ecotoxicol. Environ. Saf. 2021, 216, 112218. [Google Scholar] [CrossRef] [PubMed]
- Ji, Q.; Li, K.; Wang, Y.; Liang, R.; Feng, J.; Li, R.; Zhu, D.Z. Swimming performance of a pelagic species in the Yangtze River under different exposure modes of the total dissolved gas supersaturation. Conserv. Physiol. 2022, 10, coac047. [Google Scholar] [CrossRef]
- Weitkamp, D.E. Total Dissolved Gas Supersaturation Biological Effects, Review of Literature 1980–2007; Parametrix: Bellevue, WA, USA, 2008. [Google Scholar]
- Mathias, J.A.; Barica, J. Gas supersaturation as a cause of early spring mortality of stocked trout. Can. J. Fish. Aquat. Sci. 1985, 42, 268–279. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, N.; Lee, J.Y.; Kang, H.; Sung, M.; Yu, Y.B.; Kim, K.; Je, J.Y.; Kim, H.W.; Kang, J.C.; et al. Case report: Mass mortality of olive flounder (Paralichthys olivaceus) caused by acute gas bubble disease. J. Fish Pathol. 2021, 34, 255–259. [Google Scholar]
Figure 1.
The Dadu river system, showing the positions of the Dagangshan dam, experiment site, Longtoushi dam and Hanyuan fish breeding station; the arrow indicates the direction of river flow.
Figure 1.
The Dadu river system, showing the positions of the Dagangshan dam, experiment site, Longtoushi dam and Hanyuan fish breeding station; the arrow indicates the direction of river flow.

Figure 2.
Typical symptoms of GBD; bubbles were observed in (a) the pectoral fin, (b) the anal fin, and (c) the dorsal fin, while (d) shows corneal emphysema.
Figure 2.
Typical symptoms of GBD; bubbles were observed in (a) the pectoral fin, (b) the anal fin, and (c) the dorsal fin, while (d) shows corneal emphysema.

Figure 3.
Survival probability of S. prenanti confined in 0–1 m, 1–2 m, 2–3 m water depths as a function of exposure duration during the dam spill season in 2016 and 2017; (a) survival curve for six scenarios; (b) survival probability with 95% confidence interval using Cox proportional hazard model for S. prenanti at various water depths.
Figure 3.
Survival probability of S. prenanti confined in 0–1 m, 1–2 m, 2–3 m water depths as a function of exposure duration during the dam spill season in 2016 and 2017; (a) survival curve for six scenarios; (b) survival probability with 95% confidence interval using Cox proportional hazard model for S. prenanti at various water depths.

Figure 4.
TDG-induced effects on fish survival and growth: (a) survival curves of adult S. prenanti for the initial period of TDG supersaturated exposure (6 d) and subsequent equilibrated water observation (59 d), with the control group in the equilibrated water for the entire observation period (65 d); (b) changes in the relative weight growth rate of S. prenanti in equilibrated water after 113 d, 184 d, 228 d, and 272 d of recovery, with the average weight of fish from the treatment group and the control group at 65 d considered to be the initial weight. Asterisks indicate significance for p < 0.05.
Figure 4.
TDG-induced effects on fish survival and growth: (a) survival curves of adult S. prenanti for the initial period of TDG supersaturated exposure (6 d) and subsequent equilibrated water observation (59 d), with the control group in the equilibrated water for the entire observation period (65 d); (b) changes in the relative weight growth rate of S. prenanti in equilibrated water after 113 d, 184 d, 228 d, and 272 d of recovery, with the average weight of fish from the treatment group and the control group at 65 d considered to be the initial weight. Asterisks indicate significance for p < 0.05.

Figure 5.
Survival characteristics of re-introduced and newly introduced adult S. prenanti in 0–1 m, 1–2 m, and 2–3 m depths: (a) survival curve for six scenarios; (b) survival probability with 95% confidence interval using the Cox proportional hazards model for the exposure group and the re-exposure group.
Figure 5.
Survival characteristics of re-introduced and newly introduced adult S. prenanti in 0–1 m, 1–2 m, and 2–3 m depths: (a) survival curve for six scenarios; (b) survival probability with 95% confidence interval using the Cox proportional hazards model for the exposure group and the re-exposure group.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Prevalence of S. prenanti with different GBD symptoms during the TDG supersaturation exposure experiments in 2016 and 2017; 40 adult S. prenanti were used in each scenario in 2016 while 30 adult fish were used in each scenario in 2017.
Table 1.
Prevalence of S. prenanti with different GBD symptoms during the TDG supersaturation exposure experiments in 2016 and 2017; 40 adult S. prenanti were used in each scenario in 2016 while 30 adult fish were used in each scenario in 2017.
| GBD Signs | 2016 | 2017 | ||||
|---|---|---|---|---|---|---|
| 0–1 m | 1–2 m | 2–3 m | 0–1 m | 1–2 m | 2–3 m | |
| Dorsal fin emphysema | 53% | 5% | 5% | 57% | 40% | 3% |
| Tail fin emphysema | 50% | 10% | 3% | 17% | 17% | 3% |
| Tail fin bleeding | 45% | 15% | 13% | 7% | 10% | 0% |
| Pectoral fin bleeding | 43% | 18% | 3% | 13% | 10% | 7% |
| Anal fin emphysema | 43% | 8% | 0% | 17% | 17% | 3% |
| Anal fin bleeding | 38% | 18% | 5% | 3% | 7% | 0% |
| Pelvic fin bleeding | 35% | 8% | 3% | 3% | 3% | 0% |
| Pectoral fin bleeding | 25% | 8% | 3% | 7% | 3% | 0% |
| Dorsal fin bleeding | 25% | 10% | 5% | 3% | 10% | 0% |
| Gill congestion | 3% | 0% | 3% | 3% | 3% | 0% |
| Cutaneous lesions | 0% | 0% | 0% | 3% | 0% | 0% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yuan, Q.; Li, K.; Wang, Y.; Ji, Q.; Liang, R. Tolerance and Growth of Adult Schizothorax prenanti Exposed and Re-Exposed to Supersaturated Gas Downstream of a Dam. Water 2022, 14, 2501. https://doi.org/10.3390/w14162501
AMA Style
Yuan Q, Li K, Wang Y, Ji Q, Liang R. Tolerance and Growth of Adult Schizothorax prenanti Exposed and Re-Exposed to Supersaturated Gas Downstream of a Dam. Water. 2022; 14(16):2501. https://doi.org/10.3390/w14162501
Chicago/Turabian StyleYuan, Quan, Kefeng Li, Yuanming Wang, Qianfeng Ji, and Ruifeng Liang. 2022. "Tolerance and Growth of Adult Schizothorax prenanti Exposed and Re-Exposed to Supersaturated Gas Downstream of a Dam" Water 14, no. 16: 2501. https://doi.org/10.3390/w14162501
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.