
Abstract
People respond more slowly in two-choice tasks when either a previous stimulus feature or the previous response repeats in partial repetition trials than when (a) both repeat in complete repetition trials or (b) both alternate in complete alternation trials. The binding account posits that such partial repetition costs index a memory-retrieval conflict, which occurs because partial repetition trials trigger the retrieval of a previous stimulus feature or response that conflicts with a current stimulus feature or response. However, such costs may additionally reflect a simple decision-making heuristic that uses the repetition or alternation of a previous stimulus feature as a “signal” to bias response selection toward a repetition or an alternation of the previous response. To determine whether signaling contributes to partial repetition costs, we employed a four-choice task. Here, a stimulus feature repetition still signals a response repetition, but a stimulus feature alternation does not signal which of the three remaining responses to make. Consistent with an influence of signaling, we sometimes observed complete repetition advantages without complete alternation advantages. Exploratory analyses further revealed that partial repetition costs measured more broadly were smaller in the four-choice task than in a matched two-choice task. These findings suggest that partial repetition costs index a mixture of binding and signaling.
Similar content being viewed by others

Introduction
The ability to act in a controlled manner is crucial for goal-directed behavior. Current views posit that the ability to exert control over action, or action control, depends on both (1) top-down, voluntary processes that map goal-relevant stimulus features to responses (Miller & Cohen, 2001) and (2) recent experiences with both relevant and irrelevant stimulus features and responses (Hommel, 1998). For example, a driver’s ability to press the brake when approaching a red stoplight may depend on both (1) knowledge of the correct stimulus-response (S-R) mapping (i.e., red means press the brake) and (2) recent experiences with red stoplights (e.g., braking as usual or accelerating if a police officer recently waved the driver through a red light).
Findings from the partial repetition cost (PRC) task provide a laboratory example of how recent experience influences action control (Hommel, 1998, 2007; Hommel & Colzato, 2004; Huffman et al., 2020). Each trial of this task consists of three successive events (Fig. 1). First, a Response Cue (e.g., a “1” or a “2”) at fixation instructs participants to prepare a left or a right keypress. Second, participants respond to a Cued Response Stimulus (e.g., a blue or green square that appears above or below fixation) by making the prepared keypress (e.g., left). Third, participants respond to a Discrimination Response Stimulus (e.g., a blue or green square that appears above or below fixation) by employing a pre-specified stimulus-response (S-R) mapping. This S-R mapping involves the task-relevant stimulus feature (e.g., color – make a left or right keypress if a blue or a green square appears, respectively) but not the task-irrelevant stimulus feature (e.g., location – top or bottom). Responses to the Discrimination Response Stimulus are slower and/or less accurate when (1) a stimulus feature (e.g., color) repeats and the response (e.g., left keypress) alternates, or vice versa, than when (2) a stimulus feature and the response both repeat or both alternate (Hommel, 1998, 2007). This data pattern manifests as a two-way interaction between (1) whether a stimulus feature repeats or alternates and (2) whether the response repeats or alternates. This interaction appears separately for the task-relevant and task-irrelevant stimulus features.

Example trial from the partial repetition cost (PRC) task. The response cue (e.g., “1”) appears before the Cued Response Stimulus (e.g., a green square) which, in turn, appears before the Discrimination Response Stimulus (e.g., a blue square)
The binding hypothesis
The binding hypothesis provides an influential explanation of this interaction (Hommel, 1998, 2004; Hommel et al., 2001). In this view, participants create a memory of the stimulus-response (S-R) integration episode corresponding to the Cued Response Stimulus. This episodic memory – or event file – typically stores the stimulus and response features that occurred as a collection of independent, binary bindings (e.g., between the color blue and a left keypress, the location “top” and a left keypress, etc.). Critically, repeating a feature (e.g., the color blue) from one of these bindings in the Discrimination Response Stimulus triggers the retrieval of the associated feature (e.g., left keypress) from the same binding.
What is the impact of such retrieval? If the stimulus feature and the response feature from a previous binding repeat (e.g., the color and the response repeat; complete repetition trials), participants respond quickly and accurately because the retrieved features match the current features. If no features repeat (e.g., the color and the response both change; complete alternation trials), no retrieval occurs but participants respond quickly and accurately because the process of creating a new binding is fast and efficient. Finally, if one feature repeats and the other alternates (e.g., the color repeats and the response changes or vice-versa; partial repetition trials), participants respond more slowly and make more errors. In these trials, a repeated feature (e.g., the color blue) triggers the retrieval of the associated feature (e.g., left keypress) from the same binding. However, the associated feature (e.g., left keypress) differs from the current feature (e.g., right keypress) with which the repeated feature (e.g., the color blue) now appears. This leads to a conflict, which may be resolved by “unbinding” the repeated feature (e.g., the color blue) from the associated feature (e.g., left keypress) and forming an updated binding between the repeated feature (e.g., the color blue) and the current feature (e.g., right keypress) (Hommel, 2004). Critically, this conflict impairs performance in partial repetition trials relative to complete repetition and complete alternation trials, resulting in “partial repetition costs” (Fig. 2).

Idealized partial repetition costs related to a color by response interaction in the two-choice partial repetition cost (PRC) task. Mean response time (RT) to the second colored square is relatively short when both color and response repeat in complete repetition trials (bottom left) or both change in complete alternation trials (bottom right). In contrast, mean RT to the second colored square is relatively long when (1) the color changes and the response repeats (top left) or (2) the color repeats and the response changes (top right) in partial repetition trials
The binding hypothesis further posits that a response (e.g., left keypress) can be stored in more than one binding (e.g., a binding with a color, a binding with a location, etc.) (Giesen & Rothermund, 2014). This allows repetitions of different features of the Cued Response Stimulus (e.g., its color, its location, etc.) to independently trigger the retrieval of the associated response. Consistent with this view, partial repetition costs that stem from repetitions and alternations of distinct stimulus features (e.g., colors and locations) typically sum additively (Hommel, 1998; Huffman et al., 2020).
Recent views of binding as an explanation of partial repetition costs
The explanation of partial repetition costs provided by the binding hypothesis has changed in three important ways since its inception. First, some recent views posit that partial repetition costs index the effects of episodic retrieval on performance (e.g., response conflict) in the absence of unbinding costs (Frings et al., 2020; Schmidt et al., 2016). These views still predict a two-way interaction between (1) whether a stimulus feature repeats or alternates and (2) whether the response repeats or alternates. However, the interaction stems solely from the costs of experiencing heightened conflict in partial repetition trials, rather than from additional costs associated with updating a previous binding (Hommel, 2004).
Second, some recent views posit that event files also store abstract, contextual features that are not immediately observable within a trial (e.g., task sets, control settings, stimulus-response mappings, etc.) (Dignath et al., 2019; Egner, 2014; Grant et al., 2020). Abstract features are bound and retrieved in the same way as concrete stimulus and response features. Thus, repeating an abstract feature (e.g., the task) triggers the retrieval of the previous trial’s event file even when no concrete features repeat.
Third, one recent view (Dutzi & Hommel, 2009) posits an additional influence on performance in complete alternation trials. In this view, a stimulus feature alternation biases response selection toward a response alternation via a process called “integrated competition” (Duncan, 1996; Duncan et al., 1997). It is unclear, though, whether integrated competition applies to binding phenomena. Bindings exist in memory (i.e., within event files), but the integrated competition hypothesis “deals only with the restricted domain of perceived visual objects” (Duncan, 1996, page 572). The restricted domain of this hypothesis stems from the fact that it is an extension of the biased competition model of attention (Desimone & Duncan, 1995), which explains how visual attention biases inhibitory interactions between co-occurring objects that are competing to drive a visual cortical neuron’s activity. Such interactions, however, do not occur when different objects appear sequentially rather than simultaneously (Beck & Kastner, 2005, 2007, 2009; Luck et al., 1997). Therefore, since a 500-ms interval separates the Cued Response Stimulus and the Discrimination Response Stimulus in the PRC task, it appears unlikely that integrated competition biases response selection toward a response alternation. Nonetheless, as we describe next, a different mechanism may allow a stimulus feature alternation to bias response selection toward a response alternation.
The signaling hypothesis
Some researchers argue that partial repetition costs in two-alternative forced-choice (two-choice) tasks – including the PRC task – partly reflect the influence of a simple decision-making heuristic (Açkay & Hazeltine, 2007; Fletcher & Rabbitt, 1978; Huffman et al., 2020; Kingstone, 1992; Krummenacher et al., 2009; Müller & Krummenacher, 2006; Notebaert & Soetens, 2003; Schoepper et al., 2020; Tollner et al., 2008). To our knowledge, Fletcher and Rabbitt (1978) were the first to make this suggestion. They tested the hypothesis that, after extended practice, participants code each stimulus as a repetition or an alternation of the previous stimulus, rather than in terms of its specific identity (e.g., the letter B or the letter Y). Specifically, they tested whether participants use such codes to implement the following strategy. If the stimulus repeats, prepare for a response repetition. If the stimulus alternates, prepare for a response alternation.
To test their hypothesis, Fletcher and Rabbitt (1978) inserted a dot pattern stimulus (8% of trials) into a two-choice task that maps two letters (e.g., B and Y) onto two responses (e.g., left and right). The authors instructed participants to choose freely between the two possible responses when the dot pattern appeared. They reasoned that if participants select a response based on whether the current stimulus is a repetition or an alternation of the previous stimulus, they should produce a response alternation when the infrequent dot pattern stimulus appears.
Consistent with this “signaling” hypothesis,Footnote 1 participants exhibited exactly such a bias after extended practice, and three additional findings shed light on its nature. First, the bias did not appear in a task that maps more than one stimulus to each response, consistent with the view that a signaling heuristic is less useful under such conditions. Second, the bias was stronger when the two letters were highly discriminable than when they were less discriminable, suggesting that more discriminable signals exert a stronger influence on response selection. Third, the bias was stronger when the interval between the previous response and the next stimulus was 300 ms relative to 1,200 ms, suggesting that a representation of the previous stimulus decays with time. These effects are not clearly predicted by the explanation of partial repetition costs provided by the binding hypothesis, wherein only the repetition of a previous feature triggers the retrieval of a previous event file. Therefore, these findings suggest the operation of a distinct mechanism.
Other data suggest that signaling influences performance even when participants receive little practice and more time separates consecutive trials. First, in visual search tasks, switching between (a) target-defining features (e.g., red vs. blue) within a single dimension (e.g., color) or (b) target-defining dimensions (e.g., color and location) engenders a bias to switch away from the previous response (e.g., Tollner et al., 2008). Second, computational models suggest that sequential trial effects in two-choice tasks arise partly from representing each stimulus as a repetition or an alternation of the previous stimulus, rather than solely in terms of its specific identity (e.g., A or B) (Jones et al., 2013). Specifically, Jones et al. (2013, page 657) argue that
“. . individual trials are represented both as concrete events (specifically, by the physical response that was executed) and more abstractly as repetitions and alternations. These representations correspond to two possible strategies for performing the task: one can identify the stimulus and give the appropriate response, or one can identify whether the stimulus changed from the previous trial and accordingly change or repeat the previous response (Fletcher & Rabbitt, 1978).”
Third, in some versions of the Simon task, the nature of sequential trial effects indicates that participants code identical consecutive responses as “repetitions” independent of their concrete spatial locations (i.e., left and right) (Wuhr & Heuer, 2015). These findings further suggest that participants code individual stimuli and responses as repetitions and alternations of previous stimuli and responses, rather than solely in terms of their specific identities, thereby enabling signaling to influence performance.
The binding plus signaling hypothesis
The findings above suggest that partial repetition costs may index a mixture of binding and signaling, rather than binding alone. This mixture may influence performance via four sequential processes. First, related to binding, participants form a memory – or event file – of each stimulus-response integration episode. Second, repeating a previous feature (e.g., the task) in the next trial triggers the retrieval of the previous event file, which leads to partial repetition costs. Third, related to signaling, each current stimulus feature (e.g., color, location, etc.) is compared to the corresponding, retrieved previous stimulus feature (e.g., color, location, etc.). Fourth, the outcome of each comparison – repetition or alternation – independently biases response selection toward a response repetition or a response alternation (cf., Notebaert & Soetens, 2003), which independently increases partial repetition costs for distinct stimulus features. In sum, the binding plus signaling hypothesis posits that binding and signaling each lead to better performance when a stimulus feature (e.g., color) and the response (e.g., left keypress) (a) both repeat (the complete repetition advantage) or (b) both alternate (the complete alternation advantage) relative to when (c) one changes while the other repeats.
The present study
The goal of the present study is to determine whether partial repetition costs index binding alone or binding plus signaling by comparing such costs in two- and four-choice versions of the PRC task (Hommel, 1998). To our knowledge, the binding hypothesis does not predict a decrease in partial repetition costs as the number of stimulus and response alternatives increases.Footnote 2 In line with this view, partial repetition costs related to bindings between task-relevant features (e.g., different responses) remain robust in four-choice tasks (see Frings et al., 2020, for a review).
In contrast, the binding plus signaling hypothesis predicts smaller partial repetition costs in a four-choice (vs. two-choice) PRC task, because a stimulus feature alternation does not signal which of the three alternative responses to make in a four-choice task (Açkay & Hazeltine, 2007). Selecting an alternative response can, therefore, neither facilitate performance in (a) complete alternation trials nor interfere with performance in (b) partial repetition trials wherein a stimulus feature alternates and the response repeats. Consequently, “signal-free” Footnote 3 complete alternation advantages – defined by contrasting these two trial types – should be relatively small as they reflect binding but not signaling. The binding plus signaling hypothesis does, however, still predict a large performance decrement in (a) partial repetition trials wherein a stimulus feature (e.g., color) repeats and the response alternates relative to (b) complete alternation trials. Since a stimulus feature repetition signals a response repetition even in a four-choice task, signaling can still interfere with performance in partial repetition trials wherein a stimulus feature repeats.
Finally, we note two additional considerations. First, partial repetition costs in the PRC task are larger for task-relevant stimulus features than for task-irrelevant stimulus features (Hommel, 1998, 2007). Thus, we will assess such costs separately for (1) the task-relevant stimulus feature (i.e., color) and (2) the task-irrelevant stimulus feature (i.e., location). Second, precisely because partial repetition costs in the PRC task are larger for task-relevant (vs. task-irrelevant) stimulus features, we may observe significant “signal-free” complete alternation advantages in a four-choice PRC task for (1) the task-relevant stimulus feature but not (2) the task-irrelevant stimulus feature. This outcome would suggest that binding alone is insufficient to produce “signal-free” complete alternation advantages for the task-irrelevant stimulus feature, especially if such advantages are robust in an otherwise-matched two-choice PRC task. In other words, this outcome would suggest that reducing the influence of signaling by using a four-choice PRC task eliminates complete alternation advantages for the task-irrelevant stimulus feature.
Experiment 1
In Experiment 1, we conducted an online study of a two-choice PRC task. Here, we sought to establish – in an online setting – the effect sizes for the partial repetition costs and complete alternation advantages that Huffman et al. (2020) reported in relation to (1) a task-relevant stimulus feature (one of two possible colors) and (2) a task-irrelevant stimulus feature (one of two possible locations). As we described earlier, both hypotheses make the same predictions for a two-choice PRC task. We reasoned, however, that establishing the sizes of these effects would prove useful for a critical follow-up experiment involving a four-choice version of the same online task that has four task-relevant colors – rather than just two – and two task-irrelevant locations.Footnote 4
Methods
Participants
Based on power analyses conducted with G*Power 3.1.9.2 (Faul et al., 2007), we sought usable data from 40 participants. The analyses employed an alpha level of 0.05 and the effect sizes for the interactions in mean response time (RT) between (1) color and response (\( {\eta}_p^2 \) = 0.81) and (2) location and response (\( {\eta}_p^2 \) = 0.29) that Huffman et al. (2020), Experiment 2) observed. The results showed that 40 participants would provide over 99% power for observing an interaction between color and response and over 97% power for observing an interaction between location and response. Note that each of the two factors in these interactions had only two levels. Thus, the power analyses were equivalent to those for paired t-tests.
Forty-one students from the Psychology Department subject pool at the University of Michigan participated for course credit. Students could participate only if they indicated via a subject pool questionnaire that they were not taking prescription or recreational psychoactive drugs, did not have any neurological or psychiatric disorders, and had no history of serious head injury or seizures. We excluded the data from one of the 41 participants who performed the task with less than 75% accuracy, leaving 40 participants for the final analyses (11 male, 29 female; mean age, 19 years: age range, 18–22 years). The University of Michigan’s Behavioral Sciences Institutional Review Board (IRB) determined that the study was exempt from oversight. Nonetheless, we obtained informed consent in accordance with the Declaration of Helsinki and, more generally, conducted the study in accordance with the regulations of the IRB.
Stimuli and apparatus
Three empty white boxes arranged vertically (i.e., an empty three-box array, height = 364 pixels, width = 135 pixels) remained on the screen at all times except for briefly after an incorrect response (see Experimental design). Three additional stimuli appeared in each trial: a response cue (“1” or “2”; 44 × 27 pixels) in the central box, a Cued Response Stimulus (blue or green) in the top or bottom box (121 × 135 pixels), and a Discrimination Response Stimulus (blue or green) in the top or bottom box (121 × 135 pixels). All stimuli appeared in white on a black background. Participants pressed the Z or M key to respond in each trial. We programmed the experiment in PsychoPy (Peirce et al., 2019), converted it to an online format on Pavlovia (https://pavlovia.org/), and presented it to participants via a web browser outside of the lab.
Experimental design
Each trial included six successive stimuli (Fig. 1). First, there was an empty three-box array (duration, 2,000 ms). Second, the response cue (1 or 2) appeared in the central box (duration, 500 ms). Third, there was an empty three-box array (duration, 500 ms). Fourth, the Cued Response Stimulus (blue or green) appeared in the top or bottom box (duration, until response). Fifth, there was an empty three-box array (duration, 500 ms). Sixth, and finally, the Discrimination Response Stimulus (blue or green) appeared in the top or bottom box (duration, until response). Each of the 32 possible combinations of response cue (1 or 2), Cued Response Stimulus color (blue or green), Cued Response Stimulus location (top or bottom), Discrimination Response Stimulus color (blue or green), and Discrimination Response Stimulus location (top or bottom) appeared once in a random order in each 32-trial block.
We instructed participants to make two responses in each trial. When the Cued Response Stimulus appeared, we asked participants to press the Z key with their left index finger if the preceding response cue was a “1” or the M key with their right index finger if the preceding response cue was a “2”. When the Discrimination Response Stimulus appeared, we asked participants to press the Z key with their left index finger to indicate one stimulus color (e.g., blue) and the M key with their right index finger to indicate the other stimulus color (e.g., green). We counterbalanced the key-color mapping for the latter stimulus across participants.
If a participant responded incorrectly to either the Cued Response Stimulus or the Discrimination Response Stimulus, an error message appeared immediately after the incorrect response. The message also described the correct S-R mapping for the appropriate stimulus (Cued Response or Discrimination Response) as a reminder of the task instructions. After 3,000 ms, the program moved on to the next scheduled event (i.e., to the Discrimination Response Stimulus in the same trial or to the Cued Response Stimulus at the beginning of the next trial).
Procedure
After reading a brief description of the study, participants provided informed consent by clicking “1” to proceed with the experiment. Next, they read the task instructions, which described the sequence of stimuli that would appear in each trial and the stimulus-response (S-R) mapping for (1) the Cued Response Stimulus and (2) the Discrimination Response Stimulus. The instructions indicated that participants should respond as quickly as possible without making mistakes. Participants completed a 32-trial practice block followed by eight 32-trial test blocks. In total, each of the two-way interactions that we investigated (i.e., between color and response and between location and response) was associated with 64 complete repetition trials, 64 partial repetition trials wherein the color repeated and the response alternated, 64 partial repetition trials wherein the color alternated and the response repeated, and 64 complete alternation trials.
Data analyses
We employed separate repeated-measured analyses of variance (ANOVAs) in JASP 0.14.1.0 (JASP Team, 2020) to analyze mean RT and mean error rate (ER) related to the Discrimination Response Stimulus. The ANOVAs included three factors: color (repeat, alternate), location (repeat, alternate), and response (repeat, alternate). In the analysis of mean RT, we excluded trials with outlier RTs (i.e., RTs more than 3 SnsFootnote 5 from their condition means) and errors (involving either response). In the analysis of mean ER, we excluded trials with outlier RTs and trials wherein participants responded incorrectly to the Cued Response Stimulus. Overall accuracy was 93.1%. The percentage of trials that were RT outliers was 7.1%.
We conducted a series of planned comparisons in JASP to assess the complete alternation advantage separately for each of the two typical interactions in the present PRC task: (1) the interaction between color and response and (2) the interaction between location and response. More specifically, we conducted paired, two-tailed t-tests to compare performance in complete alternation trials to performance in each type of partial repetition trial. We reasoned that observing better performance in complete alternation trials than in each type of partial repetition trial would indicate the typical complete alternation advantages that appear in the two-choice PRC task. We conducted analogous analyses to assess the complete repetition advantages in our data. We report standardized effect sizes using \( {\eta}_p^2 \) (for F-tests) and Cohen’s dz (for paired t-tests).
Transparency and openness
We report our rationale for all sample sizes, manipulations, dependent measures, and data exclusions, and follow journal article reporting standards (JARS) (Kazak, 2018). We used JASP 0.14.1.0 (JASP Team, 2020) to analyze the data. The task scripts, data analysis scripts, and raw data are available on the Open Science Framework (https://osf.io/5a4rd). The study design was not pre-registered.
Results
Mean response time (RT)
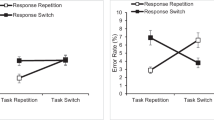
We observed two significant interactions. First, as expected, there was an interaction between color and response, F(1, 39) = 114.329, p < 0.001, \( {\eta}_p^2 \) = 0.75 (Fig. 3a). When the color repeated, mean RT was shorter when the response repeated (598 ms) than when the response alternated (641 ms). When the color alternated, mean RT was longer when the response repeated (666 ms) than when the response alternated (594 ms). Further analyses revealed that both indices of the complete alternation advantage were significant. Mean RT was shorter in complete alternation trials than in partial repetition trials wherein (a) the color alternated and the response repeated, t(39) = - 6.20, p < 0.001, dz = - 0.98, and (b) the color repeated and the response alternated, t(39) = - 4.79, p < 0.001, dz = - 0.76. Analogously, both indices of the complete repetition advantage were significant. Mean RT was shorter in complete repetition trials than in partial repetition trials wherein (a) the color alternated and the response repeated, t(39) = - 9.30, p < 0.001, dz = - 1.47, and (b) the color repeated and the response alternated, t(39) = - 5.36, p < 0.001, dz = - 0.85.

Partial repetition costs in the two-choice partial repetition cost (PRC) task. Panels a and c (top) show the interaction between color and response. Panels b and d (bottom) show the interaction between location and response. The left panels (a and b) indicate mean response time while the right panels (c and d) indicate mean error rate. Error bars indicate ± 1 SEM as calculated in JASP
Second, as expected, there was an interaction between location and response, F(1, 39) = 8.209, p < 0.007, \( {\eta}_p^2 \) = 0.17 (Fig. 3b). When the location repeated, mean RT was equivalent when the response repeated (625 ms) than when the response alternated (629 ms). When the location alternated, mean RT was longer when the response repeated (639 ms) than when the response alternated (605 ms). Further analyses revealed that both indices of the complete alternation advantage were significant. Mean RT was shorter in complete alternation trials than in partial repetition trials wherein (a) the location alternated and the response repeated, t(39) = - 4.41, p < 0.001, dz = - 0.70, and (b) the location repeated and the response alternated, t(39) = - 2.93, p = 0.006, dz = - 0.46. In contrast, neither index of the complete repetition advantage was significant. Mean RT did not differ between complete repetition trials and partial repetition trials wherein (a) the location alternated and the response repeated, t(39) = - 1.61, p = 0.12, dz = - 0.26, and (b) the location repeated and the response alternated, t(39) = - 0.32, p = 0.76, dz = - 0.05. Although the lack of a complete repetition advantage deviates from the typical pattern of partial repetition costs, it resembles the pattern of data that Huffman et al. (2020), Experiment 2) observed in the same task. No other effects were significant (all p > 0.08), although there was a trend toward a main effect of response (p = 0.087).
Mean error rate (ER)
We observed two significant main effects. First, there was a main effect of color, F(1, 39) = 6.92, p = 0.012, \( {\eta}_p^2 \) = 0.15, because mean ER was lower when the color repeated (4.1%) than when the color alternated (5.4%). Second, there was a main effect of response, F(1, 39) = 9.032, p = 0.005, \( {\eta}_p^2 \) = 0.19, because mean ER was lower when the response alternated (3.9%) than when the response repeated (5.6%).
We also observed three two-way interactions. First, as in the mean RT data, there was an interaction between color and response, F(1, 39) = 76.11, p < 0.001, \( {\eta}_p^2 \) = 0.66 (Fig. 3c). When the color repeated, mean ER was lower when the response repeated (1.9%) than when the response alternated (6.3%). When the color alternated, mean ER was higher when the response repeated (9.4%) than when the response alternated (1.4%). Further analyses revealed that both indices of the complete alternation advantage were significant. Mean ER was lower in complete alternation trials than in partial repetition trials wherein (a) the color alternated and the response repeated, t(39) = - 7.13, p < 0.001, dz = - 1.13, and (b) the color repeated and the response alternated, t(39) = - 7.72, p < 0.001, dz = - 1.22. Analogously, both indices of the complete repetition advantage were significant. Mean ER was lower in complete repetition trials than in partial repetition trials wherein (a) the color alternated and the response repeated, t(39) = - 7.14, p < 0.001, dz = - 1.13, and (b) the color repeated and the response alternated, t(39) = - 6.53, p < 0.001, dz = - 1.03.
Second, and also as in the mean RT data, there was an interaction between location and response, F(1, 39) = 26.45, p < 0.001, \( {\eta}_p^2= \)0.40 (Fig. 3d). When the location repeated, mean ER was lower when the response repeated (4.1%) than when the response alternated (5.7%). When the location alternated, mean ER was higher when the response repeated (7.2%) than when the response alternated (2.0%). Further analyses revealed that both indices of the complete alternation advantage were significant. Mean ER was lower in complete alternation trials than in partial repetition trials wherein (a) the location alternated and the response repeated, t(39) = - 5.27, p < 0.001, dz = - 0.83, and (b) the location repeated and the response alternated, t(39) = - 4.87, p < 0.001, dz = - 0.77. Analogously, both indices of the complete repetition advantage were significant. In particular, mean ER was lower in complete repetition trials than in partial repetition trials wherein (a) the location alternated and the response repeated, t(39) = - 3.64, p < 0.001, dz = - 0.58, and (b) the location repeated and the response alternated, t(39) = - 2.08, p = 0.044, dz = - 0.33.
Third, and unrelated to the present hypotheses, there was an interaction between color and location, F(1, 39) = 10.98, p = 0.002, \( {\eta}_p^2 \)= 0.220. As opposed to the normal “partial repetition cost” pattern, performance was worse in complete repetition and complete alternation trials than in partial repetition trials. When the color repeated, mean ER was higher (not lower) when the location repeated (5.1%) than when it alternated (3.1%). Analogously, when the color alternated, mean ER was higher (not lower) when the location alternated (6.1%) than when the location repeated (4.7%). We note that interactions between a task-relevant stimulus feature and a task-irrelevant stimulus feature that differ from the typical “partial repetition cost” pattern appear sporadically in the literature (e.g., Hommel & Colzato, 2004). However, the mechanisms underlying these effects remain unclear. No other effects were significant (all p > 0.21).
Discussion
In line with both of the hypotheses under investigation, we observed significant partial repetition costs and complete alternation advantages for the task-relevant feature (color) and the task-irrelevant feature (location) in a two-choice PRC task similar to that of Huffman et al. (2020), Experiment 2). Critically, we established the sizes of these effects in an online testing environment while distinguishing between the two types of partial repetition trials. As we describe next, this allowed us to calculate an appropriate sample size for investigating the extent to which each of these effects also appears in a four-choice version of the same online task.
Experiment 2
In Experiment 2, we investigated whether the partial repetition costs and complete alternation advantages that we observed in the two-choice PRC task of Experiment 1 generalize to a four-choice PRC task. To our knowledge, the binding hypothesis predicts they will generalize as there is no stated reason why partial repetition costs should vary with the number of stimulus and response alternatives (Hommel, 1998). In contrast, the binding plus signaling hypothesis predicts they will not generalize – at least not fully. As we explained in the Introduction, it predicts a smaller advantage in a four-choice (vs. two-choice) task for (a) complete alternation trials relative to (b) partial repetition trials wherein a stimulus feature alternates and the response repeats. That is, as the influence of signaling should be reduced in a four-choice (vs. two-choice) PRC task, the binding plus signaling hypothesis predicts relatively small “signal-free” complete alternation advantages.
Since partial repetition costs are larger for task-relevant as compared to task-irrelevant stimulus features, (Hommel, 2004), one possible outcome is that we will observe “signal-free” complete alternation advantages for (a) the task-relevant stimulus feature but not (b) the task-irrelevant stimulus feature. This outcome would suggest that the contribution of binding to partial repetition costs related to the task-irrelevant stimulus feature is not sufficient on its own (i.e., without signaling) to produce a “signal-free” complete alternation advantage. Therefore, such an outcome would be consistent with the binding plus signaling hypothesis
Finally, we note that the binding plus signaling hypothesis also predicts smaller partial repetition costs – as indexed by smaller two-way interactions between (1) color and response and (2) location and response – in a four-choice PRC task than in a two-choice PRC task. Later, we conduct exploratory, across-experiment analyses to investigate this additional prediction.
Methods
Participants
Based on power analyses conducted with G*Power 3.1.9.2 (Faul et al., 2007), we sought usable data from 66Footnote 6 healthy participants. The analyses employed an alpha level of 0.05 and the effect sizes for the (1) color by response (\( {\eta}_p^2 \) = 0.75) and (2) location by response (\( {\eta}_p^2 \) = 0.17) interactions in the mean RT from Experiment 1. The results indicated that 65 participants would provide over 99% power for observing an interaction between color and response and over 97% power for observing an interaction between location and response. They also showed that, for each of these two-way interactions, this sample size would provide over 97% power for observing a significant difference in performance between complete alternation trials and each type of partial repetition trial. Finally, the results showed that this sample size would provide over 97% power for observing the corresponding effects in mean ER.
Seventy students from the Psychology Department subject pool at the University of Michigan participated for course credit. Students could participate only if they met the same inclusion criteria as in Experiment 1. We excluded the data from four participants who performed the task with less than 75% accuracy, leaving 66 participants (26 males, 40 females; mean age, 19 years: age range, 18–22 years). The University of Michigan’s Behavioral Sciences Institutional Review Board (IRB) determined that the study was exempt from oversight. Nonetheless, we obtained informed consent in accordance with the Declaration of Helsinki and, more generally, conducted the study in accordance with the regulations of the IRB.
Stimuli and apparatus
The stimuli and apparatus were identical to those in Experiment 1 with four exceptions that were necessary to create a four-alternative forced-choice (4-AFC) task. First, the response cue was one of four (vs. two) possible digits (i.e., “1”, “2”, “3”, and “4”). Second, the Cued Response Stimulus appeared in one of four (vs. two) possible colors (blue, red, yellow, or green). Third, the Discrimination Response Stimulus appeared in one of four (vs. two) possible colors (blue, red, yellow, or green). Fourth, participants pressed one of four (vs. two) possible keys (Z, C, B, or M).
Experimental design
The experimental design was identical to that in Experiment 1 with three exceptions. First, each of the 256 (vs. 32) possible combinations of Response Cue (“1”, “2”, “3”, “4”), Cued Response Stimulus color (blue, red, yellow, green), Cued Response Stimulus location (top or bottom), Discrimination Response Stimulus color (blue, red, yellow, green), and Discrimination Response Stimulus location (top or bottom) appeared once randomly. To keep the block length consistent with that in Experiment 1, we divided these 256 trials into eight 32-trial blocks. Second, when the Cued Response Stimulus appeared in each trial, participants made one of four (vs. two) responses. Specifically, they pressed the Z key with their left middle finger, the C key with their left index finger, the B key with their right index finger, and the M key with their right middle finger if the preceding Response Cue was a “1”, “2”, “3”, or “4”, respectively. Third, when the Discrimination Response Stimulus appeared, participants also made one of four (vs. two) responses. Half of the participants pressed the Z key with their left middle finger, the C key with their left index finger, the B key with their right index finger, and the M key with their right middle finger if the Discrimination Response Stimulus was blue, red, yellow, or green, respectively. The other participants pressed Z for green, C for yellow, B for red, and M for blue.
Procedure
The procedure was identical to that in Experiment 1 with one exception: there were 256 unique trials, rather than 32. Thus, although participants completed a 32-trial practice block followed by eight 32-trial test blocks, each test block did not contain an equal number of trials from each condition. Rather, we randomized the entire sequence of 256 test trials at the beginning of the experiment, and participants proceeded through the sequence in 32-trial blocks. In total, each of the two-way interactions that we investigated (i.e., between color and response and between location and response) was associated with 16 complete repetition trials, 48 partial repetition trials wherein the color repeated and the response alternated, 48 partial repetition trials wherein the color alternated and the response repeated, and 144 complete alternation trials.
Data analyses
The data analyses were identical to those in Experiment 1.Footnote 7 Overall accuracy was 92.3%, and the percentage of trials that were RT outliers was 7.7%.
Transparency and openness
The transparency and openness were the same as in Experiment 1 with one exception. We pre-registered Experiment 2 on the Open Science Framework (https://osf.io/5tw7e).
Results
Mean RT
We observed one significant main effect. Specifically, there was a main effect of response, F(1, 65) = 6.95, p = 0.01, \( {\eta}_p^2= \)0.091, because mean RT was shorter when the response repeated (908 ms) than when the response alternated (938 ms). A trend toward a main effect of color did not achieve significance, F(1, 65) = 3.39, p = 0.07, \( {\eta}_p^2=0.050. \)
We also observed a pair of significant two-way interactions. First, as in Experiment 1, there was an interaction between color and response, F(1, 65) = 22.16, p < 0.001, \( {\eta}_p^2= \)0.25 (Fig. 4a). When the color repeated, mean RT was shorter when the response repeated (866 ms) than when the response alternated (954 ms). When the color alternated, mean RT was longer when the response repeated (950 ms) than when the response alternated (922 ms). Critically, there was only a non-significant trend toward a “signal-free” complete alternation advantage: mean RT was numerically (but not significantly) shorter in complete alternation trials than in partial repetition trials wherein the color alternated and the response repeated, t(65) = - 1.98, p = 0.052, dz = - 0.24. Mean RT was also shorter in complete alternation trials than in partial repetition trials wherein the color repeated and the response alternated, t(65) = - 2.78, p = 0.007, dz = - 0.34. Finally, both indices of the complete repetition advantage were significant. Mean RT was shorter in complete repetition trials than in partial repetition trials wherein (a) the color alternated and the response repeated, t(65) = - 3.51, p < 0.001, dz = - 0.43, and (b) the color repeated and the response alternated, t(65) = - 4.53, p < 0.001, dz = - 0.56.

Partial repetition costs in the four-choice partial repetition cost (PRC) task. Panels a and c (top) show the interaction between color and response. Panels b and d (bottom) show the interaction between location and response. The left panels (a and b) indicate mean response time while the right panels (c and d) indicate mean error rate. Error bars indicate ± 1 SEM as calculated in JASP
Second, as in Experiment 1, there was an interaction between location and response, F(1, 65) = 7.09, p = 0.01, \( {\eta}_p^2= \)0.098 (Fig. 4b). When the location repeated, mean RT was shorter when the response repeated (884 ms) than when the response alternated (944 ms). When the location alternated, mean RT was equivalent when the response repeated (931 ms) and when the response alternated (932 ms). Critically, unlike in Experiment 1, neither index of the complete alternation advantage was significant. We did not observe a “signal-free” complete alternation advantage as mean RT did not differ between complete alternation trials and partial repetition trials wherein the location alternated and the response repeated, t(65) = 0.052, p = 0.96, dz = 0.006. Mean RT also did not differ between complete alternation trials and partial repetition trials wherein the location repeated and the response alternated, t(65) = - 1.49, p = 0.14, dz = - 0.18. In contrast, both indices of the complete repetition advantage were significant. Mean RT was shorter in complete repetition trials than in partial repetition trials wherein (a) the location alternated and the response repeated, t(65) = - 2.26, p = 0.027, dz = - 0.28, and (b) the location repeated and the response alternated, t(65) = - 3.71, p < 0.001, dz = - 0.45. No other effects were significant (all p > 0.13).
Mean ER
We observed a significant main effect of color, F(1, 65) = 5.95, p = 0.017, \( {\eta}_p^2= \)0.084. This main effect occurred because mean ER was lower when the color alternated (4.6%) than when the color repeated (5.7%). There was a trend toward a main effect of location, but it did not achieve conventional levels of significance, F(1, 65) = 3.57, p = 0.063, \( {\eta}_p^2=0.052. \)
We also observed a pair of two-way interactions. First, as in the mean RT data, we observed an interaction between color and response, F(1, 65) = 23.11, p < 0.001, \( {\eta}_p^2= \)0.26 (Fig. 4c). When the color repeated, mean ER was lower when the response repeated (4.2%) than when the response alternated (7.1%). When the color alternated, mean ER was higher when the response repeated (5.7%) than when the response alternated (3.6%). Critically, as in Experiment 1, both indices of the complete alternation advantage were significant. We observed a “signal-free” complete alternation advantage because mean ER was lower in complete alternation trials than in partial repetition trials wherein the color alternated and the response repeated, t(65) = - 3.88, p < 0.001, dz = - 0.48. Mean ER was also lower in complete alternation trials than in partial repetition trials wherein the color repeated and the response alternated, t(65) = - 5.190, p < 0.001, dz = - 0.64. Finally, both indices of the complete repetition advantage were significant. Mean ER was lower in complete repetition trials than in partial repetition trials wherein (a) the color alternated and the response repeated, t(65) = - 2.14, p = 0.037, dz = - 0.26, and (b) the color repeated and the response alternated, t(65) = - 3.25, p = 0.002, dz = - 0.40.
Second, as in the mean RT data, there was an interaction between location and response, F(1, 65) = 6.57, p = 0.013, \( {\eta}_p^2= \)0.092 (Fig. 4d). When the location repeated, mean ER was lower when the response repeated (4.9%) than when the response alternated (6.3%). When the location alternated, mean ER was higher when the response repeated (5.0%) than when the response alternated (4.4%). Critically, we did not observe a “signal-free” complete alternation advantage. That is, mean ER did not differ between complete alternation trials and partial repetition trials wherein the location alternated and the response repeated, t(65) = - 1.13, p = 0.26, dz = - 0.14. As in Experiment 1, however, mean ER was lower in complete alternation trials than in partial repetition trials wherein the location repeated and the response alternated, t(65) = - 3.44, p = 0.001, dz = - 0.42. Further analyses revealed that, unlike in Experiment 1, neither index of the complete repetition advantage was significant. Mean ER did not differ between complete repetition trials and partial repetition trials wherein (a) the location alternated and the response repeated, t(65) = - 0.26, p = 0.80, dz = - 0.032, or (b) the location repeated and the response alternated, t(65) = - 1.91, p = 0.061, dz = - 0.24.
Finally, and unexpectedly, we observed a three-way interaction among color, location, and response, F(1, 65) = 4.38, p = 0.04, \( {\eta}_p^2= \)0.063. Further inspection revealed especially high error rates when both the color and the location repeated but the response alternated. Thus, we speculate that participants found it difficult to inhibit the previous response to the Cued Response Stimulus when its color and its location both repeated in the Discrimination Response Stimulus (see Hommel & Colzato, 2004 for an analogous three-way interaction and interpretation).
Discussion
In Experiment 2, we used a four-choice PRC task to distinguish between the binding and binding plus signaling hypotheses. In line with an influence of binding, we observed a significant “signal-free” complete alternation advantage for the task-relevant stimulus feature (color) in the mean ER data (but not in the mean RT data). Furthermore, in line with a reduced influence of signaling, we did not observe significant “signal-free” complete alternation advantages for the task-irrelevant feature (location) in either the mean RT or the mean ER data, even though such advantages were significant in the two-choice PRC task of Experiment 1. These findings suggest that binding and signaling each contribute to partial repetition costs. Thus, they appear more consistent with the binding plus signaling hypothesis than with the binding hypothesis.
Exploratory analyses
The absence of a “signal-free” complete alternation advantage for the task-irrelevant feature in the four-choice PRC task could simply reflect the significant (trending) main effect of response in the mean RT (mean ER) data. Difficulty with alternating (vs. repeating) the response would impair performance to a greater extent in complete alternation trials than in partial repetition trials wherein the response repeats. Thus, the absence of a “signal-free” complete alternation advantage may index a main effect of response, rather than a reduction of signaling.
To investigate this possibility, we conducted exploratory across-experiment analyses. The aim here was to determine whether the two-way interactions between (1) color and response and (2) location and response – which constitute the most critical signatures of binding in some views (Frings et al., 2020; Hommel, 2004; Schmidt et al., 2016) – were significantly smaller in the four-choice (vs. two-choice) PRC task. We also determined whether any such reductions were associated with (1) smaller complete alternation advantages coupled with (2) preserved complete repetition advantages. Indeed, the latter effect would be consistent with the view that a stimulus feature repetition still “signals” a response repetition in a four-choice task. Critically, observing smaller two-way interactions in the four-choice (vs. two-choice) PRC task would favor the binding plus signaling account over the main effect of response account by indicating that other critical signatures of binding are, in fact, reduced when signaling is not possible.
Methods
We prepared the data to test these predictions as follows. First, we excluded trials as in the main analyses. Second, to control for large between-experiment differences in mean RT, we converted each participant’s trial-specific RT values into trial-specific z scores and computed each participant’s mean z-scored RT (mean z-RT) in every condition (Aschenbrenner & Balota, 2017). The z-scoring approach controls for the fact that standard deviation increases with overall RT. In other words, this approach accounts for the fact that a given difference in mean RT (e.g., 50 ms) is more meaningful when it comes from an RT distribution that has a relatively low standard deviation (e.g., 300 ms) as compared to a relatively high standard deviation (e.g., 500 ms). Third, we employed each participant’s condition-specific mean ER data without transformations because overall accuracy was similar in Experiments 1 (93.1%) and 2 (92.3%). Since we did not transform the mean ER data, the partial repetition costs in these data are identical to those in Experiments 1 and 2 (see Figs. 2 and 3). Consequently, Fig. 5 in the next section presents only the partial repetition costs that appeared in the mean z-RT data. Finally, after preparing the data in these ways, we tested each of the three predictions above.

Exploratory analyses of differences between partial repetition costs in the two-choice and four-choice partial repetition cost (PRC) tasks. Panels a and c (top) show the interaction between color and response while panels b and d (bottom) show the interaction between location and response. In addition, panels a and b (left) show the mean z-RT data from the two-choice PRC task while panels c and d (right) show the mean z-RT data from the four-choice PRC task. Error bars indicate ± 1 SEM as calculated in JASP
First, we determined whether the interactions between (1) color and response and (2) location and response were smaller in the four-choice PRC task of Experiment 2 than in the two-choice PRC task of Experiment 1. To this end, we conducted separate mixed ANOVAs on the mean z-RT data and the mean ER data. Each ANOVA consisted of three within-participants factors – color (repeat, alternate), location (repeat, alternate), and response (repeat, alternate) – and one between-participants factor – experiment (1, 2).
Second, we determined whether either index of the complete alternation advantage was smaller in the four-choice PRC task than in the two-choice PRC task. To this end, we conducted two pairs of independent (two-tailed) t-tests: one pair for the interaction between color and response and a second pair for the interaction between location and response. Each individual t-test assessed whether one of the two indices of the complete alternation advantage was smaller in the four-choice (vs. two-choice) PRC task. We conducted analogous t-tests on the mean z-RT and mean ER data.
Finally, we determined whether either index of the complete repetition advantage was smaller in the four-choice PRC task than in the two-choice PRC task. We did so by conducting t-tests that were analogous to those described in the prior paragraph. That is, we compared performance in complete repetition trials to performance in each type of partial repetition trial, separately for the interactions between (1) color and response and (2) location and response. We report standardized effect sizes using \( {\eta}_p^2 \) (for F-tests) and Cohen’s ds (for independent t-tests).
Results
Effects related to the interaction among experiment, color, and response
Consistent with the binding plus signaling hypothesis, we observed a significant interaction among experiment, color, and response (mean z-RT: F(1, 104) = 12.81, p < 0.001, \( {\eta}_p^2= \)0.11; mean ER: F(1, 104) = 18.18, p < 0.001, \( {\eta}_p^2= \)0.15). This three-way interaction occurred because the interaction between color and response was smaller in the four-choice PRC task than in the two-choice PRC task (e.g., for the mean z-RT data, see Fig. 5a and c). Follow-up analyses revealed that the complete alternation advantage was smaller in the four-choice PRC task, which controlled for signaling, than in the standard two-choice PRC task (e.g., the difference between the two points on the right side of Fig. 5c is smaller than the corresponding difference in Fig. 5a). Specifically, the difference in performance between (a) complete alternation trials and (b) partial repetition trials wherein the color alternated and the response repeated was smaller in the four-choice (vs. two-choice) PRC task (mean z-RT: t(104) = - 5.93, p < 0.001, ds = - 1.19; mean ER: t(104) = - 5.24, p < 0.001, ds = - 1.05).
Our second index of the complete alternation advantage related to color was also smaller in the four-choice (vs. two-choice) PRC task (e.g., for the mean z-RT data, the solid line in Fig. 5c has a shallower slope than the solid line in Fig. 5a). More concretely, the difference in performance between (a) complete alternation trials and (b) partial repetition trials wherein the color repeated and the response alternated was smaller in the four-choice (vs. two-choice) PRC task. This reduction was significant only in the mean z-RT data (mean z-RT: t(104) = - 3.59, p < 0.001, ds = - 0.72; mean ER: t(104) = - 1.30, p = 0.20, ds = - 0.26). Nonetheless, it further supports the binding plus signaling hypothesis because a reduction of signaling in complete alternation trials could reduce both indices of the complete alternation advantage.
Finally, we determined whether either index of the complete repetition advantage was smaller in the four-choice PRC task than in the two-choice PRC task. Consistent with the binding plus signaling hypothesis, there was no difference in the degree to which performance was better in (a) complete repetition trials than in (b) partial repetition trials wherein the color repeated and the response alternated (mean z-RT: t(104) = - 0.008, p = 0.994, ds = - 0.002 ; mean ER: t(104) = - 1.23, p = 0.22, ds = - 0.25). In the mean z-RT data, for example, the difference between the lower left and upper left points does not differ in Fig. 5a and c. Analogously, there was no difference in the degree to which performance was better in (a) complete repetition trials than in (b) partial repetition trials wherein the color alternated and the response repeated in the mean z-RT data, t(104) = - 1.77, p = 0.079, ds = - 0.36. Specifically, the difference between the lower left and upper right points does not differ in Fig. 5a and c. In the mean ER data, however, this index of the complete repetition advantage was significantly smaller in the four-choice (vs. two-choice) PRC task, t(104) = - 5.11, p < 0.001, ds = - 1.024. We speculate that less signaling of the incorrect response in partial repetition trials wherein the color alternated and the response repeated reduced this index of the complete repetition advantage in the four-choice PRC task.
Effects related to the interaction among experiment, location, and response
Consistent with the binding plus signaling hypothesis, we observed a significant interaction among experiment, location, and response (mean z-RT: F(1, 104) = 4.60, p = 0.034, \( {\eta}_p^2= \)0.042; mean ER: F(1, 104) = 10.86, p = 0.001, \( {\eta}_p^2= \)0.095). This three-way interaction occurred because the interaction between location and response was smaller in the four-choice PRC task than in the two-choice PRC task (e.g., for the mean z-RT data, see Fig. 5b and d). Follow-up analyses revealed that our measure of the “signal-free” complete alternation advantage was smaller in the four-choice PRC task, which controlled for signaling, than in the standard two-choice PRC task (e.g., the difference between the two points on the right side of Fig. 5d is smaller than the corresponding difference in Fig. 5b). In other words, the difference in performance between (a) complete alternation trials and (b) partial repetition trials wherein the location alternated and the response repeated was smaller in the four-choice (vs. two-choice) PRC task (mean z-RT: t(104) = - 4.33, p < 0.001, ds = - 0.87; mean ER: t(104) = - 3.21, p = 0.002, ds = - 0.64).
Our second index of the complete alternation advantage related to location was also smaller in the four-choice (vs. two-choice) PRC task (e.g., for the mean z-RT data, the solid line in Fig. 5d has a shallower slope than the solid line in Fig. 5b). More concretely, the difference in performance between (a) complete alternation trials and (b) partial repetition trials wherein the location repeated and the response alternated was smaller in the four-choice (vs. two-choice) PRC task (mean z-RT: t(104) = - 3.33, p = 0.001, ds = - 0.67; mean ER: t(104) = - 3.26, p = 0.001, ds = - 0.65). This further supports the signaling hypothesis. Indeed, a reduction of signaling in complete alternation trials could reduce both indices of the complete alternation advantage.
Finally, we determined whether either index of the complete repetition advantage was smaller in the four-choice PRC task than in the two-choice PRC task. There was no difference in the degree to which performance was better in (a) complete repetition trials than in (b) partial repetition trials wherein the location repeated and the response alternated (mean z-RT: t(104) = 1.19, p = 0.24, ds = 0.24; mean ER: t(104) = - 0.207, p = 0.84, ds = - 0.04). For example, in the mean z-RT data, the difference between the lower left and upper left points does not differ in Fig. 5b and d. Analogously, there was no difference in the degree to which performance was better in (a) complete repetition trials than in (b) partial repetition trials wherein the location alternated and the response repeated in the mean z-RT data, t(104) = - 0.47, p = 0.64, ds = - 0.093. Specifically, the difference between the lower left and upper right points does not differ in Fig. 5b and d. In the mean ER data, however, this index of the complete repetition advantage was smaller in the four-choice (vs. two-choice) PRC task, t(104) = - 2.76, p = 0.007, ds = - 0.55). As we suggested earlier, less signaling of the incorrect response in partial repetition trials wherein the location alternated and the response repeated may have reduced this index of the complete repetition advantage in the four-choice PRC task.
Summary
The exploratory analyses yielded three principal results. First, the critical interactions between (1) color and response and (2) location and response were smaller in the four-choice PRC task than in the two-choice PRC task. Second, the smaller two-way interactions described above indexed reduced complete alternation advantages in the four-choice (vs. two-choice) PRC task. Third, in most cases, complete repetition advantages did not differ between the two tasks. These findings favor the binding plus signaling hypothesis over the main effect of response account by showing that other critical signatures of binding – the two-way interactions above – are reduced when signaling is not possible. Thus, they further suggest that signaling contributes to partial repetition costs, and that this contribution is smaller in the four-choice (vs. two-choice) PRC task.
General discussion
We sought to distinguish between the binding and binding plus signaling accounts of partial repetition costs in the PRC task. In line with an independent influence of binding on such costs, we observed a “signal-free” complete alternation advantage for the task-relevant feature in the mean ER data of the four-choice PRC task. In line with an independent influence of signaling on such costs, we did not observe “signal-free” complete alternation advantages for the task-irrelevant feature in the four-choice PRC task, even though the same contrasts were significant in the two-choice PRC task wherein signaling could occur. Further, exploratory analyses revealed that the two-way interactions between (1) color and response and (2) location and response – the critical signatures of binding in modern views (Frings et al., 2020; Schmidt et al., 2016) – were smaller in the four-choice (vs. two-choice) PRC task. These findings indicate that several signatures of binding are reduced when signaling is not possible and, therefore, suggest that binding and signaling each contribute to partial repetition costs in the PRC task. Thus, our findings appear more consistent with the binding plus signaling hypothesis than with the binding hypothesis.
Implications for the binding hypothesis
The present findings indicate a unique contribution of binding to partial repetition costs. First, we observed a “signal-free” complete alternation advantage related to the task-relevant stimulus feature (color) in the four-choice PRC task. In other words, we observed a complete alternation advantage even when controlling for the influence of a simple signaling heuristic (Açkay & Hazeltine, 2007; Fletcher & Rabbitt, 1978; Huffman et al., 2020; Kingstone, 1992; Krummenacher et al., 2009; Müller & Krummenacher, 2006; Notebaert & Soetens, 2003; Schoepper et al., 2020; Tollner et al., 2008). Second, we still observed interactions between (1) color and response and (2) location and response in the four-choice task of Experiment 2. As we stated earlier, such interactions are the critical signatures of binding in many views (e.g., Frings et al., 2020). Therefore, the present findings provide novel support for the view that binding-related processes influence action control in the PRC task (Frings et al., 2020; Hommel, 1998).
At the same time, we observed that two critical signatures of binding – (1) the complete alternation advantage and (2) partial repetition costs related to color and location – were smaller in the four-choice (vs. two-choice) PRC task. As we described earlier, some views of binding appear unable to explain this outcome. It is important to consider, however, whether other views of binding can explain the smaller partial repetition costs in our four-choice PRC task.
Recent findings from Hilchey and colleagues raise this possibility (Hilchey et al., 2017a, 2017b). In each trial of the authors’ two-choice tasks, participants made a lateralized (left or right) response to indicate the color (red or blue) of a square that appeared to the left or right of fixation, or at fixation. The authors reported smaller partial repetition costs (i.e., location by response interactions) at fixation than at the left and right locations. Critically, as in our four-choice PRC task, these reductions reflected smaller complete alternation advantages coupled with preserved complete repetition advantages. This outcome indicates that the selective reductions of the complete alternation advantage that we observed in our four-choice task can also appear in two-choice tasks. Thus, a factor other than reduced signaling may drive these reductions.
Hilchey et al. (2017b, page 898) provide the following speculative explanation:
“It is conceivable that, in the context of lateralized manual responding, lateralized target locations afford a more effective retrieval cue of the prior location-response code than a stimulus in central vision. If there is merit to this notion, we would expect location-response repetition effects to break down or become smaller in these tasks when the response set is not spatially-defined (e.g., Hommel, 2007; Hommel, 2011).”
This explanation suggests that partial repetition costs might be smaller in the four-choice (vs. two-choice) PRC task, because structural (including spatial) relationships between stimulus features and responses are more poorly defined when two responses (vs. one) are mapped to each hand.
Poorly defined relationships between stimulus features and responses may sometimes reduce partial repetition costs, but this view appears unlikely to explain either Hilchey and colleague’s findings or the present results. First, if repeating a central (vs. peripheral) target location less effectively triggers the retrieval of the previous location-response binding, then the complete repetition advantage should be smaller for central (vs. peripheral) targets. Hilchey and colleagues, however, observed a robust complete repetition advantage at fixation even while the complete alternation advantage vanished. The absence of a complete alternation advantage at fixation, but not in the periphery, may index other effects such as greater difficulty with switching attention from a peripheral location to a central location than vice versa. Indeed, it is more difficult to switch from a harder task (e.g., attending to the periphery) to an easier task (e.g., attending to fixation) than the other way around (Wylie & Allport, 2000). Second, the view that relationships between stimulus features and responses are more poorly defined in our four-choice (vs. two-choice) PRC task predicts a reduction of both the complete repetition advantage and the complete alternation advantage, rather than the selective reduction of the complete alternation advantage that we observed. Third, prior studies have revealed robust partial repetition costs and complete alternation advantages in four-choice tasks that map two responses to each hand, possibly because the associated bindings involved task-relevant features (i.e., responses) that receive heightened attention (Moeller & Frings, 2019, 2021).
Still, to test the structural account above, we conducted an exploratory analysis using the data from the four-choice PRC task. Specifically, we determined whether mean RT in complete alternation trials was slower when the response alternation involved a finger on (1) the same hand versus (2) the opposite hand (as in the two-choice PRC task). We reasoned that such a finding would support the view that structural relationships between stimulus features and responses are weaker within a hand than between hands. Contrary to this view, we did not observe a significant difference between the two mean RTs above when either the color (p > 0.06) or the location (p > 0.40) alternated with the response. Moreover, the numerical trend ran in the opposite direction in both cases. This outcome further suggests that the structural account described earlier cannot explain why increasing the number of stimulus-response alternatives from two to four reduces the complete alternation advantage without changing the complete repetition advantage.Footnote 8
Implications for the binding plus signaling hypothesis
The present findings suggest that partial repetition costs in the PRC task index a mixture of binding and signaling. This conclusion has important implications for the interpretation of previous findings from the two-choice PRC task (Hommel et al., 2011; Hommel & Frings, 2020; Zmigrod et al., 2013). For example, consider prior data indicating that partial repetition costs vary with the duration of the inter-trial-interval (Hommel & Frings, 2020). Although researchers interpreted this finding as indicating the gradual decay of event files over time, this finding could also reflect reduced signaling (cf., Fletcher & Rabbitt, 1978). Clearly distinguishing between binding and signaling in future studies of the PRC task may, therefore, provide a more fine-grained view of how recent experience influences action control. We discuss a possible method for accomplishing this objective next.
An important difference between the binding and binding plus signaling hypotheses is that only the latter hypothesis posits that a stimulus feature alternation biases response selection toward a response alternation in a two-choice task.Footnote 9 Since the present findings are consistent with this hypothesis, future studies could investigate whether it is possible to measure response activation related to signaling in the two-choice PRC task. To accomplish this objective, researchers could monitor the lateralized readiness potential (LRP) – a change in voltage on the scalp surface that is measured with electroencephalography (EEG) – during task performance. EEG studies of the LRP leverage the fact that primary motor regions in the left and right hemispheres execute opposite-side (i.e., contralateral) right- and left-hand responses, respectively. Indeed, since the LRP indexes the difference in neural activity between left and right primary motor regions, it allows researchers to measure the relative activation of contralateral right and left hand responses, respectively (Coles et al., 1988).
The binding plus signaling hypothesis makes interesting predictions for the two-choice PRC task. First, similar to the binding hypothesis, it predicts a deflection of the LRP toward the hemisphere contralateral to the previous response when a task-irrelevant stimulus feature repeats (signaling a response repetition). Second, and critically, it predicts a deflection of the LRP away from the hemisphere contralateral to the previous response when a task-irrelevant stimulus feature alternates (signaling a response alternation). The binding hypothesis does not predict this effect as there is no retrieval of the previous response when a stimulus feature alternates. Therefore, observing this effect would provide further support for the binding plus signaling hypothesis. It would also extend findings from other tasks showing that signaling can account for effects of repeating or alternating task-relevant information on the LRP (Tollner et al., 2008).
Observing electrophysiological evidence for an influence of signaling on partial repetition costs in the 2-AFC PRC task would highlight the need for a more detailed account of this phenomenon. Some current views suggest that sequential effects in 2-AFC choice RT tasks reflect implicit expectations about upcoming stimuli and responses (M. Jones et al., 2013). Such expectations are based on trial-by-trial estimates of (1) the proportion of trials in which each of the two responses is required and (2) whether the upcoming stimulus is likely to be a repetition or an alternation of the previous stimulus. Integrating the present version of the signaling hypothesis with these types of sequential learning could be a fruitful avenue for future research. Such an integration, if successful, would provide a more detailed account of this hypothesis than many other accounts offer (Açkay & Hazeltine, 2007; Fletcher & Rabbitt, 1978; Huffman et al., 2020; Kingstone, 1992; Krummenacher et al., 2009; Müller & Krummenacher, 2006; Notebaert & Soetens, 2003; Schoepper et al., 2020; Tollner et al., 2008).
Relationship of the present findings to prior data that appear inconsistent with signaling
Some data appear inconsistent with a contribution of signaling to partial repetition costs. First, Frings et al. (2007) reported that retrieving the previous response is more efficient when both the previous distractor and the previous target repeat than when only the previous target repeats. The authors argued that this finding weighs against a qualitative signaling mechanism that retrieves the previous response only if all previous trial stimulus features repeat in the current trial. They acknowledged, however, that it does not rule out a quantitative signaling mechanism wherein retrieval varies in a graded fashion with the degree of similarity between previous and current stimuli. Such a quantitative mechanism could resemble the current view of signaling, wherein each of several independent comparisons between current and previous stimulus features biases response selection toward a response repetition or a response alternation. Consistent with this view, the complete repetition advantage is especially large when all of the features of a multi-dimensional stimulus repeat relative to only some features (Bertelson, 1963).
Second, Frings and Rothermund (2011) reported partial repetition costs in a flanker task even when an unrelated yellow circle that required a unique response separated consecutive flanker task trials. The authors argued that signaling could not produce such costs, because the flanker stimulus always differed from the circle stimulus in the previous trial. Thus, a signaling heuristic would always bias response selection toward a response alternation. This does not, however, rule out signaling based on the most recent trial of the flanker task, as task repetitions trigger the retrieval of event files not only from the preceding trial but also from two trials back (Mayr et al., 2003; Spape & Hommel, 2008; Wendt et al., 2006).
Third, Hilchey and colleagues reported that interactions between (1) whether a location repeats or alternates and (2) whether the response repeats or alternates (i.e., location by response interactions) do not occur if participants need not attend to space in order to plan a response (e.g., left key or right key) (Hilchey et al., 2020). The authors argued that attention to space facilitates the retrieval of event files that store bindings between stimulus locations and response locations. Consequently, location by response interactions should not appear when attention to spatial processing is unnecessary to plan a response. One might initially interpret the absence of such interactions as providing evidence against signaling because stimulus feature repetitions and alternations do not bias response selection toward response repetitions and alternations, respectively. However, this is not the case. As the binding plus signaling hypothesis is dependent on event file retrieval, it predicts that signaling will not occur in the absence of such retrieval.
Broader implications
Our exploratory findings indicating smaller partial repetition costs in the four-choice (vs. two-choice) PRC task have important implications for the broader literature on action control. These findings suggest that partial repetition costs may be smaller in a wide variety of greater-than-two-choice tasks. Consistent with this possibility, current views of action control posit that the same processes that give rise to partial repetition costs in the PRC task contribute to a variety of other cognitive phenomena such as repetition priming, negative priming, distractor-response binding, congruency sequence effects, and task switching (for a review, see Frings et al., 2020).
Our findings supporting the binding plus signaling hypothesis resolve a paradox in the binding literature. If the task is simply another feature bound within the event file (Dignath et al., 2019; Egner, 2014; Frings et al., 2020), then repeating the task should trigger the retrieval of the previous response not only when a stimulus feature repeats but also when a stimulus feature alternates. Such retrieval should aid performance in complete repetition trials but impair performance in complete alternation trials. Thus, contrary to many findings, performance should always be better in complete repetition trials than in complete alternation trials. The binding plus signaling hypothesis resolves this paradox by positing that the outcome of each comparison (i.e., repetition or alternation) between a current stimulus feature (e.g., color, location, etc.) and the corresponding previous stimulus feature (e.g., color, location, etc.) biases response selection toward (i.e., “signals”) a response repetition or a response alternation. This can explain why performance is not always better in complete repetition trials than in complete alternation trials.
The present findings are also interesting in light of recent views wherein event files store abstract relationships between different stimuli. For example, these views posit that event files store whether a distractor and a subsequent target in a temporal flanker task are the same (i.e., congruent) or different (i.e., incongruent) (Egner, 2014; Weissman et al., 2016). Encoding such relationships requires comparing two stimuli, which has an obvious parallel to our view of signaling. In particular, we posit that signaling involves determining whether each stimulus feature in the current trial (e.g., color, location, etc.) is a repetition or an alternation of the corresponding stimulus feature in the previous trial via a series of independent comparisons.Footnote 10.Given this parallel, future studies might investigate whether overlapping processes contribute to the effects of signaling and congruency on performance. If they do, then signaling may produce sequential trial effects that are analogous to those produced by congruency (e.g., the congruency sequence effect) (Gratton et al., 1992). Recent findings appear consistent with this possibility (Hazeltine et al., in preparation).
Limitations
One may wonder whether differences in the relative frequency with which complete alternation and partial repetition trials appear lead to strategies that engender a smaller complete alternation advantage in the four-choice (vs. two-choice) PRC task. Specifically, complete alternation trials associated with the color by response and location by response interactions appear more frequently than partial repetition trials in the four-choice PRC task but less frequently than partial repetition trials in the two-choice PRC task. If participants prepare for each trial type in proportion to its frequency of occurrence, then the complete alternation advantage should be larger in the four-choice PRC task. However, we observed a smaller complete alternation advantage in the four-choice (vs. two-choice) PRC task. Thus, this account cannot easily explain our findings.
One may also wonder why evidence for a complete alternation advantage in the four-choice PRC task appeared for the task-relevant stimulus feature (i.e., color) but not for the task-irrelevant feature (i.e., location). The binding hypothesis suggests that the task-irrelevant stimulus feature is bound to the response to a lesser degree than the task-relevant stimulus feature (Hommel, 1998, 2007). In contrast, the signaling hypothesis suggests that the task-irrelevant stimulus feature might serve as a less effective signal for a response alternation because it receives less attention than the task-relevant stimulus feature. Given that the present findings suggest that both binding and signaling contribute to partial repetition costs, future studies could aim to distinguish between these not mutually exclusive possibilities.
Finally, one might ask why a complete alternation advantage appeared in the mean ER data of the four-choice PRC task but not in the mean RT data. This pattern mirrors data indicating that stimulus feature alternations sometimes facilitate quick responding less than they facilitate accurate responding (Notebaert & Soetens, 2003). In the four-choice PRC task, participants may rely on time-consuming decision-making processes in complete alternation trials – rather than on a decision-making heuristic – resulting in a relatively long mean RT. Although relying on such processes should also increase mean ER, mean ER may remain relatively low because the memory-retrieval conflicts that characterize partial repetition trials do not occur. Thus, reducing the influence of signaling in the four-choice PRC task may increase mean RT more than mean ER.
Conclusion
The present findings support the view that binding and signaling each contribute to partial repetition costs in the PRC task. Therefore, future studies investigating the relative contributions of binding and signaling to partial repetition costs in a wide variety of cognitive tasks may provide a more nuanced view of how recent experience influences action control.
Notes
We use the term “signaling” to emphasize that a repeating or alternating stimulus feature “signals” the cognitive system to bias response selection toward a response repetition or a response alternation, respectively. Functionally, the signaling hypothesis is similar to the “bypass rule” (Frings et al., 2007), which was originally formulated to explain related findings from earlier two-choice tasks (Fletcher & Rabbitt, 1978).
Dutzi and Hommel’s (2009) integrated competition hypothesis might predict this finding but integrated competition appears unlikely to influence partial repetition costs for the reasons we discussed earlier.
In this context, “signal-free” means that a stimulus feature alternation does not signal a single response alternative.
We use two locations in the four-choice PRC task of Experiment 2 to investigate whether increasing the number of S-R alternatives for the task-relevant feature (color) from two to four influences the complete alternation advantage for the task-irrelevant feature (location) while keeping the two irrelevant locations identical to those in Experiment 1.
In our pre-registration, we indicated a sample size of 65 healthy participants. However, we increased this number to 66 so that we could have an equal number of participants (i.e., 33) in each of the two S-R mapping conditions.
To further test this structural account, one could determine whether partial repetition costs in the two-choice PRC task are smaller when the task maps both responses to the same hand than when the task maps one response to each hand as is usually the case. Weighing against this possibility, the typical Simon effect depends on the relative locations of the two keys, rather than on the effectors that participants use to press those keys (Hommel, 2011).
Although Dutzi and Hommel’s (2009) integrated competition view might also predict this result, this account is unlikely to predict this result in the present two-choice PRC task for the reasons we provided in the Introduction.
Future studies could investigate whether the degree to which repeating or alternating a particular stimulus feature contributes to signaling increases with the feature’s task-relevance or perceptual salience. Such a result would be consistent with data showing that these factors influence binding (e.g., Hommel, 2004).
References
Açkay, C., & Hazeltine, E. (2007). Conflict monitoring and feature overlap: two sources of sequential modulations. Psychonomic Bulletin and Review, 14, 742–748.
Aschenbrenner, A. J., & Balota, D. A. (2017). Dynamic adjustments of attentional control in healthy aging. Psychology and Aging, 32(1), 1–15. https://doi.org/10.1037/pag0000148
Beck, D. M., & Kastner, S. (2005). Stimulus context modulates competition in human extrastriate cortex. Nature Neuroscience, 8(8), 1110–1116. https://doi.org/10.1038/nn1501
Beck, D. M., & Kastner, S. (2007). Stimulus similarity modulates competitive interactions in human visual cortex. Journal of Vision, 7(2), 19 11-12. https://doi.org/10.1167/7.2.19
Beck, D. M., & Kastner, S. (2009). Top-down and bottom-up mechanisms in biasing competition in the human brain. Vision Research, 49(10), 1154–1165. https://doi.org/10.1016/j.visres.2008.07.012
Bertelson, P. (1963). S-R relationships and reaction times to new versus repeated signals in a serial task. Journal of Experimental Psychology, 65, 478–484.
Coles, M. G., Gratton, G., & Donchin, E. (1988). Detecting early communication: Using measures of movement potentials to illuminate human information processing. Biological Psychology, 26, 69–89.
Desimone, R., & Duncan, J. (1995). Neural mechanisms of selective visual attention. Annual Review of Neuroscience, 18, 193–222.
Dignath, D., Hommel, B., Johannsen, L., & Kiesel, A. (2019). Reconciling cognitive-control and episodic-retrieval accounts of sequential conflict modulation: Binding of control-states into event-files. Journal of Experimental Psychology: Human Perception and Performance, 45(9), 1265–1270.
Duncan, J. (1996). Cooperating brain systems in selective perception and action. In T. Inui & J. L. McClelland (Eds.), Attention and Performance XVI (pp. 549–576). MIT Press.
Duncan, J., Humphreys, G., & Ward, R. (1997). Competitive brain activity in visual attention. Current Opinion in Neurobiology, 7(2), 255–261. https://doi.org/10.1016/s0959-4388(97)80014-1
Dutzi, I. B., & Hommel, B. (2009). The microgenesis of action-effect binding. Psychological Research, 73(3), 425–435. https://doi.org/10.1007/s00426-008-0161-7
Egner, T. (2014). Creatures of habit (and control): a multi-level learning perspective on the modulation of congruency effects. Frontiers in Psychology, 5, 1247. https://doi.org/10.3389/fpsyg.2014.01247
Faul, F., Erdfelder, E., Lang, A. G., & Buchner, A. (2007). G*Power 3: a flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior Research Methods, 39(2), 175–191.
Fletcher, B., & Rabbitt, P. M. (1978). The changing pattern of perceptual analytic strategies and response selection with practice in a two-choice reaction time task. Quarterly Journal of Experimental Psychology, 30(3), 417–427. https://doi.org/10.1080/00335557843000025
Frings, C., & Rothermund, K. (2011). To be or not to be...included in an event file: integration and retrieval of distractors in stimulus-response episodes is influenced by perceptual grouping. Journal of Experimental Psychology: Learning, Memory, and Cognition, 37(5), 1209–1227. https://doi.org/10.1037/a0023915
Frings, C., Rothermund, K., & Wentura, D. (2007). Distractor repetitions retrieve previous responses to targets. Quarterly Journal of Experimental Psychology (Hove), 60(10), 1367–1377. https://doi.org/10.1080/17470210600955645
Frings, C., Hommel, B., Koch, I., Rothermund, K., Dignath, D., Giesen, C., ... Philipp, A. (2020). Binding and Retrieval in Action Control (BRAC). Trends in Cognitive Science, 24(5), 375-387. https://doi.org/10.1016/j.tics.2020.02.004
Giesen, C., & Rothermund, K. (2014). Distractor repetitions retrieve previous responses and previous targets: experimental dissociations of distractor-response and distractor-target bindings. Journal of Experimental Psychology: Learning, Memory, and Cognition, 40(3), 645–659. https://doi.org/10.1037/a0035278
Grant, L. D., Cookson, S. L., & Weissman, D. H. (2020). Task sets serve as boundaries for the congruency sequence effect. Journal of Experimental Psychology. Human Perception and Performance, 46, 798–812. https://doi.org/10.1037/xhp0000750
Gratton, G., Coles, M. G. H., & Donchin, E. (1992). Optimizing the use of information: strategic control of activation and responses. Journal of Experimental Psychology: General, 4, 480–506.
Hilchey, M. D., Rajsic, J., Huffman, G., & Pratt, J. (2017a). Intervening response events between identification targets do not always turn repetition benefits into repetition costs. Attention, Perception, & Psychophysics, 79(3), 807–819. https://doi.org/10.3758/s13414-016-1262-9
Hilchey, M. D., Rajsic, J., Huffman, G., & Pratt, J. (2017b). Response-mediated spatial priming despite perfectly valid target location cues and intervening response events. Visual Cognition, 25, 888–902.
Hilchey, M. D., Rajsic, J., & Pratt, J. (2020). When do response-related episodic retrieval effects co-occur with inhibition of return? Attention, Perception, & Psychophysics, 82(6), 3013–3032. https://doi.org/10.3758/s13414-020-02020-3
Hommel, B. (1998). Event Files: Evidence for Automatic Integration of Stimulus-Response Episodes. Visual Cognition, 5(1/2), 183–216.
Hommel, B. (2004). Event files: feature binding in and across perception and action. Trends in Cognitve Science, 8(11), 494–500. https://doi.org/10.1016/j.tics.2004.08.007
Hommel, B. (2007). Feature integration across perception and action: event files affect response choice. Psychological Research, 71(1), 42–63. https://doi.org/10.1007/s00426-005-0035-1
Hommel, B. (2011). The Simon effect as tool and heuristic. Acta Psychologica, 136(2), 189–202. https://doi.org/10.1016/j.actpsy.2010.04.011
Hommel, B., & Colzato, L. (2004). Visual attention and the temporal dynamics of feature integration. Visual Cognition, 11(4), 483–521.
Hommel, B., & Frings, C. (2020). The disintegration of event files over time: Decay or interference? Psychonomic Bulletin and Review, 27(4), 751–757. https://doi.org/10.3758/s13423-020-01738-3
Hommel, B., Müsseler, J., Aschersleben, G., & Prinz, W. (2001). The Theory of Event Coding (TEC): a framework for perception and action planning. Behavioral and Brain Sciences, 24(5), 849–878.
Hommel, B., Kray, J., & Lindenberger, U. (2011). Feature integration across the lifespan: stickier stimulus-response bindings in children and older adults. Frontiers in Psychology, 2, 268. https://doi.org/10.3389/fpsyg.2011.00268
Huffman, G., Hilchey, M. D., Weidler, B. J., Mills, M., & Pratt, J. (2020). Does feature-based attention play a role in the episodic retrieval of event files? Journal of Experimental Psychology: Human Perception and Performance, 46(3), 241–251. https://doi.org/10.1037/xhp0000709
JASP Team. (2020). JASP (Version 0.14.1.0): https://jasp-stats.org/.
Jones, P. R. (2019). A note on detecting statistical outliers in psychophysical data. Attention, Perception, & Psychophysics, 81(5), 1189–1196. https://doi.org/10.3758/s13414-019-01726-3
Jones, M., Curran, T., Mozer, M. C., & Wilder, M. H. (2013). Sequential effects in response time reveal learning mechanisms and event representations. Psychological Review, 120(3), 628–666. https://doi.org/10.1037/a0033180
Kazak, A. E. (2018). Editorial: Journal article reporting standards. American Psychologist, 73(1), 1–2. https://doi.org/10.1037/amp0000263
Kingstone, A. (1992). Combining expectancies. Quarterly Journal of Experimental Psychology, 44A(1), 69–104.
Krummenacher, J., Müller, H. J., Zehetleitner, M., & Geyer, T. (2009). Dimension- and space-based intertrial effects in visual pop-out search: modulation by task demands for focal-attentional processing. Psychological Research, 73, 186–197.
Luck, S. J., Chelazzi, L., Hillyard, S. A., & Desimone, R. (1997). Neural mechanisms of spatial selective attention in areas V1, V2, and V4 of macaque visual cortex. Journal of Neurophysiology, 77(1), 24–42. https://doi.org/10.1152/jn.1997.77.1.24
Mayr, U., Awh, E., & Laurey, P. (2003). Conflict adaptation effects in the absence of executive control. Nature Neuroscience, 6, 450–452.
Miller, E. K., & Cohen, J. D. (2001). An integrative theory of prefrontal cortex function. Annual Review of Neuroscience, 24, 167–202.
Moeller, B., & Frings, C. (2019). From simple to complex actions: Response-response bindings as a new approach to action sequences. Journal of Experimental Psychology: General, 148(1), 174–183. https://doi.org/10.1037/xge0000483
Moeller, B., & Frings, C. (2021). Response-response bindings do not decay for 6 seconds after integration: A case for bindings' relevance in hierarchical action control. Journal of Experimental Psychology. Human Perception and Performance, 47(4), 508–517. https://doi.org/10.1037/xhp0000897
Müller, H. J., & Krummenacher, J. (2006). Locus of dimension weighting: Preattentive or postselective? Visual Cognition, 14(4-8), 490–513.
Notebaert, W., & Soetens, E. (2003). The influence of irrelevant stimulus changes on stimulus and response repetition effects. Acta Psychologica, 112(2), 143–156. https://doi.org/10.1016/s0001-6918(02)00080-x
Peirce, J., Gray, J. R., Simpson, S., MacAskill, M., Hochenberger, R., Sogo, H., ... Lindelov, J. K. (2019). PsychoPy2: Experiments in behavior made easy. Behavioral Research Methods, 51(1), 195-203. https://doi.org/10.3758/s13428-018-01193-y
Rousseeuw, P. J., & Crouz, C. (1993). Alternatives to the Median Absolute Deviation. Journal of the American Statistical Association, 88(424), 1273–1283.
Schmidt, J. R., De Houwer, J., & Rothermund, K. (2016). The Parallel Episodic Processing (PEP) model 2.0: A single computational model of stimulus-response binding, contingency learning, power curves, and mixing costs. Cognitive Psychology, 91, 82–108. https://doi.org/10.1016/j.cogpsych.2016.10.004
Schoepper, L. M., Hilchey, M. D., Lappe, M., & Frings, C. (2020). Detection versus discrimination: The limits of binding accounts in action control. Attention, Perception, & Psychophysics, 82(4), 2085–2097. https://doi.org/10.3758/s13414-019-01911-4
Spape, M. M., & Hommel, B. (2008). He said, she said: episodic retrieval induces conflict adaptation in an auditory Stroop task. Psychonomic Bulletin and Review, 15(6), 1117–1121. https://doi.org/10.3758/PBR.15.6.1117
Tollner, T., Gramann, K., Müller, H. J., Kiss, M., & Eimer, M. (2008). Electrophysiological markers of visual dimension changes and response changes. Journal of Experimental Psychology: Human Perception and Performance, 34(3), 531–542. https://doi.org/10.1037/0096-1523.34.3.531
Weissman, D. H., Hawks, Z. W., & Egner, T. (2016). Different levels of learning interact to shape the congruency sequence effect. Journal of Experimental Psychology: Learning, Memory, and Cognition, 42(4), 566–583. https://doi.org/10.1037/xlm0000182
Wendt, M., Kluwe, R. H., & Peters, A. (2006). Sequential modulations of interference evoked by processing task-irrelevant stimulus features. Journal of Experimental Psychology: Human Perception and Performance, 32(3), 644–667. https://doi.org/10.1037/0096-1523.32.3.644
Wuhr, P., & Heuer, H. (2015). The coding of repetitions and alternations in action sequences: spatial or relational? Psychological Research, 79(3), 432–445. https://doi.org/10.1007/s00426-014-0579-z
Wylie, G., & Allport, A. (2000). Task switching and the measurement of "switch costs". Psychological Research, 63(3-4), 212–233. https://doi.org/10.1007/s004269900003
Zmigrod, S., de Donneville, L. M. J., Colzato, L., Swaab, H., & Hommel, B. (2013). Cognitive control of feature bindings: evidence from children with autistic spectrum disorder. Psychological Research, 77(2), 147–154.
Acknowledgements
The authors thank Emma Carter, Samantha Cerpa, and Raisa Dorzbach for assistance with creating the online tasks.
Code availability
The task and data analysis scripts for Experiments 1 and 2 are available on the Open Science Framework (https://osf.io/5a4rd).
Funding
The authors received no funding to conduct this study.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors have no relevant financial or non-financial interests to disclose.
Ethics approval
The University of Michigan’s Behavioral Sciences Institutional Review Board (IRB) determined that the study was exempt from oversight. Nonetheless, we obtained informed consent in accordance with the Declaration of Helsinki and, more generally, conducted the study in accordance with the regulations of the IRB.
Consent to participate
All subjects gave consent to participate via an online form before the online study began.
Consent for publication
All subjects were informed that we intended to publish the study’s results in the informed consent document. No subjects requested that their data be excluded from publication.
Additional information
Open practices statement
The task scripts, data analysis scripts, and raw data from Experiments 1 and 2 are available on the Open Science Framework (https://osf.io/5a4rd). We also pre-registered Experiment 2 on the Open Science Framework (https://osf.io/5tw7e).
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Weissman, D.H., Grant, L.D., Koch, I. et al. Partial repetition costs index a mixture of binding and signaling. Atten Percept Psychophys 85, 505–524 (2023). https://doi.org/10.3758/s13414-022-02539-7
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-022-02539-7