



Introduction
Following their arrival in 1519, the Spanish conquistadors took less than two years to conquer Aztec central Mexico; however, it took a further two decades to secure their rule over Yucatán and its Maya population (Figure 1). By the end of 1540, as part of the third Spanish colonisation campaign of Yucatán, Conquistador Francisco de Montejo ‘the Younger’ founded the villa of San Francisco de Campeche (Valencia Reference Valencia1939). This coastal Hispanic town replaced the nearby Maya settlement of Ah Kin Pech, after which it was named (Valencia Reference Valencia1939; Scholes & Roys Reference Scholes and Roys1996) (Figure 2: left). Its first formal church was erected on the central plaza to serve “Spaniards, mestizos, mulattos, blacks, Indian naboríos and seven other Indian tribes” (López de Cogolludo Reference López de Cogolludo1954: 386–87) (Figure 2: right). Later, this early parish church and its cemetery served all those baptised settlers who were not part of any of the surrounding district's congregations: visitors and recent immigrants, together with the residents of the houses that lined the blocks surrounding the main plaza. The latter area was, from the beginning of the settlement, reserved for Spanish residents, their servants and harbour workers (Antochiw Reference Antochiw, Tiesler, Zabala and Cucina2010; Price et al. Reference Price2012).

Figure 1. Colonial sources attest that Juan Cortés, slave of the soldier Juan Sedeño, was the first African registered in New Spain. He was part of the military consortium led by Hernán Cortés in 1519 (Bishop Diego Durán; image courtesy of Arqueología Mexicana/Raices).

Figure 2. Locations of (left) the town of Campeche (map by the authors) and (right) the old Novohispanic church within the fortified central plaza of San Francisco de Campeche in 1604 (source: Historical Military Service of Madrid, Spain).
Located on the coast of the Gulf of Mexico, the early Hispanic town of Campeche thus provides a unique opportunity to examine the intermingling of previously distinct cultures and populations. Declared a UNESCO World Heritage Site in 2000, its historic centre stands as a monumental landmark of the beginnings of Hispanic society. Among the archaeological discoveries within the town's early colonial core is a cemetery associated with the foundations of an early colonial church (Tiesler et al. Reference Tiesler, Zabala and Cucina2010). Excavations in 2000 (see below), recovered 129 interments from this complex. These were examined through a combination of standard archaeological and anthropological methods, and isotope analyses. Here, we present new genome-wide ancient DNA (aDNA) data for 10 of these individuals and evaluate and discuss the results alongside the previously published datasets. In doing so, we identify the areas where these data sources complement or contradict each other, and the implications of combining and interpreting multiple types of information about the life histories of people in the past.
The plaza cemetery and the early colonial population of Campeche
The early colonial parish church at Campeche was probably established only a few years after the town was founded, and remained in use until the present-day cathedral replaced it in 1680 (Tiesler et al. Reference Tiesler, Zabala and Cucina2010). At that point, the church was demolished to its foundations and the present-day cathedral was built nearby. Memory of the original parish church and associated cemetery faded as the central square and town grew. It was not until 2000 that archaeological rescue excavations, led by Mexico's Instituto Nacional de Antropología e Historia (INAH), revived the memory of the early parish church and its deceased colonial parishioners. These investigations identified 129 burials (Figure 3; see also online supplementary information (OSM)1). The analysis of bioarchaeological, archaeometric and conventional archaeological evidence, as well as historical records, indicates that this cemetery was in use for at least 140 years, from the mid-sixteenth to late seventeenth century (Tiesler et al. Reference Tiesler, Zabala and Cucina2010; Price et al. Reference Price2012; see also OSM1, 3 & 4). Following Catholic tradition, the densely packed burials took the form of simple, extended inhumations. Most of the articulated skeletons showed evidence for disturbance by subsequent occupations and a number of the burials featured purely commingled remains. Only two of the 129 individuals were found with associated personal items (Tiesler & Zabala Reference Tiesler, Zabala, Tiesler, Zabala and Cucina2010; Tiesler et al. Reference Tiesler, Zabala and Cucina2010).

Figure 3. Explorations at the main plaza of Campeche: left) burial distribution; middle) excavation in progress in 2000 (photograph courtesy of Instituto Nacional de Antropología e Historia (INAH)); right) grid of excavation area and test pits (courtesy of INAH & Laboratorio de Bioarqueología/Autonomous, University of Yucatan).
Conventional and archaeometric analyses were coordinated at the Laboratory of Bioarchaeology at the Autonomous University of Yucatán, Mexico (see OSM2). Macroscopic analysis using standard metric and non-metric techniques was combined with histological, isotopic and molecular studies (OSM1–3) to inform the reconstruction of the individuals’ age-at-death, sex estimation, geographical provenance, biological ancestry, living conditions, health, nutrition and artificial body modifications (Tiesler et al. Reference Tiesler, Zabala and Cucina2010). Macroscopic assessments revealed both male and female adolescents and adults, the majority of whom died before reaching their fifth decade. The combined studies point to elevated rates of metabolic and inflammatory conditions, as evidenced by cranial porotic hyperostosis, dental enamel hypoplasia and periosteal reactions. Developmental physiological stress seems to have been a common feature, especially among the local population—even surpassing those recorded for their pre-Hispanic Maya peers (Cucina Reference Cucina, Tiesler, Zabala and Cucina2010; Tiesler et al. Reference Tiesler2015).
The geographical origin and potential biological affinity of the individuals from the Campeche cemetery were previously examined through analysis of isotope values (conducted by a team at the University of Wisconsin in Madison, led by T. Douglas Price; see OSM3), and body modifications and dental morphological traits were scored on the permanent dentition (see OSM2). The profiling of strontium (87Sr/86Sr) and oxygen isotopes and comparisons with respective baseline data suggest that a considerable segment of the population consisted of first-generation immigrants from near and far, including at least a dozen individuals from West Africa (Figure 4). The results of isotope analyses correlate with biomorphological assessments: all individuals with signs of cultural head shaping were determined to be locals, while four cases of dental chipping—an African practice—were identified as non-locals (Price et al. Reference Price, Tiesler and Burton2006). The combination of all these lines of evidence culminated in an edited volume and article (Tiesler et al. Reference Tiesler, Zabala and Cucina2010; Price et al. Reference Price2012). The resulting understanding of individual life histories and layered assessments of the overall burial population from the cemetery have fostered an integrated, yet hypothetical, understanding of life and death in early colonial Campeche. It suggests that individuals of Native American, European and African origin were buried side by side, and potentially together with individuals of mixed heritage. Here, a decade later, we return to the Campeche cemetery population and open a new chapter of exploration through the presentation of novel palaeogenomic information.

Figure 4. Schematic plan of grave locations surrounding the early colonial church on Campeche's central plaza (figure by the authors).
Palaeogenetic analyses
To explore the early Hispanic cemetery and population of colonial Campeche in more detail, we can look to aDNA analysis. The first author (Tiesler) previously attempted, unsuccessfully, to extract aDNA data from the Campeche cemetery population in 2001 and 2007. These attempts, however, were prior to the subsequent exponential growth in aDNA studies and the discovery that the petrosal portion of the temporal bone often generates better results than other skeletal elements (Pinhasi et al. Reference Pinhasi2015). Encouraged by these developments, Tiesler decided to once again attempt aDNA analyses and selected the 10 petrous portions from the Campeche population, determined on the basis of visual inspection most likely to have the best preservation. The goals of this analysis were to:
1) determine whether it was possible to generate genome-wide data from the petrous portions of the Campeche individuals;
2) compare results from aDNA to the existing biomorphological and biomolecular data; and
3) explore any patterns evident in the aDNA data (OSM5).
We met our first goal and generated working data from all 10 of the petrosal portions selected for analysis (we also radiocarbon dated two of the individuals: see Figure 5, Table S2 and OSM4). Table 1 provides basic data from the analysis of these individuals (additional details are provided in Table S1). We then performed a suite of standard aDNA analyses to achieve our second and third goals. Determinations of genetic sex, mitochondrial haplogroups and Y-chromosome haplogroups were made for all individuals. No close genetic relatives were detected among the 10 individuals (up to third-degree relatives). Mitochondrial lineages for most individuals were either B2 or A2 (haplogroup lineages remain unchanged when restricting to reads with a characteristic damage signal of aDNA), which are both common among Indigenous Maya populations (González-Martín et al. Reference González-Martín2015; Söchtig et al. Reference Söchtig2015). Notable mitochondrial haplogroup outliers were noted for the individual from Burial 11–1, who has an L3 mitochondrial haplogroup, and Burial 52–1, who has an X2 haplogroup. These haplogroups are most common in African and Eurasian populations respectively (Reidla et al. Reference Reidla2003; Soares et al. Reference Soares2012), which correlates with whole genome data for these two individuals.

Figure 5. Plotted radiocarbon distributions of two individuals from the Campeche churchyard dated as part of this research (dates calibrated in OxCal v4.3.2, using the IntCal13 atmospheric curve; Reimer et al. Reference Reimer2013; Bronk Ramsey Reference Bronk Ramsey2017) (figure by the authors).
Table 1. Summary data from ancient DNA analysis of 10 Campeche individuals, with previous biomorphological assessments in brackets. See Table S1 for additional data. Notes: SNP = single nucleotide polymorphism; M = male; F = female; NID = not identified; EUR = European; AFR = African; AME = Native American.

We performed ADMIXTURE clustering analysis (Figure 6), principal component analysis (PCA) (Figure 7) and qpAdm analysis (Table S4) to provide insights into the population structure of the Campeche individuals and their relationship with previously reported data. These analyses reveal that, while most of the individuals cluster together as an unadmixed Native Campeche population, there are outliers, including one unadmixed European individual and one unadmixed African individual. We conducted qpAdm analysis to model, formally, the ancestry of the Campeche individuals. We attempted to model each of the individuals as a mixture of Mixe, Yoruba or French-related ancestry. These well-studied modern groups have a long history of genetic isolation from each other, with little mixture from other populations prior to European colonisation (Mendoza-Revilla et al. Reference Mendoza-Revilla2022), therefore allowing us to assess whether an individual's ancestry was more correlated with African, European or Native American populations. We found that individuals had ancestry proportions consistent with being entirely European (Burial 52–1), entirely sub-Saharan African (Burial 11–1), or entirely Native American (the remaining eight individuals) (Table S4).

Figure 6. Unsupervised ADMIXTURE clustering plots of the 10 individuals relative to highly diverged modern populations (for modern sample information, see Table S3 in the OSM). The small proportion of Han-related ancestry detected for Burial 30 at K = 4 is statistical noise due to the much lower dataset size (only approximately 40 000 SNPs) for this individual. The Native American-related ancestry detected for the modern French DG individual at K = 4 is also probably an artefact of the data (figure by the authors).

Figure 7. Principal components analysis of ancient individuals projected onto the principal components inferred from modern individuals consisting of Han Chinese, African (Yoruba and Mende), European (French and Finnish) and Native American (Pima, Chane and Mixe) individuals (for modern sample information, see Table S3 in the OSM).
A neighbour-joining tree with outgroup-f3 statistics (Raghavan et al. Reference Raghavan2014) of the form f3(Mbuti; Pop1, Pop2) was also created. This shows that the Campeche individuals were all more closely related to contemporary Maya individuals from southern Mexico than to Indigenous groups from central or northern Mexico (Figure 8). This suggests that these individuals came from the immediate region rather than from further afield in Mesoamerica.

Figure 8. Neighbour joining tree from outgroup-f3 statistics (above), and location of modern and ancient individuals used (below). Ancient Campeche individuals are most closely related to modern Maya, the group in closest geographic proximity (figure by the authors).
To determine whether there is any detectable substructure in the eight individuals consistent with Native American ancestry, we computed statistics of the form [D(Campeche.individual.A, Campeche.individual.B; Campeche.individual.C; YRI.SG)]. This method examines whether a test individual “Campeche.individual.C” shares more genetic alleles with “Campeche.individual.A” or “Campeche.individual.B”, using modern West Africans from Nigeria as an outgroup baseline (YRI.SG). Of all 252 (= 9 × 8 × 7/2) unique statistics of this type involving comparisons of the individuals inferred to have broadly Native American ancestry, the largest |Z|-score is 2.63, which is not significant after correcting for the number of hypotheses tested (p = 0.0087, Bonferroni corrected to p = 0.89). Thus, the eight Campeche individuals with typical Native American ancestry are unstructured within the limits of our resolution; in other words, these eight individuals could not be split into subgroups.
Individual profiles
Most of the 10 individuals analysed here—all but Burial 11–1 and Burial 52–1—have local, Indigenous population genetic profiles. ADMIXTURE analysis at K = 4 distinguishes the Native Campechean individuals from highly diverged modern populations (Figure 6). These eight individuals plot on the Native American portion of the PCA (Figure 7) and can be modelled as unadmixed Native Americans in qpAdm analysis (Table S4). Additionally, these individuals have either A2 or B2 mitochondrial haplogroups, which are the two most common haplogroups in contemporary Maya populations (González-Martín et al. Reference González-Martín2015; Söchtig et al. Reference Söchtig2015). The Y-chromosome haplogroups (Table 1) for males are also typical of Native American populations.
Indigenous burials
All eight individuals with Indigenous ancestry have isotopic values within the local range (Price et al. Reference Price2012). These individuals were recovered from different areas of the cemetery, and most showed disarticulation or other types of disturbance from post-funerary intrusions in the increasingly crowded cemetery (Figure 3; see also Figures S1–S2). Each individual was interred in a supine position (i.e. extended, on their back), in a simple, narrow pit, with the head orientated towards the setting sun. This was the uniform pattern of Catholic primary deposition within the cemetery, reflecting the Hispanic tradition of the early colonies (Tiesler & Zabala Reference Tiesler, Zabala, Tiesler, Zabala and Cucina2010). The burial arrangements and location of all eight Native individuals suggest that they had been baptised, allowing their admittance to the sanctified burial grounds after death. A number of the remains showed signs of chronic periostosis affecting the lower legs and feet, and porotic hyperostosis and enamel hypoplasia in the skull. Additionally, given the diagnostic macroscopic and histological features of postcranial segments (see OSM1), Burial 78 (either the individual genetically studied for this article, or another individual contained within this multiple, commingled burial) suffered from venereal syphilis (Rodríguez Pérez Reference Rodríguez Pérez, Tiesler, Zabala and Cucina2010). The sclerotic (dense) bone quality of a left fibular bone (evident macroscopically and radiographically) and the evocative histomorphological traits—namely delimitation banding (Grenzstreifen) and pillow formation (Polster)—suggest a long sequence of repetitive, active episodes of venereal treponematosis (Schultz Reference Schultz and Ortner2003).
Burial 11–1
The two remaining individuals studied here have entirely non-Native ancestry. Previous bioanthropological assessment of Burial 11–1 suggested that this individual was a female of African ancestry who died at a young age, probably in her early twenties (similar to most first-generation African immigrants in the overall Campeche series; see Tiesler et al. Reference Tiesler, Zabala and Cucina2010). Her strontium isotope ratio was one of the highest measured in the cemetery, indicating that she was not born in the Yucatán peninsula (see below in our use of gendered pronouns for Burials 11–1 and 52–1). She also had stable carbon and nitrogen isotope values suggesting that C3 plants (such as rice) constituted most of her childhood diet, which is consistent with early life residence somewhere in West Africa (Price et al. Reference Price2012). Her skeleton was found disarticulated and incomplete, indicating that she had been reburied. Her final resting place was 17m south-west of the main church entrance (Figure S2). This area was among the outer segments of the cemetery, and the burials here were closer to the ground surface than those nearer to the church. The outlying areas are likely to have harboured the latest interments of the cemetery, dating to the last decades of its use in the seventeenth century (Tiesler et al. Reference Tiesler, Zabala and Cucina2010). Genome-wide and mitochondrial (L3 haplogroup) data for Burial 11–1 are consistent with an African origin (Figures 6 & 7 and Table S4). The high-resolution genetic analysis allows us to determine that Burial 11–1 had negligible European or Native American ancestry (OSM5). While additional statistical analysis did not reveal a specific geographical origin for Burial 11–1 (OSM5), we were able to determine that her ancestry was broadly West African.
Burial 52–1
Previous bioanthropological analyses identified Burial 52–1 as an adult, middle-aged male, who was buried adjacent to the church wall (Tiesler et al. Reference Tiesler, Zabala and Cucina2010: 214 & 222). He was probably one of the first generation to be buried in the cemetery of the rapidly expanding town. Concave wear was observed between his right canine and first premolar. This indicates that a round, abrasive object was habitually kept at the same spot between the two arches, suggesting regular use of a tobacco pipe (Figure 9). Dental epigenetic traits are suggestive of sub-Saharan African or European ancestry (Scott & Turner 1997), but the 87Sr/86Sr isotopic value is not. Instead, this value from bone apatite is consistent with the local Campeche values, suggesting he had lived at or near Campeche for some time before death. Lead isotope values from this individual's tooth enamel, however, suggest he was born and spent at least part of his childhood in Europe, probably south-western Spain, which correlates with the aDNA results (Price et al. Reference Price2012). Our new genetic analysis indicates that this individual had unmixed European ancestry, and projection onto the PCA produced in Lazaridis et al. (Reference Lazaridis2016) suggests western Mediterranean ancestry (Figure 10). He had a Y-chromosome haplogroup of I2a2a1b, which is common in European populations. Thus, this individual was probably a European colonist who lived and died in Campeche but spent at least part of his youth in Europe—most likely Spain.
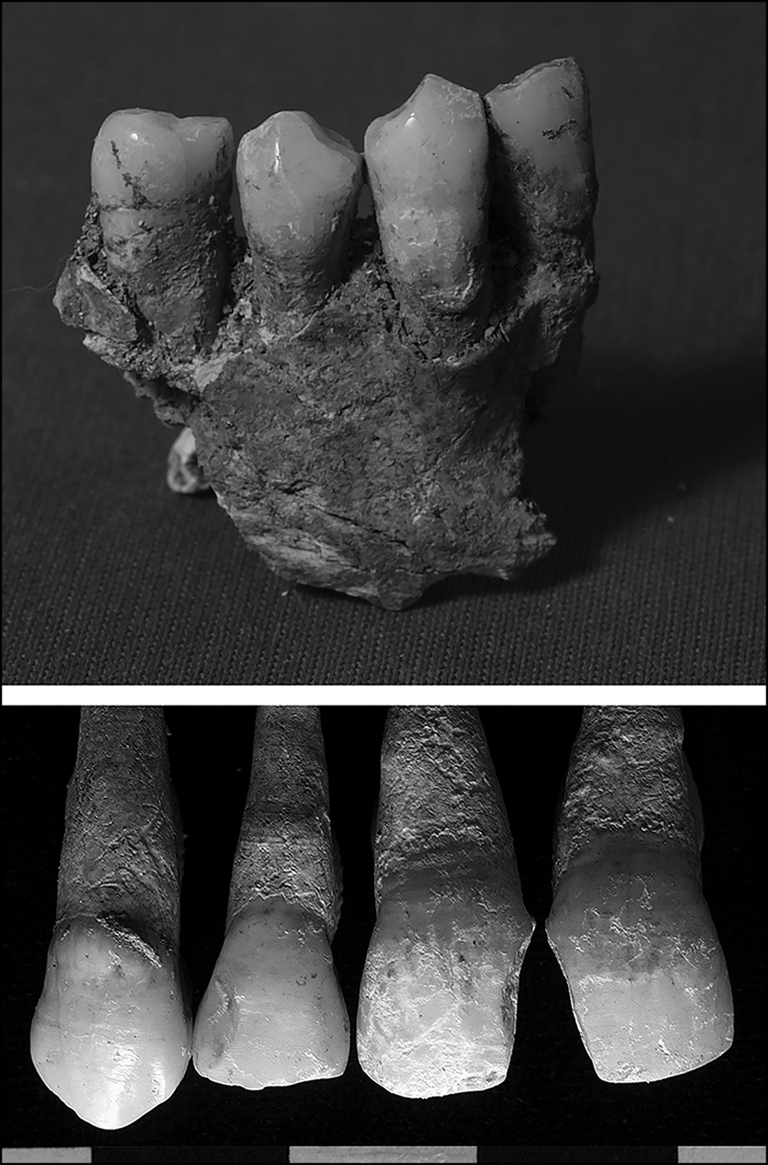
Figure 9. Ethnically distinctive cultural dental modifications: above) the lower dentition of male Burial 52–1 shows concave wear facets between the canine and first premolar, suggesting regular pipe use; below) cleft from chipping in two adjacent central incisors from a collective ossuary within the church perimeter (photographs by the authors).
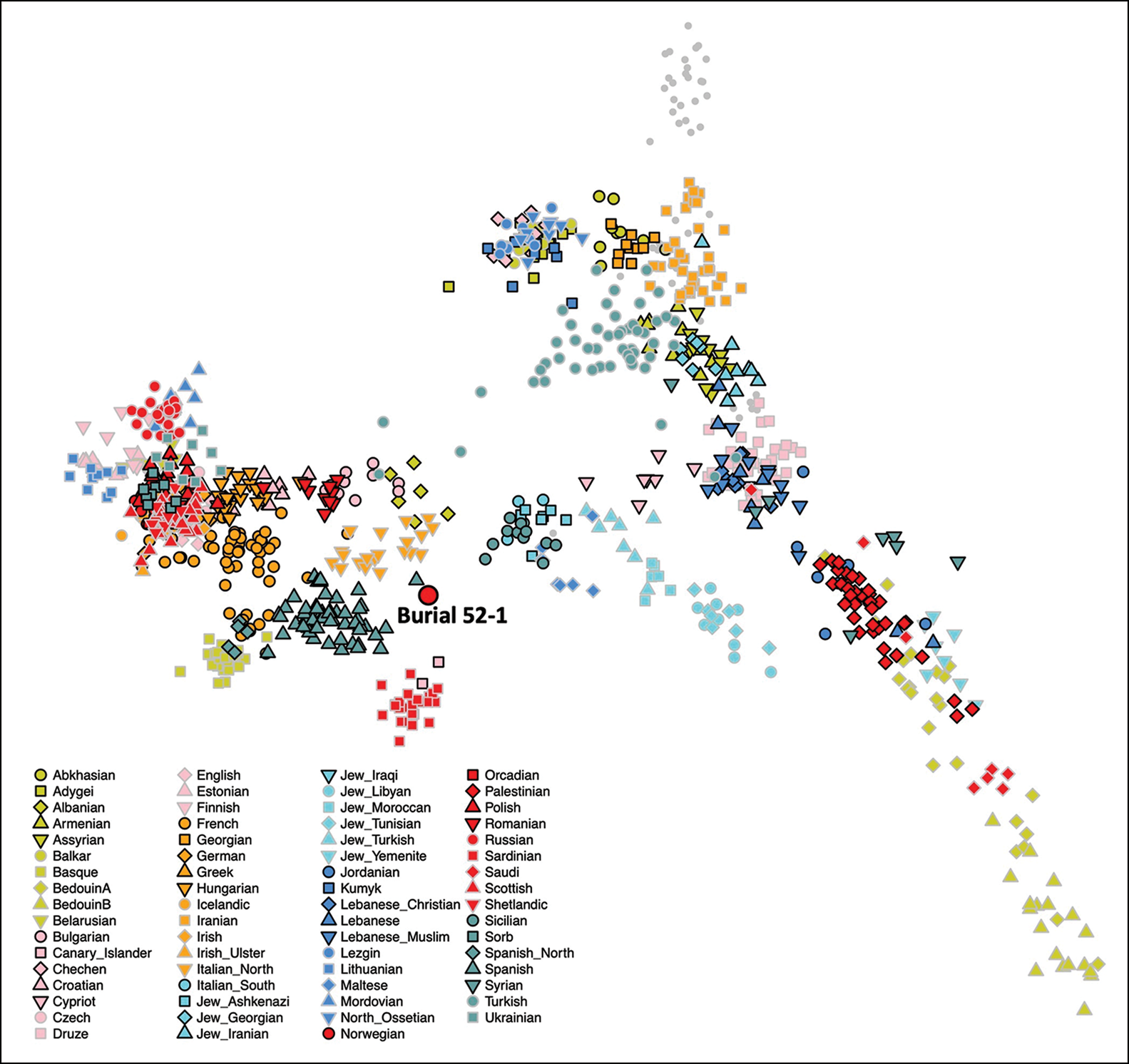
Figure 10. Burial 52–1 projected onto the Eurasian PCA from Lazaridis et al. (Reference Lazaridis2016: 420). This demonstrates that Burial 52–1 is genetically closest to individuals from modern-day Spain and northern Italy, suggesting an overall north-western Mediterranean ancestry (figure by the authors, based on data from Lazaridis et al. (Reference Lazaridis2016)).
We use singular gendered pronouns when discussing Burial 11–1 and 52–1 instead of the non-gendered ‘they’ because these individuals were part of multiple burials, thus the plural ‘they’ could mistakenly conflate all individuals in the burial when we are, in fact, referring only to the individual studied. Additionally, we are using ‘he’ and ‘she’ only in reference to measured X and Y-chromosome ratios, and recognise that the individuals may not have identified themselves in these gendered binaries while alive five centuries ago.
Burial 128 ass.
We provide dedicated discussion of Burial 128 ass. here not because of genetic ancestry—our analyses suggest that this individual was unambiguously Native American—but because of the inconsistency between our new genetic analyses and the previous macroscopic and isotopic analyses. This was a disturbed but largely articulated skeleton that was accompanied by isolated elements of at least three additional individuals. It is notable that segments of the cranium were found associated with the mandible but were not articulated and were therefore not drawn in the taphonomic illustration of the skeleton during excavation (Figure S3). Conventional analyses determined that the articulated individual—which we identify here as Burial 128–1—was probably male, based on robusticity, pelvic morphology and the elevated overall body length, as measured and calculated on-site (Tiesler & Zabala Reference Tiesler, Zabala, Tiesler, Zabala and Cucina2010). The previous isotopic study has also demonstrated that, like Burial 52–1, Burial 128–1 had a (femoral) bone apatite 87Sr/86Sr value that is consistent with local Campeche values (Price et al. Reference Price2012). The individual was therefore a Campeche resident during the last years of life. The carbon isotope ratios in this individual's tooth enamel (formed earlier in life), however, point to a dietary regimen that lacked C4 plants and therefore was very different compared with the typical Campeche diet. Additionally, the lead isotope values (measured in one enamel sample from the lower, articulated dental arch) are consistent with residence in south-west Spain during early childhood. Like Burial 52–1, these values, along with interment inside the church (probably denoting special status), led Price et al. (Reference Price2012) to classify the main individual of Burial 128 as a European migrant to the Americas (see Figure S3b).
Unlike Burial 52–1, however, palaeogenetic results for Burial 128 are inconsistent with both the isotopic values and the macroscopic sex estimation; our present analyses of the petrous portion—bagged and labelled in the field as part of this individual—demonstrate unadmixed Native American ancestry. We currently have no convincing explanation for this contradiction, other than that the sampled petrous portion (recorded in the field as contiguous to the mandible and the post cranium) belonged to a different, isolated adult (labelled here as ‘Burial 128 ass.’); we are unsure how these individuals became commingled. This highlights a methodological pitfall when using spatial association and morphological similarity to individualise disturbed assemblages such as this one. It also demonstrates how genetic analyses can make important contributions to the archaeological interpretations of highly commingled assemblages.
Discussion
There are noteworthy trends amongst the 10 Campeche individuals from whom we obtained aDNA. We have identified three distinct genetic groupings. Most were Indigenous ‘Campechanos’; however, we also documented one African female and a European male. None of the individuals within these groups has evidence of admixture from one another, despite living in close proximity and being interred in the same cemetery. While our sample size is relatively small, this lack of admixture is somewhat of a surprise, because the individuals were recovered from varied parts of the cemetery, with activity spanning 140 years—or some five to seven generations; some admixture, therefore, might have been expected over this length of time. Additionally, our selection of individuals for genetic study was based solely on the level of skeletal preservation rather than archaeological context. Indeed, Table 1 shows that most of the previous biomorphological assessments produced uncertain or indeterminate results. In other words, our sampling strategy was not designed to study individuals that we believed might harbour only one particular ancestry component based on phenotype or archaeological context.
Generation of these new aDNA data highlights the value of comparing the results of different types of investigation. Combined analysis of distinct datasets can capture the wide-ranging dynamics of a newly globalised world and generate perspectives about the past that are absent in documentary accounts (Tiesler et al. Reference Tiesler, Zabala and Cucina2010). This is especially true for colonial contexts, in which the voices of Indigenous peoples and Africans, as well as children and women, tend to be severely underrepresented, leading to a skewed perspective on the past. Multiple studies have combined archaeological, isotopic and DNA data to examine prehistoric Eurasia (e.g. Haak et al. Reference Haak2008; Knipper et al. Reference Knipper2017; Mittnik et al. Reference Mittnik2019); comparable studies that have been successfully conducted in the Americas are few, and especially so for the Maya Lowlands, where poor tissue preservation has hampered the recovery of aDNA. Thus, our ability to generate robust palaeogenomic data is highly encouraging for future studies of colonial-era sites in the Americas.
Points of tension between the historical accounts and the archaeological and palaeogenetic evidence are apparent from our study of the early colonial cemetery at Campeche, where settlers from different parts of the world and of varying social status were buried in close proximity. Spanish historical records inform us that, when founded in 1540, the villa of Campeche harboured 30 ‘residents’, with some 40 residents by 1562 and 100 in 1615 (Anonymous 1877: 370–71). These ‘residents’ are implied to have been Spaniards—either newcomers or local residents born in the colonies. These figures, however, exclude Native Americans, free Blacks and individuals of mixed African and European ancestry, who were not usually considered to be part of the resident population, as they lived at the edges of towns in so-called barrio neighbourhoods (Antochiw Reference Antochiw, Tiesler, Zabala and Cucina2010; Tiesler & Zabala Reference Tiesler, Zabala, Tiesler, Zabala and Cucina2010). In the planned Hispanic towns of New Spain, different ‘castas’ would live and die in their own ethnically distinct parishes, also called barrios. The social groups settled in this barrio system were characterised by acutely contrasting economic and living conditions (Martínez Reference Martínez2006). This situation prevailed in colonial Campeche itself, once its ethnically segregated neighbourhoods with their respective churches were in place (Antochiw Reference Antochiw, Tiesler, Zabala and Cucina2010).
No systematic information exists concerning the social segregation of cemeteries in New Spain. Theoretically, baptised parishioners could be admitted to sanctified burial grounds (Tiesler & Zabala Reference Tiesler, Zabala, Tiesler, Zabala and Cucina2010). In practice, however, documentary sources and the archaeological evidence indicate that circum-Caribbean Novohispanic urban cemeteries were typically segregated according to status and ethnicity (Stojanowsky Reference Stojanowsky2005). Further burial segregation could be expressed architecturally and by internal spatial divides (Jaén et al. Reference Jaén, Romano and Bautista2017; Pérez-Castro Reference Pérez-Castro2019).
While historical documents paint a picture of social demarcation and segregation, our palaeogenetic research demonstrates and confirms previous findings (Tiesler et al. Reference Tiesler, Zabala and Cucina2010), which show that individuals from populations who were segregated in most other aspects of life were indeed buried in the same Catholic cemetery. Most of the deceased seem to have been interred in austere, earthen pits, but were soon disturbed due to overcrowding and a general lack of organisation, which disturbed those previously interred.
This study also highlights how genetic inferences can sometimes conflict with other lines of investigation, such as dental ethnic assignment, conventional sex estimation, isotope analyses, or macroscopic identification of skeletal individuals in commingled and otherwise disturbed assemblages (such as the case of Burial 128). Only rough, conventional sex estimations could be made for six of the individuals studied here and could not be made at all for the other four due to commingling and poor skeletal preservation (Table 1). Genetic determinations, however, could be made for all 10 individuals. We view such instances as opportunities to re-evaluate discrete lines of evidence from different fields in order to provide a more critical, and therefore robust, understanding of the past.
Ancestry assessments founded on the classification of non-metric dental traits appear to be less secure than genetic assessment. Our results show some of the previously published biomorphological ancestry assessments (e.g. Tiesler & Zabala Reference Tiesler, Zabala, Tiesler, Zabala and Cucina2010; Tiesler et al. Reference Tiesler, Zabala and Cucina2010) to be inconsistent with palaeogenetic ancestry. An additional inconsistent observation relates to the nature of poorly preserved and disturbed secondary assemblages, especially when the sampled material originated from different anatomical parts, such as dental arches and petrous portions, in the case of Burial 128. Conversely, macroscopically determined cultural body modifications suitable for discerning origins, such as dental chipping or filing, as well as head shaping, were consistent with the palaeogenetic results, although they were of less diagnostic value.
The quality of aDNA preservation presented here is also notable. Until recently, acquisition of high numbers of single nucleotide polymorphisms (SNPs) from individuals buried in tropical environments, such as that of Campeche, was difficult and rare, as demonstrated by previous failed attempts to achieve results. Yet we have been able to produce working genomic data for all individuals in this study, with SNP counts of more than 500 000 for all but one individual. Moreover, the recovered genetic material is relatively uncontaminated by modern DNA and shows the typical damage profiles for aDNA samples (Figure S4). All genetic data in this study were procured through processing of disarticulated petrous bone in an aDNA clean room (see OSM5). These results highlight the effectiveness of using the petrous bone (Pinhasi et al. Reference Pinhasi2015, Reference Pinhasi2019; Hansen et al. Reference Hansen2017) for the recovery of aDNA in tropical environments.
Conclusion
In this study, we have combined new palaeogenetic data for 10 individuals previously subjected to a suite of conventional analytical methods in order to establish more detail about their life histories. These combined analyses now provide key details about the multi-ethnic nature of the people living in early colonial Campeche, where previously disparate groups mixed for the first time and lived under Hispanic rule. Singly or collectively, these genetic profiles add a new ‘human’ layer to our understanding of a crucial era and location in the history of New Spain. The spatial distribution of the locals and immigrants of different ancestries across the Campeche cemetery confirms a prompt and forced integration of all segments into the new Novohispanic social fabric. Our analysis of these 10 individuals found no evidence of mestizaje (mixing across groups). Our sample size is small but seems to indicate that although they were buried together, different groups in early colonial, urban, European hubs maintained some degree of separation in life. Future palaeogenetic analyses of the burial populations from other colonial sites, such as the San Román or San Francisco parishes, would undoubtedly shed new light on specific ancestries and any evidence of biological admixture. While the San Román neighbourhood was reportedly established and occupied by the Nahuas, whom Montejo brought from his encomienda in Azcapotzalco, the parishioners of the San Francisco neighbourhood were local Maya (López de Cogolludo Reference López de Cogolludo1954). The INAH has recently recovered human remains from both parish cemeteries. In this same vein, we are confident that future scrutiny of additional individuals from the central cemetery of Campeche will provide an even more nuanced picture of the timing of population mixing. We hope this research provides a starting point for further studies in this direction in the Americas and beyond.
Acknowledgements
This research ensued from the archaeological rescue project Registry, Conservation, Restoration and Study of the Archaeological and Human Remains Found in the Main Square of Campeche (directed by Heber Ojeda, Carlos Miguel Huitz and Vera Tiesler). Permission to study the Campeche individuals was granted by INAH's Consejo de Arqueología. We are grateful to all who have supplemented and aided this research: Pilar Zabala Aguirre, Carlos Huitz, Heber Ojeda, Michel Antochiw, Andrea Cucina, Carlos Vidal, Adriana Velázquez Morlet, Víctor Acuña, Enrique Morett, and the two anonymous reviewers of previous versions of this article.
Funding statement
D.R. is an Investigator of the Howard Hughes Medical Institute and his ancient DNA laboratory work was supported by an Allen Discovery Center grant and by grant 61220 from the John Templeton Foundation. N.N. is supported by a National Institute of General Medical Sciences (GM007753) fellowship.
Data availability statement
The aligned DNA sequences are available through the European Nucleotide Archive, accession no. PRJEB50901. Genotype data used in the analysis are freely available at https://reich.hms.harvard.edu/datasets. Any other relevant data are available from the corresponding author upon reasonable request.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.15184/aqy.2022.79.













