
Description of Aliinostoc alkaliphilum sp. nov. (Nostocales, Cyanobacteria), a New Bioactive Metabolite-Producing Strain from Salina Verde (Pantanal, Brazil) and Taxonomic Distribution of Bioactive Metabolites in Nostoc and Nostoc-like Genera

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Strains, Microbiological Media and Growth Conditions
2.1.1. Cyanobacteria
2.1.2. Bacteria and Fungi
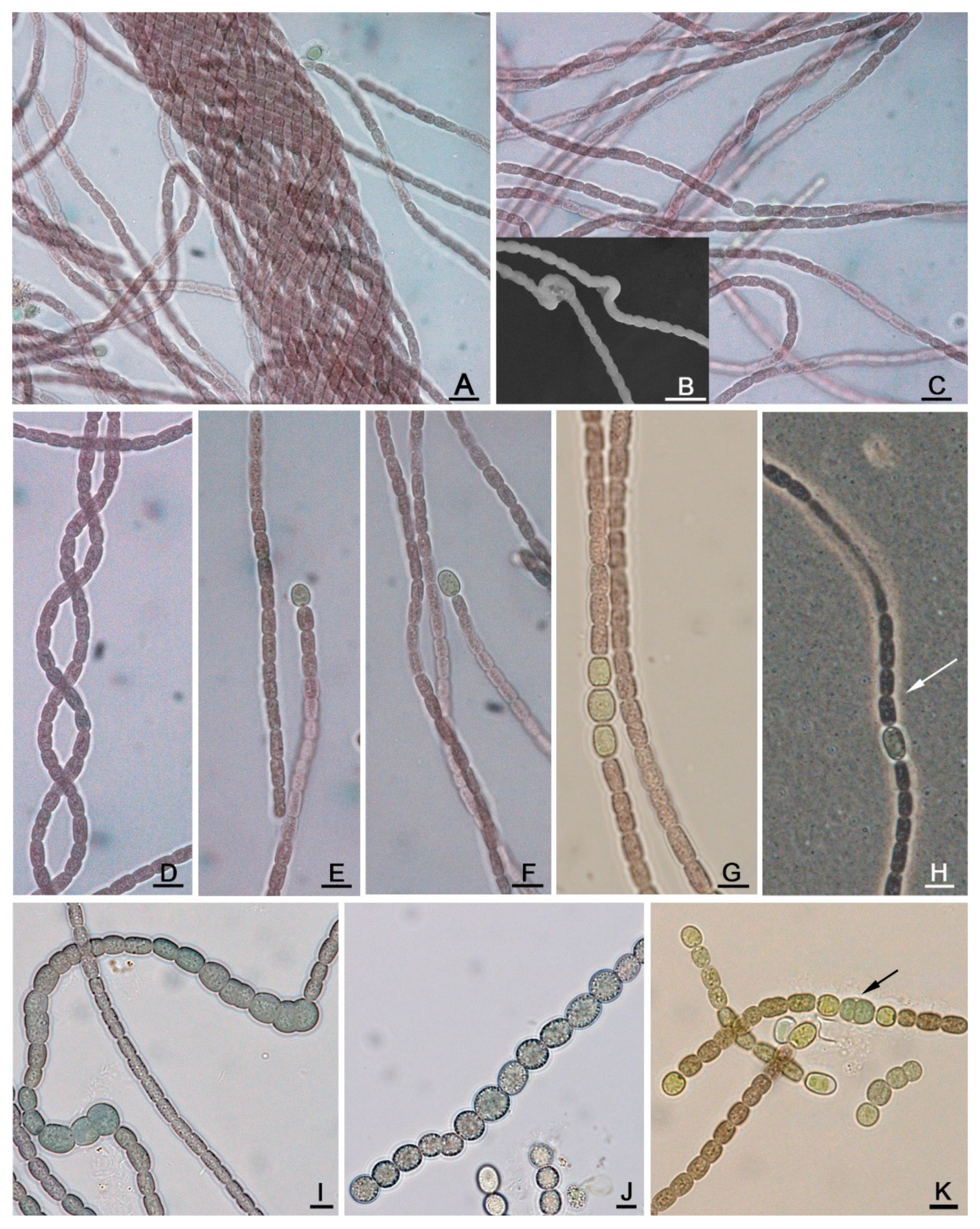
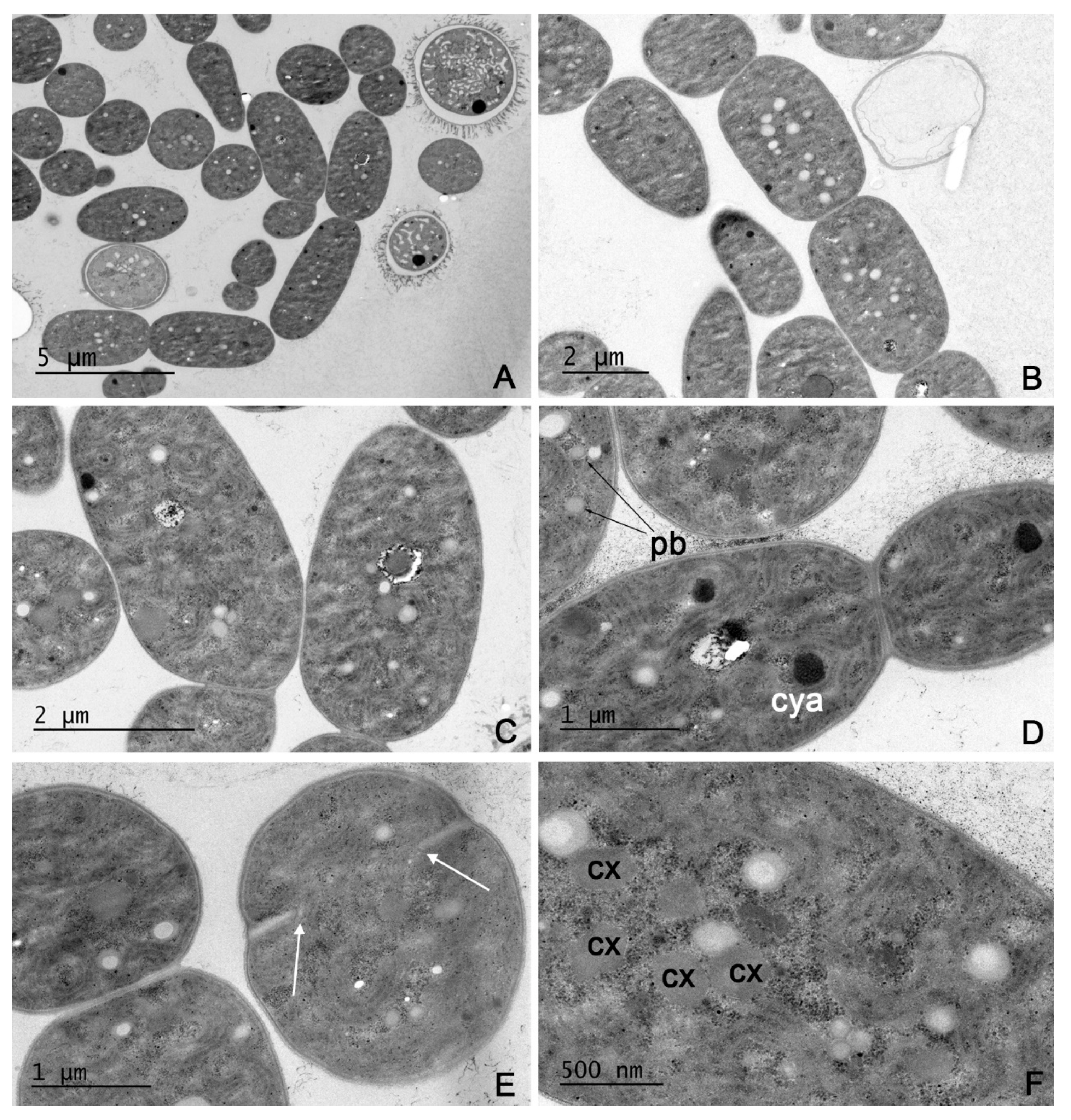
2.2. Microscopy
2.3. Molecular Characterization
2.3.1. DNA Extraction, PCR Amplification and Sequencing
2.3.2. Phylogenetic Analysis of 16S rRNA
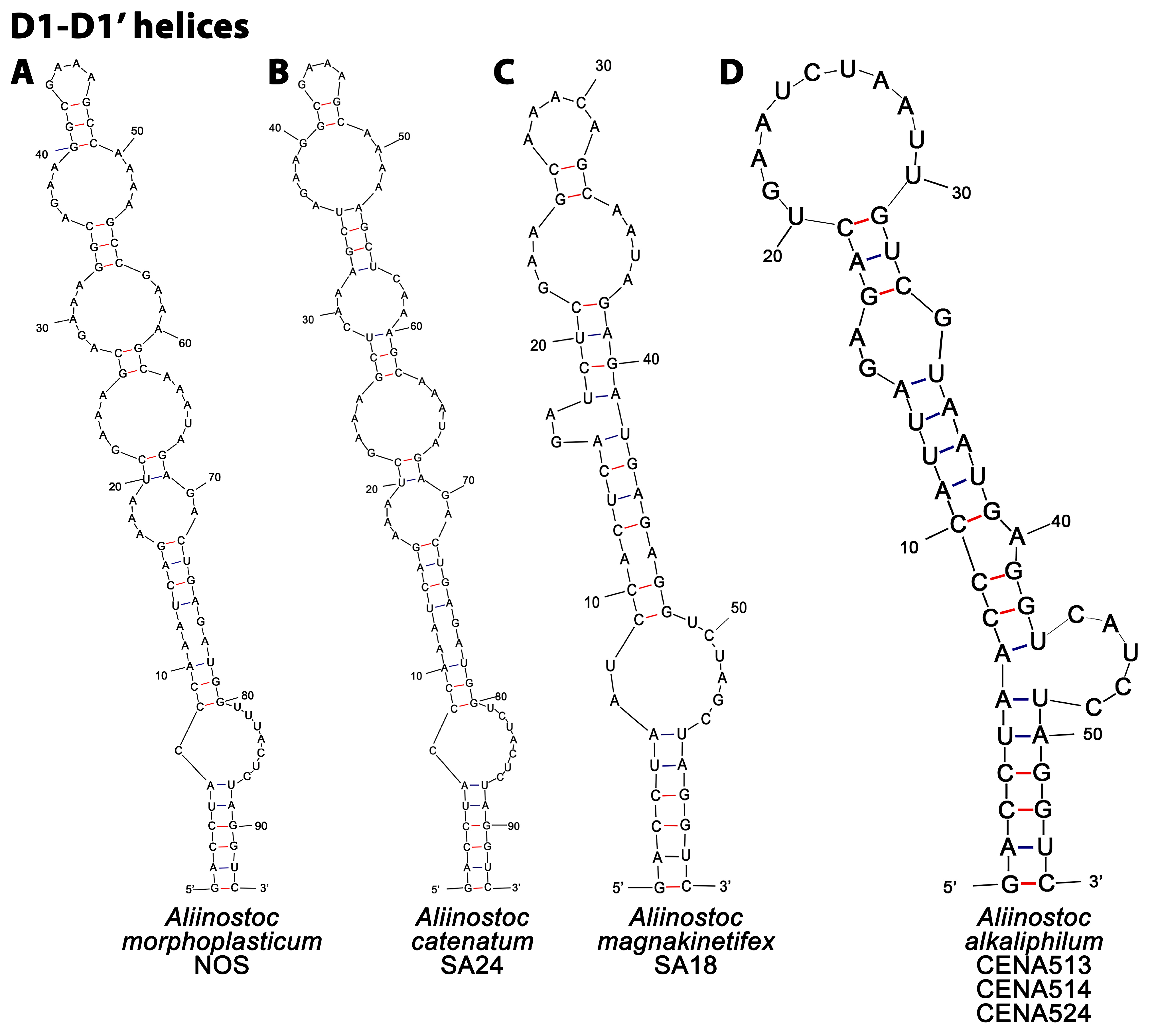
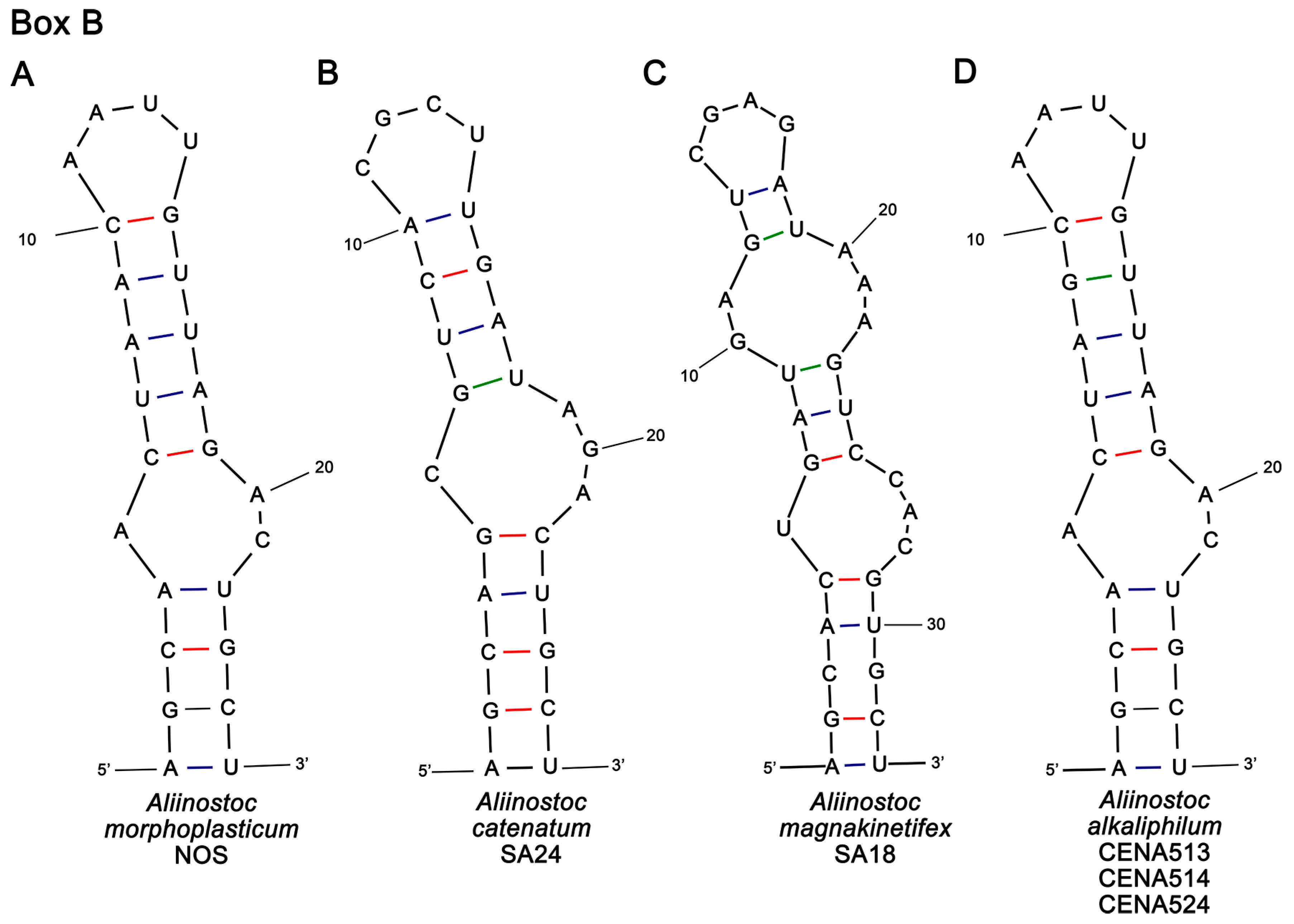
2.3.3. Determination of 16S-23S rRNA ITS Secondary Structures
2.4. Bioassays
2.4.1. Preparation of Extracts
2.4.2. Antimicrobial Disk Diffusion Susceptibility Testing
2.5. Liquid Chromatography
2.5.1. Fractionation of Extracts by High-Performance Liquid Chromatography (HPLC)
2.5.2. Chemical Analysis and Identification of Bioactive Metabolites
2.5.3. Identification of Bioactive Metabolite-Producing ‘Nostoc’ Strains
3. Results
3.1. Taxonomic Analysis
Description of Aliinostoc alkaliphilum sp. nov. under the Provisions of the International Code of Nomenclature for Algae, Fungi and Plants
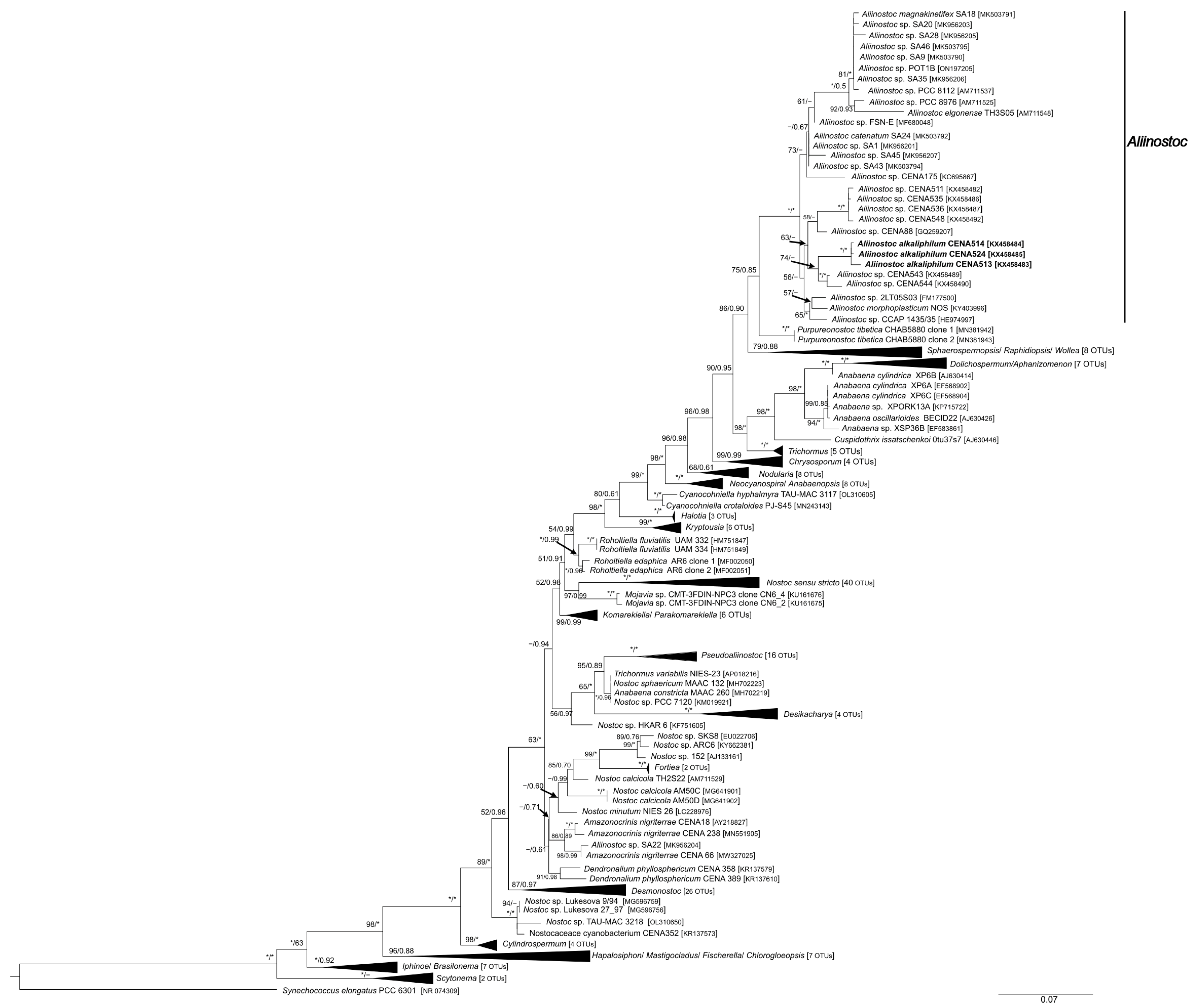
3.2. Phylogenetic Analyses and 16S-23S ITS Secondary Structures of Aliinostoc
3.3. Antimicrobial Susceptibility Testing and Chemical Analysis
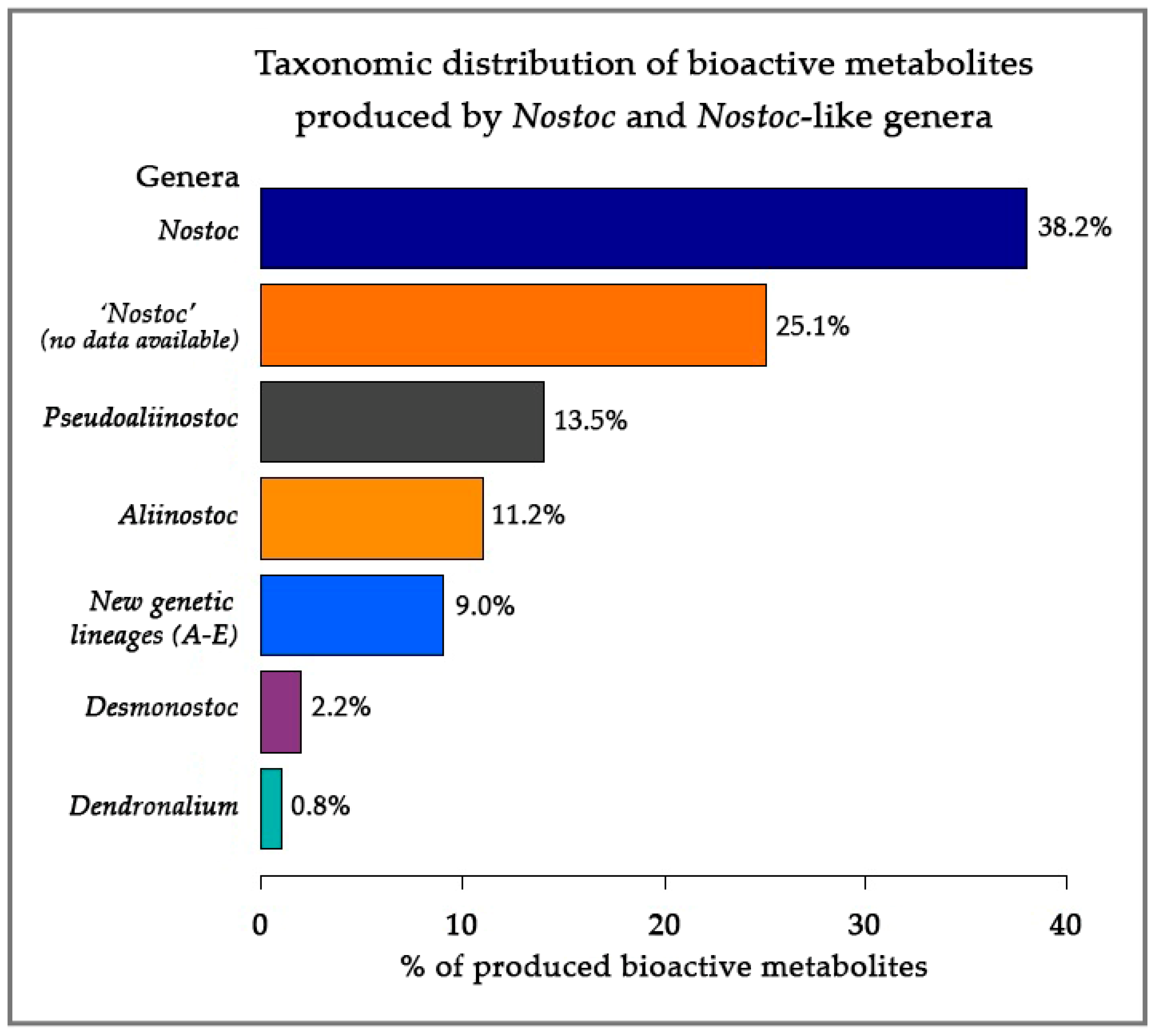
3.4. Taxonomic Distribution of Bioactive Metabolite-Producing Nostoc-like Strains
4. Discussion
4.1. Molecular Evaluation
4.2. Morphology and Ecology of Aliinostoc
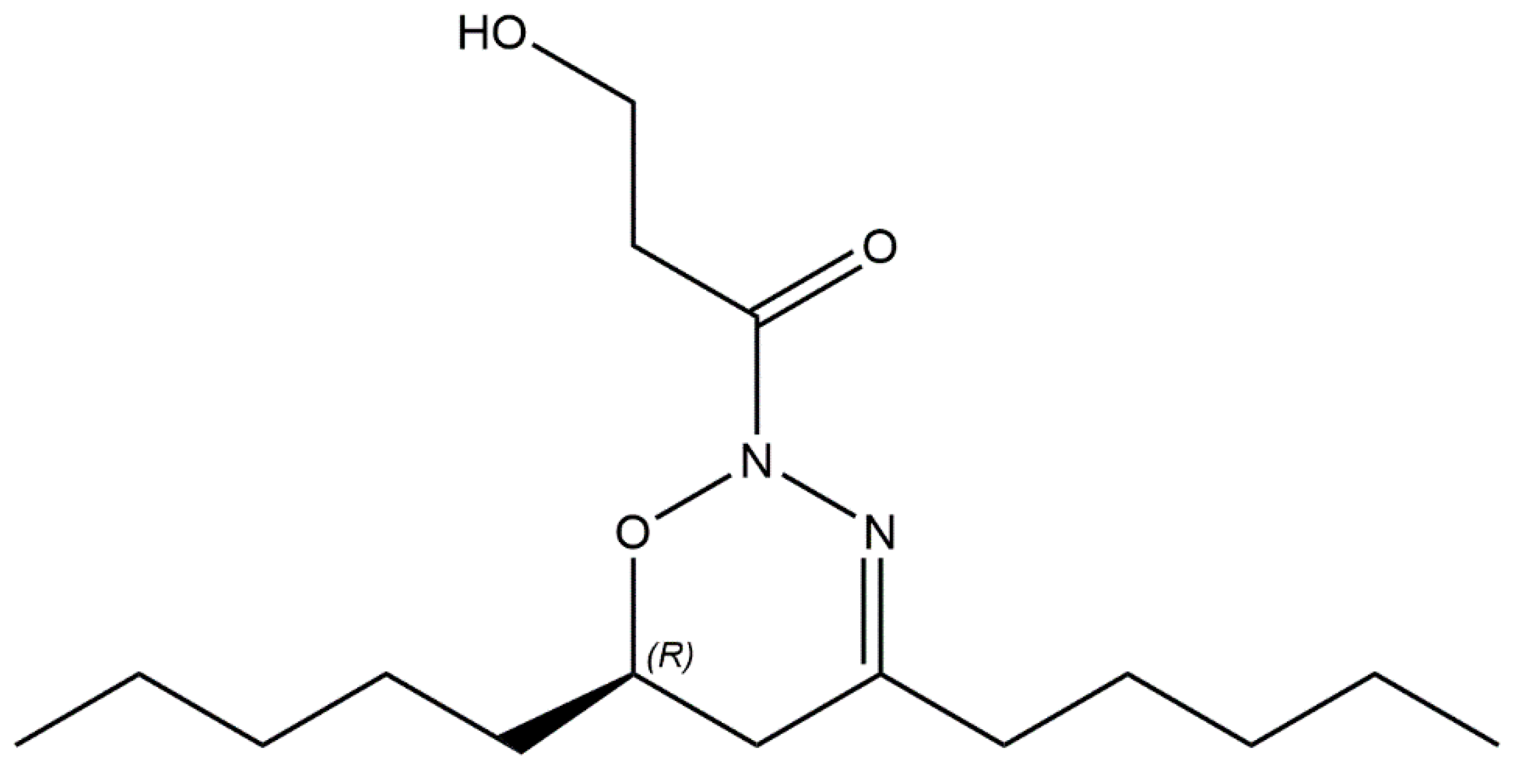
4.3. Bioactive Metabolites Produced by Aliinostoc alkaliphilum
4.4. Taxonomic Distribution of Bioactive Metabolites
4.4.1. Is genus Nostoc Rich in Bioactive Compounds?
4.4.2. Bioactive Metabolites from Nostoc-like Genera
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Whitton, B.A.; Potts, M. Introduction to the Cyanobacteria. In Ecology of Cyanobacteria II; Whitton, B.A., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 1–13. [Google Scholar]
- Swain, S.S.; Paidesetty, S.K.; Padhy, R.N. Antibacterial, antifungal and antimycobacterial compounds from cyanobacteria. Biomed. Pharmacother. 2017, 90, 760–776. [Google Scholar] [CrossRef] [PubMed]
- Mazard, S.; Penesyan, A.; Ostrowski, M.; Paulsen, I.T.; Egan, S. Tiny microbes with a big impact: The role of cyanobacteria and their metabolites in shaping our future. Mar. Drugs 2016, 14, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.K.; Tiwari, S.P.; Rai, A.K.; Mohapatra, T.M. Cyanobacteria: An Emerging source for drug discovery. J. Antibiot. 2011, 64, 401–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, M.R.; Pinto, E.; Torres, M.A.; Dörr, F.; Mazur-Marzec, H.; Szubert, K.; Tartaglione, L.; Dell’Aversano, C.; Miles, C.O.; Beach, D.G.; et al. CyanoMetDB, a comprehensive public database of secondary metabolites from cyanobacteria. Water Res. 2021, 196, 1–12. [Google Scholar] [CrossRef]
- Leão, P.N.; Engene, N.; Antunes, A.; Gerwick, W.H.; Vasconcelos, V. The chemical ecology of cyanobacteria. Nat. Prod. Rep. 2012, 29, 372–391. [Google Scholar] [CrossRef] [Green Version]
- Engene, N.; Gunasekera, S.P.; Gerwick, W.H.; Paul, V.J. Phylogenetic inferences reveal a large extent of novel biodiversity in chemically rich tropical marine cyanobacteria. Appl. Environ. Microbiol. 2013, 79, 1882–1888. [Google Scholar] [CrossRef] [Green Version]
- Engene, N.; Rottacker, E.C.; Kaštovský, J.; Byrum, T.; Choi, H.; Ellisman, M.H.; Komárek, J.; Gerwick, W.H. Moorea producens gen. nov., sp. nov. and Moorea bouillonii comb. nov., tropical marine cyanobacteria rich in bioactive secondary metabolites. Int. J. Syst. Evol. Microbiol. 2012, 62, 1171–1178. [Google Scholar] [CrossRef]
- Engene, N.; Choi, H.; Esquenazi, E.; Rottacker, E.C.; Ellisman, M.H.; Dorrestein, P.C.; Gerwick, W.H. Underestimated biodiversity as a major explanation for the perceived rich secondary metabolite capacity of the cyanobacterial genus Lyngbya. Environ. Microbiol. 2011, 13, 1601–1610. [Google Scholar] [CrossRef] [Green Version]
- Engene, N.; Tronholm, A.; Paul, V.J. Uncovering cryptic diversity of Lyngbya: The new tropical marine cyanobacterial genus Dapis (Oscillatoriales). J. Phycol. 2018, 54, 435–446. [Google Scholar] [CrossRef]
- Engene, N.; Paul, V.J.; Byrum, T.; Gerwick, W.H.; Thor, A.; Ellisman, M.H. Five chemically rich species of tropical marine cyanobacteria of the genus Okeania gen. nov. (Oscillatoriales, Cyanoprokaryota). J. Phycol. 2013, 49, 1095–1106. [Google Scholar] [CrossRef]
- Engene, N.; Tronholm, A.; Salvador-Reyes, L.A.; Luesch, H.; Paul, V.J. Caldora penicillata gen. nov., comb. nov. (Cyanobacteria), a pantropical marine species with biomedical relevance. J. Phycol. 2015, 51, 670–681. [Google Scholar] [CrossRef] [Green Version]
- Komárek, J.; Kaštovský, J.; Mareš, J.; Johansen, J.R. Taxonomic classification of Cyanoprokaryotes (cyanobacterial genera) 2014, using a polyphasic approach. Preslia 2014, 86, 295–335. [Google Scholar]
- Hoffmann, L.; Komárek, J.; Kaštovský, J. System of Cyanoprokaryotes (Cyanobacteria)–State in 2004. Arch. Hydrobiol. Suppl. Algol. Stud. 2005, 117, 95–115. [Google Scholar] [CrossRef]
- Johansen, J.R.; Casamatta, D.A. Recognizing cyanobacterial diversity through adoption of a new species paradigm. Arch. Hydrobiol. Suppl. Algol. Stud. 2005, 117, 71–93. [Google Scholar] [CrossRef]
- Řeháková, K.; Johansen, J.R.; Casamatta, D.A.; Xuesong, L.; Vincent, J. Morphological and molecular characterization of selected desert soil cyanobacteria: Three species new to science including Mojavia pulchra gen. et sp. nov. Phycologia 2007, 46, 481–502. [Google Scholar] [CrossRef]
- Saraf, A.G.; Dawda, H.G.; Singh, P. Desikacharya gen. nov., a phylogenetically distinct genus of cyanobacteria along with the description of two new species, Desikacharya nostocoides sp. nov. and Desikacharya soli sp. nov., and reclassification of Nostoc thermotolerans to Desikacharya thermotolerans comb. nov. Int. J. Syst. Evol. Microbiol. 2019, 69, 307–315. [Google Scholar]
- Hrouzek, P.; Lukešová, A.; Mareš, J.; Ventura, S. Description of the cyanobacterial genus Desmonostoc gen. nov. including D. muscorum comb. nov. as a distinct, phylogenetically coherent taxon related to the genus Nostoc. Fottea 2013, 13, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Genuário, D.; Vaz, M.; Hentschke, G.; Sant’Anna, C.; Fiore, M. Halotia gen. nov., a phylogenetically and physiologically coherent cyanobacterial genus isolated from marine coastal environments. Int. J. Syst. Evol. Microbiol. 2015, 65, 633–675. [Google Scholar] [CrossRef] [Green Version]
- Hentschke, G.S.; Johansen, J.R.; Pietrasiak, N.; Rigonato, J.; Fiore, M.F.; Sant’Anna, C.L. Komarekiella atlantica gen. et sp. nov. (Nostocaceae, Cyanobacteria): A new subaerial taxon from the atlantic rainforest and Kauai, Hawaii. Fottea 2017, 17, 178–190. [Google Scholar] [CrossRef] [Green Version]
- Bagchi, S.N.; Dubey, N.; Singh, P. Phylogenetically distant clade of nostoc-like taxa with the description of Aliinostoc gen. nov. and Aliinostoc morphoplasticum sp. nov. Int. J. Syst. Evol. Microbiol. 2017, 67, 3329–3338. [Google Scholar] [CrossRef]
- Cai, F.; Li, X.; Yang, Y.; Jia, N.; Huo, D.; Li, R. Compactonostoc shennongjiaensis gen. & sp. nov. (Nostocales, Cyanobacteria) from a wet rocky wall in china. Phycologia 2019, 58, 200–210. [Google Scholar]
- Soares, F.; Ramos, V.; Trovão, J.; Cardoso, S.M.; Tiago, I.; Portugal, A. Parakomarekiella sesnandensis gen. et sp. nov. (Nostocales, Cyanobacteria) Isolated from the Old Cathedral of Coimbra, Portugal (UNESCO World Heritage Site). Eur. J. Phycol. 2021, 56, 301–315. [Google Scholar] [CrossRef]
- Lee, N.J.; Bang, S.-D.; Kim, T.; Ki, J.S.; Lee, O.M. Pseudoaliinostoc sejongens gen. & sp. nov. (Nostocales, Cyanobacteria) from floodplain soil of the geum river in korea based on polyphasic approach. Phytotaxa 2021, 479, 55–70. [Google Scholar]
- Sivonen, K. Cyanobacterial Toxins. In Encyclopedia of Microbiology; Schaechter, M., Ed.; Elsevier: Oxford, UK, 2009; pp. 290–307. [Google Scholar]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management, 2nd ed.; Chorus, I., Welker, M., Eds.; CRC Press, on Behalf of the World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Liu, L.; Jokela, J.; Wahlsten, M.; Nowruzi, B.; Permi, P.; Zhang, Y.Z.; Xhaard, H.; Fewer, D.P.; Sivonen, K. Nostosins, trypsin inhibitors isolated from the terrestrial cyanobacterium Nostoc sp. strain FSN. J. Nat. Prod. 2014, 77, 1784–1790. [Google Scholar] [CrossRef]
- Humisto, A.; Herfindal, L.; Jokela, J.; Karkman, A.; Bjørnstad, R.; Choudhury, R.R.; Sivonen, K. Cyanobacteria as a source for novel anti-leukemic compounds. Curr. Pharm. Biotechnol. 2016, 17, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Dittmann, E.; Gugger, M.; Sivonen, K.; Fewer, D.P. Natural product biosynthetic diversity and comparative genomics of the cyanobacteria. Trends Microbiol. 2015, 23, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Nowruzi, B.; Haghighat, S.; Fahimi, H.; Mohammadi, E. Nostoc cyanobacteria species: A new and rich source of novel bioactive compounds with pharmaceutical potential. J. Pharm. Health Serv. Res. 2018, 9, 5–12. [Google Scholar] [CrossRef]
- Thuan, N.H.; An, T.T.; Shrestha, A.; Canh, N.X.; Sohng, J.K.; Dhakal, D. Recent advances in exploration and biotechnological production of bioactive compounds in three cyanobacterial genera: Nostoc, Lyngbya, and Microcystis. Front. Chem. 2019, 7, 1–33. [Google Scholar] [CrossRef]
- Grant, W.D. Alkaline systems and biodiversity. In Extremophiles; Gerday, C., Glansdorff, N., Eds.; Eolss Publishers: Oxford, UK, 2006; pp. 21–38. [Google Scholar]
- Sorokin, D.Y.; Berben, T.; Melton, E.D.; Overmars, L.; Vavourakis, C.D.; Muyzer, G. Microbial diversity and biogeochemical cycling in soda lakes. Extremophiles 2014, 18, 791–809. [Google Scholar] [CrossRef] [Green Version]
- Andreote, A.P.D.; Dini-Andreote, F.; Rigonato, J.; Machineski, G.S.; Souza, B.C.E.; Barbiero, L.; Rezende-Filho, A.T.; Fiore, M.F. Contrasting the genetic patterns of microbial communities in soda lakes with and without cyanobacterial bloom. Front. Microbiol. 2018, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Genuário, D.B.; Vaz, M.G.M.V.; Santos, S.N.; Kavamura, V.N.; Melo, I.S. Cyanobacteria from Brazilian Extreme environments: Toward functional exploitation. In Microbial Diversity in the Genomic Era; Das, S., Dash, H.R., Eds.; Academic Press: London, UK, 2019; pp. 265–284. [Google Scholar]
- Andreote, A.P.D.; Vaz, M.G.M.V.; Genuário, D.B.; Barbiero, L.; Rezende-Filho, A.T.; Fiore, M.F. Nonheterocytous cyanobacteria from Brazilian saline-alkaline lakes. J. Phycol. 2014, 50, 675–684. [Google Scholar] [CrossRef]
- Genuário, D.B.; Andreote, A.P.D.; Vaz, M.G.M.V.; Fiore, M.F. Heterocyte-forming cyanobacteria from Brazilian saline-alkaline lakes. Mol. Phylogenet. Evol. 2017, 109, 105–112. [Google Scholar] [CrossRef]
- Kabirnataj, S.; Nematzadeh, G.A.; Talebi, A.F.; Saraf, A.; Suradkar, A.; Tabatabaei, M.; Singh, P. Description of novel species of Aliinostoc, Desikacharya and Desmonostoc using a polyphasic approach. Int. J. Syst. Evol. Microbiol. 2020, 70, 3413–3426. [Google Scholar] [CrossRef]
- Kotai, J. Instructions for preparation of modified nutrient solution Z8 for algae. Nor. Inst. Water Res. 1972, 11, 1–5. [Google Scholar]
- Rippka, R. Isolation and purification of cyanobacteria. In Methods in Enzymology 167; Academic Press: San Diego, CA, USA, 1988; pp. 3–27. [Google Scholar]
- Leber, A.L. Antifungal disk diffusion susceptibility testing. In Clinical Microbiology Procedures Handbook; Leber, A.L., Ed.; ASM Press: Washington, DC, USA, 2016; Volume 2. [Google Scholar]
- Romo, S.; Perez-Martinez, C. The use of immobilization in alginate beads for long-term storage of Pseudanabaena galeata (Cyanobacteria) in the Laboratory. J. Phycol. 1997, 33, 1073–1076. [Google Scholar] [CrossRef]
- Reynolds, E.S. The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J. Cell Biol. 1963, 17, 208–212. [Google Scholar] [CrossRef] [Green Version]
- Boyer, S.L.; Johansen, J.R.; Flechtner, V.R.; Howard, G.L.; Bliss, F. Phylogeny and genetic variance in terrestrial Microcoleus (Cyanophyceae) species based on sequence analysis of the 16S rRNA gene and associated 16S–23S ITS region. J. Phycol. 2002, 38, 1222–1235. [Google Scholar] [CrossRef]
- Taton, A.; Grubisic, S.; Brambilla, E.; De Wit, R.; Wilmotte, A. Cyanobacterial diversity in natural and artificial microbial mats of lake Fryxell (McMurdo Dry Valleys, Antarctica): A morphological and molecular approach. Appl. Environ. Microbiol. 2003, 69, 5157–5169. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Automat. Contr. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, 232–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Kimura, M. A Simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Iteman, I.; Rippka, R.; De Marsac, N.T.; Herdman, M. Comparison of Conserved structural and regulatory domains within divergent 16S rRNA-23S rRNA spacer sequences of cyanobacteria. Microbiology 2000, 146, 1275–1286. [Google Scholar] [CrossRef] [Green Version]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Disk Susceptibility Tests; Approved Standard—Eleventh Edition. CLSI Document M02-A11, 11th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- Voráčová, K.; Hájek, J.; Mareš, J.; Urajová, P.; Kuzma, M.; Cheel, J.; Villunger, A.; Kapuscik, A.; Bally, M.; Novák, P.; et al. The cyanobacterial metabolite nocuolin A is a natural oxadiazine that triggers apoptosis in human cancer cells. PLoS ONE 2017, 12, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Saraf, A.; Dawda, H.G.; Suradkar, A.; Behere, I.; Kotulkar, M.; Shaikh, Z.M.; Kumat, A.; Batule, P.; Mishra, D.; Singh, P. Description of two new species of Aliinostoc and one new species of Desmonostoc from india based on the polyphasic approach and reclassification of Nostoc punensis to Desmonostoc punense comb. nov. FEMS Microbiol. Lett. 2018, 365, 1–10. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Goebel, B.M. Taxonomic Note: A place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. Int. J. Syst. Bacteriol. 1994, 44, 846–849. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Oh, H.S.; Park, S.C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef]
- Wilmotte, A.; Neefs, J.M.; De Wachter, R. Evolutionary affiliation of the marine nitrogen-fixing cyanobacterium Trichodesmium sp. strain NIBB 1067, derived by 16S ribosomal RNA sequence analysis. Microbiology 1994, 140, 2159–2164. [Google Scholar] [CrossRef] [Green Version]
- Boyer, S.L.; Flechtner, V.R.; Johansen, J.R. Is the 16S-23S rRNA internal transcribed spacer region a good tool for use in molecular systematics and population genetics? A case study in cyanobacteria. Mol. Biol. Evol. 2001, 18, 1057–1069. [Google Scholar] [CrossRef] [Green Version]
- Johansen, J.R.; Kovacik, L.; Casamatta, D.A.; Fučiková, K.; Kaštovský, J. Utility of 16S-23S ITS sequence and secondary structure for recognition of intrageneric and intergeneric limits within cyanobacterial taxa: Leptolyngbya corticola sp. nov. (Pseudanabaenaceae, Cyanobacteria). Nov. Hedwigia 2011, 92, 283–302. [Google Scholar] [CrossRef]
- Berrendero Gómez, E.; Johansen, J.R.; Kaštovský, J.; Bohunická, M.; Čapková, K. Macrochaete gen. nov. (Nostocales, Cyanobacteria), a taxon morphologically and molecularly distinct from Calothrix. J. Phycol. 2016, 52, 638–655. [Google Scholar] [CrossRef]
- Alvarenga, D.O.; Andreote, A.P.D.; Branco, L.H.Z.; Fiore, M.F. Kryptousia macronema gen. nov., sp. nov. and Kryptousia microlepis sp. nov., nostocalean cyanobacteria isolated from phyllospheres. Int. J. Syst. Evol. Microbiol. 2017, 67, 3301–3309. [Google Scholar] [CrossRef] [Green Version]
- Martins, M.D.; Rigonato, J.; Taboga, S.R.; Branco, L.H.Z. Proposal of Ancylothrix gen. nov., a new genus of Phormidiaceae (Cyanobacteria, Oscillatoriales) based on a polyphasic approach. Int. J. Syst. Evol. Microbiol. 2016, 66, 2396–2405. [Google Scholar] [CrossRef]
- Martins, M.D.; Branco, L.H.Z. Potamolinea gen. nov. (Oscillatoriales, Cyanobacteria): A phylogenetically and ecologically coherent cyanobacterial genus. Int. J. Syst. Evol. Microbiol. 2016, 66, 3632–3641. [Google Scholar] [CrossRef]
- Pietrasiak, N.; Reeve, S.; Osorio-Santos, K.; Lipson, D.A.; Johansen, J.R. Trichotorquatus gen. nov.—A new genus of soil cyanobacteria discovered from American drylands 1. J. Phycol. 2021, 57, 886–902. [Google Scholar] [CrossRef]
- Machado-de-Lima, N.M.; Dornelles Martins, M.; Branco, L.H. Description of a tropical new species of Wilmottia (Oscillatoriales, Cyanobacteria) and considerations about the monophyly of W. Murrayi. Phytotaxa 2017, 307, 43–54. [Google Scholar] [CrossRef]
- Johansen, J.R.; Mareš, J.; Pietrasiak, N.; Bohunická, M.; Zima, J.; Štenclová, L.; Hauer, T. Highly divergent 16S rRNA sequences in ribosomal operons of Scytonema hyalinum (Cyanobacteria). PLoS ONE 2017, 12, e0186393. [Google Scholar] [CrossRef] [PubMed]
- Buch, B.; Martins, M.D.; Branco, L.H.Z. A widespread cyanobacterium supported by polyphasic approach: Proposition of Koinonema pervagatum gen. & sp. nov. (Oscillatoriales). J. Phycol. 2017, 53, 1097–1105. [Google Scholar] [PubMed]
- González-Fuente, A.M.; Hernández-Cabanillas, A.; Maderuelo-Corral, S.; Ortega-Doménech, M.; Rosero-Valencia, D.F.; Rumbero-Sánchez, Á.; Tena-Pérez, V. Dihydrooxadiazine Compounds for Treating Infections and Cancer. European Patent EP3156400B1, 15 May 2019. [Google Scholar]
- Arunkumar, K.; Selvapalam, N.; Rengasamy, R. The antibacterial compound sulphoglycerolipid 1-0 palmitoyl-3-0(6′-sulpho-α-quinovopyranosyl)-glycerol from Sargassum wightii Greville (Phaeophyceae). Bot. Mar. 2005, 48, 441–445. [Google Scholar] [CrossRef] [Green Version]
- El Baz, F.K.; El Baroty, G.S.; Abd El Baky, H.H.; Abd El-Salam, O.I.; Ibrahim, E.A. Structural characterization and biological activity of sulfolipids from selected marine algae. Grasas Y Aceites 2013, 64, 561–571. [Google Scholar]
- Logan, N.A.; De Vos, P. Bacillus. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.B., Rainey, F., Kämpfer, P., Trujillo, M., Chun, J., DeVos, P., Hedlund, B., Dedysh, S., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2015; pp. 1–163. [Google Scholar]
- Tidgewell, K.; Clark, B.R.; Gerwick, W.H. The natural products chemistry of cyanobacteria. In Comprehensive Natural Products II: Chemistry and Biology-Volume 2; Mander, L., Liu, H.-W., Eds.; Elsevier: Oxford, UK, 2010; pp. 141–188. [Google Scholar]
- Tronholm, A.; Engene, N. Moorena gen. nov., a valid name for “Moorea Engene et al” nom. inval. (Oscillatoriaceae, Cyanobacteria). Not. Algarum 2019, 122, 1–2. [Google Scholar]
- Trimurtulu, G.; Ohtani, I.; Patterson, G.M.L.; Moore, R.E.; Corbett, T.H.; Valeriote, F.A.; Demchik, L. Total Structures of cryptophycins, potent antitumor depsipeptides from the blue-green alga Nostoc sp. strain GSV 224. J. Am. Chem. Soc. 1994, 116, 4729–4737. [Google Scholar] [CrossRef]
- Magarvey, N.A.; Beck, Z.Q.; Golakoti, T.; Ding, Y.; Huber, U.; Hemscheidt, T.K.; Abelson, D.; Moore, R.E.; Sherman, D.H. Biosynthetic characterization and chemoenzymatic assembly of the cryptophycins. potent anticancer agents from cyanobionts. ACS Chem. Biol. 2006, 1, 766–779. [Google Scholar] [CrossRef]
- Kang, H.S.; Santarsiero, B.D.; Kim, H.; Krunic, A.; Shen, Q.; Swanson, S.M.; Chai, H.; Kinghorn, A.D.; Orjala, J. Merocyclophanes A and B, antiproliferative cyclophanes from the cultured terrestrial cyanobacterium Nostoc sp. Phytochemistry 2012, 79, 109–115. [Google Scholar] [CrossRef] [Green Version]
- May, D.S.; Chen, W.L.; Lantvit, D.D.; Zhang, X.; Krunic, A.; Burdette, J.E.; Eustaquio, A.; Orjala, J. Merocyclophanes C and D from the cultured freshwater cyanobacterium Nostoc sp. (UIC 10110). J. Nat. Prod. 2017, 80, 1073–1080. [Google Scholar] [CrossRef] [Green Version]
- Kampa, A.; Gagunashvili, A.N.; Gulder, T.A.M.; Morinaka, B.I.; Daolio, C.; Godejohann, M.; Miao, V.P.W.; Piel, J.; Andrésson, Ó.S. Metagenomic natural product discovery in lichen provides evidence for a family of biosynthetic pathways in diverse symbioses. Proc. Natl. Acad. Sci. USA 2013, 110, 3129–3137. [Google Scholar] [CrossRef] [Green Version]
- Shishido, T.K.; Popin, R.V.; Jokela, J.; Wahlsten, M.; Fiore, M.F.; Fewer, D.P.; Herfindal, L.; Sivonen, K. Dereplication of natural products with antimicrobial and anticancer activity from Brazilian cyanobacteria. Toxins 2019, 12, 12. [Google Scholar] [CrossRef] [Green Version]
- Genuário, D.B.; Silva-Stenico, M.E.; Welker, M.; Beraldo Moraes, L.A.; Fiore, M.F. Characterization of a microcystin and detection of microcystin synthetase genes from a Brazilian isolate of Nostoc. Toxicon 2010, 55, 846–854. [Google Scholar] [CrossRef]
- Armstrong, L.; Vaz, M.G.M.V.; Genuário, D.B.; Fiore, M.F.; Debonsi, H.M. Volatile compounds produced by cyanobacteria isolated from mangrove environment. Curr. Microbiol. 2019, 76, 575–582. [Google Scholar] [CrossRef]
- Jokela, J.; Heinilä, L.M.P.; Shishido, T.K.; Wahlsten, M.; Fewer, D.P.; Fiore, M.F.; Wang, H.; Haapaniemi, E.; Permi, P.; Sivonen, K. Production of high amounts of hepatotoxin nodularin and new protease inhibitors pseudospumigins by the Brazilian benthic Nostoc sp. CENA543. Front. Microbiol. 2017, 8, 1963. [Google Scholar] [CrossRef]
- Shishido, T.K.; Jokela, J.; Fewer, D.P.; Wahlsten, M.; Fiore, M.F.; Sivonen, K. Simultaneous production of anabaenopeptins and namalides by the cyanobacterium Nostoc sp. CENA543. ACS Chem. Biol. 2017, 12, 2746–2755. [Google Scholar] [CrossRef]
- Bui, H.T.N.; Jansen, R.; Pham, H.T.L.; Mundt, S. Carbamidocyclophanes A-E, chlorinated paracyclophanes with cytotoxic and antibiotic activity from the vietnamese cyanobacterium Nostoc sp. J. Nat. Prod. 2007, 70, 499–503. [Google Scholar] [CrossRef]
- Chlipala, G.E.; Sturdy, M.; Krunic, A.; Lantvit, D.D.; Shen, Q.; Porter, K.; Swanson, S.M.; Orjala, J. Cylindrocyclophanes with proteasome inhibitory activity from the cyanobacterium Nostoc sp. J. Nat. Prod. 2010, 73, 1529–1537. [Google Scholar] [CrossRef] [Green Version]
- Preisitsch, M.; Harmrolfs, K.; Pham, H.T.; Heiden, S.E.; Füssel, A.; Wiesner, C.; Pretsch, A.; Swiatecka-Hagenbruch, M.; Niedermeyer, T.H.; Müller, R.; et al. Anti-MRSA-acting carbamidocyclophanes H–L from the vietnamese cyanobacterium Nostoc sp. CAVN2. J. Antibiot. 2015, 68, 165–177. [Google Scholar] [CrossRef]
- May, D.S.; Kang, H.-S.; Santarsiero, B.D.; Krunic, A.; Shen, Q.; Burdette, J.E.; Swanson, S.M.; Orjala, J. Ribocyclophanes A–E, glycosylated cyclophanes with antiproliferative activity from two cultured terrestrial cyanobacteria. J. Nat. Prod. 2018, 81, 572–578. [Google Scholar] [CrossRef]
- Preisitsch, M.; Niedermeyer, T.H.J.; Heiden, S.E.; Neidhardt, I.; Kumpfmüller, J.; Wurster, M.; Harmrolfs, K.; Wiesner, C.; Enke, H.; Müller, R.; et al. Cylindrofridins A–C, linear cylindrocyclophane-related alkylresorcinols from the cyanobacterium Cylindrospermum stagnale. J. Nat. Prod. 2016, 79, 106–115. [Google Scholar] [CrossRef]
- Preisitsch, M.; Heiden, S.E.; Beerbaum, M.; Niedermeyer, T.H.J.; Schneefeld, M.; Herrmann, J.; Kumpfmüller, J.; Thürmer, A.; Neidhardt, I.; Wiesner, C.; et al. Effects of halide ions on the carbamidocyclophane biosynthesis in Nostoc sp. CAVN2. Mar. Drugs 2016, 14, 21. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Kang, H.-S.; Krunic, A.; Chlipala, G.E.; Cai, G.; Chen, W.-L.; Franzblau, S.G.; Swanson, S.M.; Orjala, J. Carbamidocyclophanes F and G with anti-Mycobacterium tuberculosis activity from the cultured freshwater cyanobacterium Nostoc sp. Tetrahedron Lett. 2014, 55, 686–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, B.S.; Chen, J.L.; Patterson, G.M.L.; Moore, R.E.; Brinen, L.S.; Kato, Y.; Clardy, J. [7.7]Paracyclophanes from blue-green algae. J. Am. Chem. Soc. 1990, 112, 4061–4063. [Google Scholar] [CrossRef]
- Moore, B.S.; Chen, J.-L.; Patterson, G.M.L.; Moore, R.E. Structures of cylindrocyclophanes A-F. Tetrahedron 1992, 48, 3001–3006. [Google Scholar] [CrossRef]
- Chen, J.L.; Moore, R.E.; Patterson, G.M.L. Structures of nostocyclophanes A-D. J. Org. Chem. 1991, 56, 4360–4364. [Google Scholar] [CrossRef]
- Bobzin, S.C.; Moore, R.E. Biosynthetic origin of [7.7]Paracyclophanes from Cyanobacteria. Tetrahedron 1993, 49, 7615–7626. [Google Scholar] [CrossRef]
- Mattila, A.; Andsten, R.-M.; Jumppanen, M.; Assante, M.; Jokela, J.; Wahlsten, M.; Mikula, K.M.; Sigindere, C.; Kwak, D.H.; Gugger, M.; et al. Biosynthesis of the bis-prenylated alkaloids muscoride A and B. ACS Chem. Biol. 2019, 14, 2683–2690. [Google Scholar] [CrossRef]
- Nagatsu, A.; Kajitani, H.; Sakakibara, J. Muscoride A: A new oxazole peptide alkaloid from freshwater cyanobacterium Nostoc muscorum. Tetrahedron Lett. 1995, 36, 4097–4100. [Google Scholar] [CrossRef]
- Cheel, J.; Hájek, J.; Kuzma, M.; Saurav, K.; Smýkalová, I.; Ondráčková, E.; Urajová, P.; Vu, D.L.; Faure, K.; Kopecký, J.; et al. Application of HPCCC combined with polymeric resins and HPLC for the separation of cyclic lipopeptides muscotoxins A-C and their antimicrobial activity. Molecules 2018, 23, 2653. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Name | D1-D1′ | Box B |
|---|---|---|
| Aliinostoc morphoplasticum NOS | 93 | 25 |
| Aliinostoc catenatum SA24 | 93 | 26 |
| Aliinostoc magnakinetifex SA18 | 60 | 33 |
| Aliinostoc alkaliphilum (all three strains) | 54 | 25 |
| Crude Extracts of A. alkaliphilum Strains | Controls | ||||
|---|---|---|---|---|---|
| Strain Name | CENA513T | CENA514 | CENA524 | Positive * | Negative ** |
| S. aureus | + | + | + | + | − |
| B. cereus | + | − | − | + | − |
| A. flavus | + | + | + | + | − |
| Mucor sp. | + | + | + | + | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Christodoulou, M.; Jokela, J.; Wahlsten, M.; Saari, L.; Economou-Amilli, A.; Fiore, M.d.F.; Sivonen, K. Description of Aliinostoc alkaliphilum sp. nov. (Nostocales, Cyanobacteria), a New Bioactive Metabolite-Producing Strain from Salina Verde (Pantanal, Brazil) and Taxonomic Distribution of Bioactive Metabolites in Nostoc and Nostoc-like Genera. Water 2022, 14, 2470. https://doi.org/10.3390/w14162470
Christodoulou M, Jokela J, Wahlsten M, Saari L, Economou-Amilli A, Fiore MdF, Sivonen K. Description of Aliinostoc alkaliphilum sp. nov. (Nostocales, Cyanobacteria), a New Bioactive Metabolite-Producing Strain from Salina Verde (Pantanal, Brazil) and Taxonomic Distribution of Bioactive Metabolites in Nostoc and Nostoc-like Genera. Water. 2022; 14(16):2470. https://doi.org/10.3390/w14162470
Chicago/Turabian StyleChristodoulou, Maria, Jouni Jokela, Matti Wahlsten, Lyudmila Saari, Athena Economou-Amilli, Marli de Fatima Fiore, and Kaarina Sivonen. 2022. "Description of Aliinostoc alkaliphilum sp. nov. (Nostocales, Cyanobacteria), a New Bioactive Metabolite-Producing Strain from Salina Verde (Pantanal, Brazil) and Taxonomic Distribution of Bioactive Metabolites in Nostoc and Nostoc-like Genera" Water 14, no. 16: 2470. https://doi.org/10.3390/w14162470