
Abstract
There is considerable evidence that salient items can be suppressed in order to prevent attentional capture. However, this evidence has relied almost exclusively on paradigms using color singletons as salient distractors. It is therefore unclear whether other kinds of salient stimuli, such as abrupt onsets, can also be suppressed. Using an additional singleton paradigm optimized for detecting oculomotor suppression, we directly compared color singletons with abrupt onsets. Participants searched for a target shape (e.g., green diamond) and attempted to ignore salient distractors that were either abrupt onsets or color singletons. First eye movements were used to assess whether salient distractors captured attention or were instead suppressed. Initial experiments using a type of abrupt onset from classic attentional capture studies (four white dots) revealed that abrupt onsets strongly captured attention whereas color singletons were suppressed. After controlling for important differences between the onsets and color singletons – such as luminance and color – abrupt-onset capture was reduced but not eliminated. We ultimately conclude that abrupt onsets are not suppressed like color singletons.
Similar content being viewed by others

Introduction
Visual attention often seems to be automatically drawn to salient stimuli. For example, when searching for a parking space in a crowded lot, one might involuntarily shift attention toward a uniquely colored sign (a color singleton; Theeuwes, 1992) or the flashing hazard lights of a car (an abrupt onset; Todd & Van Gelder, 1979; Yantis & Jonides, 1984). Extensive research efforts have been made to better understand this type of visual distraction, known as attentional capture. Recent evidence has suggested that participants can learn to suppress salient items in order to prevent attentional capture (Luck et al., 2021). However, this evidence has relied almost exclusively on paradigms using color singletons as distracting stimuli. The current study therefore evaluates suppression of abrupt onsets in a manner that allows direct comparison of color singletons and abrupt onsets. As will be seen, abrupt onsets are not suppressed like color singletons, even after systematically controlling for important differences between the two types of salient stimuli.
Early theories of attentional capture
Early studies of attentional capture supported stimulus-driven accounts, which claim that certain kinds of salient distractors automatically capture attention, even when entirely task-irrelevant (Franconeri & Simons, 2003; Theeuwes, 1992; Yantis & Jonides, 1984). For example, Theeuwes (1992) used an additional singleton paradigm in which participants searched for a unique target shape (e.g., a circle) in a field of homogenous distractor shapes (e.g., diamonds) then made a speeded button-press to indicate the orientation of a line inside the target. On some trials, one of the non-target items was a color singleton that was uniquely colored from the other items. The key finding was that response times (RTs) were slowed when the singleton was present compared to when it was absent, even though participants knew the singleton was entirely task-irrelevant (a singleton-presence cost). This was taken as evidence that attention was automatically directed to the singleton distractor, which slowed detection of the target. Similarly, other studies supporting stimulus-driven accounts have suggested that eye movements are automatically directed toward salient items (Belopolsky et al., 2008; Theeuwes et al., 1998, 1999).
Goal-driven accounts, in contrast, posit that salient distractors do not capture attention unless they match the attentional control settings of the observer (Burnham, 2007; Folk et al., 1992; Lien et al., 2010). For example, if an observer is searching for a red item, other red items will capture attention; but a green color singleton – no matter how salient – will not capture attention because it mismatches the attentional set. To account for apparent capture by singletons in the paradigms mentioned above, goal-driven theorists have suggested that the aforementioned search tasks encouraged an attentional set for salience more broadly. In this paradigm, the target is typically a shape singleton in an otherwise homogenous field of distractor shapes. Thus, the target can be found by broadly searching for any type of singleton (singleton detection mode; Bacon & Egeth, 1994; Pashler, 1988), leaving observers vulnerable to capture by color singletons. As evidence of this vulnerability, when singleton detection mode is discouraged by using heterogenous shapes as distractors, color singletons fail to capture attention (feature search mode; see Fig. 1; Bacon & Egeth, 1994; Gaspelin et al., 2015, 2017; Leber & Egeth, 2006; Ruthruff et al., 2020). These findings suggest that capture is modulated by the top-down attentional control settings of the observer rather than by the salience of the stimulus.

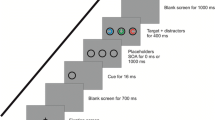
Stimuli and procedure for Experiment 1. Each search array contained a target and non-singleton distractors. On distractor-absent trials, all distractors were presented in the target color. On singleton-distractor trials, a color singleton distractor was presented at a random non-target location. On onset-distractor trials, an abrupt-onset distractor was presented at a non-target location
The signal suppression hypothesis
A potential resolution to the attentional capture debate has been proposed in the form of a hybrid model of attentional capture (for a review, see Luck et al., 2021). The signal suppression hypothesis claims that physically salient stimuli automatically generate a salience signal, but that observers can supersede this salience signal under certain circumstances (Gaspelin & Luck, 2018c; Gaspelin et al., 2019; Sawaki & Luck, 2010). This model is hybrid in that it makes two simultaneous predictions, one that stems from stimulus-driven accounts and one that stems from goal-driven accounts. First, the signal suppression hypothesis predicts that salient stimuli will capture attention if they are not suppressed, consistent with stimulus-driven accounts. However, the signal suppression hypothesis also predicts that salient stimuli can, under certain conditions, be successfully suppressed to prevent attentional capture, consistent with goal-driven accounts.
One line of support for the signal suppression hypothesis has come from studies of eye movements. For example, Gaspelin et al. (2017) had participants perform an additional singleton paradigm and used the destinations of first eye movements to infer attentional capture by the salient distractor. When the target was a shape singleton, inducing singleton detection mode, first eye movements were more likely to be directed to the singleton distractor than to the average non-singleton distractor (an oculomotor capture effect). This finding suggests that color singletons capture overt attention when they cannot be suppressed. Crucially, when the target was presented amongst distractors with heterogeneous shapes, inducing feature search mode, first eye movements were less likely to be directed to singleton distractors than to non-singleton distractors (an oculomotor suppression effect), indicating that observers actually benefited from singleton presence. This finding was taken to suggest that color singletons can be pre-emptively suppressed to prevent attentional capture (see also Gaspelin et al., 2019; Gaspelin & Luck, 2018a).
Further supporting the signal suppression hypothesis, converging evidence of color singleton suppression has been obtained under a variety of circumstances. For example, additional evidence of singleton suppression comes from demonstrations that unexpected probe letters are reported less accurately at singleton distractor locations than at non-singleton distractor locations (Chang & Egeth, 2019; Gaspelin et al., 2015). Interestingly, this suppression effect occurs even at large set sizes that maximize singleton salience (Stilwell & Gaspelin, 2021) and when multiple objects are presented in a to-be-ignored color (Lien et al., 2021; Stilwell & Vecera, 2022). Additional evidence of suppression comes from studies demonstrating that initial capture by color singletons is reduced as participants gain experience with specific features or likely locations of the color singleton, which is consistent with the notion that participants are learning to apply some suppressive process to salient items (Adam et al., 2021; Adam & Serences, 2021; Gaspelin et al., 2019; Gaspelin & Luck, 2018b; Stilwell & Vecera, 2019; Vatterott & Vecera, 2012; Wang & Theeuwes, 2018; Won et al., 2019, 2022; Won & Geng, 2020). Finally, many electrophysiological studies indicate that color singletons do not attract attention, as indexed by the N2pc component, and instead elicit electrophysiological indices of suppression (the PD component; Drisdelle & Eimer, 2021; Feldmann-Wüstefeld et al., 2020, 2021; Gaspar & McDonald, 2014; Gaspelin & Luck, 2018b; Sawaki & Luck, 2010; Stilwell et al., 2022; van Moorselaar & Slagter, 2019; but see Forschack et al., 2022; Kerzel & Burra, 20202020). In short, there has been an abundance of evidence that observers can learn to suppress salient items and that this suppression can be used to prevent attentional capture.
Can abrupt onsets also be suppressed?
Although the signal suppression hypothesis has gained considerable support from the studies cited above, they have focused almost exclusively on color singletons. This makes it unclear whether other kinds of salient stimuli, such as abrupt onsets, can be suppressed in a similar manner. If other kinds of salient stimuli cannot be suppressed, this would represent an important limitation of the signal suppression hypothesis (Gaspelin & Luck, 2018a).
There have been many previous studies of abrupt-onset capture, which makes it challenging to comprehensively review the entire literature. For example, Gaspelin et al. (2016) identified 104 published experiments on abrupt-onset capture. They found that the results were highly mixed, with approximately 75% producing evidence of capture and 25% not producing evidence of capture. The results of this literature review hint that onsets capture attention and are therefore not easily suppressed like color singletons. However, many of these studies were not designed to answer our specific question of whether abrupt onsets can be suppressed like color singletons. To address this question, a study would need to meet four basic requirements (see also Wöstmann et al., 2022).
-
1.
Direct comparison: The study must directly compare capture by abrupt onsets and color singletons within the same paradigm.
-
2.
Task-irrelevant: Salient distractors must be task-irrelevant, because participants are unlikely to suppress stimuli that are task-relevant. In particular, the search task should encourage use of feature-search mode (Bacon & Egeth, 1994).
-
3.
Antipredictive: Salient distractors should be antipredictive of the target location. If a salient distractor can appear at the target location, participants may be reluctant to suppress it because this would often result in counterproductive suppression of the target (Roque et al., 2016).
-
4.
Baseline: The study must permit an assessment of whether the salient distractor was suppressed below baseline (e.g., Gaspelin et al., 2015, 2017). This typically involves using a metric, such as eye movements or probe performance, that compares attention to a salient item with attention to non-salient (baseline) items.
To our knowledge, these requirements have never before been met by a single study. Below, however, we review some examples of studies of abrupt-onset capture from various paradigms and relate them to these requirements.
Several studies have shown that abrupt onsets produce capture effects in the spatial cuing paradigm (e.g., Burnham, 2020; Folk & Remington, 2015; Gabbay et al., 2019; Gaspelin et al., 2016; Lamy et al., 2018; Lamy & Egeth, 2003; Maxwell et al., 2021; Ruthruff et al., 2019, 2020; Zivony & Lamy, 2018) and the irrelevant feature paradigm (Franconeri & Simons, 2003; Jonides & Yantis, 1988; Yantis & Jonides, 1984). Most of the studies, however, failed to meet requirement #1 because they did not also include color singletons (Gabbay et al., 2019; Gaspelin et al., 2016; Lamy et al., 2018; Maxwell et al., 2021; Zivony & Lamy, 2018), allowing no direct comparison between onsets and color singletons. Additionally, none of these studies met requirement #3 because the salient distractor could appear at the target location, which may have discouraged suppression of the salient item.
Other studies have used the oculomotor capture paradigm to study capture by abrupt onsets. In this paradigm, participants generated saccades to uniquely colored items and an irrelevant distractor sometimes unexpectedly appeared at a non-target location (Theeuwes et al., 1999; see also Belopolsky et al., 2008). In these studies, first eye movements were frequently directed to abrupt-onset distractors (e.g., on 16–40% of trials), and this was initially taken as evidence that the abrupt onsets automatically captured attention. A shortcoming of many of these studies is that the abrupt onset may have been made task-relevant (requirement #2), by either encouraging participants to use singleton-detection mode (see Wu & Remington, 2003) or making the onset a task-relevant color (Becker et al., 2017; Becker & Lewis, 2015; Fuchs et al., 2013; Goller et al., 2016; Ludwig & Gilchrist, 2003; Weichselbaum et al., 2014). Furthermore, none of these studies directly compared processing of the salient distractor to some baseline level of processing to evaluate whether salient items were suppressed (requirement #4).
In summary, there have been many studies of abrupt-onset capture, but none have addressed the specific question of whether abrupt onsets can be suppressed under the same conditions in which color singletons are suppressed. If abrupt onsets cannot be suppressed like color singletons, this would represent an important limitation of the signal suppression hypothesis (e.g., see Luck et al., 2021).
Experiment 1: Baseline
The purpose of Experiment 1 was to determine whether abrupt onsets can be suppressed under the same conditions that have previously revealed suppression of color singletons. We set out to design a study that meets requirements 1–4 as described above. As shown in Fig. 1, participants performed an additional singleton paradigm that was adapted for eye tracking based upon Gaspelin et al. (2017). On each trial, participants searched for a specific target shape (e.g., a diamond) amongst a heterogeneous set of distractor shapes (e.g., circles, hexagons, and triangles) and reported the tilt of a line inside the target. We randomly intermixed three distractor conditions: color singleton, abrupt onset, and distractor absent. We begin by using the same abrupt onset stimuli – four white dots – that have been employed in a very large number of previous studies (e.g., Folk et al., 1992).
Importantly, our approach allows for a direct comparison of oculomotor capture between abrupt onset and color singleton distractors within the same experimental paradigm. Based upon past experiments (Gaspelin et al., 2017, 2019; Gaspelin & Luck, 2018a), we expect that first saccades to color singletons will be suppressed. The key question is whether abrupt onsets can also be suppressed or instead will capture overt attention. If onsets can be suppressed below baseline, then first saccades should be less likely to be directed to the salient abrupt-onset distractor than to the average non-salient distractor. If onsets capture attention, however, then first saccades should be more frequently directed to the salient abrupt-onset distractor than to the average non-salient distractor.
Method
Participants
An a priori sample of 32 participants was established based upon previous studies of oculomotor suppression. Assuming that the magnitude of oculomotor suppression is similar to that in previous studies (dz = 1.58; Gaspelin et al., 2017, Experiment 2), this sample size should allow over .999 power to detect an oculomotor suppression effect.
Participants were undergraduate students from State University of New York at Binghamton who participated for course credit. One participant was replaced due to a manual response accuracy 3.5 standard deviations below the group mean (i.e., less than 86%), leaving a final sample of 32 participants (22 women and 10 men, mean age = 19.0 years). All participants demonstrated normal color vision on an Ishihara color vision test and self-reported normal or corrected-to-normal visual acuity. All experiments in this study were approved for ethical considerations by an institutional review board.
Apparatus
Stimuli were presented with PsychToolbox (Brainard, 1997) for Matlab on an Asus VG245H LCD monitor at a viewing distance of 100 cm. A photosensor was used to measure the timing delay of the video system (12 ms), and this delay was subtracted from all latency values stated in this paper. An SR Research Eye Link 1000+ desk-mounted eye tracker measured monocular eye position from the right eye at 500 Hz. The Eye Link Toolbox was used to interface the stimulus presentation system and eye-tracking system (Cornelissen et al., 2002).
Stimuli and procedure
The search display contained six shapes arranged in a notional circle that was 4.5° in radius, with distance calculated between the center of the display and the center of each shape (see Fig. 1). Shapes consisted of circles (0.9° in diameter), diamonds (1.0° in diameter), hexagons (0.9° in diameter), and triangles (0.9° in base and height). The shapes were drawn in photometrically isoluminant colors: green (30.0 cd/m2, x = .30, y = .63) and red (30.0 cd/m2, x = .63, y = .33). Each shape contained a gray line (0.2° in length and 0.03° in thickness), slanted 45° to either the left or right. These lines were too small to be visible from central fixation, necessitating that the participants directly fixated the target to identify the line orientation. A gray fixation cross (30.0 cd/m2; 0.3° × 0.3° in diameter) was presented at the center of the screen in the fixation screen and search array. In onset-distractor trials, four bright white onset dots (313.0 cd/m2; 0.2° × 0.2° in diameter) were presented around a randomly chosen non-salient distractor. All stimuli were presented on a black background.
Each search display contained a target shape (e.g., a diamond) and five distractors with different shapes (e.g., circles, hexagons, and triangles). The target color (red vs. green) and target shape (circle vs. diamond) were constant for each participant’s entire experimental session and counterbalanced across participants. Distractor shapes were randomly generated as hexagons, triangles, and the unselected target shape (circle in the diamond-target condition and diamond in the circle-target condition). By holding the target shape constant for each participant and using heterogeneous distractor shapes, this experiment encouraged the use of feature search mode and decreased motivation to intentionally search for singletons (Bacon & Egeth, 1994; Gaspelin et al., 2017; Leber & Egeth, 2006). On every trial, the target location was randomly selected. On onset-distractor and singleton-distractor trials, one location was randomly selected as the salient distractor, with the exception that it was never the target location. The singleton distractor appeared in a distinct color from the other distractors, while the onset appeared as four white circles surrounding a randomly chosen distractor. Participants were instructed to ignore both onset and singleton distractors as they would never appear at the target location. Because onsets and singletons were both presented as task-irrelevant distractors, attentional capture could be directly measured by comparing the first eye movements to the salient distractors with the baseline level of first eye movements to non-salient distractors (Adams & Gaspelin, 2021; Gaspelin et al., 2017; Talcott & Gaspelin, 2020).
The experiment consisted of three trial types that were randomly interspersed over the course of the experimental session. On onset-distractor trials (one-third of trials), an abrupt-onset distractor appeared at a non-target location, consisting of dots that suddenly appeared around one item for 100 ms before suddenly disappearing. On singleton-distractor trials (one-third of trials), a color singleton distractor appeared at a non-target location, consisting of a shape drawn in a unique color from the rest of the search items. Finally, on distractor-absent trials (one-third of trials), all non-target shapes were non-salient distractors. Across all trial types, participants were tasked with locating the target shape as quickly as possible, then executing a speeded manual button-press to indicate the orientation of the line inside (left or right tilted) on a gamepad (left or right trigger buttons, respectively).
Each trial began with a blank screen for 1,000 ms. Next, a fixation cross with placeholder shapes at each of the search locations was presented (see Fig. 1). Each placeholder was a combination of all the potential shapes (diamond, circle, hexagon, triangle) and tilted lines (left vs. right) at each location. This technique was based upon Lamy and Egeth (2003) and served the purpose of eliminating onset transients in the search array (see also Franconeri & Simons, 2003; Yantis & Jonides, 1984). That is, this approach caused the appearance of the search array to consist of abruptly offsetting line segments, which should reduce any attentional set for onsets to the greatest degree possible (Yantis & Jonides, 1984). To initiate a trial, participants were required to maintain gaze position within 1.5° of the fixation cross for 500 ms. Once this criterion was met, the search array then appeared until a manual response was made, or until 2,000 ms had elapsed (the timeout period).
Each experimental block consisted of 60 trials. The first block was a practice block and this was followed by eight regular blocks (480 total regular trials). If participants took more than 2,000 ms to respond, they were presented with a 500-ms timeout display (“Too Slow”). If an incorrect response was made, a 200-Hz tone sounded for 500 ms. At the end of each block, participants were provided with feedback on mean response time (RT) and accuracy. These block breaks also warned participants whose accuracy fell below 90%.
Data analysis
Saccades were analyzed using techniques similar to those of previous studies of oculomotor capture (Adams & Gaspelin, 2021; Gaspelin et al., 2017, 2019; Gaspelin & Luck, 2018a; Leonard & Luck, 2011; Talcott & Gaspelin, 2020). Saccades were defined by a minimal eye velocity threshold of 30° per second and a minimum acceleration threshold of 9,500 °/sec2. To identify the destination of the first saccade, an annulus was defined around the search array, with an inner radius of 1.5° from fixation and an outer radius of 7.5° from fixation. The first saccade on each trial was then defined as the first eye movement landing within the annulus. The nearest search item was then selected as the first saccade destination. This effectively creates wedge-shaped interest areas around each search item (Leonard & Luck, 2011). Saccadic latency was measured as the start time of the first saccade that landed within the annulus.
The first experimental block was excluded as a practice. Trials with RTs less than 200 ms (0.0% of trials) or an RT timeout (greater than 2,000 ms; 0.3% of trials) were excluded from all analyses, as well as trials in which participants did not move their eyes from central fixation (0.2%) and trials with abnormal saccade latencies (less than 50 ms or greater than 1,000 ms, comprising 1.1% of trials). Trials with incorrect responses (3.0%) were omitted from RT analyses. In total, 4.1% of trials were excluded.
For analyses of variance (ANOVAs), Greenhouse-Geisser corrected p values are reported to avoid issues of sphericity. In analyzing saccadic destination, we avoided directly comparing salient distractors, non-salient distractors, and targets due to interdependence issues. In other words, within a given condition, an increase in first eye movements to one search item is automatically accompanied by a decrease in first eye movements to the other search items. Therefore, following Gaspelin and Luck (2018a), we used t tests to compare percentages of saccades in independent conditions. Cohen’s dz is used for within-subject t tests, whereas Cohen’s ds is used for between-subject t tests.
Results
Manual responses
If the salient item captures attention, then mean RT should be slower on trials in which the salient distractor is present than trials in which it is absent (distractor presence cost; Theeuwes, 1992). If the salient item is suppressed, however, then mean RT should be faster on distractor-present trials than distractor-absent trials (distractor presence benefit; Chang & Egeth, 2019; Gaspelin et al., 2015).
As depicted in Fig. 2a, manual RTs were slower in onset-distractor trials (915 ms) than distractor-absent trials (884 ms), whereas RTs were faster in singleton-distractor trials (873 ms) than distractor-absent trials. To formally analyze this pattern, a one-way repeated-measures ANOVA was conducted on manual RTs with the factor of salient distractor type (onset, singleton, and absent). This resulted in a main effect of salient distractor type, F(2, 62) = 34.06, p < .001, η2 = .524. Preplanned t tests indicated that manual RTs were slower in onset-distractor trials than distractor-absent trials: a 31-ms distractor presence cost, t(31) = 5.51, p < .001, dz = 0.97. Manual RTs were faster in singleton-distractor trials than distractor-absent trials: an 11-ms distractor presence benefit, t(31) = 2.66, p = .012, dz = 0.47. Manual RTs were significantly slower in onset-distractor trials than singleton-distractor trials, t(31) = 7.21, p < .001, dz = 1.28.

Results from Experiment 1. (a) Manual response times (RTs) by salient distractor type (absent, singleton, onset). (b) Percentage of first saccades to each search item by salient distractor type. (c) Oculomotor capture effects by salient distractor type (abrupt onsets vs. color singletons). In all figures, error bars indicate within-subject 95% confidence intervals (Morey, 2008)
The same one-way ANOVA was conducted on manual error rates with the within-subject factor of salient distractor type (onset, singleton, and absent). There was not a reliable main effect of salient distractor type, F(2, 62) = 0.09, p = .914, η2 = .003. Participants did not commit many errors during this task, as evidenced by an overall manual error rate of only 3.0%.
In summary, the RT results suggest that onset distractors captured attention whereas color singleton distractors were suppressed. In the following sections, we also discuss our primary dependent measure, which more directly indexes overt attentional capture versus suppression: the destination of first saccades.
First saccade destination
Salient distractor absent
Table 1 shows, for each experiment, the percentages of first eye movements to each search item for trials in which the salient distractor was absent. Although we have no a priori hypotheses about these trials, we have included them for the sake of transparency.
Salient distractor present
For trials in which a salient distractor was present, Fig. 2b depicts the percentage of first eye movements to each search item (target, non-salient distractor, salient distractor) for both distractor types (onset and singleton). The percentage of first eye movements to the non-salient distractors was divided by the number of non-salient distractors to give a per item estimate. As can be seen, first saccades were less likely to be directed to the singleton distractor than to the average non-salient distractor. However, first saccades were more likely to be directed to the onset distractor than the average non-salient distractor.
A two-way within-subjects ANOVA compared the percentage of first saccades to each search item (target, non-salient distractor, salient distractor) as a function of salient distractor type (onset vs. singleton). This analysis revealed main effects of salient distractor type, F(1, 31) = 119.68, p < .001, ηp2 = .794, and search item, F(2, 62) = 77.46, p < .001, ηp2 = .714. There was also a significant interaction between salient distractor type and search item, F(2, 62) = 114.20, p < .001, ηp2 = .786. Preplanned t tests compared the percentage of first eye movements to each search item on singleton-distractor trials versus onset-distractor trials. These analyses demonstrated a significantly greater percentage of first eye movements to the salient distractor on onset-distractor trials (30%) than on singleton-distractor trials (8%), t(31) = 12.26, p < .001, dz = 2.17. First saccades were less likely to be directed to the target on onset-distractor trials (38%) than singleton-distractor trials (46%), t(31) = 6.29, p < .001, dz = 1.11. Furthermore, first saccades were less likely to be directed to the non-salient distractor on onset-distractor trials (8%) than on singleton-distractor trials (12%), t(31) = 10.10, p < .001, dz = 1.79.
The key question is whether abrupt onsets capture overt attention or are instead suppressed. To assess this, a difference score was calculated comparing the percentage of first saccades to the salient distractor minus the percentage of first saccades to the average non-salient distractor (Adams & Gaspelin, 2021; Gaspelin et al., 2017; Gaspelin & Luck, 2018a). This oculomotor capture effect was computed for each distractor type (onset vs. singleton) and is depicted in Fig. 2c. A positive score indicates that the salient distractor attracted attention more than the typical non-salient distractor (oculomotor capture), whereas a negative score indicates that the salient distractor attracted attention less than the typical non-salient distractor (oculomotor suppression). Onset distractors produced a large positive score (22%), indicating that onsets captured overt attention, t(31) = 8.39, p < .001, ds = 1.48. Singleton distractors, however, produced a negative score (-4%), indicating that singletons were suppressed, t(31) = 3.47, p = .002, ds = 0.61. A paired-samples t test revealed that oculomotor capture effects were indeed significantly larger for onset distractors (22%) than singleton distractors (-4%), t(31) = 12.32, p < .001, dz = 2.18.
Altogether, the oculomotor results indicate that color singleton distractors were suppressed (Gaspelin et al., 2017; Gaspelin & Luck, 2018b) whereas abrupt onsets captured attention.
Learning effects: Oculomotor capture across blocks
We also investigated whether participants learned to suppress salient items across the experimental session. Previous evidence suggests that observers learn to generate fewer saccades to repeatedly presented salient distractors, for both abrupt onsets (Bonetti & Turatto, 2019; Pascucci & Turatto, 2015; Turatto et al., 2018a, b) and color singletons (Anderson & Mrkonja, 2021; De Tommaso & Turatto, 2019; Gaspelin & Luck, 2018b; Vatterott & Vecera, 2012). The details of an exploratory analysis of these learning effects are reported in the Online Supplemental Material (OSM) and are depicted in Fig. 3. As can be seen, both onset and singleton capture decreased as a function of task experience. The most important finding is that, although oculomotor capture by abrupt onsets declined across blocks, it was never eliminated.

Oculomotor capture effects as a function of block. Oculomotor capture effects gradually declined across the experimental session for both stimulus types (onsets and color singletons). However, capture by abrupt onsets was never completely eliminated. Error bars indicate between-subject standard error of the mean
Saccadic latency by first saccade destination
We had no a priori hypotheses about saccadic latency but have included them for the sake of completeness (Table 2). A more detailed exploratory analysis is reported in the OSM. To briefly summarize, saccadic latencies were generally faster when the first saccade was directed to the salient distractor than when it was directed to the target or non-salient distractor. But this pattern did not significantly differ as a function of distractor type (onset vs. singleton).
Discussion
Experiment 1 compared oculomotor capture by color singletons and abrupt onsets. Consistent with previous studies (Gaspelin et al., 2017, 2019; Gaspelin & Luck, 2018a), saccades to color singletons were suppressed (4% oculomotor suppression effect). Interestingly, abrupt onsets did not produce the same pattern of results. Instead, first eye movements were more likely to be directed to the onset distractor than the average non-salient distractor: a 22% oculomotor capture effect. This initial finding suggests that abrupt onsets cannot be suppressed like color singletons, which is broadly consistent with previous studies indicating that abrupt onsets can capture attention under circumstances where color singletons can be ignored (Franconeri & Simons, 2003; Lamy & Egeth, 2003; Ruthruff et al., 2020; Yantis & Jonides, 1984).
One question left unresolved by Experiment 1 is whether onsets were not suppressed simply because they were intermixed with color singletons on a trial-by-trial basis. For instance, participants might have difficulty suppressing more than one type of salient stimulus at a time. To explain the findings of Experiment 1 in this manner, one would have to additionally assume that participants chose to focus primarily on suppressing color singletons. Arguing against this hypothesis, several studies have demonstrated that observers can establish templates for multiple search targets (Beck et al., 2012; Grubert & Eimer, 2015; Moore & Osman, 1993) and multiple singleton colors (Chang & Egeth, 2019, 2021; Won & Geng, 2018). However, no previous study has specifically examined whether templates for onset and singleton distractors can be established at the same time. We therefore conducted a separate control experiment (N = 32) that used the same procedure as in Experiment 1, except that the salient distractor type was blocked. One half of the experiment contained abrupt onsets only and one half of the experiment contained color singletons only. There was a 19% oculomotor capture effect by abrupt onsets, which is similar in magnitude to the oculomotor capture effect produced by onsets in Experiment 1 (22%), t(62) = 0.88, p = .382, ds = 0.22. There was also a 4% oculomotor suppression effect for singleton distractors, which is similar in magnitude to the oculomotor suppression effect for singletons in Experiment 1 (4%), t(62) = 0.07, p = .943, ds = 0.02. These results indicate that abrupt onsets capture attention even when repeatedly and predictably presented without other salient distractors. Thus, there is no evidence that the failure to suppress abrupt onsets reflects a difficulty in simultaneously suppressing more than one type of salient object (see also Won & Geng, 2018).
Experiment 2: Offsets
A straightforward conclusion from Experiment 1 is that abrupt onsets are powerful and can overwhelm the suppressive processes used to prevent attentional capture by color singletons. However, in using the classic abrupt-onset stimuli (brief appearance of four bright white dots), Experiment 1 introduced several differences between the onsets and singleton distractors (e.g., offsetting vs. remaining visible, bright vs. dim, and color). It is unclear to what degree these differences account for the power of abrupt onsets. Therefore, in the following experiments (Experiments 2–4), we progressively eliminate each of these differences. As will be seen, we reveal that each difference, by itself, is important; nevertheless, abrupt onsets captured attention even after eliminating all three of the differences.
The first difference we investigate is that, in Experiment 1, the abrupt onsets also offsetted shortly thereafter (as in many previous studies; see Folk et al., 1992), whereas color singletons remained visible until response. This offset might make it especially difficult to suppress the onset stimulus. For one thing, it might be difficult to apply suppressive processes to a salient item that has disappeared. Additionally, the offset itself might be a salient transient (Miller, 1989), providing a second chance for a transient to capture attention and trigger an eye movement to that location.
Experiment 2 therefore used the same white onset dots as in Experiment 1, except without immediate offsets. If offsets contribute to onset capture, then oculomotor capture effects by abrupt onsets should decrease in Experiment 2 relative to Experiment 1. Alternatively, if offsets play no important role in capture, then onset capture effects should be similar in Experiments 1 and 2.
Method
All methods were identical to Experiment 1, except for the following changes.
A new sample of 32 participants (20 women and 12 men, mean age = 19.1 years) was collected. One participant was replaced due to abnormally low manual response accuracy (3.5 standard deviations below the group mean) and one participant was replaced due to a saccadic latency 3.5 standard deviations above the group mean (i.e., more than 452 ms). The onset distractors appeared with the surrounding search array until a response was made. In other words, onsets no longer disappeared 100 ms after the appearance of the search array, as in Experiment 1. The same trial exclusion criteria from Experiment 1 were used. We removed from analysis all trials with RTs less than 200 ms (0.0%) or no response at all (0.3% of trials), as well as trials in which participants made no eye movements from central fixation (0.7%) and trials with abnormal saccade latencies (less than 50 ms or greater than 1,000 ms; 0.8% of trials). Trials with incorrect responses (2.4%) were omitted from RT analyses. Altogether, 3.3% of trials were excluded.
Results
To summarize, the results of Experiment 2 replicated Experiment 1 by demonstrating suppression of color singletons but capture by abrupt onsets. Importantly, capture effects from onsets were reliably smaller in Experiment 2 than in Experiment 1, indicating that offsets do enhance capture by abrupt onsets.
Manual responses
Figure 4a depicts manual RTs for each salient distractor type (onset, singleton, absent). A one-way repeated-measures ANOVA was conducted on manual RTs with the factor of salient distractor type. This resulted in a main effect of salient distractor type, F(2, 62) = 19.98, p < .001, η2 = .392. Preplanned t tests indicated that manual RTs were slower in onset-distractor trials (880 ms) than distractor-absent trials (867 ms): a 13-ms distractor presence cost, t(31) = 2.36, p = .025, dz = 0.42. Manual RTs were faster in singleton-distractor trials (850 ms) than distractor-absent trials (867 ms): a 17-ms distractor presence benefit, t(31) = 4.49, p < .001, dz = 0.79. Manual RTs were significantly slower in onset-distractor trials than in singleton-distractor trials, t(31) = 6.47, p < .001, dz = 1.14.

Results from Experiment 2. (a) Manual response times (RTs) by salient distractor type (absent, singleton, onset). (b) Percentage of first saccades to each search item by salient distractor type. (c) Oculomotor capture effects by salient distractor type
We conducted the same one-way repeated-measures ANOVA on manual error rates with the factor of salient distractor type (onset, singleton, absent). There was not a reliable main effect of salient distractor type, F(2, 62) = 0.06, p = .939, η2 = .002. As in Experiment 1, manual error rates were generally low in this task, averaging only 2.4%.
First saccade destination
Figure 4b depicts the percentage of first eye movements to each search item (target, non-salient distractor, salient distractor) on onset-distractor trials and singleton-distractor trials. The basic pattern of results resembles that of Experiment 1. On singleton-distractor trials, first saccades were less likely to be directed to the salient distractor than the average non-salient distractor. On onset-distractor trials, however, first saccades were more likely to be directed to the onset distractor than the average non-salient distractor.
A two-way within-subjects ANOVA compared the percentage of first saccades as a function of salient distractor type (onset, singleton) and search item (target, non-salient distractor, salient distractor). This analysis revealed a main effect of salient distractor type, F(1, 31) = 88.45, p < .001, ηp2 = .740, and a main effect of search item, F(2, 62) = 78.31, p < .001, ηp2 = .716. There was also a significant interaction between salient distractor type and search item, F(2, 62) = 59.51, p < .001, ηp2 = .658. Preplanned t tests compared the percentage of first eye movements to each search item on singleton-distractor trials and onset-distractor trials. These analyses demonstrated a greater percentage of first eye movements to the salient distractor on onset-distractor trials (23%) than singleton-distractor trials (6%), t(31) = 9.18, p < .001, dz = 1.62. Further indicating capture by the onset distractor, first saccades were less likely to be directed to the target on onset-distractor trials (41%) than singleton-distractor trials (47%), t(31) = 4.11, p < .001, dz = 0.73. First saccades were less likely to be directed to the non-salient item on onset-distractor trials (9%) than singleton-distractor trials (12%), t(31) = 9.08, p < .001, dz = 1.61.
The key question is whether onsets captured attention or were instead suppressed. As depicted in Fig. 4c, oculomotor capture effects were calculated for each salient distractor type (singleton and onset). A paired-samples t test indicated that oculomotor capture effects were indeed larger for onset distractors (14%) than singleton distractors (-6%), t(31) = 9.58, p < .001, dz = 1.69. A one-sample t test confirmed that the 14% oculomotor capture effect by onsets was significantly greater than zero, t(31) = 5.81, p < .001, ds = 1.03, and the 6% oculomotor suppression effect for singletons was significantly below zero, t(31) = 7.07, p < .001, ds = 1.25. These results clearly indicate that color singleton distractors were suppressed, but that onset distractors were not suppressed.
Experiment 1 versus Experiment 2
Experiment 1 and 2 used identical methods, except that Experiment 2 removed the offsets from onset distractors. We therefore compared the magnitude of onset-capture effects and singleton-suppression effects between the two experiments using independent-samples t tests. Onset-capture effects were indeed significantly larger in Experiment 1 (22%) than Experiment 2 (14%), t(62) = 2.05, p = .044, ds = 0.51. Meanwhile, singleton-suppression effects were similar in Experiment 1 (4%) and Experiment 2 (6%), t(62) = 1.44, p = .154, ds = 0.36. These results suggest that immediate offsets do play an important role in the magnitude of capture by abrupt onsets.
Learning effects: Oculomotor capture across blocks
As in Experiment 1, we assessed whether oculomotor capture by onset and singleton distractors decreased as a function of task experience with an exploratory analysis in the OSM. Onset- and singleton-capture effects again decreased as a function of task experience (Fig. 3). These results indicate that oculomotor capture by salient distractors does decrease with task experience, replicating the results of the first experiment and previous studies (Pascucci & Turatto, 2015; Turatto et al., 2018a, b; Turatto & Pascucci, 2016). However, onset capture was never completely eliminated, suggesting that onsets could not be suppressed regardless of task experience.
Saccadic latency by first saccade destination
We had no key predictions about saccadic latencies in relation to onset capture. But, for the sake of completeness, they are included in Table 2 and a detailed analysis is reported in the OSM. To summarize here, saccadic latencies were generally slower when the first saccade was directed to the target than the salient distractor or non-salient distractor (Donk & van Zoest, 2008; Gaspelin et al., 2017; van Zoest et al., 2004). However, this pattern was very similar for onsets and singletons.
Discussion
Experiment 2 controlled one major difference between the onsets and singletons from Experiment 1: offsets. Instead of presenting onsets for only 100 ms before offsetting, as in Experiment 1 and many prior demonstrations of onset capture (Folk et al., 1992; Gabbay et al., 2019; Gaspelin et al., 2016; Lamy et al., 2018), onsets remained present until response. Color singletons were suppressed, just as in Experiment 1. Importantly, onsets still captured attention, albeit significantly less strongly than in Experiment 1 (22% in Exp. 1 vs. 14% in Exp. 2). These results demonstrate that the offset transients do add to onset capture, but cannot fully explain why onsets are not suppressed and instead capture attention.
Experiment 3: Luminance
Experiment 3 addressed a further difference between the onset and singleton distractor: luminance. Traditionally, demonstrations of onset capture have used abrupt onsets with much higher luminance than other display elements (Folk et al., 1992; Gaspelin et al., 2016; Lamy et al., 2018). This luminance increase might greatly increase the power of the abrupt onset transient by virtue of higher contrast with respect to the background. It is therefore important to assess whether abrupt onsets can capture attention even when they do not have higher luminance than other display elements. This was the goal of Experiment 3.
Experiment 3 retained the key change introduced in Experiment 2 – presenting the abrupt onsets without a subsequent abrupt offset – while also controlling for luminance. Specifically, Experiment 3 replicated the methods of Experiment 2 except with gray onsets that were photometrically isoluminant with the color singletons, instead of the highly luminant white onsets from Experiments 1 and 2. The main question is whether onset capture will still remain. If capture by onsets in Experiments 1 and 2 was due to high luminance, it should be eliminated in the current experiment. If capture by onsets in Experiments 1 and 2 was due primarily to the abruptness of the onset, then it should remain in the current experiment.
Method
All procedures were identical to Experiment 2, except for the following changes.
First, a new sample of 32 participants was collected (16 women and 16 men with a mean age of 20.4 years). Second, the onset stimuli were gray dots that were photometrically isoluminant (30.0 cd/m2, x = .54, y = .36) with the other display items.
The same trial exclusion criteria from Experiment 1 were again used here. We removed trials with RTs less than 200 ms (0.0% of trials) or an RT timeout (1.0% of trials) from all analyses, as well as trials in which participants made no eye movements from central fixation (0.5%) and trials with abnormal saccade latencies (< 50 ms or > 1,000 ms; 1.5% of trials). Trials with incorrect responses (3.4%) were omitted from RT analyses. Altogether, 4.9% of trials were excluded.
Results
To briefly summarize, Experiment 3 replicated Experiment 2 by demonstrating onset capture. However, onset capture effects were reliably lower in Experiment 3 than in Experiment 2, indicating that high luminance enhances onset capture.
Manual responses
As depicted in Fig. 5a, manual RTs were much slower in onset-distractor trials (962 ms) than singleton-distractor trials (937 ms) and distractor-absent trials (955 ms). A one-way repeated-measures ANOVA was conducted on manual RT with the factor of salient distractor type (onset, singleton, and absent). This produced a main effect of salient distractor type, F(2, 62) = 11.14, p < .001, η2 = .264. Preplanned t tests indicated that manual RTs were not significantly slower on onset-distractor trials than distractor-absent trials, t(31) = 1.27, p = .213, dz = 0.23. Manual RTs on singleton-distractor trials were faster than manual RTs on distractor-absent trials: an 18-ms singleton presence benefit, t(31) = 3.25, p = .003, dz = 0.57. Manual RTs were significantly slower on onset-distractor trials than singleton-distractor trials, t(31) = 4.65, p < .001, dz = 0.82.

Results from Experiment 3. (a) Manual response times (RTs) by salient distractor type (absent, singleton, onset). (b) Percentage of first saccades to each search item by salient distractor type. (c) Oculomotor capture effects by salient distractor type
A one-way repeated-measures ANOVA was conducted on manual error rates with the factor of salient distractor type (onset, singleton, absent). There was not a reliable main effect of salient distractor type, F(2, 62) = 0.23, p = .794, η2 = .007. Manual error rates were similar regardless of salient distractor type (averaging only 3.4%).
Overall, these patterns replicate Experiments 1 and 2 and are consistent with the interpretation that attention was captured by the onset.
First saccade destination
As in Experiments 1 and 2, we examined the percentage of first eye movements to each search item (target, non-salient distractor, salient distractor) on onset distractor and singleton-distractor trials (Fig. 5b). As can be seen, first saccades were less likely to be directed to the singleton distractor than non-salient distractors. In contrast, first saccades were more likely to be directed to the onset distractor than non-salient distractors.
We first compared the percentage of first saccades to each search item. A two-way within-subjects ANOVA was conducted on the percentage of first saccades as a function of salient distractor type (onset, singleton) and search item (target, non-salient distractor, salient distractor). This analysis revealed a main effect of salient distractor type, F(2, 62) = 23.04, p < .001, ηp2 = .426, a main effect of search item, F(2, 62) = 49.24, p < .001, ηp2 = .614, and a significant interaction between salient distractor type and search item, F(2, 62) = 30.59, p < .001, ηp2 = .497. Preplanned t tests compared the percentage of first eye movements to each search item on singleton-distractor trials and onset-distractor trials. This demonstrated a greater proportion of first eye movements to the salient distractor on onset-distractor trials (18%) than singleton-distractor trials (8%), t(31) = 6.04, p < .001, dz = 1.07. First saccades were less likely to be directed to the target on onset-distractor trials (40%) than singleton-distractor trials (43%), t(31) = 3.40, p = .002, dz = 0.60, and distractor-absent trials (46%), t(31) = 4.52, p < .001, dz = 0.80. First saccades were also less likely to be directed to the non-salient item on onset-distractor trials (10%) than singleton-distractor trials (12%), t(31) = 5.07, p < .001, dz = 0.90, and distractor-absent trials (11%), t(31) = 2.33, p = .027, dz = 0.41. In contrast, first saccades were more likely to be directed to the non-salient item on singleton-distractor trials than distractor-absent trials, t(31) = 3.62, p = .001, dz = 0.64.
Oculomotor capture effects for each salient distractor type (singleton and onset) are depicted in Fig. 5c. As can be seen, oculomotor capture effects were clearly larger on onset-distractor trials than singleton-distractor trials. Preplanned t tests compared oculomotor capture effects for each salient distractor type. Oculomotor capture effects were larger on onset-distractor trials (8%) than singleton-distractor trials (-4%), t(31) = 6.10, p < .001, dz = 1.08. Separate one-sample t tests analyzed whether capture effects were significantly different from zero, which would indicate capture if the difference was positive or suppression if the difference was negative. Onset-distractor trials produced an 8% oculomotor capture effect, t(31) = 4.41, p < .001, ds = 0.78. Singleton-distractor trials produced the opposite pattern: a 4% oculomotor suppression effect, t(31) = 2.13, p = .041, ds = 0.38. These results further confirm that onset distractors captured attention, whereas singleton distractors were suppressed, replicating the basic pattern of results from Experiments 1 and 2.
Experiment 2 versus Experiment 3
Experiments 2 and 3 used identical methods, except that the onset was photometrically isoluminant with other display items in Experiment 3. Thus, we again conducted an exploratory analysis comparing the magnitude of onset-capture effects and singleton-suppression effects between the two experiments using independent-samples t tests. Crucially, this comparison confirmed that onset-capture effects were indeed significantly larger in Experiment 2 (14%) than Experiment 3 (8%), t(62) = 2.22, p = .030, ds = 0.56. Meanwhile, singleton-suppression effects were similar in Experiment 2 (6%) and Experiment 3 (4%), t(62) = 1.16, p = .252, ds = 0.29. These results suggest that high luminance plays an important role in onset capture.
Learning effects: Oculomotor capture across blocks
We evaluated block-by-block decreases in oculomotor capture by onset and singleton distractors with an exploratory analysis reported in the OSM. Singleton distractors captured attention more weakly as participants gained more experience with the task (Fig. 3). However, abrupt-onset capture was never completely eliminated.
Saccadic latency by first saccade destination
Table 2 depicts saccadic latency by destination (target, non-salient distractor, salient distractor) for each salient distractor type (onset, singleton). We conducted a detailed analysis of saccadic latencies in the OSM. To summarize here, saccadic latencies were generally slower when the first saccade was directed to the target than to the singleton distractor or non-singleton distractor (Donk & van Zoest, 2008; Gaspelin et al., 2017; van Zoest et al., 2004). However, this pattern of results did not vary as a function of salient distractor type.
Discussion
The key change in Experiment 3 was that we matched onset and singleton distractors for luminance with the other search items. Instead of using the traditional bright white abrupt onsets (which are also luminance singletons), we presented gray abrupt onsets that were photometrically isoluminant with other search items. This change reduced the amount of capture relative to Experiment 2, confirming that high luminance contributes to the power of abrupt onsets. That being said, onsets still captured attention under the same luminance levels that allowed color singletons to be suppressed.
Experiment 4: Color
Experiments 1–3 demonstrated capture by abrupt-onset distractors under the same circumstances in which color singletons were suppressed. However, the onsets in these experiments were always white or gray, as in most previous studies of abrupt-onset capture (Folk et al., 1992, 1994; Folk & Remington, 1998; Gaspelin et al., 2016; Lamy et al., 2018; Ruthruff et al., 2019; Zivony & Lamy, 2018). Experiment 4 was designed to determine whether this difference in color is sufficient to explain why abrupt onsets captured attention while color singletons were suppressed.
As depicted in Fig. 6a, the abrupt-onset distractor is now drawn in the same color as the singleton distractor. This manipulation allows us to assess whether color affects capture by abrupt-onset distractors. Note that we also retained the changes introduced in Experiment 2 (presenting abrupt onsets without offsets) and Experiment 3 (presenting onsets with the same luminance as the color singletons).

Stimuli and results from Experiment 4. Panel A depicts salient distractor types (absent, onset, singleton-plus-onset, singleton). Panel B depicts manual response times (RTs) by salient distractor type. Panel C depicts percent of first saccades to each search item by salient distractor type. Panel C depicts oculomotor capture effects by salient distractor type
Experiment 4 also added a new condition to address an ancillary question regarding suppression: Given that color singletons are suppressed so successfully, what would happen if the abrupt onset was itself part of a color singleton? To answer this question, we combined the singleton and abrupt onset at the same location (see Fig. 6a), pitting onset capture against color singleton suppression. If singleton suppression can override onset capture, we should see suppression of this singleton-plus-onset. If, however, singleton suppression cannot override onset capture, we should see capture by the singleton-plus-onset.
Method
All procedures were identical to Experiment 3, except for the following changes.
First, we collected a new sample of 32 participants (22 women and 10 men, mean age = 19.0 years). One participant was replaced due to low manual response accuracy (3.5 standard deviations below the group mean; less than 86%), one participant was replaced for making too few eye movements (i.e., less than 75% of trials), and one participant was replaced due to a slow average saccadic latency (2.5 standard deviations above the group mean; more than 455 ms).
Second, there were four distractor types: singleton, onset, singleton-plus-onset, and absent (Fig. 6a). Singleton distractor and distractor-absent trials were identical to Experiments 1–3. On onset-distractor trials, onsets were drawn in the same color as the singleton distractor to allow for a direct comparison with the singleton distractor condition. In addition, we included a new singleton-plus-onset condition, in which the salient location contained an onset (drawn in the singleton color) and the search item within the onset cue was also singleton colored. This allowed us to assess whether singleton suppression was sufficient to override attentional capture by a task-irrelevant abrupt onset. Each trial type was equally probable and selected at random.
The same trial exclusion criteria from Experiment 1 were again used here. We removed trials with RTs less than 200 ms (0.0% of trials) or an RT timeout (0.4% of trials) from all analyses, as well as trials in which participants made no eye movements from central fixation (0.2%) and trials with abnormal saccade latencies (< 50 ms or > 1,000 ms; 1.4% of trials). Trials with incorrect responses (3.0%) were omitted from RT analyses. Altogether, 4.3% of trials were excluded.
Results
To briefly summarize, Experiment 4 demonstrated capture by the onset distractor, whereas the singleton-plus-onset distractor was suppressed. This indicates that singletons can be suppressed while onsets cannot.
Manual responses
As depicted in Fig. 6b, manual RTs were slower on distractor-absent trials (912 ms) than onset-distractor trials (908 ms), singleton-distractor trials (887 ms), and singleton-plus-onset-distractor trials (892 ms). A one-way repeated-measures ANOVA was conducted on manual RTs with the factor of salient distractor type (onset, singleton, singleton-plus-onset, absent). This resulted in a main effect of salient distractor type, F(3, 93) = 10.31, p < .001, η2 = .250. Preplanned t tests indicated that manual RTs were faster in singleton-distractor trials than distractor-absent trials: a 25-ms distractor presence benefit, t(31) = 3.92, p < .001, dz = 0.69. Interestingly, manual RTs were not significantly slower on onset-distractor trials than on distractor-absent trials, t(31) = 0.76, p = .455, dz = 0.13.
An additional question in Experiment 4 was whether singleton suppression could override onset capture. Preplanned t tests confirmed that manual RTs were indeed faster in singleton-plus-onset-distractor trials than distractor-absent trials: a 20-ms distractor presence benefit, t(31) = 3.49, p = .001, dz = 0.62. Furthermore, manual RTs on singleton-plus-onset-distractor trials were reliably faster than on onset-distractor trials, t(31) = 3.68, p < .001, dz = 0.65. These patterns indicate that onset distractors at singleton locations did not induce the typical RT costs associated with attentional capture, indicating that singleton suppression superseded onset capture.
A one-way repeated-measures ANOVA was conducted on manual error rates with the factor of salient distractor type (onset, singleton, absent). There was not a reliable main effect of salient distractor type, F(2, 62) = 0.10, p = .959, η2 = .003. Manual error rates were similar regardless of salient distractor type (averaging only 3.0%).
First saccade destination
Figure 6c depicts the percentage of first eye movements to each search item (target, non-salient distractor, salient distractor) in onset distractor, singleton distractor, and singleton-plus-onset-distractor trials. The key result pertains to the singleton-plus-onset-distractor trials: first saccades were less likely to be directed to the singleton-plus-onset distractor than the average non-salient distractor. This pattern of results indicates that singleton-plus-onset distractors were suppressed below baseline levels, similar to the singleton distractor.
We first compared the percentage of first saccades to each search item. A two-way within-subjects ANOVA was conducted on the percentage of first saccades as a function of salient distractor type (onset, singleton, singleton-plus-onset) and search item (target, non-salient distractor, salient distractor). This analysis revealed a main effect of salient distractor type, F(2, 62) = 16.57, p < .001, ηp2 = .348, and a main effect of search item, F(2, 62) = 61.63, p < .001, ηp2 = .665. There was also a significant interaction between salient distractor type and search item, F(2, 62) = 19.37, p < .001, ηp2 = .385. Preplanned t tests compared the percentage of first eye movements to each search item on singleton distractor, onset distractor, and singleton-plus-onset-distractor trials. This indicated a greater percentage of first eye movements to the salient distractor on onset-distractor trials (15%) than singleton-plus-onset-distractor trials (7%), t(31) = 6.14, p < .001, dz = 1.09, and singleton-distractor trials (6%), t(31) = 5.95, p < .001, dz = 1.05. In contrast, there was a similar percentage of first eye movements to the salient item on singleton-plus-onset-distractor trials and singleton-distractor trials, t(31) = 1.41, p = .170, dz = 0.25.
The key question in this experiment was whether the onset distractor would continue to capture attention or would instead be suppressed. To answer this question, we calculated oculomotor capture effects for each salient distractor type (onset, singleton, singleton-plus-onset). As depicted in Fig. 6d, the onset distractors produced a clear capture effect. A one-way within-subjects ANOVA was used to compare oculomotor capture effects for each salient distractor type (onset, singleton, singleton-plus-onset). There was a main effect of salient distractor type, F(2, 62) = 34.65, p < .001, η2 = .528. Preplanned t tests confirmed that oculomotor capture effects were larger on onset-distractor trials (4%) than singleton-plus-onset-distractor trials (-6%), t(31) = 6.51, p < .001, dz = 1.15, and singleton-distractor trials (-7%), t(31) = 6.04, p < .001, dz = 1.07. Oculomotor capture effects were similar on singleton-plus-onset-distractor trials and singleton-distractor trials, t(31) = 1.04, p = .305, dz = 0.18. Separate one-sample t tests were conducted in each condition to analyze whether capture and suppression effects were significantly different from zero percent. Onset distractor trials produced a 4% oculomotor capture effect, t(31) = 2.24, p = .033, ds = 0.40, whereas singleton-distractor trials produced a 7% oculomotor suppression effect, t(31) = 7.99, p < .001, ds = 1.41. Singleton-plus-onset-distractor trials also produced a 6% oculomotor suppression effect, t(31) = 5.15, p < .001, ds = 0.91.
Across-experiment comparison of onset capture
Experiments 1–4 used identical methods to assess several key factors in onset capture: offsets, luminance, and color. To illustrate the cumulative impact of these factors, Fig. 7 compares oculomotor capture effects from abrupt onsets across Experiments 1–4, as each difference between onsets and singletons was removed. An exploratory one-way ANOVA with the between-subject factor of experiment (Exps. 1–4) yielded a significant main effect, F(3, 124) = 14.01, p < .001, η2 = .253. Between-subject t tests then compared oculomotor capture effects between successive experiments. Onset capture effects were significantly larger in Experiment 1 (22%) than Experiment 2 (14%), t(62) = 2.05, p = .044, ds = 0.51, and larger in Experiment 2 than Experiment 3 (8%), t(62) = 2.22, p = .030, ds = 0.56. Onset capture effects were numerically, but not significantly, larger in Experiment 3 than Experiment 4 (4%), t(62) = 1.79, p = .078, ds = 0.45. Taken together, these results indicate that rapid offsets, high luminance, and onset color all contribute to the magnitude of onset capture. Additionally, the cumulative decrease between Experiment 1 (22%) and Experiment 4 (4%) was very large, t(62) = 6.05, p < .001, ds = 1.51. Ultimately, the results suggest that onset capture is highly sensitive to several presentation factors (luminance, number of transients, color) and this will be important to consider in future studies. It is important to highlight, however, that onset-capture effects were never completely eliminated, even when onsets were closely matched to color singletons.

Oculomotor capture effects (i.e., the difference score between the percentage of first eye movements to non-salient and salient distractors) by abrupt onsets in Experiments 1–4. As can be seen, the oculomotor capture effects gradually decreased as a result of controlling for offsets (Exp. 2), luminance (Exp. 3), and color (Exp. 4, onset condition). Error bars indicate between-subject standard error of the mean
Learning effects: Oculomotor capture across blocks
As in the previous experiments, we compared onset and singleton capture with an exploratory analysis reported in the online supplemental material. As depicted in Fig. 3, onset and singleton-plus-onset capture decreased as a function of task experience. However, onset capture never decreased below baseline levels, indicating that observers were unable to suppress capture by onsets even when they were matched in color to the color singletons.
Saccadic latency by first saccade destination
Table 2 depicts saccadic latency by destination (target, non-salient distractor, salient distractor) for each distractor type (onset, singleton, singleton-plus-onset). One participant was excluded from this analysis for never fixating the singleton distractor, yielding a missing value for the analysis. We had no key predictions about saccadic latencies, but analyzed them for the sake of completeness in the OSM. To summarize here, saccadic latencies were generally slower when the first saccade was directed to the target than to the salient distractor or non-salient distractor (Donk & van Zoest, 2008; Gaspelin et al., 2017; van Zoest et al., 2004). However, this pattern did not differ as a function of salient distractor type.
Discussion
Experiment 4 investigated whether an abrupt onset would still capture attention even when presented in the same color as the color singleton. The results confirmed that, despite eliminating this final difference, abrupt onsets still attracted more eye movements than did non-salient distractors. This occurred despite the fact that participants were successful at suppressing that same color when it formed a color singleton.
Interestingly, we succeeded at eliminating the power of an abrupt onset to capture attention only by presenting it as part of the suppressed color singleton (see the singleton-plus-onset condition in Fig. 6a). Thus, the strong suppression of a color singleton can overwhelm the tendency of an onset to capture attention.
General discussion
There has been much debate as to whether salient items can automatically capture attention. A potential resolution to this debate is the signal suppression hypothesis, which proposes that observers can learn to suppress salient distractors to prevent attentional capture (Gaspelin & Luck, 2018c). The signal suppression hypothesis has garnered much support from studies of color singletons (Adam et al., 2021; Adam & Serences, 2021; Chang & Egeth, 2019; Drisdelle & Eimer, 2021; Feldmann-Wüstefeld et al., 2020; Gaspar & McDonald, 2014; Gaspelin et al., 2015; Gaspelin & Luck, 2018a, b; Sawaki & Luck, 2010; Stilwell & Gaspelin, 2021; Stilwell & Vecera, 2019; van Moorselaar & Slagter, 2019; Vatterott & Vecera, 2012; Wang & Theeuwes, 2018; Won et al., 2019, 2022; Won & Geng, 2020). However, it is currently unknown whether other kinds of salient stimuli, such as abrupt onsets, can also be suppressed below baseline levels.
Some previous studies suggest that abrupt onsets cannot be suppressed to the same degree as color singletons (e.g., Franconeri & Simons, 2003; Jonides & Yantis, 1988; Ruthruff et al., 2020; Schreij et al., 2008, 2010a, b; Theeuwes et al., 1998). But, with respect to the present research question, these studies have some important shortcomings. Namely, many did not assess attentional suppression of abrupt onsets using the paradigms that have been optimized to enable and detect singleton suppression. It is therefore unclear whether onsets captured attention due to their bottom-up salience, or to some other aspect of the task design. For example, these studies may have inadvertently boosted capture either by allowing salient items to appear at the target location (Jonides & Yantis, 1988; Lamy et al., 2018; Schreij et al., 2008; Yantis & Jonides, 1984) or by encouraging participants to search broadly for salient objects via singleton detection mode (Belopolsky et al., 2008; Irwin et al., 2000; Theeuwes et al., 1998). Additionally, many of these studies did not control for unnecessary differences between singletons and onsetting stimuli. For example, unlike color singletons, onsets have typically been presented with at least some of the following features: rapid offsets after initial onset, higher luminance than other items in the search array, and lack of chromaticity (i.e., gray or white color).
The current study therefore sought to compare suppression of abrupt onsets and color singletons in a paradigm that has previously produced suppression of color singletons (Gaspelin et al., 2017, 2019). In Experiment 1, participants searched for a target shape amongst heterogenous distractor shapes, promoting feature search mode. Three types of trials were randomly intermixed: singleton distractor, onset distractor, and distractor absent. First saccades were used to classify whether the salient distractor captured overt attention or was instead suppressed below baseline levels. The results indicated that saccades to color singleton distractors were suppressed below baseline, replicating previous studies (Gaspelin et al., 2017, 2019; Gaspelin & Luck, 2018a). In sharp contrast, saccades to abrupt-onset distractors were strongly enhanced (a 22% oculomotor capture effect for onsets, as opposed to a 4% oculomotor suppression effect for singletons). This suggests that abrupt onsets could not be suppressed like color singletons.
Experiments 2–4 progressively eliminated important differences between abrupt onsets and color singletons to determine what factors contribute to onset capture. Experiment 2 replicated Experiment 1, except that onsets no longer disappeared after 100 ms to make the time-course identical to that of color singletons. Onset-capture effects were reduced in Experiment 2 (no offset) compared to Experiment 1 (offset), indicating that immediate offsets independently enhanced the magnitude of onset-capture effects. This may be because immediate offsets offer a second transient with the potential to trigger attentional shifts (Miller, 1989) or because it is difficult to suppress salient signals that quickly disappear before inhibitory processes can be enabled. In any case, the key finding is that abrupt onsets continued to capture attention whereas color singletons were suppressed.
Experiment 3 also equated singletons and onsets for luminance by using gray onsets that were photometrically isoluminant with the surrounding search items. Abrupt onsets again produced oculomotor capture effects, but these capture effects were smaller than those produced by bright abrupt onsets in Experiments 1 and 2. This finding suggests that bright onsets (which are frequently employed in studies of onsets; Folk et al., 1992; Gaspelin et al., 2016; Lamy et al., 2018; Schreij et al., 2008; Zivony & Lamy, 2018) have substantially more power than relatively dim onsets.
Experiment 4 assessed whether abrupt onsets would capture attention even when presented in the exact same color that is suppressed when presented as a color singleton (see Fig. 6a). Crucially, the abrupt onsets continued to capture attention. These results indicate that abrupt onsets cannot be suppressed like color singletons, even when matched for color and luminance. We also investigated what would happen when we pitted onset capture against singleton suppression, by placing the abrupt-onset dots around a color singleton shape in the exact same color (see the singleton-plus-onset condition in Fig. 6a), so that the compound object was itself a color singleton. These singleton-plus-onset distractors were suppressed below baseline levels, roughly to the same degree as the color singleton without the abrupt onset. Thus, color singleton suppression can, at least under some conditions, override abrupt-onset capture.
Taken together, our results indicate that abrupt onsets are not suppressed like color singletons. In all experiments, onsets captured attention, whereas singletons were suppressed. This pattern fits with previous claims that onsets are more likely to capture attention than color singletons and other salient stimuli (Franconeri & Simons, 2003; Jonides & Yantis, 1988; Ruthruff et al., 2020). Here, we have confirmed that this finding holds even when (1) onsets and color singletons are well matched, and (2) they are presented in a paradigm conducive to strong suppression.
The magnitude of onset-capture effects and contributing factors
Although abrupt-onset capture was not suppressed below baseline levels, we demonstrated that it was sensitive to several factors: rapid offsets, high luminance, and color (Fig. 7; see Across-experiment comparison of onset capture). Despite not being strictly required by the traditional definition of abrupt onsets, most or all of these elements have been used in classic demonstrations of onset capture (Belopolsky et al., 2008; Folk et al., 1992; Folk & Remington, 1998; Franconeri et al., 2005; Franconeri & Simons, 2003; Gabbay et al., 2019; Gaspelin et al., 2016; Jonides & Yantis, 1988; Lamy et al., 2018; Lamy & Egeth, 2003; Maxwell et al., 2021; Mounts, 2000; Ruthruff et al., 2019, 2020; Theeuwes et al., 1998, 1999; Yantis & Jonides, 1984; Zivony & Lamy, 2018). These factors might explain why some studies find large capture effects from abrupt onsets and others do not (Becker et al., 2017; Becker & Lewis, 2015; Wu & Remington, 2003). For example, a stronger transient (high contrast, with both an onset and an offset) may increase the attentional priority of the abrupt onsets, enhancing their ability to compete with the target stimulus for attention (Gabbay et al., 2019; Lamy et al., 2018). The current study found support for this possibility by demonstrating a cumulative decrease in oculomotor capture as each difference between onsets and singletons was removed.
Learning to ignore across the experimental session
All of the present experiments showed evidence of learning effects across the session. Oculomotor capture effects from abrupt onsets tended to decrease in magnitude as participants gained experience with the task (see OSM for details). This decrease was particularly pronounced in Experiments 1 and 2, which yielded the strongest capture effects by abrupt onsets (e.g., from 32% to 20% in Experiment 1). A similar decrease was also observed for color singletons, which became more strongly suppressed across the experimental session. Altogether, these results fit well with previous claims that participants can gradually learn to ignore salient stimuli based upon their feature values and/or their overall presence in search displays (Anderson & Mrkonja, 2021; Bonetti & Turatto, 2019; De Tommaso & Turatto, 2019; Gaspelin & Luck, 2018b; Pascucci & Turatto, 2015; Stilwell & Vecera, 2019; Turatto et al., 2018a, b; Turatto & Pascucci, 2016; Vatterott & Vecera, 2012; Won et al., 2019; Won & Geng, 2020). For example, Turatto et al. (2018a) demonstrated that abrupt onsets capture attention less across the experimental session and this learning effect remained even in follow-up assessments several days later. In fact, some recent evidence from Won and Geng (2020) suggests that passive viewing of displays with salient distractors may be sufficient to reduce capture by salient items. Interestingly, although the data suggest that participants were learning to ignore onsets across the session in the current study, capture by abrupt onsets was never fully eliminated.
Future directions
Can abrupt onsets ever be suppressed? The current study found that abrupt onsets were not suppressed, despite matching the properties of onsets and color singletons. It should also be highlighted that the task was specifically designed to discourage capture by abrupt onsets. For example, the target was made non-salient to reduce the likelihood of participants using singleton detection mode (Bacon & Egeth, 1994). Similarly, premasks were used to make the search items offset and to reduce the likelihood of a display-wide attentional set for suddenly appearing stimuli (Burnham, 2007). We also used abrupt onsets that never appeared at the target location to eliminate any incentive to attend abrupt onsets.
That being said, there is always the possibility that onsets can be suppressed under conditions we did not study. There might be something about our task that encouraged an attentional set for abrupt onsets. For example, Schönhammer and Kerzel (2018) found that white onset cues could be ignored when the search display contained only the target and a single white distractor, so that the distractors more closely resembled the cue display, thereby encouraging participants to suppress the white distractor (though see also Experiment 4 of Ruthruff et al., 2019, which reported a contrasting result using a more difficult visual search). It is also possible that mixing abrupt onsets and color singletons may have made them more difficult to suppress. However, it should be noted that an early control experiment (described in the Discussion section of Experiment 1) argued against this possibility. In sum, although we were unable to achieve suppression of abrupt onsets even under seemingly favorable conditions, there is still a need for future research looking for the boundary conditions under which abrupt onsets can be suppressed.
There is also the question of how the observed oculomotor capture effect relates to covert attentional processes. The current study showed that overt eye movements were captured by abrupt onsets. However, some previous studies have prohibited eye movements during search and have found no evidence of covert attentional shifts to onsets via event-related potentials (ERPs) such as the N2pc component (Goller et al., 2020; Lien et al., 2008). This seems to indicate that covert attention may not always be captured by abrupt onsets. However, a major challenge to studying abrupt onsets with ERPs is that abrupt onsets are, by definition, an imbalance in stimulus energy. The sudden appearance of a lone stimulus in a given hemifield will cause large sensory-level imbalances between the contralateral and ipsilateral ERP waveforms and these imbalances will make any N2pc-like component (or lack thereof) difficult to interpret (Luck, 2012). It should be highlighted that such issues are not a problem with studies of overt eye movements, which is why we chose them as a dependent measure in the current study. Future studies are needed to develop methods to study ERPs from abrupt onsets that circumvent the energy imbalance problem.
Abrupt onsets are a dynamic stimulus that changes over time, whereas color singletons are static throughout the display period. Therefore, another important question is whether other types of dynamic salient stimuli can capture attention or can be suppressed below baseline. Several studies have suggested that moving stimuli (or at least the onset of moving stimuli) automatically capture attention (Abrams & Christ, 2003; Al-Aidroos et al., 2010). For example, according to the behavioral urgency hypothesis (Franconeri & Simons, 2003), certain types of motion capture attention more frequently than other salient stimuli because they have higher behavioral relevance. However, there are also studies indicating that certain kinds of dynamic motion can be ignored (Folk et al., 1992). This will be an important question for future studies of attentional capture.
Finally, it is worth clarifying the mechanism of suppression. Original formulations of the signal suppression hypothesis proposed that a generalized saliency signal was suppressed and that this suppression occurred independently of feature values (Sawaki & Luck, 2010). But subsequent studies demonstrated that color singletons are likely suppressed based upon their specific color value, (e.g., red), rather than a global saliency signal (Gaspelin & Luck, 2018b; see also Chang & Egeth, 2019; Cunningham & Egeth, 2016; Vatterott & Vecera, 2012). In addition, multiple studies have also demonstrated that participants can suppress search items even when they are non-salient (Lien et al., 2021; Stilwell & Vecera, 2022). In light of these findings, the signal suppression hypothesis has been revised to propose that suppression of color singletons likely occurs due to learned suppression of simple features like color or luminance (Luck et al., 2021). It remains to be seen whether participants can learn to suppress a saliency signal without knowing the specific feature value (see Vatterott et al., 2018; Won et al., 2019), and this issue deserves further exploration in future research. Another important question is whether the suppression is a direct response to salience (i.e., an attempt to deal with a potent distractor), or whether it is merely distinguishing target features from non-target features (Lien et al., 2021; Stilwell & Vecera, 2019). Relatedly, Gaspelin and Luck (2018b, Exp. 4) found that salient distractors in a novel color captured overt attention, which seems to indicate that without feature-based suppression, salient items generate a saliency signal that captures attention. This would seem to indicate that feature-based suppression is necessary to prevent capture.
Conclusions
The current study indicates that abrupt onsets cannot be suppressed to the same extent as color singletons. Bright, white onsets captured attention strongly. Even when onsets were matched to color singletons in terms of offsets, luminance, and color, they still captured attention. In contrast, color singletons were suppressed below baseline in all experiments. Thus, these results demonstrate that, compared to color singletons, abrupt onsets have a greater inherent ability to capture attention and resist suppression.
References
Abrams, R. A., & Christ, S. E. (2003). Motion onset captures attention. Psychological Science, 14(5), 427–432. https://doi.org/10.1111/1467-9280.01458
Adam, K. C. S., & Serences, J. T. (2021). History modulates early sensory processing of salient distractors. Journal of Neuroscience, 41(38), 8007–8022. https://doi.org/10.1523/JNEUROSCI.3099-20.2021
Adam, K. C. S., Patel, T., Rangan, N., & Serences, J. T. (2021). Classic visual search effects in an additional singleton task: An open dataset. Journal of Cognition, 4(1), 34. https://doi.org/10.5334/joc.182
Adams, O. J., & Gaspelin, N. (2021). Introspective awareness of oculomotor attentional capture. Journal of Experimental Psychology: Human Perception and Performance, 47(3), 442–459. https://doi.org/10.1037/xhp0000898
Al-Aidroos, N., Guo, R. M., & Pratt, J. (2010). You can’t stop new motion: Attentional capture despite a control set for colour. Visual Cognition, 18(6), 859–880. https://doi.org/10.1080/13506280903343085
Anderson, B. A., & Mrkonja, L. (2021). Oculomotor feedback rapidly reduces overt attentional capture. Cognition, 217, 104917. https://doi.org/10.1016/j.cognition.2021.104917
Bacon, W. F., & Egeth, H. E. (1994). Overriding stimulus-driven attentional capture. Perception & Psychophysics, 55(5), 485–496. https://doi.org/10.3758/BF03205306
Beck, V. M., Hollingworth, A., & Luck, S. J. (2012). Simultaneous control of attention by multiple working memory representations. Psychological Science, 23, 887–898. https://doi.org/10.1177/0956797612439068
Becker, S. I., & Lewis, A. J. (2015). Oculomotor capture by irrelevant onsets with and without color contrast: Oculomotor capture by onsets. Annals of the New York Academy of Sciences, 1339(1), 60–71. https://doi.org/10.1111/nyas.12685
Becker, S. I., Lewis, A. J., & Axtens, J. E. (2017). Top-down knowledge modulates onset capture in a feedforward manner. Psychonomic Bulletin & Review, 24(2), 436–446. https://doi.org/10.3758/s13423-016-1134-2
Belopolsky, A. V., Kramer, A. F., & Theeuwes, J. (2008). The role of awareness in processing of oculomotor capture: Evidence from event-related potentials. Journal of Cognitive Neuroscience, 20(12), 2285–2297. https://doi.org/10.1162/jocn.2008.20161
Bonetti, F., & Turatto, M. (2019). Habituation of oculomotor capture by sudden onsets: Stimulus specificity, spontaneous recovery and dishabituation. Journal of Experimental Psychology: Human Perception and Performance, 45(2), 264–284. https://doi.org/10.1037/xhp0000605
Brainard, D. H. (1997). The Psychophysics Toolbox. Spatial Vision, 10(4), 433–436. https://doi.org/10.1163/156856897X00357
Burnham, B. R. (2007). Displaywide visual features associated with a search display’s appearance can mediate capture. Psychonomic Bulletin & Review, 14(3), 392–422. https://doi.org/10.3758/BF03194082
Burnham, B. R. (2020). Evidence for early top-down modulation of attention to salient visual cues through probe detection. Attention, Perception, & Psychophysics, 82(3), 1003–1023. https://doi.org/10.3758/s13414-019-01850-0
Chang, S., & Egeth, H. E. (2019). Enhancement and suppression flexibly guide attention. Psychological Science, 30(12), 1724–1732. https://doi.org/10.1177/0956797619878813
Chang, S., & Egeth, H. E. (2021). Can salient stimuli really be suppressed? Attention, Perception, & Psychophysics, 83(1), 260–269. https://doi.org/10.3758/s13414-020-02207-8
Cornelissen, F. W., Peters, E. M., & Palmer, J. (2002). The Eyelink Toolbox: Eye tracking with MATLAB and the Psychophysics Toolbox. Behavior Research Methods, Instruments, & Computers, 34(4), 613–617. https://doi.org/10.3758/BF03195489
Cunningham, C. A., & Egeth, H. E. (2016). Taming the white bear: Initial costs and eventual benefits of distractor inhibition. Psychological Science, 27(4), 476–485. https://doi.org/10.1177/0956797615626564
De Tommaso, M., & Turatto, M. (2019). Learning to ignore salient distractors: Attentional set and habituation. Visual Cognition, 27(3–4), 214–226. https://doi.org/10.1080/13506285.2019.1583298
Drisdelle, B. L., & Eimer, M. (2021). PD components and distractor inhibition in visual search: New evidence for the signal suppression hypothesis. Psychophysiology, 00, e13878. https://doi.org/10.1111/psyp.13878
Donk, M., & van Zoest, W. (2008). Effects of salience are short-lived. Psychological Science, 19(7), 733–739. https://doi.org/10.1111/j.1467-9280.2008.02149.x
Feldmann-Wüstefeld, T., Busch, N. A., & Schubö, A. (2020). Failed suppression of salient stimuli precedes behavioral errors. Journal of Cognitive Neuroscience, 32(2), 367–377.
Feldmann-Wüstefeld, T., Weinberger, M., & Awh, E. (2021). Spatially guided distractor suppression during visual search. Journal of Neuroscience, 41(14), 3180. https://doi.org/10.1523/JNEUROSCI.2418-20.2021
Folk, C., & Remington, R. (1998). Selectivity in distraction by irrelevant featural singletons: Evidence for two forms of attentional capture. Journal of Experimental Psychology: Human Perception and Performance, 24(3), 847–858. https://doi.org/10.1037//0096-1523.24.3.847
Folk, C. L., & Remington, R. W. (2015). Unexpected abrupt onsets can override a top-down set for color. Journal of Experimental Psychology: Human Perception and Performance, 41(4), 1153–1165. https://doi.org/10.1037/xhp0000084
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992). Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology: Human Perception and Performance, 18(4), 1030–1044. https://doi.org/10.1037//0096-1523.18.4.1030
Folk, C. L., Remington, R. W., & Wright, J. H. (1994). The structure of attentional control: Contingent attentional capture by apparent motion, abrupt onset, and color. Journal of Experimental Psychology: Human Perception & Performance, 20(2), 317–329.
Forschack, N., Gundlach, C., Hillyard, S., & Müller, M. M. (2022). Electrophysiological evidence for target facilitation without distractor suppression in two-stimulus search displays. Cerebral Cortex. https://doi.org/10.1093/cercor/bhab450
Franconeri, S. L., & Simons, D. J. (2003). Moving and looming stimuli capture attention. Perception & Psychophysics, 65(7), 999–1010. https://doi.org/10.3758/bf03194829
Franconeri, S. L., Hollingworth, A., & Simons, D. J. (2005). Do new objects capture attention? Psychological Science, 16(4), 275–281. https://doi.org/10.1111/j.0956-7976.2005.01528.x
Fuchs, I., Theeuwes, J., & Ansorge, U. (2013). Exogenous attentional capture by subliminal abrupt-onset cues: Evidence from contrast-polarity independent cueing effects. Journal of Experimental Psychology: Human Perception and Performance, 39(4), 974–988. https://doi.org/10.1037/a0030419
Gabbay, C., Zivony, A., & Lamy, D. (2019). Splitting the attentional spotlight? Evidence from attentional capture by successive events. Visual Cognition, 27(5-8), 518–536. https://doi.org/10.1080/13506285.2019.1617377
Gaspar, J. M., & McDonald, J. J. (2014). Suppression of salient objects prevents distraction in visual search. Journal of Neuroscience, 34(16), 5658–5666. https://doi.org/10.1523/JNEUROSCI.4161-13.2014
Gaspelin, N., & Luck, S. J. (2018a). Distinguishing among potential mechanisms of singleton suppression. Journal of Experimental Psychology: Human Perception and Performance, 44(4), 626–644. https://doi.org/10.1037/xhp0000484
Gaspelin, N., & Luck, S. J. (2018b). Combined electrophysiological and behavioral evidence for the suppression of salient distractors. Journal of Cognitive Neuroscience, 30(9), 1265–1280. https://doi.org/10.1162/jocn_a_01279
Gaspelin, N., & Luck, S. J. (2018c). The role of inhibition in avoiding distraction by salient stimuli. Trends in Cognitive Sciences, 22(1), 79–92. https://doi.org/10.1016/j.tics.2017.11.001
Gaspelin, N., Leonard, C. J., & Luck, S. J. (2015). Direct evidence for active suppression of salient-but-irrelevant sensory inputs. Psychological Science, 26(11), 1740–1750. https://doi.org/10.1177/0956797615597913
Gaspelin, N., Ruthruff, E., & Lien, M. C. (2016). The problem of latent attentional capture: Easy search conceals capture by task-irrelevant abrupt onsets. Journal of Experimental Psychology: Human Perception and Performance, 42(8), 1104–1120. https://doi.org/10.1037/xhp0000214
Gaspelin, N., Leonard, C. J., & Luck, S. J. (2017). Suppression of overt attentional capture by salient-but-irrelevant color singletons. Attention, Perception, & Psychophysics, 79(1), 45–62. https://doi.org/10.3758/s13414-016-1209-1
Gaspelin, N., Gaspar, J. M., & Luck, S. J. (2019). Oculomotor inhibition of salient distractors: Voluntary inhibition cannot override selection history. Visual Cognition, 27(3/4), 227–246. https://doi.org/10.1080/13506285.2019.1600090
Goller, F., Ditye, T., & Ansorge, U. (2016). The contribution of color to attention capture effects during search for onset targets. Attention, Perception, & Psychophysics, 78(3), 789–807. https://doi.org/10.3758/s13414-015-1053-8
Goller, F., Schoeberl, T., & Ansorge, U. (2020). Testing the top-down contingent capture of attention for abrupt-onset cues: Evidence from cue-elicited N2pc. Psychophysiology, 57(11), e13655. https://doi.org/10.1111/psyp.13655
Grubert, A., & Eimer, M. (2015). Rapid parallel attentional target selection in single-color and multiple-color visual search. Journal of Experimental Psychology: Human Perception and Performance, 41(1), 86–101. https://doi.org/10.1037/xhp0000019
Irwin, D. E., Colcombe, A. M., Kramer, A. F., & Hahn, S. (2000). Attentional and oculomotor capture by onset, luminance, and color singletons. Vision Research, 40, 1443–1458.
Jonides, J., & Yantis, S. (1988). Uniqueness of abrupt visual onset in capturing attention. Perception & Psychophysics, 43(4), 346–354. https://doi.org/10.3758/BF03208805
Kerzel, D., & Burra, N. (2020). Capture by context elements, not attentional suppression of distractors, explains the PD with small search displays. Journal of Cognitive Neuroscience, 32(6), 1170–1183. https://doi.org/10.1162/jocn_a_01535
Lamy, D., & Egeth, H. E. (2003). Attentional capture in singleton-detection and feature-search modes. Journal of Experimental Psychology: Human Perception and Performance, 29(5), 1003–1020. https://doi.org/10.1037/0096-1523.29.5.1003
Lamy, D., Darnell, M., Levi, A., & Bublil, C. (2018). Testing the attentional dwelling hypothesis of attentional capture. Journal of Cognition, 1(1), 1–16. https://doi.org/10.5334/joc.48
Leber, A. B., & Egeth, H. E. (2006). It’s under control: Top-down search strategies can override attentional capture. Psychonomic Bulletin & Review, 13(1), 132–138. https://doi.org/10.3758/BF03193824
Leonard, C. J., & Luck, S. J. (2011). The role of magnocellular signals in oculomotor attentional capture. Journal of Vision, 11(13), 1–20. https://doi.org/10.1167/11.13.11
Lien, M.-C., Ruthruff, E., Goodin, Z., & Remington, R. W. (2008). Contingent attentional capture by top-down control settings: Converging evidence from event-related potentials. Journal of Experimental Psychology: Human Perception and Performance, 34(3), 509–530. https://doi.org/10.1037/0096-1523.34.3.509
Lien, M.-C., Ruthruff, E., & Johnston, J. C. (2010). Attentional capture with rapidly changing attentional control settings. Journal of Experimental Psychology: Human Perception and Performance, 36(1), 1–16. https://doi.org/10.1037/a0015875
Lien, M.-C., Ruthruff, E., & Hauck, C. (2021). On preventing attention capture: Is singleton suppression actually singleton suppression? Psychological Research. https://doi.org/10.1007/s00426-021-01599-y
Luck, S. J. (2012). Electrophysiological correlates of the focusing of attention within complex visual scenes: N2pc and related ERP components. In S. J. Luck & E. S. Kappenman (Eds.), The Oxford handbook of event-related potential components (pp. 329–360). Oxford University Press.
Luck, S. J., Gaspelin, N., Folk, C. L., Remington, R. W., & Theeuwes, J. (2021). Progress toward resolving the attentional capture debate. Visual Cognition, 29(1), 1–21. https://doi.org/10.1080/13506285.2020.1848949
Ludwig, C. J. H., & Gilchrist, I. D. (2003). Goal-driven modulation of oculomotor capture. Perception & Psychophysics, 65(8), 1243–1251. https://doi.org/10.3758/BF03194849
Maxwell, J. W., Gaspelin, N., & Ruthruff, E. (2021). No identification of abrupt onsets that capture attention: Evidence against a unified model of spatial attention. Psychological Research, 85(5), 2119–2135. https://doi.org/10.1007/s00426-020-01367-4
Miller, J. (1989). The control of attention by abrupt visual onsets and offsets. Perception & Psychophysics, 45(6), 567–571. https://doi.org/10.3758/BF03208064
Moore, C. M., & Osman, A. M. (1993). Looking for two targets at the same time: One search or two? Perception & Psychophysics, 53(4), 381–390. https://doi.org/10.3758/BF03206781
Morey, R. D. (2008). Confidence intervals from normalized data: A correction to Cousineau (2005). Tutorials in Quantitative Methods for Psychology, 4(2), 61–64. https://doi.org/10.20982/tqmp.04.2.p061
Mounts, J. R. W. (2000). Attentional capture by abrupt onsets and feature singletons produces inhibitory surrounds. Perception & Psychophysics, 62(7), 1485–1493. https://doi.org/10.3758/BF03212148
Pascucci, D., & Turatto, M. (2015). The distracting impact of repeated visible and invisible onsets on focused attention. Journal of Experimental Psychology: Human Perception and Performance, 41(3), 879–892. https://doi.org/10.1037/xhp0000025
Pashler, H. E. (1988). Cross-dimensional interaction and texture segregation. Perception & Psychophysics, 43(4), 307–318. https://doi.org/10.3758/BF03208800
Roque, N. A., Wright, T. J., & Boot, W. R. (2016). Do different attention capture paradigms measure different types of capture? Attention, Perception, & Psychophysics, 78(7), 2014–2030. https://doi.org/10.3758/s13414-016-1117-4
Ruthruff, E., Kuit, D., Maxwell, J. W., & Gaspelin, N. (2019). Can capture by abrupt onsets be suppressed? Visual Cognition, 27(3-4), 279–290. https://doi.org/10.1080/13506285.2019.1604593
Ruthruff, E., Faulks, M., Maxwell, J. W., & Gaspelin, N. (2020). Attentional dwelling and capture by color singletons. Attention, Perception, & Psychophysics, 82, 3048–3064. https://doi.org/10.3758/s13414-020-02054-7
Sawaki, R., & Luck, S. J. (2010). Capture versus suppression of attention by salient singletons: Electrophysiological evidence for an automatic attend-to-me signal. Attention, Perception, & Psychophysics, 72(6), 1455–1470. https://doi.org/10.3758/APP.72.6.1455
Schönhammer, J. G., & Kerzel, D. (2018). Optimal task-sets override attentional capture by rare cues. Journal of Experimental Psychology: Human Perception and Performance, 44(5), 681–692. https://doi.org/10.1037/xhp0000483
Schreij, D., Owens, C., & Theeuwes, J. (2008). Abrupt onsets capture attention independent of top-down control settings. Perception & Psychophysics, 70(2), 208–218. https://doi.org/10.3758/PP.70.2.208
Schreij, D., Theeuwes, J., & Oliver, C. N. L. (2010a). Irrelevant onsets cause inhibition of return regardless of attentional set. Attention, Perception & Psychophysics, 72(7), 1725–1729. https://doi.org/10.3758/APP.72.7.1725
Schreij, D., Theeuwes, J., & Olivers, C. N. L. (2010b). Abrupt onsets capture attention independent of top-down control settings II: Additivity is no evidence for filtering. Attention, Perception, & Psychophysics, 72(3), 672–682. https://doi.org/10.3758/APP.72.3.672
Stilwell, B. T., & Gaspelin, N. (2021). Attentional suppression of highly salient color singletons. Journal of Experimental Psychology: Human Perception & Performance, 47(10), 1313–1328. https://doi.org/10.1037/xhp0000948
Stilwell, B. T., & Vecera, S. P. (2019). Cued distractor rejection disrupts learned distractor rejection. Visual Cognition, 27(3), 327–342. https://doi.org/10.1080/13506285.2018.1564808
Stilwell, B. T., & Vecera, S. P. (2022). Testing the underlying processes leading to learned distractor rejection: Learned oculomotor avoidance. Attention, Perception, & Psychophysics. https://doi.org/10.3758/s13414-022-02483-6
Stilwell, B. T., Egeth, H. E., & Gaspelin, N. (2022). Electrophysiological evidence that highly salient color singletons can be proactively suppressed. Journal of Cognitive Neuroscience. https://doi.org/10.1162/jocn_a_01827
Talcott, T. N., & Gaspelin, N. (2020). Prior target locations attract overt attention during search. Cognition, 201, 104282. https://doi.org/10.1016/j.cognition.2020.104282
Theeuwes, J. (1992). Perceptual selectivity for color and form. Perception & Psychophysics, 51(6), 599–606. https://doi.org/10.3758/BF03211656
Theeuwes, J., Kramer, A. F., Hahn, S., & Irwin, D. E. (1998). Our eyes do not always go where we want them to go: Capture of the eyes by new objects. Psychological Science, 9(5), 379–385. https://doi.org/10.1111/1467-9280.00071
Theeuwes, J., Kramer, A. F., Hahn, S., Irwin, D. E., & Zelinsky, G. J. (1999). Influence of attentional capture on oculomotor control. Journal of Experimental Psychology: Human Perception and Performance, 25(6), 1595–1608. https://doi.org/10.1037/0096-1523.25.6.1595
Todd, J. T., & Van Gelder, P. (1979). Implications of a transient-sustained dichotomy for the measurement of human performance. Journal of Experimental Psychology: Human Perception and Performance, 5(4), 625–638. https://doi.org/10.1037/0096-1523.5.4.625
Turatto, M., & Pascucci, D. (2016). Short-term and long-term plasticity in the visual-attention system: Evidence from habituation of attentional capture. Neurobiology of Learning and Memory, 130, 159–169. https://doi.org/10.1016/j.nlm.2016.02.010
Turatto, M., Bonetti, F., & Pascucci, D. (2018a). Filtering visual onsets via habituation: A context-specific long-term memory of irrelevant stimuli. Psychonomic Bulletin & Review, 25(3), 1028–1034. https://doi.org/10.3758/s13423-017-1320-x
Turatto, M., Bonetti, F., Pascucci, D., & Chelazzi, L. (2018b). Desensitizing the attention system to distraction while idling: A new latent learning phenomenon in the visual attention domain. Journal of Experimental Psychology: General, 147(12), 1827–1850. https://doi.org/10.1037/xge0000503
van Moorselaar, D., & Slagter, H. A. (2019). Learning what is irrelevant or relevant: Expectations facilitate distractor inhibition and target facilitation through distinct neural mechanisms. Journal of Neuroscience, 39(35), 6953–6967. https://doi.org/10.1523/JNEUROSCI.0593-19.2019
van Zoest, W., Donk, M., & Theeuwes, J. (2004). The role of stimulus-driven and goal-driven control in saccadic visual selection. Journal of Experimental Psychology: Human Perception and Performance, 30(4), 746–759. https://doi.org/10.1037/0096-1523.30.4.746
Vatterott, D. B., & Vecera, S. P. (2012). Experience-dependent attentional tuning of distractor rejection. Psychonomic Bulletin & Review, 19(5), 871–878. https://doi.org/10.3758/s13423-012-0280-4
Vatterott, D. B., Mozer, M. C., & Vecera, S. P. (2018). Rejecting salient distractors: Generalization from experience. Attention, Perception, & Psychophysics, 80(2), 485–499. https://doi.org/10.3758/s13414-017-1465-8
Wang, B., & Theeuwes, J. (2018). Statistical regularities modulate attentional capture. Journal of Experimental Psychology: Human Perception and Performance, 44(1), 13–17. https://doi.org/10.1037/xhp0000472
Weichselbaum, H., Fuchs, I., & Ansorge, U. (2014). Oculomotor capture by supraliminal and subliminal onset singletons: The role of contrast polarity. Vision Research, 100, 1–7. https://doi.org/10.1016/j.visres.2014.03.014
Won, B. Y., & Geng, J. J. (2018). Learned suppression for multiple distractors in visual search. Journal of Experimental Psychology: Human Perception and Performance, 44(7), 1128–1141. https://doi.org/10.1037/xhp0000521
Won, B.-Y., & Geng, J. J. (2020). Passive exposure attenuates distraction during visual search. Journal of Experimental Psychology, 149(10), 1987–1995. https://doi.org/10.1037/xge0000760
Won, B., Kosoyan, M., & Geng, J. J. (2019). Evidence for second-order singleton suppression based on probabilistic expectations. Journal of Experimental Psychology: Human Perception and Performance, 45(1), 125–138. https://doi.org/10.1037/xhp0000594
Won, B.-Y., Venkatesh, A., Witkowski, P. P., Banh, T., & Geng, J. J. (2022). Memory precision for salient distractors decreases with learned suppression. Psychonomic Bulletin & Review, 29(1), 169–181. https://doi.org/10.3758/s13423-021-01968-z
Wöstmann, M., Störmer, V. S., Obleser, J., Addleman, D. A., Andersen, S. K., Gaspelin, N., Geng, J. J., Luck, S. J., Noonan, M. P., Slagter, H. A., & Theeuwes, J. (2022). Ten simple rules to study distractor suppression. Progress in Neurobiology, 213, 102269. https://doi.org/10.1016/j.pneurobio.2022.102269
Wu, S.-C., & Remington, R. (2003). Characteristics of covert and overt visual orienting: Evidence from attentional and oculomotor capture. Journal of Experimental Psychology: Human Perception & Performance, 29(5), 1050–1067. https://doi.org/10.1037/0096-1523.29.5.1050
Yantis, S., & Jonides, J. (1984). Abrupt visual onsets and selective attention: Evidence from visual search. Journal of Experimental Psychology: Human Perception and Performance, 10(5), 601–621. https://doi.org/10.1037/0096-1523.10.5.601
Zivony, A., & Lamy, D. (2018). Contingent attentional engagement: Stimulus- and goal-driven capture have qualitatively different consequences. Psychological Science, 29(12), 1930–1941. https://doi.org/10.1177/0956797618799302
Open Practices Statement
The stimulus presentation programs, original data, and data summaries for all experiments are available on the Open Science Framework at the following link: https://osf.io/852qe/?view_only=6d501ad27d624d768f351bd14addc749. None of the experiments were preregistered.
Author Note
This project was made possible by National Science Foundation Grant BCS-2045624 to N.G. Stimulus presentation programs, original data, and data summaries are available on OSF.io (https://osf.io/852qe/?view_only=6d501ad27d624d768f351bd14addc749).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
ESM 1
(DOCX 27 kb)
Rights and permissions
About this article

Cite this article
Adams, O.J., Ruthruff, E. & Gaspelin, N. Oculomotor suppression of abrupt onsets versus color singletons. Atten Percept Psychophys 85, 613–633 (2023). https://doi.org/10.3758/s13414-022-02524-0
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-022-02524-0