
Abstract
Glucocorticoid hormones were discovered to have use as potent anti-inflammatory and immunosuppressive therapeutics in the 1940s and their continued use and development have successfully revolutionized the management of acute and chronic inflammatory diseases. However, long-term use of glucocorticoids is severely hampered by undesirable metabolic complications, including the development of type 2 diabetes mellitus. These effects occur due to glucocorticoid receptor activation within multiple tissues, which results in inter-organ crosstalk that increases hepatic glucose production and inhibits peripheral glucose uptake. Despite the high prevalence of glucocorticoid-induced hyperglycaemia associated with their routine clinical use, treatment protocols for optimal management of the metabolic adverse effects are lacking or underutilized. The type, dose and potency of the glucocorticoid administered dictates the choice of hypoglycaemic intervention (non-insulin or insulin therapy) that should be provided to patients. The longstanding quest to identify dissociated glucocorticoid receptor agonists to separate the hyperglycaemic complications of glucocorticoids from their therapeutically beneficial anti-inflammatory effects is ongoing, with selective glucocorticoid receptor modulators in clinical testing. Promising areas of preclinical research include new mechanisms to disrupt glucocorticoid signalling in a tissue-selective manner and the identification of novel targets that can selectively dissociate the effects of glucocorticoids. These research arms share the ultimate goal of achieving the anti-inflammatory actions of glucocorticoids without the metabolic consequences.
Key points
-
Glucocorticoid drugs are widely prescribed to treat inflammatory diseases and to prevent organ transplant rejection yet they promote hyperglycaemia and diabetes mellitus.
-
Glucocorticoid-induced hyperglycaemia manifests directly via glucocorticoid signalling in metabolic organs and tissues (liver, adipose tissue, muscle, bone and pancreatic β-cells) and indirectly via inter-organ hormone and metabolite flux.
-
No single consensus exists with respect to optimal screening frequency for glucocorticoid-induced hyperglycaemia due to context-dependent factors, although analysis of postprandial glucose (not fasting glucose) is recommended.
-
Hypoglycaemic agents (such as insulin sensitizers and insulin) can provide satisfactory glucose control but need to be tailored for differences in the dose and type of glucocorticoid used.
-
Developing selective glucocorticoid modulators that dissociate the diabetogenic from the anti-inflammatory effects of glucocorticoids continues to be challenging; however, new pharmacological targets that prevent diabetogenic effects are on the horizon.
Similar content being viewed by others
Introduction
Cortisol is a human glucocorticoid hormone that is essential for survival under periods of stress1. In 1949, exogenous glucocorticoid supplementation for patients with rheumatoid arthritis was shown to provide pain relief2. This discovery led to the recognition that glucocorticoids have potent anti-inflammatory and immunosuppressive effects and fuelled their use as therapeutics in a variety of immune-associated disorders and to prevent organ transplant rejection3. At present, ~1% of the general population receive oral glucocorticoids, with this prevalence increasing to 2.5% among older adults aged 70–79 years4,5. Glucocorticoid use has gained prominence during the COVID-19 pandemic as dexamethasone, one of the most potent glucocorticoid drugs available on the market, is effective in the treatment of patients with severe COVID-19 (ref.6) (Box 1).
Despite excellent therapeutic efficacy for the treatment of inflammatory disorders, long-term glucocorticoid therapy is limited by a wide range of adverse effects7. Relevant to this Review, glucocorticoids precipitate the development of hyperglycaemia, insulin resistance, dyslipidaemia, central adiposity and hepatic steatosis. Over time, these metabolic effects result in diabetes mellitus (alternately referred to as ‘steroid diabetes’), one of the most well-established adverse effects of hyperglucocorticoidism8. Globally, oral glucocorticoid use is linked to 2% of cases of new-onset diabetes mellitus9. When glucocorticoids were administered for 1–7 days to healthy individuals, insulin resistance and impaired glucose homeostasis were observed10,11. Furthermore, a meta-analysis found that, in patients without pre-existing diabetes mellitus who were treated with glucocorticoids for more than 1 month, the incidence of glucocorticoid-induced hyperglycaemia is ~32% and the incidence of diabetes mellitus is 19%12. In patients with pre-existing diabetes mellitus, hyperglycaemia can be exacerbated by starting glucocorticoid treatment13. Of note, in Cushing syndrome, endogenous overproduction of cortisol produces similar complications to those of iatrogenic hypercortisolism14. The prevalence of diabetes mellitus among patients with endogenous Cushing syndrome is 20–45%, with an additional 10–30% of patients exhibiting impaired glucose tolerance15.
Despite the widespread use of glucocorticoids and their tight association with hyperglycaemia and diabetes mellitus, a firm understanding of the underlying mechanisms and strategies to mitigate and manage these adverse effects are lacking. The actions of glucocorticoids are predominately mediated by the glucocorticoid receptor, which is localized in the cytoplasm of target tissues16. Upon glucocorticoid binding, the glucocorticoid receptor translocates to the nucleus and can directly bind as a homodimer to DNA sequences called glucocorticoid-response elements (GREs), which are located throughout the genome17. This binding to GREs is facilitated by co-regulatory proteins and histone modifiers. In addition, the ligand-bound glucocorticoid receptor can tether to other transcription factors to indirectly upregulate or downregulate gene expression18. Studies into mechanisms and treatments to minimize or prevent glucocorticoid-induced diabetes mellitus are ongoing and eagerly anticipated to improve the clinical experience of glucocorticoid users13.
In this Review, we summarize the epidemiology of glucocorticoid-induced hyperglycaemia and our current understanding of the diabetogenic actions of glucocorticoids within different organs that contribute to the development of glucocorticoid-induced diabetes mellitus. We then discuss current diagnostic criteria and treatment options based on the type and schedule of glucocorticoid and the degree of hyperglycaemia established. Finally, we provide novel insights into promising preclinical approaches and potential targets on the horizon that could eliminate the diabetogenic effects of glucocorticoid drugs while preserving their anti-inflammatory effects.
Epidemiology
Exposure to even a single dose of exogenous glucocorticoids is associated with an abnormal elevation in blood concentrations of glucose in patients with or without pre-existing diabetes mellitus19,20. Indeed, glucocorticoids exacerbate hyperglycaemia in a dose-dependent manner irrespective of prior history of diabetes mellitus21,22. A retrospective review reported that 56% of hospitalized patients without a history of diabetes mellitus experienced at least one episode of hyperglycaemia (defined as blood glucose levels ≥200 mg/dl) after corticosteroid treatment23. In patients treated with glucocorticoids for respiratory disease24, renal disease25, cancer26, solid organ transplant27 and rheumatoid arthritis28, the incidence of new-onset glucocorticoid-induced diabetes mellitus ranges between 15% and 40%. Moreover, the incidence of glucocorticoid-induced hyperglycaemia might be underestimated because of a reliance on fasting blood levels of glucose for diagnosis (discussed later).
Effects of drug type and dosing regimen
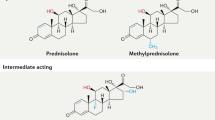
The relative risk of any individual developing glucocorticoid-induced hyperglycaemia and diabetes mellitus is difficult to predict owing to the combination of doses, variable glucocorticoid potencies, routes of administration and duration of treatment that are used for different diseases. For the purpose of this Review, we use the following definitions: acute treatment, up to 24 h; short-term treatment, between 24 h to 2 weeks; long-term treatment, 2 weeks to 6 months or longer). Glucocorticoid preparations on the market vary in their pharmacological properties and duration of anti-inflammatory action29. Relative to short-acting hydrocortisone (half-life of 8 h), intermediate-acting prednisolone (half-life of 16–36 h) and methylprednisolone (half-life of 18–40 h), which are the most commonly used glucocorticoids, are 4–5-fold more potent in their anti-inflammatory effect13,29. The long-acting dexamethasone and betamethasone (both with a half-life of 36–54 h) are 25-fold more potent than hydrocortisone13,29.
All common routes of administration for glucocorticoids (oral, inhalation and topical) are linked to an increased risk of hyperglycaemia when prescribed at high doses22,30,31,32, yet oral corticosteroids confer the greatest risk due to their systemic exposure33. Local injections of glucocorticoids (intra-articular or intra-lesional) also increase hyperglycaemia in patients with controlled diabetes mellitus34,35. Glucocorticoids exert dose-dependent effects as demonstrated by odds ratios for eventual hypoglycaemic intervention (over a 90-day period) that ranged between 1.7 and 10.3 for daily oral hydrocortisone-equivalent doses starting at 20 mg per day and increasing to ≥120 mg per day, respectively22. In patients who underwent an organ transplant, the risk of developing post-transplant diabetes mellitus was also found to increase by 5% per 0.01 mg/kg per day increase in prednisolone dose administered by intravenous injection36. Inhaled corticosteroids used at high doses (fluticasone at ≥1,000 µg per day) increased the incidence of diabetes mellitus by 34% in patients with respiratory disease30. A positive correlation was also observed between the relative potency of a prescribed topical glucocorticoid and the incidence of diabetes mellitus in case–control studies of adults in Denmark and the UK31. Dosing regimen also contributes substantially to the precipitation of diabetes mellitus as the incidence of hyperglycaemia was 50% more frequent in patients with haematological diseases who received long-term continuous administration of glucocorticoids over 6-weeks compared with cyclic administration (5 days on, 15 days off)37. Patients without a history of diabetes mellitus who experienced multiple episodes of hyperglycaemia after glucocorticoid therapy were found to be on comparatively longer corticosteroid regimens than those who experienced ≤1 episode of hyperglycaemia23.
Other risk factors
Many factors are associated with the susceptibility of patients on glucocorticoid therapy to develop hyperglycaemia and diabetes mellitus (Box 2). Older age (>60 years) is tightly linked to an increased risk of glucocorticoid-induced diabetes mellitus due to declining pancreatic β-cell function and increased glucose intolerance with age24,25,38. High BMI (>25 kg/m2)38, abdominal obesity and hypertriglyceridaemia39 are other independent risk factors that contribute to glucocorticoid-induced diabetes mellitus25. In addition, prior reductions in insulin sensitivity or deficient glucose-stimulated insulin secretion (GSIS)40,41, prior glucose intolerance or impaired fasting levels of glucose42,43, and high HbA1c values (≥6.0%)44 are further indicators of increased risk of glucocorticoid-induced diabetes mellitus.
The extent to which a family history of diabetes mellitus contributes to new-onset glucocorticoid-induced diabetes mellitus is still not clear. Several studies do report a positive link to a family history of diabetes mellitus45,46,47. In a study that compared a cohort of patients with glucocorticoid-induced diabetes mellitus to patients with type 2 diabetes mellitus (T2DM), a family history of diabetes mellitus was found in 35% versus 65% of patients, respectively48. In 2021, specific single-nucleotide polymorphisms were shown to correlate with the development of hyperglycaemia in children receiving dexamethasone for the treatment of acute lymphoblastic leukaemia49. Mechanistically, the single-nucleotide polymorphisms modified the genomic binding of the glucocorticoid receptor to metabolic genes in adipose and liver cells49. Although these data are intriguing, they will need to be replicated in larger cohorts.
Other less well-studied risk factors include impaired renal function (estimated glomerular filtration rate <40 ml/min/1.73 m2)44,47, presence of hypertension50, cytomegalovirus infection36, current smoking38, and race and ethnicity51. No consistent sex differences were observed in glucocorticoid-induced insulin resistance in healthy individuals52 or hospital inpatients44. However, a trend was observed towards an increased incidence of glucocorticoid-induced hyperglycaemia in hospitalized men38. The incidence of glucocorticoid-induced hyperglycaemia might also be confounded by the use of concomitant immunosuppressive or diuretic medications36,50. Patients who underwent an organ transplant and received calcineurin inhibitors concurrent with glucocorticoid therapy would be expected to show more severe glucose intolerance than those not on calcineurin inhibitors due to the suppressive effect of calcineurin inhibitors on insulin production48. Similarly, the concurrent use of the immunosuppressant mycophenolate mofetil with high-dose prednisolone treatment (≥1 mg/kg per day) in patients with systemic lupus erythematosus was associated with the development of diabetes mellitus, possibly explained by pancreatic β-cell dysfunction and impaired insulin secretion50. Thus, several predictors beyond glucocorticoid potency, dose and dosing schedule should be considered when estimating the risk of developing glucocorticoid-induced diabetes mellitus.
Pathophysiology
The development of glucocorticoid-induced hyperglycaemia involves stimulation of hepatic glucose production and increased adipose tissue lipolysis followed by the development of whole-body insulin resistance as well as impaired production and secretion of insulin by pancreatic β-cells. Many of these effects are mediated by direct actions of the glucocorticoid receptor, which binds to specific target genes and controls their transcriptional expression. As described in this section, the tissue-specific effects of glucocorticoids contribute to amplifying their whole-body diabetogenic effect in vivo through inter-organ signalling (Fig. 1).

Glucocorticoids increase appetite and promote the intake of high-calorie (high-fat and/or high-sugar) ‘comfort food’, which indirectly promotes obesity and diabetes mellitus. Glucocorticoids upregulate the transcriptional and functional activity of neuropeptide Y (NPY)–agouti-related peptide (AgRP) neurons in the arcuate nucleus of the hypothalamus and promote leptin resistance. Skeletal muscle atrophy results from glucocorticoid-mediated protein degradation and decreased protein synthesis in myocytes, and glucocorticoids also decrease glucose uptake into these cells. In the liver, glucocorticoids act directly to upregulate enzymes involved in gluconeogenesis and promote hepatic insulin resistance, which together accelerate the development of hyperglycaemia. Furthermore, glucocorticoids synergize with insulin to stimulate non-esterified fatty acid (NEFA) uptake by hepatocytes and triglyceride synthesis in the liver, which causes hepatic steatosis. In adipose tissue, glucocorticoids increase adipogenesis, de novo lipogenesis and triglyceride synthesis as well as lipid uptake and storage. Concurrently, glucocorticoids facilitate lipolysis, which promotes the futile cycling of lipids. Glucocorticoids also decrease glucose uptake into adipocytes. Acute exposure of pancreatic β-cells to glucocorticoids can stimulate insulin secretion and β-cell hyperplasia to counterbalance glucocorticoid-induced insulin resistance and to maintain plasma levels of glucose within the physiological range. However, long-term exposure to glucocorticoids can interfere with insulin biosynthesis and secretion and induce β-cell apoptosis. Osteocalcin is secreted by osteoblasts and circulating osteocalcin from bone promotes insulin secretion by β-cells. Glucocorticoids suppress the expression of osteocalcin, thereby indirectly inhibiting insulin secretion. The increase in circulating levels of amino acids from muscle breakdown and NEFAs and glycerol from adipose tissue lipolysis provide substrates to the liver for gluconeogenesis. High plasma levels of NEFAs also accumulate ectopically in skeletal muscle, liver and β-cells, which further exacerbates insulin resistance. Thick solid arrows indicate effects; thin solid arrows indicate a transition in time; dashed arrows indicate secreted factors.
Liver
Hepatic gluconeogenesis
The liver serves as a central node that links nutritional and hormonal cues to whole-body glucose homeostasis. When circulating glucose is scarce, the liver maintains euglycaemia by increasing gluconeogenesis and glycogenolysis. During a period of stress, glucocorticoids promote increased liver glucose output to ensure that the organism has enough glucose to fuel the brain and survive53. In response to increased circulating levels of glucocorticoids, the liver upregulates enzymes involved in de novo glucose synthesis. Actions of glucocorticoids on other tissues accelerate the shuttling of gluconeogenic precursors to the liver through protein catabolism in skeletal muscle54 and lipolysis in white adipose tissue (WAT)55, which further drive hepatic gluconeogenesis.
At the molecular level, the glucocorticoid–glucocorticoid receptor complex binds DNA in hepatocytes to activate gene networks involved in the hepatic metabolism of carbohydrates. For gluconeogenesis, GREs are located in the respective promoters of the genes encoding two critical enzymes, phosphoenolpyruvate carboxykinase (PEPCK, encoded by PCK1) and glucose-6-phosphatase (G6Pase, encoded by G6PC1)56,57. Activation of these glucocorticoid receptor target genes is only maximized when there is concurrent recruitment of other factors to the regulatory regions on DNA. For example, the co-regulator peroxisome proliferator-activated receptor coactivator 1α (PGC1α) and transcription factor Forkhead box protein O1 (FOXO1) are both directly regulated by glucocorticoids and synergistically potentiate activation of PEPCK53,58.
Hepatic insulin resistance
A second key pathway contributing to glucocorticoid-induced hyperglycaemia is the development of hepatic insulin resistance. Under normal physiological conditions, insulin potently suppresses hepatic gluconeogenesis59. However, in the presence of glucocorticoids, hepatic insulin resistance occurs, which enables gluconeogenesis to proceed unopposed due to the loss of insulin-mediated repression53,60. Mechanistically, the phosphorylation of downstream signalling messengers of the insulin cascade (that is, insulin receptor substrates (IRSs), phosphatidylinositol 3-kinase (PI3K) and protein kinase B (PKB)–AKT) was found to be impaired in rats treated with glucocorticoids61. Despite glucocorticoid-induced insulin resistance occurring within the gluconeogenic pathway, the pro-lipogenic effect of insulin remains intact62. Compensatory hyperinsulinaemia occurs in response to glucocorticoid-induced hyperglycaemia and, when this condition is paired with increased hepatic glucose delivery (from glucocorticoid-induced peripheral insulin resistance), synergistic stimulation of de novo lipogenesis and subsequent hepatic steatosis occurs, which in turn exacerbates hepatic insulin resistance in a vicious feed-forward cycle in both humans and animal models63,64. Increased liver uptake of circulating non-esterified fatty acids (NEFAs) in the presence of glucocorticoids further precipitates gluconeogenesis and hepatic steatosis63,64. Glucocorticoids also stimulate the biosynthesis of ceramide in the liver, a sphingolipid that is highly associated with hepatic insulin resistance65.
Skeletal muscle
Muscle insulin signalling and glucose uptake
Skeletal muscle is the primary site where insulin-stimulated glucose uptake and glycogen storage occur postprandially66. Glucocorticoids can directly disrupt insulin signalling in skeletal muscle by downregulating the expression and phosphorylation of IRS1, PI3K and PKB–AKT as shown in rats67,68,69. This effect results in concomitant inhibition of insulin-induced recruitment of glucose transporter 4 (GLUT4) to the cell surface of skeletal muscle70. In mouse myotubes, GREs were identified in numerous genes that affect insulin signalling (such as Pik3r1, which encodes the p85α subunit of PI3K71). Increases in p85α abundance disrupt its normal interaction with IRS1. This disruption in insulin signalling indirectly decreases glycogen synthase kinase 3 phosphorylation69, which clinically manifests as decreased levels of muscle glycogen occurring in patients who receive glucocorticoids after transplantation72. Thus, glucocorticoid exposure dampens skeletal muscle insulin sensitivity and glucose uptake, which contributes to the development of glucocorticoid-induced diabetes mellitus.
Development of myopathy
Patients under long-term exposure to glucocorticoid drugs (the length of time until muscle effects occur depends on dose but is generally 2–4-weeks) experience extensive skeletal muscle atrophy that results in the development of myopathy73. Muscle mass is reduced by the combined effect of glucocorticoids to promote protein degradation within skeletal muscle while blunting protein synthesis54. Glucocorticoids act by abolishing the transport of amino acids into muscle and also inhibit myogenesis by downregulating myogenin, therefore limiting protein synthesis54. Furthermore, the stimulatory actions of insulin and insulin-like growth factor 1 (IGF1) on muscle anabolism are repressed by glucocorticoids through the inhibition of PKB–AKT phosphorylation and mammalian target of rapamycin (mTOR) phosphorylation54,74. Synchronously, proteolysis is facilitated by glucocorticoid-mediated upregulation of atrophy-related gene networks in C2C12 mouse myoblasts, including Fbxo32 (encoding atrogin 1)75 and Trim63 (encoding MuRF1)76, members of the ubiquitin–proteasome system. Myostatin, a secreted factor that promotes muscle catabolism, is concurrently upregulated by glucocorticoids and attenuates the proliferation and differentiation of myoblasts54. The effects of glucocorticoids on skeletal muscle were verified in healthy individuals who were treated for 3–6 days with high doses of glucocorticoid; in these individuals, serum levels of amino acids were elevated due to enhanced muscle proteolysis and blunted protein anabolism77,78.
Increases in circulating levels of amino acids also indirectly hinder insulin-stimulated glucose transport and glycogen synthesis in muscle tissue79. As described later, glucocorticoids also increase the circulation of NEFAs from adipose tissue lipolysis55 and the ectopic accumulation of NEFAs in skeletal muscle further exacerbates insulin resistance80. Moreover, the elevation of serum amino acids and NEFAs from glucocorticoid exposure provides substrates to the liver for gluconeogenesis.
White adipose tissue
Aberrant adipose tissue distribution and truncal obesity are hallmark clinical characteristics of patients with hypercortisolism81. Lipid redistribution from peripheral to abdominal adipose depots suggests that glucocorticoids induce depot-dependent adipose tissue breakdown and expansion, although the mechanisms are not fully understood82. Central obesity is linked to insulin resistance and the development of T2DM due to the anatomical location of hypertrophic WAT near the liver and direct drainage of NEFAs and pro-inflammatory factors into the portal circulation83. Glucocorticoids promote adipogenesis, triglyceride synthesis and adipose hypertrophy, preferentially in the abdominal adipose depot84. Of note, glucocorticoids are also an essential component of the differentiation cocktail used to transform preadipocytes into mature adipocytes in vitro84. In mouse 3T3-L1 adipocytes, GREs are present near a wide range of genes involved in adipogenesis, de novo lipogenesis, triglyceride synthesis, and lipid transport and storage85. In addition, the transcription factor Forkhead box A3 (a direct target of glucocorticoid receptor) binds to promoters of glucocorticoid receptor-regulated genes related to lipid metabolism and facilitates glucocorticoid receptor binding via chromatin remodelling, which leads to adipose expansion86.
Lipid metabolism
Although de novo lipogenesis is only a minor contributor to the overall pool of triglycerides in adipose tissue87, glucocorticoids can amplify lipogenesis in the presence of insulin88. The additive or synergistic effect of glucocorticoids and insulin has also been shown to increase the expression and activity of lipoprotein lipase, an enzyme that hydrolyses circulating triglyceride-rich lipoproteins and provides local free fatty acids that are taken up by adipocytes82. Glucocorticoids have a greater effect on lipoprotein lipase in visceral versus subcutaneous adipose depots, thereby promoting lipid accumulation specifically in the abdominal region84.
Intriguingly, despite contributing to lipid expansion, glucocorticoids concurrently increase triglyceride hydrolysis through upregulation of hormone-sensitive lipase and monoacylglycerol lipase, which promotes the futile cycling of lipids85,89. The expression of genes involved in lipolysis is regulated by the direct binding of glucocorticoid receptors to GREs within these genes or indirectly via other transcription factors, including FOXO1 and FOXO3 (refs82,85). Glucocorticoids also promote lipolysis through upregulation of ANGPTL4, which encodes angiopoietin-like 4, an adipokine that stimulates the production of intracellular cAMP in adipocytes82. NEFA release is amplified by the permissive action of glucocorticoids on β-adrenergic-stimulated lipolysis, which occurs concurrently with the loss of insulin-mediated inhibition of lipolysis due to insulin resistance84. Although some discrepant results have been reported in the literature (probably due to the concentration, duration of glucocorticoid treatment, adipose depot and/or species differences), short-term infusions of glucocorticoids (6–11 h) in humans increase serum levels of NEFA and glycerol90,91. Increased levels of NEFA and glycerol promote hyperglycaemia by accelerating hepatic gluconeogenesis and impairing liver and muscle insulin sensitivity from ectopic lipid accumulation.
Insulin sensitivity
The effects of glucocorticoids on adipose tissue insulin sensitivity are species dependent. In rodent models, glucocorticoids decrease the phosphorylation of factors downstream of insulin receptor (IRS1, PI3K and PKB–AKT) and decrease insulin-receptor stimulated GLUT4 translocation92,93. By contrast, increased insulin signalling and glucose uptake were observed with short-term (from 24 h to 7 days) glucocorticoid administration to human subcutaneous adipocytes94,95. Notably, in humans, the subcutaneous adipose depot is comparatively sensitive to insulin relative to the insulin-resistant visceral adipose depot, independent of glucocorticoid use84,96.
Glucocorticoids also indirectly affect whole-body insulin sensitivity by regulating the levels of adipokines secreted by WAT. For example, leptin and resistin are upregulated by glucocorticoids in WAT and promote insulin resistance8,84.
WAT inflammation
Another consideration is the effect of glucocorticoids on adipose tissue inflammation. Adipose tissue macrophages are associated with insulin resistance in obesity97. As glucocorticoids are globally anti-inflammatory, it is interesting that glucocorticoid treatment induces adipocyte hypertrophy and insulin resistance without increasing M1-like pro-inflammatory macrophage infiltration in adipose tissue under conditions of high-fat diet-induced obesity in mice97. Interestingly, glucocorticoids were shown to influence mouse primary peritoneal macrophage polarization in vitro, increasing M2-like macrophage markers (Cd206) and decreasing M1-like macrophage markers (Il6)98. In line with this observation, corticosterone treatment in mice promoted M2-macrophage infiltration into visceral (but not subcutaneous) adipose tissue, which was linked to impaired insulin sensitivity under a normal chow diet98. Further studies are warranted to better understand the mechanism of this macrophage–adipose tissue crosstalk.
Other adipose depots
Thermogenic adipose tissue
Brown adipocytes in brown adipose tissue (BAT) differ morphologically and functionally from white adipocytes in WAT by the presence of a high number of mitochondria, which promote non-shivering thermogenesis through the expression of UCP1. A third type of adipocyte, known as beige (or ‘brite’) adipocytes, is derived from the white adipocyte lineage and located within WAT, yet also expresses UCP1 (ref.99).
A growing body of evidence, primarily from animal models, has demonstrated the potential of targeting BAT and the browning of WAT to combat obesity and T2DM99. In rodents, glucocorticoids are known inhibitors of BAT and beige adipose tissue function that act, in part, to suppress energy expenditure by downregulating Ucp1 (refs100,101). Suppression of Ucp1 is enhanced by the overexpression of 11β-HSD1 in mouse primary brown adipocytes102. This suppressive action of glucocorticoids is mediated by a regulatory region between −4 kb and 0 kb upstream of the Ucp1 transcription start site as shown in mouse primary brown adipocytes103. Glucocorticoids can also indirectly suppress Ucp1 by inhibiting adrenocorticotropic hormone (ACTH) secretion via feedback inhibition on the hypothalamic–pituitary–adrenal axis, and ACTH has been found to promote UCP1-dependent respiration in mouse primary brown adipocytes104. Whether the glucocorticoid-mediated downregulation of Ucp1 gene expression translates to a true decrease in the thermogenic capacity of BAT remains to be clarified103. Intriguingly, acute glucocorticoid treatment in humans (three doses over 24 h) increased BAT activation (assessed by measuring supraclavicular skin temperature and cold-induced 18F-fluorodeoxyglucose uptake in BAT depots assessed by PET–CT), whereas long-term treatment (>2 weeks) suppressed glucose uptake in BAT compared with control individuals105. Whether reversing glucocorticoid-associated suppression of BAT and WAT browning is sufficient to improve insulin sensitivity and obesity in humans remains to be determined.
Bone marrow adipose tissue
Adipocytes residing within the bone marrow have gained attention in the past decade as they were found to have a permissive role in contributing to metabolic diseases. Bone marrow adipose tissue (BMAT) is hypothesized to be metabolically distinct from BAT and WAT and accounts for ~70% of bone marrow volume, where BMAT critically functions to fill the long bone cavities in adult humans106. However, the metabolic contribution of BMAT to whole-body metabolism remains poorly understood. Lipid content in bone marrow of premenopausal women is inversely correlated with whole-body insulin sensitivity107. Moreover, decreased BMAT insulin responsiveness was reported in patients with obesity and T2DM108. Glucocorticoids promote BMAT lipid accumulation without affecting glucose uptake into BMAT adipocytes, re-emphasizing the site-specific actions of glucocorticoids in adipose depots109. A better understanding of the effect of glucocorticoids in BMAT remains an exciting avenue for further investigation.
Pancreatic β-cells
Pancreatic β-cell dysfunction underlies the progression to T2DM110. Under acute physiological and/or pathological stress, where the demand for insulin is increased, β-cells adapt by enhancing their functional capacity to maintain euglycaemia within a narrow range. Long-term β-cell stress can result in systemic insulin resistance, which is later superseded by β-cell failure and consequent hyperglycaemia110. Studies that investigate the effects of glucocorticoids in isolated primary islets or in vitro systems are therefore unable to account for the adaptive responses observed in vivo.
In vitro and ex vivo studies
An inhibitory role of glucocorticoids on β-cell function was demonstrated using in vitro cell lines and rodent-derived pancreatic islets after acute (minutes to hours) treatment with glucocorticoids111,112. In rat insulinoma INS-1E cells, glucocorticoids had a minimal effect on basal insulin release but GSIS was greatly reduced112. The underlying mechanism is unclear as evidence of direct inhibition of the insulin secretory cascade is still under debate113. Under physiological conditions, the uptake of glucose by β-cells causes an increase in intracellular ATP levels that triggers closure of the ATP-sensitive K+ channels followed by depolarization of the β-cell membrane. This change in voltage enables the influx of Ca2+ and subsequent exocytosis of insulin granules114. Glucocorticoids are thought to interfere at several stages in this process by limiting glucose uptake and oxidation, facilitating futile cycling of glucose, repolarizing the β-cell membrane by enhancing K+ channel activity and in turn attenuating Ca2+ influx, and amplifying α2-adrenergic signalling74,113,115.
Apart from directly reducing insulin secretion, glucocorticoids also elicit inhibitory effects on insulin biosynthesis by inducing endoplasmic reticulum stress, which ultimately triggered apoptosis of INS-1E cells treated with glucocorticoids112. Glucocorticoids might also increase cytotoxicity by accelerating the production of reactive oxygen species, which can also induce β-cell apoptosis116. Despite the potential for glucocorticoid-induced β-cell apoptosis, glucocorticoid-induced diabetes mellitus in humans was found to be more related to insulin resistance and β-cell dysfunction rather than to decreased β-cell mass117.
In vivo studies
Studies to identify the direct effects of glucocorticoids in vivo are particularly challenging to interpret owing to compensatory factors induced by long-term glucocorticoid exposure. A single oral dose of glucocorticoids acutely impaired insulin secretion in healthy individuals, whereas short-term treatment (up to 2 weeks) resulted in hyperinsulinaemia along with subtle changes in glycaemia111,118. Healthy β-cells can increase insulin secretion to counterbalance glucocorticoid-induced insulin resistance as shown by increased β-cell function in rodent islets after in vivo glucocorticoid challenge (13 days)111. Dexamethasone treatment of rats (5 days) also increases Ca2+-dependent protein kinase Cα (PKCα) signalling in islets, enhances GSIS capacity, and increases docking and exocytosis of preformed insulin granules119. Hyperinsulinaemia induced by dexamethasone treatment in rats (5 days) is accompanied by the expansion of β-cell mass in a dose-dependent manner120. However, in susceptible populations (such as those with obesity or a family history of T2DM), where β-cells are already overworked, the compensation by β-cells to a glucocorticoid challenge is lost41,46,121. Short-term (2–5 days) exposure to glucocorticoids in these individuals decreases β-cell insulin secretion and whole-body glucose disposal41,46,121. In agreement with these findings, GSIS was impaired with long-term (24 days) glucocorticoid treatment of obese Wistar and Zucker (fa/fa) rats, which led to the development of hyperglycaemia74,122.
Interpreting the long-term effects of glucocorticoids on β-cells can also be confounded by the effect of the underlying inflammatory condition that the glucocorticoids are prescribed to treat74,123. Intriguingly, high serum levels of cortisol (still within physiological range) are positively associated with β-cell defects and suppressed insulin secretion in the general population and in patients with Cushing syndrome124,125. In addition to these mechanisms by which glucocorticoids directly induce β-cell failure, glucocorticoids also indirectly promote β-cell lipotoxicity due to high plasma levels of NEFAs74.
Bone
Osteoporosis is one of the most well-established adverse effects of long-term glucocorticoid exposure (>3 months, osteoporosis risk depends on the dose)126. Glucocorticoid-induced osteoporosis results from the dual action of glucocorticoids on the suppression of bone-forming osteoblasts and the differentiation and maturation of bone-resorbing osteoclasts127. The effects of glucocorticoids on bone might contribute to hyperglycaemia in part due to elevated leptin secretion from adipose tissue as leptin suppresses the expression of osteocalcin, an insulin-sensitizing hormone that is secreted by osteoblasts128. Intriguingly, mice that lack osteocalcin develop glucose intolerance and insulin resistance due to impaired pancreatic β-cell proliferation and insulin expression129. Indeed, circulating levels of osteocalcin are associated with improved glycaemic control in patients with T2DM130. Furthermore, a single dose of glucocorticoid in humans is correlated with reduced serum levels of osteocalcin131. Mice with decreased glucocorticoid signalling in osteoblasts (achieved through targeted overexpression of 11β-HSD2 to inactivate endogenous glucocorticoids) had higher levels of circulating osteocalcin and were protected against glucocorticoid-induced glucose and insulin intolerance compared with glucocorticoid-treated wild-type mice132. These findings demonstrate a partial role for the skeleton in the regulation of energy metabolism.
Brain
The development of glucocorticoid-induced obesity and diabetes mellitus is also partially mediated through appetite stimulation in the arcuate nucleus of the hypothalamus133. Glucocorticoids upregulate the mRNA expression of neuropeptide Y (NPY) and agouti-related peptide (AgRP) and increase the firing rate of their respective neurons, which stimulates appetite in rodents134,135. In mice, local administration of glucocorticoids to the arcuate nucleus was shown to induce hepatic insulin resistance through NPY release and signalling via the sympathetic nervous system136. Glucocorticoids stimulate leptin release from adipocytes into the circulation137. Furthermore, under physiological conditions, leptin suppresses appetite via leptin receptor signalling in NPY–AgRP neurons, which inhibits the release of these orexigenic peptides138. However, in rats, glucocorticoids were shown to synergistically antagonize the action of leptin by directly reducing leptin-dependent JAK–STAT signalling in the hypothalamus139. Aside from their orexigenic effects, glucocorticoids also promote increased caloric intake by shifting food preference towards high-calorie food that is high in fat or sugar (‘comfort food’). Rodents placed under restraint stress preferred lard and sucrose over normal chow140. Similarly, individuals with elevated circulating levels of cortisol (occurring after a stressful task) prefer high-calorie ‘comfort food’ over food that is lower in sugar and fat141. Over time, this extra caloric intake could contribute to a substantial increase in adiposity.
Current treatment options
Prior to starting glucocorticoid treatment, patients should be evaluated for risk factors for hyperglycaemia, including age, BMI and family history, and should be screened for pre-existing diabetes mellitus (measurements of fasting plasma glucose and HbA1c)38,142. At a minimum, plasma levels of glucose should be monitored 1–3 days after initiation of glucocorticoid treatment7,13. The frequency of monitoring plasma levels of glucose will increase if the patient is in hospital and/or has pre-existing T2DM29,143. Monitoring should continue at regular 3–6 month intervals for a year and then yearly thereafter7. The criteria for the diagnosis of glucocorticoid-induced diabetes mellitus are similar to other types of diabetes mellitus: fasting plasma glucose concentration ≥7.0 mM, random plasma concentration of glucose ≥11.1 mM, HbA1c ≥6.5% (48 mmol/mol) or plasma glucose concentration ≥11.1 mM 2 h after an oral glucose tolerance test (OGTT)144. Glucocorticoid exposure has a more notable effect on postprandial glycaemia compared with fasting glucose13. Thus, the prevalence of glucocorticoid-induced hyperglycaemia is probably underestimated because fasting plasma glucose is more frequently assessed compared with the more time-consuming OGTT145. Determination of postprandial glycaemia 2 h after lunch and/or OGTT provides the most ideal measure to diagnose glucocorticoid-induced diabetes mellitus13,146. Treatment should be initiated when plasma levels of glucose are repeatedly ≥12 mM according to the management guideline established by the Joint British Diabetes Societies29. The goal of hypoglycaemic treatment in glucocorticoid-induced hyperglycaemia is to achieve glucose levels ≤10 mM (refs29,147).
Few clinical trials have systematically investigated the efficacy of oral hypoglycaemic agents on glucocorticoid-induced hyperglycaemia. Currently, no one-size-fits-all consensus exists regarding the optimal treatment strategy for this condition owing to the myriad factors involved. Due to the slow onset of action and narrow flexibility in dose titration of many oral hypoglycaemic agents, it is difficult to match the hypoglycaemic agent to the hyperglycaemic oscillation induced by glucocorticoids13. Individualized treatments are needed that consider the type, dose, scheme and duration of action of the glucocorticoid used as well as underlying patient comorbidities, concurrent medication use and the severity of hyperglycaemia induced by glucocorticoid administration. Short- and intermediate-acting glucocorticoids (often administered transiently with a high initial dose and tapered over time) can lead to rapid and severe fluctuations in glycaemia, with peaks occurring within 4–8 h of the dose and decreasing during the night29,115. In response to this, hypoglycaemic agents with high potency and rapid onset should be used to avoid the risk of nocturnal hypoglycaemia13,29. For patients on long-acting glucocorticoids or those taking multiple daily doses, hyperglycaemia can persist over 24 h. As such, treatments are favoured that have a long duration of action and enable flexibility in dose adjustments without exacerbating other glucocorticoid-dependent adverse effects148. Treatments should be tailored to the pattern of glucocorticoid-induced hyperglycaemia by considering the type of glucocorticoid and the pharmacokinetics and pharmacodynamics of the different hypoglycaemic agents145.
Non-insulin glucose-lowering therapies
Oral hypoglycaemic drugs are generally reserved for those experiencing mild glucocorticoid-induced hyperglycaemia (fasting plasma concentration of glucose <11.1 mM) without a history of diabetes mellitus or with well-controlled diabetes mellitus13,145. Different hypoglycaemic treatment options have various advantages and disadvantages and considerations for use under different glucocorticoid treatment schemes (Table 1).
Sulfonylureas
Sulfonylureas act to increase insulin release from pancreatic β-cells by binding to and inhibiting ATP-sensitive K+ channels149. Their mechanism and immediate onset of action is an asset as an oral agent for the treatment of glucocorticoid-induced diabetes mellitus150. However, they target fasting plasma levels of glucose rather than postprandial hyperglycaemia and, if used with short-acting glucocorticoids given as a single morning dose, their long effects and narrow therapeutic window result in a high risk of nocturnal hypoglycaemia145. Therefore, sulfonylureas are not recommended for the management of glucocorticoid-induced hyperglycaemia and are only considered for patients treated with intermediate-acting or long-acting glucocorticoid preparations in two or more daily doses or intra-articular glucocorticoids13,38.
Glinides
Glinides are secretagogues that stimulate insulin secretion while exerting a more rapid onset and shorter duration of action than sulfonylureas38. As such, glinides provide improved flexibility in adapting to glucocorticoid-provoked postprandial hyperglycaemia and their use eliminates the possibility of nocturnal hypoglycaemia13. Glinides were shown to effectively manage blood levels of glucose in 14 out of 23 transplant recipients with new-onset diabetes mellitus that occurred after exposure to low dose glucocorticoids in combination with other immunosuppressants151. Retrospective studies further confirmed that glinides improved glucocorticoid-induced postprandial hyperglycaemia regardless of the dose and duration of glucocorticoid treatment (divided dose or high-dose pulse therapy)152,153. However, the frequent required dosing and cost have limited their use and the efficacy of glinides is reduced when insulin resistance is present151,154. Therefore, a combination of glinides with insulin sensitizers (metformin or thiazolidinediones) might be beneficial to control glucocorticoid-induced hyperglycaemia. Interestingly, the insulin-sensitizing thiazolidinediones (described later) induce CYP2C8 and CYP3A4 liver enzyme activity155. If thiazolidinediones are combined with a glinide, this effect might result in a potential drug–drug interaction due to metabolism by the same enzymes156.
Incretin-based therapies
Incretin hormones are peptides released by enteroendocrine cells in the gut following nutrient ingestion that stimulate insulin secretion. Glucagon-like peptide 1 (GLP1) receptor agonists and dipeptidyl peptidase 4 inhibitors (DPP4is) are incretin-based therapies that control hyperglycaemia by enhancing insulin secretion and glucose uptake into peripheral tissues, inhibiting glucagon secretion and decreasing gastric emptying115. In particular, DPP4is enable the release of insulin postprandially by elevating the circulating levels of GLP1, thus specifically targeting postprandial glucose levels38. Indeed, exenatide (a GLP1 receptor agonist) has been reported to efficaciously treat glucocorticoid-evoked postprandial hyperglycaemia in both healthy individuals157 and those with T2DM158. DPP4is are also effective for the treatment of glucocorticoid-induced new-onset diabetes mellitus, yet contradicting reports exist that question their efficacy to prevent glucocorticoid-augmented postprandial hyperglycaemia159,160,161,162. GLP1 receptor agonists are associated with more potent hypoglycaemic effects compared with DPP4is in patients with T2DM163. Of note, injectable GLP1 receptor agonists have substantial gastrointestinal adverse effects, which make them less well tolerated compared with DPP4is154. GLP1 receptor agonists should be used cautiously in patients with glucocorticoid-induced hyperglycaemia after renal transplantation owing to the potential for acute renal injury that is secondary to plasma volume contraction164.
The immediate onset of action of incretin-based therapies without risk of hypoglycaemia and their specificity towards controlling postprandial glucose could enable these drugs to become frontline hypoglycaemics for patients with glucocorticoid-induced diabetes mellitus. They are particularly suitable for patients treated with a single daily dose of intermediate-acting or long-acting glucocorticoids, either transiently or over a long-term period13. GLP1 agonists also have beneficial effects on body weight reduction and blood pressure, which suggests potential additive benefits towards glucocorticoid-induced secondary complications165. However, evidence of incretin-based therapies providing long-term efficacy towards glucocorticoid-induced hyperglycaemia remains unclear. Furthermore, potential relapse might occur as observed in patients with T2DM who were treated with sitagliptin (a DPP4i) after 6 months166. The paucity of evidence confirming the efficacy and safety of the incretin-based therapies in treating glucocorticoid-induced diabetes mellitus has limited their use and thus future studies are essential29.
Insulin-sensitizing therapies
Metformin
Metformin has long been recommended as the first-line therapy for the treatment of T2DM. The drug induces a minimal risk of hypoglycaemia and limits glucocorticoid-induced hepatic and peripheral insulin resistance. Metformin greatly improves hepatic insulin sensitivity by counteracting gluconeogenesis and lipogenesis in the liver while increasing GLP1 secretion and glucose utilization in the gastrointestinal tract, thereby lowering glycaemia167. Despite widespread use in T2DM, evidence of the effectiveness of metformin to counterbalance glucocorticoid treatment is fairly scarce. Nonetheless, glucocorticoid-mediated postprandial hyperglycaemia168 and insulin resistance169 are decreased with metformin use in patients receiving glucocorticoids who do not have diabetes mellitus. Due to its slow onset of action, metformin is considered a favourable option when low doses of intermediate-acting glucocorticoids are administered over the long term13. Metformin remains important for combination therapy with other oral hypoglycaemic drugs because it is not metabolized by cytochrome P450 enzymes, has minimal adverse effects and is low cost170. Of note, metformin is contraindicated in individuals with impaired renal function171, and should therefore be avoided in patients receiving glucocorticoids after renal transplantation.
Thiazolidinediones
Thiazolidinediones are robust insulin sensitizers and therefore affect the action of glucocorticoids on plasma levels of glucose by improving both hepatic and peripheral insulin sensitivity without inducing hypoglycaemia148. In addition to improving insulin sensitivity, thiazolidinediones have the ability to preserve β-cell function172. Similar to metformin, their slow onset of action limits the use of thiazolidinediones for long-term treatment in patients with glucocorticoid-induced hyperglycaemia38. In patients on long-term, low-dose glucocorticoid therapy, 6 months of treatment with pioglitazone statistically significantly reduced plasma glucose concentration 2 h post-OGTT, HbA1c and homeostasis model assessment of insulin resistance (HOMA-IR)173. Thiazolidinediones also improve diabetes-related parameters in patients with severe glucocorticoid-induced hyperglycaemia when administered in combination with insulin174. However, thiazolidinediones are less appealing compared with metformin owing to their overlapping adverse effects with glucocorticoids, including weight gain, increased risk of bone fractures and fluid retention145. Therefore, the benefits must be balanced against their complications when considering thiazolidinediones as treatment options for managing glucocorticoid-induced hyperglycaemia.
Therapies with other mechanisms of action
Sodium–glucose co-transporter type 2 inhibitors
Sodium–glucose co-transporter type 2 inhibitors (SGLT2is) lower plasma levels of glucose by reducing glucose reabsorption in the kidney. Their immediate onset of action without the risk of hypoglycaemia provides a novel approach to minimizing glucocorticoid-associated diabetes mellitus145. Additionally, SGLT2is reduce body weight and cardiovascular risk in patients with T2DM, thus theoretically providing the capacity to combat other adverse effects of glucocorticoids38. However, add-on dapagliflozin to existing hypoglycaemic therapy exerted minimal additional glycaemic control in patients with hyperglycaemia or T2DM when they were challenged with a short-term (5–14 days) high dose of glucocorticoid175. SGLT2is are also associated with risks of urinary tract and genital infections176, and this risk could be exacerbated by glucocorticoid-mediated immunosuppression. The risk of glucocorticoid-induced osteoporosis might also be enhanced due to the potential of SGLT2is to enhance bone resorption177. Finally, whether SGLT2is alone can alleviate glucocorticoid-induced hyperglycaemia remains to be tested.
α-Glucosidase inhibitors
α-Glucosidase inhibitors (AGIs) inhibit the breakdown of complex carbohydrates and delay the absorption of glucose in the gastrointestinal tract, thus reducing postprandial plasma levels of glucose178. As a second-line intervention to metformin, AGIs are mainly prescribed in combination with other hypoglycaemic agents owing to their non-cytochrome P450-dependent metabolism and weak hypoglycaemic effect when administered alone178,179. The combination of glinides and AGIs was shown to improve glucocorticoid-mediated postprandial hyperglycaemia in patients with rheumatoid arthritis152.
Insulin
Insulin therapy should be initiated in those patients who are treated with glucocorticoids and have persistent glycaemic oscillations ≥10 mM as recommended by the ADA143,147. Insulin should also be used when non-insulin hypoglycaemic therapy fails to adequately control glycaemia in patients treated with glucocorticoids who have pre-existing T2DM180. Insulin therapy can efficaciously provide immediate targeting of postprandial hyperglycaemia and flexibility in dosing based on food intake142. Insulin regimens are tailored to glucocorticoid dosing schedules and patient BMI and hyperglycaemic profile181 (Table 2). In patients with pre-existing diabetes mellitus who already require insulin, a 20% increment in daily insulin dose is generally required upon the addition of glucocorticoid therapy in order to achieve similar glycaemic control142. Of note, changes to the glucocorticoid dose might not immediately result in corresponding effects on blood levels of glucose, particularly in patients with pre-existing diabetes mellitus29. As such, the insulin dose used should be carefully titrated to match the glycaemic oscillation of individual patients to provide optimal glucose control.
The use of basal and bolus insulin in combination delivers insulin that most closely resembles its physiological pancreatic release to maintain euglycaemia in both fasting and postprandial states182. For inpatients with severe or persistent hyperglycaemia due to high glucocorticoid doses, multiple daily doses or long-acting glucocorticoid use, basal-bolus insulin should be initiated29,143,148. These regimens offer great flexibility in dose titration, as hospital staff can make adjustments based on preprandial glycaemic reading, anticipated carbohydrate intake and interpatient insulin sensitivity180. However, as multiple daily injections are required of long-acting basal insulin and rapid-acting bolus insulin administered preprandially183, it is impractical for the outpatient with new-onset glucocorticoid-induced hyperglycaemia to manage once discharged. Education on self-injections, stringent glucose monitoring and concern over nocturnal hypoglycaemia can be major drawbacks for insulin therapy, especially if glucocorticoids are required only over a short term154.
For outpatients who are treated with long-term, once-daily glucocorticoids, for whom only postprandial glucose is elevated, neutral protamine Hagedorn (NPH) insulin is a preferable option that can be administered at the same time as their glucocorticoid in the morning13,154. These insulin preparations have a closely aligned temporal profile (peak 4–10 h, duration of action ≥14 h) with the hyperglycaemic excursion induced by intermediate-acting glucocorticoids182. When twice-daily intermediate-acting or long-acting glucocorticoids are administered, the total dose of NPH insulin can be divided13 or substituted for long-acting basal insulin (insulin detemir or glargine), which can adequately match the pattern of glucocorticoid-evoked hyperglycaemia154. Oral hypoglycaemic agents can be added to insulin therapy when patients continue to exhibit severe or persistent hyperglycaemia (HbA1c >9%)13,181. Insulin sensitizers, such as metformin and thiazolidinediones, are the most frequently prescribed in combination with insulin that can provide further glycaemic benefits under chronic glucocorticoid exposure13,182.
Novel pharmacological targets
The unparalleled efficacy of glucocorticoids for the treatment of inflammatory diseases has led to tremendous interest by pharmaceutical companies to develop compounds that could eliminate their diabetogenic effects while preserving their anti-inflammatory functions. Two such approaches, selective glucocorticoid receptor agonists or selective glucocorticoid receptor modulators (Box 3) and 11β-HSD1 inhibitors (Box 4), have made it to clinical trials; however, neither has yet emerged as being clearly superior to glucocorticoids as currently used in practice. The limited dissociative effects of selective glucocorticoid receptor agonists or selective glucocorticoid receptor modulators and the limited efficacy of 11β-HSD1 inhibitors have challenged their continued clinical development as they have not achieved clear superiority over conventional glucocorticoids. Discoveries of novel therapeutic targets that can distinguish the beneficial effects of glucocorticoids from their secondary complications have sparked new interest in the field (Fig. 2).

In the absence of ligand, glucocorticoid receptor resides in the cytoplasm bound to chaperone proteins, including heat shock protein 90 (HSP90) and p23. Histone deacetylase 6 (HDAC6) deacetylates HSP90, thereby enabling its interaction with the co-chaperone p23, which in turn promotes the proper folding of the glucocorticoid receptor. Inhibition of HDAC6 prevents the interaction of HSP90 with p23, inhibiting glucocorticoid receptor translocation. In the presence of glucocorticoids, the glucocorticoid receptor binds to glucocorticoid-response elements (GREs) in genes related to metabolism in target tissues to exert their metabolic effects. Liver X receptor-β (LXRβ), basic helix–loop–helix transcription factor E47 and the glucocorticoid receptor arginine and glutamate-rich protein 1 (ARGLU1) are required for the full recruitment of the glucocorticoid receptor to target genes involved in gluconeogenesis. Loss of any of these factors protects against glucocorticoid-induced hyperglycaemia. Elevated ceramide levels in the portal circulation and the liver, occurring as a result of increased gene and protein expression of serine palmitoyltransferase (SPT) induced by glucocorticoids, promote hepatic gluconeogenesis. Likewise, increased serotonin synthesis (from glucocorticoid-induced TPH1 and AADC) and serotonin receptor expression promote gluconeogenesis and steatosis in liver and lipolysis in adipose tissue. LXRβ also promotes glucocorticoid-induced lipolysis increasing the flux of non-esterified fatty acids and glycerol into the liver, which exacerbates hepatic steatosis. Small-molecule inhibitors (HDAC6, SPT, PKCζ, TPH1 and AADC) or antagonists (LXRβ and serotonin receptor) have been shown to protect against glucocorticoid-induced hyperglycaemia and insulin resistance in mouse models. Thick solid arrows indicate effects; dashed arrows indicate secreted factors.
Liver X receptor-β
As members of the nuclear receptor superfamily of transcription factors, liver X receptor-α (LXRα) and LXRβ control cholesterol homeostasis by sensing intracellular cholesterol load184. Whole-body knockout of the gene encoding LXRβ (Nr1h2) (but not LXRα, encoded by Nr1h3) and/or pharmacological inhibition of LXRβ protects mice against hyperglycaemia, hyperinsulinaemia and hepatic steatosis following glucocorticoid treatment via suppression of genes involved in gluconeogenesis184,185. Notably, the immunosuppressive effects of glucocorticoids are unaltered by LXRβ antagonism. Loss of LXRβ specifically in adipose tissue was reported in a 2021 study to attenuate glucocorticoid-induced lipogenesis and lipolysis in mice and improve systemic insulin tolerance by decreasing NEFA shuttling to the liver186. Limitations of targeting LXRβ to minimize the diabetogenic effects of glucocorticoids include the development of LXRβ isoform-selective inhibitors (owing to high similarity with LXRα) and ensuring no adverse effects occur on whole-body cholesterol efflux (although this function is mainly mediated by LXRα)187.
E47
The basic helix–loop–helix transcription factor E47 is required for efficient recruitment of glucocorticoid receptors and coregulators to a subset of lipid and glucose metabolic gene promoters in the liver188. Loss of E47 specifically from rodent livers abolished the development of glucocorticoid-induced hyperglycaemia and hepatic steatosis188. Both LXRβ and E47 could plausibly synchronously regulate the recruitment of glucocorticoid receptors to target genes involved in gluconeogenesis such as Pck1. Due to the ubiquitous expression of LXRβ and E47, targeting these proteins might necessitate the development of tissue-specific antagonists.
Coregulators of glucocorticoid receptor
The loss of the glucocorticoid receptor coactivator euchromatic histone methyltransferase 2 in the liver was associated with aggravated glucocorticoid-induced hepatic insulin resistance in mice189. Another coactivator of the glucocorticoid receptor, arginine and glutamate-rich protein 1 (ARGLU1), is critical in mediating the recruitment of glucocorticoid receptors to the gluconeogenic Pck1 promoter190. Thus, targeting specific members of the glucocorticoid receptor co-regulator complex remains a fruitful area for future investigation.
Histone deacetylase 6
In the absence of ligand, the glucocorticoid receptor is bound to chaperone proteins in the cytoplasm, including heat shock protein 90 (HSP90)191. Histone deacetylase 6 (HDAC6) deacetylates HSP90, thereby enabling its interaction with the co-chaperone p23, which is essential for the ligand-binding activity of glucocorticoid receptor. Inactivation of HDAC6 results in HSP90 hyperacetylation and decreases ligand binding to the glucocorticoid receptor due to the dissociation of p23 (ref.192). Loss of HDAC6 in mice partially ameliorated glucocorticoid-induced hyperglycaemia and insulin resistance by impairing translocation of glucocorticoid receptors and subsequent glucocorticoid receptor-mediated gluconeogenesis in the liver193. In 2021, HDAC6 was discovered to deacetylate the histone arginine demethylase PRMT5, which inhibits its activity194. As PRMT5 increases chromatin accessibility near gluconeogenic promoters195, this finding could be an additional mechanism by which HDAC6 inhibition constrains glucocorticoid-induced hyperglycaemia.
Serotonin signalling
Plasma levels of serotonin (5-hydroytryptamine or 5-HT) were found to be elevated in patients with T2DM and seemingly contribute to peripheral vascular disease196. In rodents, glucocorticoids can promote insulin resistance by upregulating serotonin synthesis and serotonin receptor (5-HT2R) expression in liver and visceral adipose tissue197. In rats, treatment with a serotonin synthetic inhibitor and/or 5-HT2R antagonist reversed glucocorticoid-mediated hepatic gluconeogenesis, steatosis and insulin resistance198. A key question that remains for therapeutic targeting of the serotonin signalling pathway in this context will be whether inhibition can be achieved without altering serotonin transmission in the central nervous system.
Ceramide synthesis
The ability of glucocorticoids to upregulate the expression of genes involved in ceramide synthesis, such as that encoding serine palmitoyltransferase (Sptlc2), provides a novel approach to protect against glucocorticoid-dependent disruption in glucose homeostasis65. Ceramide-induced activation of protein kinase Cζ (PKCζ) promotes hepatic insulin resistance by inhibiting PKB–AKT downstream of the insulin receptor199. In rodents, pharmacological inhibition of serine palmitoyltransferase by myriocin or PKCζ by 2-acetyl-1,3-cyclopentanedione limits ceramide biosynthesis and signalling, which in turn reduces glucocorticoid-mediated hepatic glucose output and insulin resistance. Despite the inverse correlation between plasma levels of ceramides and improved glucose homeostasis, ceramides have a wide range of biological functions at several cellular growth checkpoints. Therefore, pharmacological targeting of ceramides will first require prudent examination of the specific roles of different ceramide species200.
Conclusions
Glucocorticoids are indispensable for their powerful immunosuppressive and anti-inflammatory effects. However, the adverse metabolic effects of long-term use, including hyperglycaemia and diabetes mellitus, have diminished their therapeutic value. The underlying molecular mechanisms of glucocorticoid-induced hyperglycaemia are complex and involve multiple organs. Organs and tissues either fail to remain responsive to insulin or promote insulin resistance via inter-organ crosstalk, which makes it difficult to predict which patients will develop adverse metabolic effects. This limitation in knowledge has resulted in the poor clinical management of hyperglycaemia and diabetes mellitus as a result of glucocorticoid exposure. Universal guidelines should be established that consider the variations in glucocorticoid therapy, genetic predisposition, comorbidities and therapeutic tolerance of individual patients. The incorporation of patient education with respect to routine glucose monitoring upon the prescription of glucocorticoids will help combat glucocorticoid-induced hyperglycaemia and diabetes mellitus. Future studies that provide a deeper understanding of the mechanism of action of glucocorticoids could lead to the development of novel therapeutic strategies to minimize the diabetogenic effects while sparing the beneficial immunosuppressive effects of glucocorticoids.
References
Sapolsky, R. M., Romero, L. M. & Munck, A. U. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr. Rev. 21, 55–89 (2000).
Hench, P. S. et al. The effect of a hormone of the adrenal cortex (17-hydroxy-11-dehydrocorticosterone; compound E) and of pituitary adrenocorticotropic hormone on rheumatoid arthritis. Proc. Staff. Meet. Mayo Clin. 24, 181–197 (1949).
Cain, D. W. & Cidlowski, J. A. Immune regulation by glucocorticoids. Nat. Rev. Immunol. 17, 233–247 (2017).
van Staa, T. P. et al. Use of oral corticosteroids in the United Kingdom. QJM 93, 105–111 (2000).
Overman, R. A., Yeh, J. Y. & Deal, C. L. Prevalence of oral glucocorticoid usage in the United States: a general population perspective. Arthritis Care Res. 65, 294–298 (2013).
WHO Rapid Evidence Appraisal for COVID-19 Therapies (REACT) Working Group et al. Association between administration of systemic corticosteroids and mortality among critically ill patients with COVID-19: a meta-analysis. JAMA 324, 1330–1341 (2020).
Liu, D. et al. A practical guide to the monitoring and management of the complications of systemic corticosteroid therapy. Allergy Asthma Clin. Immunol. 9, 30 (2013).
van Raalte, D. H. & Diamant, M. Steroid diabetes: from mechanism to treatment? Neth. J. Med. 72, 62–72 (2014).
Gulliford, M. C., Charlton, J. & Latinovic, R. Risk of diabetes associated with prescribed glucocorticoids in a large population. Diabetes Care 29, 2728–2729 (2006).
Rizza, R. A., Mandarino, L. J. & Gerich, J. E. Cortisol-induced insulin resistance in man: impaired suppression of glucose production and stimulation of glucose utilization due to a postreceptor detect of insulin action. J. Clin. Endocrinol. Metab. 54, 131–138 (1982).
Pagano, G. et al. An in vivo and in vitro study of the mechanism of prednisone-induced insulin resistance in healthy subjects. J. Clin. Invest. 72, 1814–1820 (1983).
Liu, X. X. et al. Hyperglycemia induced by glucocorticoids in nondiabetic patients: a meta-analysis. Ann. Nutr. Metab. 65, 324–332 (2014).
Perez, A. et al. Glucocorticoid-induced hyperglycemia. J. Diabetes 6, 9–20 (2014). A thorough guideline that provides insight into the management of glucocorticoid-induced hyperglycaemia, from diagnosis to treatment strategies, based on the pathophysiology observed and dosing regimens of glucocorticoids used.
Newell-Price, J., Bertagna, X., Grossman, A. B. & Nieman, L. K. Cushing’s syndrome. Lancet 367, 1605–1617 (2006).
Scaroni, C., Zilio, M., Foti, M. & Boscaro, M. Glucose metabolism abnormalities in Cushing syndrome: from molecular basis to clinical management. Endocr. Rev. 38, 189–219 (2017).
Heitzer, M. D., Wolf, I. M., Sanchez, E. R., Witchel, S. F. & DeFranco, D. B. Glucocorticoid receptor physiology. Rev. Endocr. Metab. Disord. 8, 321–330 (2007).
Grontved, L. et al. C/EBP maintains chromatin accessibility in liver and facilitates glucocorticoid receptor recruitment to steroid response elements. EMBO J. 32, 1568–1583 (2013).
Uhlenhaut, N. H. et al. Insights into negative regulation by the glucocorticoid receptor from genome-wide profiling of inflammatory cistromes. Mol. Cell 49, 158–171 (2013).
Pasternak, J. J., McGregor, D. G. & Lanier, W. L. Effect of single-dose dexamethasone on blood glucose concentration in patients undergoing craniotomy. J. Neurosurg. Anesthesiol. 16, 122–125 (2004).
Hans, P., Vanthuyne, A., Dewandre, P. Y., Brichant, J. F. & Bonhomme, V. Blood glucose concentration profile after 10 mg dexamethasone in non-diabetic and type 2 diabetic patients undergoing abdominal surgery. Br. J. Anaesth. 97, 164–170 (2006).
Slatore, C. G., Bryson, C. L. & Au, D. H. The association of inhaled corticosteroid use with serum glucose concentration in a large cohort. Am. J. Med. 122, 472–478 (2009).
Gurwitz, J. H. et al. Glucocorticoids and the risk for initiation of hypoglycemic therapy. Arch. Intern. Med. 154, 97–101 (1994).
Donihi, A. C., Raval, D., Saul, M., Korytkowski, M. T. & DeVita, M. A. Prevalence and predictors of corticosteroid-related hyperglycemia in hospitalized patients. Endocr. Pract. 12, 358–362 (2006).
Kim, S. Y. et al. Incidence and risk factors of steroid-induced diabetes in patients with respiratory disease. J. Korean Med. Sci. 26, 264–267 (2011).
Uzu, T. et al. Glucocorticoid-induced diabetes mellitus: prevalence and risk factors in primary renal diseases. Nephron Clin. Pract. 105, c54–c57 (2007).
Harris, D. et al. Glucocorticoid-induced hyperglycemia is prevalent and unpredictable for patients undergoing cancer therapy: an observational cohort study. Curr. Oncol. 20, e532–e538 (2013).
Dean, P. G., Kudva, Y. C., Larson, T. S., Kremers, W. K. & Stegall, M. D. Posttransplant diabetes mellitus after pancreas transplantation. Am. J. Transpl. 8, 175–182 (2008).
Hoes, J. N. et al. Glucose tolerance, insulin sensitivity and beta-cell function in patients with rheumatoid arthritis treated with or without low-to-medium dose glucocorticoids. Ann. Rheum. Dis. 70, 1887–1894 (2011).
Roberts, A., James, J. & Dhatariya, K., Joint British Diabetes Societies for Inpatient Care. Management of hyperglycaemia and steroid (glucocorticoid) therapy: a guideline from the Joint British Diabetes Societies (JBDS) for Inpatient Care group. Diabet. Med. 35, 1011–1017 (2018). A practice guideline from the UK that provides consensus on the management of glucocorticoid-induced hyperglycaemia and diabetes mellitus for patients in hospital and following discharge.
Suissa, S., Kezouh, A. & Ernst, P. Inhaled corticosteroids and the risks of diabetes onset and progression. Am. J. Med. 123, 1001–1006 (2010).
Andersen, Y. M. F. et al. Association between topical corticosteroid use and type 2 diabetes in two European population-based adult cohorts. Diabetes Care 42, 1095–1103 (2019).
Wu, J., Mackie, S. L. & Pujades-Rodriguez, M. Glucocorticoid dose-dependent risk of type 2 diabetes in six immune-mediated inflammatory diseases: a population-based cohort analysis. BMJ Open Diabetes Res. Care 8, e001220 (2020).
Blackburn, D., Hux, J. & Mamdani, M. Quantification of the risk of corticosteroid-induced diabetes mellitus among the elderly. J. Gen. Intern. Med. 17, 717–720 (2002).
Habib, G. & Safia, A. The effect of intra-articular injection of betamethasone acetate/betamethasone sodium phosphate on blood glucose levels in controlled diabetic patients with symptomatic osteoarthritis of the knee. Clin. Rheumatol. 28, 85–87 (2009).
Fleming, P., Drazek, L. & Shaw, J. C. Hyperglycemia following intralesional corticosteroid injection in a patient with type I diabetes mellitus. J. Cutan. Med. Surg. 18, 275–276 (2014).
Hjelmesaeth, J. et al. Glucose intolerance after renal transplantation depends upon prednisolone dose and recipient age. Transplantation 64, 979–983 (1997).
Gonzalez-Gonzalez, J. G. et al. Hyperglycemia related to high-dose glucocorticoid use in noncritically ill patients. Diabetol. Metab. Syndr. 5, 18 (2013).
Nakamura, H., Fujieda, Y., Nakamura, A. & Atsumi, T. How should rheumatologists manage glucocorticoid-induced hyperglycemia? Mod. Rheumatol. 31, 519–528 (2021). Meta-analysis that provides innovative insight into the management of glucocorticoid-induced hyperglycaemia from the point of view of rheumatology.
Shaharir, S. S., Gafor, A. H., Said, M. S. & Kong, N. C. Steroid-induced diabetes mellitus in systemic lupus erythematosus patients: analysis from a Malaysian multi-ethnic lupus cohort. Int. J. Rheum. Dis. 18, 541–547 (2015).
Wajngot, A., Giacca, A., Grill, V., Vranic, M. & Efendic, S. The diabetogenic effects of glucocorticoids are more pronounced in low- than in high-insulin responders. Proc. Natl Acad. Sci. USA 89, 6035–6039 (1992).
Larsson, H. & Ahren, B. Insulin resistant subjects lack islet adaptation to short-term dexamethasone-induced reduction in insulin sensitivity. Diabetologia 42, 936–943 (1999).
Braithwaite, S. S., Barr, W. G., Rahman, A. & Quddusi, S. Managing diabetes during glucocorticoid therapy. How to avoid metabolic emergencies. Postgrad. Med. https://doi.org/10.3810/pgm.1998.11.426 (1998).
Yang, X. et al. Fasting plasma glucose levels predict steroid-induced abnormal glucose metabolism in patients with non-diabetic chronic kidney disease: a prospective cohort study. Am. J. Nephrol. 41, 107–115 (2015).
Katsuyama, T. et al. Risk factors for the development of glucocorticoid-induced diabetes mellitus. Diabetes Res. Clin. Pract. 108, 273–279 (2015).
Depczynski, B., Daly, B., Campbell, L. V., Chisholm, D. J. & Keogh, A. Predicting the occurrence of diabetes mellitus in recipients of heart transplants. Diabet. Med. 17, 15–19 (2000).
Henriksen, J. E., Alford, F., Ward, G. M. & Beck-Nielsen, H. Risk and mechanism of dexamethasone-induced deterioration of glucose tolerance in non-diabetic first-degree relatives of NIDDM patients. Diabetologia 40, 1439–1448 (1997).
Miyawaki, Y. et al. A retrospective observational study of glucocorticoid-induced diabetes mellitus with IgA nephropathy treated with tonsillectomy plus methylprednisolone pulse therapy. PLoS One 12, e0178018 (2017).
Simmons, L. R., Molyneaux, L., Yue, D. K. & Chua, E. L. Steroid-induced diabetes: is it just unmasking of type 2 diabetes? ISRN Endocrinol. 2012, 910905 (2012).
Hu, W. et al. Individual-specific functional epigenomics reveals genetic determinants of adverse metabolic effects of glucocorticoids. Cell Metab. 33, 1592–1609 (2021). This study discovers genetic variations within the population in predicting individual-specific metabolic responses to glucocorticoid therapy, including hyperglycaemia and diabetes mellitus.
Ha, Y. et al. Glucocorticoid-induced diabetes mellitus in patients with systemic lupus erythematosus treated with high-dose glucocorticoid therapy. Lupus 20, 1027–1034 (2011).
Frazier, B., Hsiao, C. W., Deuster, P. & Poth, M. African Americans and Caucasian Americans: differences in glucocorticoid-induced insulin resistance. Horm. Metab. Res. 42, 887–891 (2010).
Binnert, C., Ruchat, S., Nicod, N. & Tappy, L. Dexamethasone-induced insulin resistance shows no gender difference in healthy humans. Diabetes Metab. 30, 321–326 (2004).
Vegiopoulos, A. & Herzig, S. Glucocorticoids, metabolism and metabolic diseases. Mol. Cell. Endocrinol. 275, 43–61 (2007). This review discusses all aspects of whole-body and tissue-specific glucocorticoid action, much of which is still not fully understood today.
Schakman, O., Gilson, H. & Thissen, J. P. Mechanisms of glucocorticoid-induced myopathy. J. Endocrinol. 197, 1–10 (2008). A thorough discussion on the muscle atrophic action of glucocorticoids and therapeutic approaches that prevent the effects of glucocorticoid-induced atrophy.
Xu, C. et al. Direct effect of glucocorticoids on lipolysis in adipocytes. Mol. Endocrinol. 23, 1161–1170 (2009).
Imai, E. et al. Characterization of a complex glucocorticoid response unit in the phosphoenolpyruvate carboxykinase gene. Mol. Cell. Biol. 10, 4712–4719 (1990).
Vander Kooi, B. T. et al. The glucose-6-phosphatase catalytic subunit gene promoter contains both positive and negative glucocorticoid response elements. Mol. Endocrinol. 19, 3001–3022 (2005).
Puigserver, P. et al. Insulin-regulated hepatic gluconeogenesis through FOXO1-PGC-1alpha interaction. Nature 423, 550–555 (2003).
Hall, R. K., Wang, X. L., George, L., Koch, S. R. & Granner, D. K. Insulin represses phosphoenolpyruvate carboxykinase gene transcription by causing the rapid disruption of an active transcription complex: a potential epigenetic effect. Mol. Endocrinol. 21, 550–563 (2007).
Andrews, R. C. & Walker, B. R. Glucocorticoids and insulin resistance: old hormones, new targets. Clin. Sci. 96, 513–523 (1999).
Rojas, F. A., Hirata, A. E. & Saad, M. J. Regulation of insulin receptor substrate-2 tyrosine phosphorylation in animal models of insulin resistance. Endocrine 21, 115–122 (2003).
Titchenell, P. M., Lazar, M. A. & Birnbaum, M. J. Unraveling the regulation of hepatic metabolism by insulin. Trends Endocrinol. Metab. 28, 497–505 (2017).
Rahimi, L., Rajpal, A. & Ismail-Beigi, F. Glucocorticoid-induced fatty liver disease. Diabetes Metab. Syndr. Obes. 13, 1133–1145 (2020).
Samuel, V. T. & Shulman, G. I. The pathogenesis of insulin resistance: integrating signaling pathways and substrate flux. J. Clin. Invest. 126, 12–22 (2016).
Holland, W. L. et al. Inhibition of ceramide synthesis ameliorates glucocorticoid-, saturated-fat-, and obesity-induced insulin resistance. Cell Metab. 5, 167–179 (2007). Key study identifying the contribution of ceramide to glucocorticoid-induced insulin resistance, which highlights the potential of targeting ceramide pharmacologically to eliminate glucocorticoid-dependent disruption in glucose homeostasis.
DeFronzo, R. A. et al. The effect of insulin on the disposal of intravenous glucose. Results from indirect calorimetry and hepatic and femoral venous catheterization. Diabetes 30, 1000–1007 (1981).
Giorgino, F., Almahfouz, A., Goodyear, L. J. & Smith, R. J. Glucocorticoid regulation of insulin receptor and substrate IRS-1 tyrosine phosphorylation in rat skeletal muscle in vivo. J. Clin. Invest. 91, 2020–2030 (1993).
Saad, M. J., Folli, F., Kahn, J. A. & Kahn, C. R. Modulation of insulin receptor, insulin receptor substrate-1, and phosphatidylinositol 3-kinase in liver and muscle of dexamethasone-treated rats. J. Clin. Invest. 92, 2065–2072 (1993).
Ruzzin, J., Wagman, A. S. & Jensen, J. Glucocorticoid-induced insulin resistance in skeletal muscles: defects in insulin signalling and the effects of a selective glycogen synthase kinase-3 inhibitor. Diabetologia 48, 2119–2130 (2005).
Weinstein, S. P., Wilson, C. M., Pritsker, A. & Cushman, S. W. Dexamethasone inhibits insulin-stimulated recruitment of GLUT4 to the cell surface in rat skeletal muscle. Metabolism 47, 3–6 (1998).
Kuo, T. et al. Genome-wide analysis of glucocorticoid receptor-binding sites in myotubes identifies gene networks modulating insulin signaling. Proc. Natl Acad. Sci. USA 109, 11160–11165 (2012).
Ekstrand, A. et al. The effect of (steroid) immunosuppression on skeletal muscle glycogen metabolism in patients after kidney transplantation. Transplantation 61, 889–893 (1996).
Gupta, A. & Gupta, Y. Glucocorticoid-induced myopathy: pathophysiology, diagnosis, and treatment. Indian J. Endocrinol. Metab. 17, 913–916 (2013).
van Raalte, D. H., Ouwens, D. M. & Diamant, M. Novel insights into glucocorticoid-mediated diabetogenic effects: towards expansion of therapeutic options? Eur. J. Clin. Invest. 39, 81–93 (2009). Seminal review that provides a thorough overview of the potential pathophysiological pathways contributing to the diabetogenic effects induced by glucocorticoids.
Sandri, M. et al. Foxo transcription factors induce the atrophy-related ubiquitin ligase atrogin-1 and cause skeletal muscle atrophy. Cell 117, 399–412 (2004).
Bodine, S. C. et al. Identification of ubiquitin ligases required for skeletal muscle atrophy. Science 294, 1704–1708 (2001).
Lofberg, E. et al. Effects of high doses of glucocorticoids on free amino acids, ribosomes and protein turnover in human muscle. Eur. J. Clin. Invest. 32, 345–353 (2002).
Short, K. R., Bigelow, M. L. & Nair, K. S. Short-term prednisone use antagonizes insulin’s anabolic effect on muscle protein and glucose metabolism in young healthy people. Am. J. Physiol. Endocrinol. Metab. 297, E1260–E1268 (2009).
Krebs, M. et al. Mechanism of amino acid-induced skeletal muscle insulin resistance in humans. Diabetes 51, 599–605 (2002).
Boden, G. & Shulman, G. I. Free fatty acids in obesity and type 2 diabetes: defining their role in the development of insulin resistance and beta-cell dysfunction. Eur. J. Clin. Invest. 32 (Suppl. 3), 14–23 (2002).
Ross, E. J. & Linch, D. C. Cushing’s syndrome–killing disease: discriminatory value of signs and symptoms aiding early diagnosis. Lancet 2, 646–649 (1982).
Wang, J. C., Gray, N. E., Kuo, T. & Harris, C. A. Regulation of triglyceride metabolism by glucocorticoid receptor. Cell Biosci. 2, 19 (2012).
Westphal, S. A. Obesity, abdominal obesity, and insulin resistance. Clin. Cornerstone 9, 23–29 (2008).
Lee, M. J., Pramyothin, P., Karastergiou, K. & Fried, S. K. Deconstructing the roles of glucocorticoids in adipose tissue biology and the development of central obesity. Biochim. Biophys. Acta 1842, 473–481 (2014). An extensive review that discusses all aspects of glucocorticoid-mediated dysfunction in adipose tissue to aid current understanding of the contribution of central adipose deposition in the development of glucocorticoid-induced diabetes mellitus and insulin resistance.
Yu, C. Y. et al. Genome-wide analysis of glucocorticoid receptor binding regions in adipocytes reveal gene network involved in triglyceride homeostasis. PLoS One 5, e15188 (2010).
Ma, X., Xu, L. & Mueller, E. Forkhead box A3 mediates glucocorticoid receptor function in adipose tissue. Proc. Natl Acad. Sci. USA 113, 3377–3382 (2016).
Strawford, A., Antelo, F., Christiansen, M. & Hellerstein, M. K. Adipose tissue triglyceride turnover, de novo lipogenesis, and cell proliferation in humans measured with 2H2O. Am. J. Physiol. Endocrinol. Metab. 286, E577–E588 (2004).
Gathercole, L. L. et al. Regulation of lipogenesis by glucocorticoids and insulin in human adipose tissue. PLoS One 6, e26223 (2011).
Harris, C. et al. Large increases in adipose triacylglycerol flux in Cushingoid CRH-Tg mice are explained by futile cycling. Am. J. Physiol. Endocrinol. Metab. 304, E282–E293 (2013).
Samra, J. S. et al. Effects of physiological hypercortisolemia on the regulation of lipolysis in subcutaneous adipose tissue. J. Clin. Endocrinol. Metab. 83, 626–631 (1998).
Djurhuus, C. B. et al. Effects of cortisol on lipolysis and regional interstitial glycerol levels in humans. Am. J. Physiol. Endocrinol. Metab. 283, E172–E177 (2002).
Buren, J., Liu, H. X., Jensen, J. & Eriksson, J. W. Dexamethasone impairs insulin signalling and glucose transport by depletion of insulin receptor substrate-1, phosphatidylinositol 3-kinase and protein kinase B in primary cultured rat adipocytes. Eur. J. Endocrinol. 146, 419–429 (2002).
Sakoda, H. et al. Dexamethasone-induced insulin resistance in 3T3-L1 adipocytes is due to inhibition of glucose transport rather than insulin signal transduction. Diabetes 49, 1700–1708 (2000).
Gathercole, L. L., Bujalska, I. J., Stewart, P. M. & Tomlinson, J. W. Glucocorticoid modulation of insulin signaling in human subcutaneous adipose tissue. J. Clin. Endocrinol. Metab. 92, 4332–4339 (2007).
Gathercole, L. L., Morgan, S. A., Bujalska, I. J., Stewart, P. M. & Tomlinson, J. W. Short- and long-term glucocorticoid treatment enhances insulin signalling in human subcutaneous adipose tissue. Nutr. Diabetes 1, e3 (2011).
Lundgren, M., Buren, J., Ruge, T., Myrnas, T. & Eriksson, J. W. Glucocorticoids down-regulate glucose uptake capacity and insulin-signaling proteins in omental but not subcutaneous human adipocytes. J. Clin. Endocrinol. Metab. 89, 2989–2997 (2004).
Patsouris, D. et al. Glucocorticoids and thiazolidinediones interfere with adipocyte-mediated macrophage chemotaxis and recruitment. J. Biol. Chem. 284, 31223–31235 (2009).
Do, T. T. H. et al. Glucocorticoid-induced insulin resistance is related to macrophage visceral adipose tissue infiltration. J. Steroid Biochem. Mol. Biol. 185, 150–162 (2019).
Harms, M. & Seale, P. Brown and beige fat: development, function and therapeutic potential. Nat. Med. 19, 1252–1263 (2013).
Poggioli, R. et al. Dexamethasone reduces energy expenditure and increases susceptibility to diet-induced obesity in mice. Obesity 21, E415–E420 (2013).
Kong, X. et al. Glucocorticoids transcriptionally regulate miR-27b expression promoting body fat accumulation via suppressing the browning of white adipose tissue. Diabetes 64, 393–404 (2015).
Liu, J. et al. Essential roles of 11beta-HSD1 in regulating brown adipocyte function. J. Mol. Endocrinol. 50, 103–113 (2013).
Luijten, I. H. N., Cannon, B. & Nedergaard, J. Glucocorticoids and brown adipose tissue: do glucocorticoids really inhibit thermogenesis? Mol. Asp. Med. 68, 42–59 (2019).
Schnabl, K., Westermeier, J., Li, Y. & Klingenspor, M. Opposing actions of adrenocorticotropic hormone and glucocorticoids on UCP1-mediated respiration in brown adipocytes. Front. Physiol. 9, 1931 (2018).
Ramage, L. E. et al. Glucocorticoids acutely increase brown adipose tissue activity in humans, revealing species-specific differences in UCP-1 regulation. Cell Metab. 24, 130–141 (2016).
Fazeli, P. K. et al. Marrow fat and bone — new perspectives. J. Clin. Endocrinol. Metab. 98, 935–945 (2013).
Ermetici, F. et al. Bone marrow fat contributes to insulin sensitivity and adiponectin secretion in premenopausal women. Endocrine 59, 410–418 (2018).
Pham, T. T. et al. Human bone marrow adipose tissue is a metabolically active and insulin-sensitive distinct fat depot. J. Clin. Endocrinol. Metab. 105, 2300–2310 (2020).
Suchacki, K. J. et al. Bone marrow adipose tissue is a unique adipose subtype with distinct roles in glucose homeostasis. Nat. Commun. 11, 3097 (2020).
Kahn, S. E. The relative contributions of insulin resistance and beta-cell dysfunction to the pathophysiology of type 2 diabetes. Diabetologia 46, 3–19 (2003).
Rafacho, A., Ortsater, H., Nadal, A. & Quesada, I. Glucocorticoid treatment and endocrine pancreas function: implications for glucose homeostasis, insulin resistance and diabetes. J. Endocrinol. 223, R49–R62 (2014).
Linssen, M. M. et al. Prednisolone-induced beta cell dysfunction is associated with impaired endoplasmic reticulum homeostasis in INS-1E cells. Cell. Signal. 23, 1708–1715 (2011).
Fichna, M. & Fichna, P. Glucocorticoids and beta-cell function. Endokrynol. Pol. 68, 568–573 (2017).
Rorsman, P. et al. The cell physiology of biphasic insulin secretion. N. Physiol. Sci. 15, 72–77 (2000).
Bonaventura, A. & Montecucco, F. Steroid-induced hyperglycemia: an underdiagnosed problem or clinical inertia? A narrative review. Diabetes Res. Clin. Pract. 139, 203–220 (2018).
Kuo, T., McQueen, A., Chen, T. C. & Wang, J. C. Regulation of glucose homeostasis by glucocorticoids. Adv. Exp. Med. Biol. 872, 99–126 (2015).
Sato, S. et al. Effects of glucocorticoid treatment on beta- and alpha-cell mass in Japanese adults with and without diabetes. Diabetes 64, 2915–2927 (2015).
van Raalte, D. H. et al. Acute and 2-week exposure to prednisolone impair different aspects of beta-cell function in healthy men. Eur. J. Endocrinol. 162, 729–735 (2010).
Rafacho, A. et al. Glucocorticoids in vivo induce both insulin hypersecretion and enhanced glucose sensitivity of stimulus-secretion coupling in isolated rat islets. Endocrinology 151, 85–95 (2010).
Rafacho, A., Cestari, T. M., Taboga, S. R., Boschero, A. C. & Bosqueiro, J. R. High doses of dexamethasone induce increased beta-cell proliferation in pancreatic rat islets. Am. J. Physiol. Endocrinol. Metab. 296, E681–E689 (2009).
Besse, C., Nicod, N. & Tappy, L. Changes in insulin secretion and glucose metabolism induced by dexamethasone in lean and obese females. Obes. Res. 13, 306–311 (2005).
Ogawa, A. et al. Roles of insulin resistance and beta-cell dysfunction in dexamethasone-induced diabetes. J. Clin. Invest. 90, 497–504 (1992).
Dessein, P. H. & Joffe, B. I. Insulin resistance and impaired beta cell function in rheumatoid arthritis. Arthritis Rheum. 54, 2765–2775 (2006).
Kamba, A. et al. Association between higher serum cortisol levels and decreased insulin secretion in a general population. PLoS One 11, e0166077 (2016).
Giordano, C. et al. Is diabetes in Cushing’s syndrome only a consequence of hypercortisolism? Eur. J. Endocrinol. 170, 311–319 (2014).
Weinstein, R. S. Clinical practice. Glucocorticoid-induced bone disease. N. Engl. J. Med. 365, 62–70 (2011).
Chotiyarnwong, P. & McCloskey, E. V. Pathogenesis of glucocorticoid-induced osteoporosis and options for treatment. Nat. Rev. Endocrinol. 16, 437–447 (2020).
Chen, S. M. et al. Dexamethasone down-regulates osteocalcin in bone cells through leptin pathway. Int. J. Med. Sci. 15, 507–516 (2018).
Lee, N. K. et al. Endocrine regulation of energy metabolism by the skeleton. Cell 130, 456–469 (2007).
Bao, Y. Q. et al. Relationship between serum osteocalcin and glycaemic variability in type 2 diabetes. Clin. Exp. Pharmacol. Physiol. 38, 50–54 (2011).
Parker, L. et al. Glucocorticoid-induced insulin resistance in men is associated with suppressed undercarboxylated osteocalcin. J. Bone Miner. Res. 34, 49–58 (2019).
Brennan-Speranza, T. C. et al. Osteoblasts mediate the adverse effects of glucocorticoids on fuel metabolism. J. Clin. Invest. 122, 4172–4189 (2012).
Sominsky, L. & Spencer, S. J. Eating behavior and stress: a pathway to obesity. Front. Psychol. 5, 434 (2014).
Perry, R. J. et al. Leptin’s hunger-suppressing effects are mediated by the hypothalamic-pituitary-adrenocortical axis in rodents. Proc. Natl Acad. Sci. USA 116, 13670–13679 (2019).
Shimizu, H. et al. Glucocorticoids increase neuropeptide Y and agouti-related peptide gene expression via adenosine monophosphate-activated protein kinase signaling in the arcuate nucleus of rats. Endocrinology 149, 4544–4553 (2008).
Yi, C. X. et al. Glucocorticoid signaling in the arcuate nucleus modulates hepatic insulin sensitivity. Diabetes 61, 339–345 (2012).
Udden, J. et al. Effects of glucocorticoids on leptin levels and eating behaviour in women. J. Intern. Med. 253, 225–231 (2003).
Xu, J. et al. Genetic identification of leptin neural circuits in energy and glucose homeostases. Nature 556, 505–509 (2018).
Ishida-Takahashi, R. et al. Rapid inhibition of leptin signaling by glucocorticoids in vitro and in vivo. J. Biol. Chem. 279, 19658–19664 (2004).
Pecoraro, N., Reyes, F., Gomez, F., Bhargava, A. & Dallman, M. F. Chronic stress promotes palatable feeding, which reduces signs of stress: feedforward and feedback effects of chronic stress. Endocrinology 145, 3754–3762 (2004).
Epel, E., Lapidus, R., McEwen, B. & Brownell, K. Stress may add bite to appetite in women: a laboratory study of stress-induced cortisol and eating behavior. Psychoneuroendocrinology 26, 37–49 (2001).
Mills, E. & Devendra, S. Steroid-induced hyperglycaemia in primary care. Lond. J. Prim. Care 7, 103–106 (2015).
Umpierrez, G. E. et al. Management of hyperglycemia in hospitalized patients in non-critical care setting: an endocrine society clinical practice guideline. J. Clin. Endocrinol. Metab. 97, 16–38 (2012).
American Diabetes Association Professional Practice Committee. 2. Classification and diagnosis of diabetes: standards of medical care in diabetes-2022. Diabetes Care 45 (Suppl. 1), S17–S38 (2022).
Tamez-Perez, H. E., Quintanilla-Flores, D. L., Rodriguez-Gutierrez, R., Gonzalez-Gonzalez, J. G. & Tamez-Pena, A. L. Steroid hyperglycemia: prevalence, early detection and therapeutic recommendations: a narrative review. World J. Diabetes 6, 1073–1081 (2015).
Burt, M. G. et al. Screening for diabetes in patients with inflammatory rheumatological disease administered long-term prednisolone: a cross-sectional study. Rheumatology 51, 1112–1119 (2012).
American Diabetes Association Professional Practice Committee et al. 16. Diabetes care in the hospital: standards of medical care in diabetes-2022. Diabetes Care 45 (Suppl. 1), S244–S253 (2022).
Radhakutty, A. & Burt, M. G. Management of endocrine disease: critical review of the evidence underlying management of glucocorticoid-induced hyperglycaemia. Eur. J. Endocrinol. 179, R207–R218 (2018). In-depth discussion of how hypoglycaemic therapies (insulin sensitizers and insulin) should be tailored towards the distinct pattern of glucocorticoid-induced hyperglycaemia.
Proks, P., Reimann, F., Green, N., Gribble, F. & Ashcroft, F. Sulfonylurea stimulation of insulin secretion. Diabetes 51 (Suppl. 3), S368–S376 (2002).
Kasayama, S., Tanaka, T., Hashimoto, K., Koga, M. & Kawase, I. Efficacy of glimepiride for the treatment of diabetes occurring during glucocorticoid therapy. Diabetes Care 25, 2359–2360 (2002).
Turk, T. et al. Repaglinide in the management of new-onset diabetes mellitus after renal transplantation. Am. J. Transplant. 6, 842–846 (2006).
Ito, S. et al. Early diagnosis and treatment of steroid-induced diabetes mellitus in patients with rheumatoid arthritis and other connective tissue diseases. Mod. Rheumatol. 24, 52–59 (2014).
Tanaka, K. et al. The effects of mitiglinide and repaglinide on postprandial hyperglycemia in patients undergoing methylprednisolone pulse therapy. Intern. Med. 57, 65–70 (2018).
Wallace, M. D. & Metzger, N. L. Optimizing the treatment of steroid-induced hyperglycemia. Ann. Pharmacother. 52, 86–90 (2018).
Sahi, J. et al. Comparative effects of thiazolidinediones on in vitro P450 enzyme induction and inhibition. Drug Metab. Dispos. 31, 439–446 (2003).
Bidstrup, T. B., Bjornsdottir, I., Sidelmann, U. G., Thomsen, M. S. & Hansen, K. T. CYP2C8 and CYP3A4 are the principal enzymes involved in the human in vitro biotransformation of the insulin secretagogue repaglinide. Br. J. Clin. Pharmacol. 56, 305–314 (2003).
van Raalte, D. H., van Genugten, R. E., Linssen, M. M., Ouwens, D. M. & Diamant, M. Glucagon-like peptide-1 receptor agonist treatment prevents glucocorticoid-induced glucose intolerance and islet-cell dysfunction in humans. Diabetes Care 34, 412–417 (2011).
Matsuo, K. et al. Evaluation of the effects of exenatide administration in patients with type 2 diabetes with worsened glycemic control caused by glucocorticoid therapy. Intern. Med. 52, 89–95 (2013).
Ohashi, N. et al. Alogliptin improves steroid-induced hyperglycemia in treatment-naive Japanese patients with chronic kidney disease by decrease of plasma glucagon levels. Med. Sci. Monit. 20, 587–593 (2014).
Yata, Y. et al. The assessment of the efficacy of dipeptidyl peptidase-4 inhibitors in patients with glucocorticoid-induced diabetes by continuous glucose monitoring. Intern. Med. 56, 2555–2562 (2017).
van Genugten, R. E. et al. Does dipeptidyl peptidase-4 inhibition prevent the diabetogenic effects of glucocorticoids in men with the metabolic syndrome? A randomized controlled trial. Eur. J. Endocrinol. 170, 429–439 (2014).
Miyawaki, Y. et al. An open-label pilot study on preventing glucocorticoid-induced diabetes mellitus with linagliptin. J. Med. Case Rep. 12, 288 (2018).
DeFronzo, R. A. et al. Effects of exenatide versus sitagliptin on postprandial glucose, insulin and glucagon secretion, gastric emptying, and caloric intake: a randomized, cross-over study. Curr. Med. Res. Opin. 24, 2943–2952 (2008).
Montada-Atin, T. & Prasad, G. V. R. Recent advances in new-onset diabetes mellitus after kidney transplantation. World J. Diabetes 12, 541–555 (2021).
Robinson, L. E., Holt, T. A., Rees, K., Randeva, H. S. & O’Hare, J. P. Effects of exenatide and liraglutide on heart rate, blood pressure and body weight: systematic review and meta-analysis. BMJ Open 3, e001986 (2013).
Tajiri, Y. et al. Long-term efficacy of sitagliptin for the treatment of type 2 diabetic patients in Japan. Endocr. J. 59, 197–204 (2012).
Rena, G., Hardie, D. G. & Pearson, E. R. The mechanisms of action of metformin. Diabetologia 60, 1577–1585 (2017).
Ochola, L. A., Nyamu, D. G., Guantai, E. M. & Weru, I. W. Metformin’s effectiveness in preventing prednisone-induced hyperglycemia in hematological cancers. J. Oncol. Pharm. Pract. 26, 823–834 (2020).
Pernicova, I. et al. Metformin to reduce metabolic complications and inflammation in patients on systemic glucocorticoid therapy: a randomised, double-blind, placebo-controlled, proof-of-concept, phase 2 trial. Lancet Diabetes Endocrinol. 8, 278–291 (2020).
Holstein, A. & Beil, W. Oral antidiabetic drug metabolism: pharmacogenomics and drug interactions. Expert Opin. Drug Metab. Toxicol. 5, 225–241 (2009).
Inzucchi, S. E., Lipska, K. J., Mayo, H., Bailey, C. J. & McGuire, D. K. Metformin in patients with type 2 diabetes and kidney disease: a systematic review. JAMA 312, 2668–2675 (2014).
Defronzo, R. A. Banting Lecture. From the triumvirate to the ominous octet: a new paradigm for the treatment of type 2 diabetes mellitus. Diabetes 58, 773–795 (2009).
Yamamoto, S., Fujimoto., H., Nakao, H., Kato, T. & Morimoto, I. Effects of pioglitazone on steroid-induced diabetes mellitus. J. Jpn. Diab. Soc. 47, 6 (2004).
Willi, S. M. et al. Effective use of thiazolidinediones for the treatment of glucocorticoid-induced diabetes. Diabetes Res. Clin. Pract. 58, 87–96 (2002).
Gerards, M. C. et al. Dapagliflozin for prednisone-induced hyperglycaemia in acute exacerbation of chronic obstructive pulmonary disease. Diabetes Obes. Metab. 20, 1306–1310 (2018).
Donnan, J. R. et al. Comparative safety of the sodium glucose co-transporter 2 (SGLT2) inhibitors: a systematic review and meta-analysis. BMJ Open 9, e022577 (2019).
Taylor, S. I., Blau, J. E. & Rother, K. I. Possible adverse effects of SGLT2 inhibitors on bone. Lancet Diabetes Endocrinol. 3, 8–10 (2015).
Hedrington, M. S. & Davis, S. N. Considerations when using alpha-glucosidase inhibitors in the treatment of type 2 diabetes. Expert Opin. Pharmacother. 20, 2229–2235 (2019).
Ranchon, F. et al. Concomitant drugs with low risks of drug-drug interactions for use in oncology clinical trials. Crit. Rev. Oncol. Hematol. 94, 189–200 (2015).
Handelsman, Y. et al. American Association of Clinical endocrinologists and American College of endocrinology — clinical practice guidelines for developing a diabetes mellitus comprehensive care plan — 2015 — Executive summary. Endocr. Pract. 21, 413–437 (2015).
Clore, J. N. & Thurby-Hay, L. Glucocorticoid-induced hyperglycemia. Endocr. Pract. 15, 469–474 (2009).
Petznick, A. Insulin management of type 2 diabetes mellitus. Am. Fam. Physician 84, 183–190 (2011).
Candido, R., Wyne, K. & Romoli, E. A review of basal-bolus therapy using insulin glargine and insulin lispro in the management of diabetes mellitus. Diabetes Ther. 9, 927–949 (2018).
Patel, R. et al. LXRbeta is required for glucocorticoid-induced hyperglycemia and hepatosteatosis in mice. J. Clin. Invest. 121, 431–441 (2011).
Patel, R. et al. Separating the anti-inflammatory and diabetogenic effects of glucocorticoids through LXRbeta antagonism. Endocrinology 158, 1034–1047 (2017).
Li, J. X. & Cummins, C. L. Disruption of adipose tissue metabolism by glucocorticoids is attenuated with LXRβ antagonism. J. Endocr. Soc. 5, A821–A822 (2021).
Alberti, S. et al. Hepatic cholesterol metabolism and resistance to dietary cholesterol in LXRbeta-deficient mice. J. Clin. Invest. 107, 565–573 (2001).
Hemmer, M. C. et al. E47 modulates hepatic glucocorticoid action. Nat. Commun. 10, 306 (2019).
Lee, R. A., Tsay, A., Chang, M., Li, D. & Wang, J. The transcriptional coactivation function of EHMT2 restricts chronic glucocorticoid exposure induced insulin resistance. J. Endocr. Soc. 4, A596 (2020).
Magomedova, L. Identification and Characterization of ARGLU1, a Novel Glucocorticoid Receptor Coactivator (University of Toronto, 2016).
Pratt, W. B., Morishima, Y., Murphy, M. & Harrell, M. Chaperoning of glucocorticoid receptors. Handb. Exp. Pharmacol. 172, 111–138 (2006).
Kovacs, J. J. et al. HDAC6 regulates Hsp90 acetylation and chaperone-dependent activation of glucocorticoid receptor. Mol. Cell 18, 601–607 (2005).
Winkler, R. et al. Histone deacetylase 6 (HDAC6) is an essential modifier of glucocorticoid-induced hepatic gluconeogenesis. Diabetes 61, 513–523 (2012).
Gomes, I. D., Ariyaratne, U. V. & Pflum, M. K. H. HDAC6 substrate discovery using proteomics-based substrate trapping: HDAC6 deacetylates PRMT5 to influence methyltransferase activity. ACS Chem. Biol. 16, 1435–1444 (2021).
Tsai, W. W. et al. PRMT5 modulates the metabolic response to fasting signals. Proc. Natl Acad. Sci. USA 110, 8870–8875 (2013).
Barradas, M. A., Gill, D. S., Fonseca, V. A., Mikhailidis, D. P. & Dandona, P. Intraplatelet serotonin in patients with diabetes mellitus and peripheral vascular disease. Eur. J. Clin. Invest. 18, 399–404 (1988).
Li, T. et al. Important role of 5-hydroxytryptamine in glucocorticoid-induced insulin resistance in liver and intra-abdominal adipose tissue of rats. J. Diabetes Investig. 7, 32–41 (2016).
Ma, S. et al. Effective treatment with combination of peripheral 5-hydroxytryptamine synthetic inhibitor and 5-hydroxytryptamine 2 receptor antagonist on glucocorticoid-induced whole-body insulin resistance with hyperglycemia. J. Diabetes Investig. 7, 833–844 (2016).
Chen, T. C. et al. The glucocorticoid-Angptl4-ceramide axis induces insulin resistance through PP2A and PKCzeta. Sci. Signal. 10, eaai7905 (2017).
Sokolowska, E. & Blachnio-Zabielska, A. The role of ceramides in insulin resistance. Front. Endocrinol. 10, 577 (2019).
Uchinuma, H., Ichijo, M., Harima, N. & Tsuchiya, K. Dulaglutide improves glucocorticoid-induced hyperglycemia in inpatient care and reduces dose and injection frequency of insulin. BMC Endocr. Disord. 20, 58 (2020).
Seelig, E. et al. Metformin prevents metabolic side effects during systemic glucocorticoid treatment. Eur. J. Endocrinol. 176, 349–358 (2017).
The RECOVERY Collaborative Group. Dexamethasone in hospitalized patients with Covid-19. N. Engl. J. Med. 384, 693–704 (2021).
Xie, Y. & Al-Aly, Z. Risks and burdens of incident diabetes in long COVID: a cohort study. Lancet Diabetes Endocrinol. https://doi.org/10.1016/S2213-8587(22)00044-4 (2022).
Gianchandani, R. et al. Managing hyperglycemia in the COVID-19 inflammatory storm. Diabetes 69, 2048–2053 (2020).
Lim, S., Bae, J. H., Kwon, H. S. & Nauck, M. A. COVID-19 and diabetes mellitus: from pathophysiology to clinical management. Nat. Rev. Endocrinol. 17, 11–30 (2021).
Kow, C. S. & Hasan, S. S. Corticosteroid-related in-hospital hyperglycemia: does it negate mortality benefits in COVID-19? Clin. Infect. Dis. 73, e2848–e2849 (2020).
Schacke, H., Berger, M., Rehwinkel, H. & Asadullah, K. Selective glucocorticoid receptor agonists (SEGRAs): novel ligands with an improved therapeutic index. Mol. Cell. Endocrinol. 275, 109–117 (2007).
Sundahl, N., Bridelance, J., Libert, C., De Bosscher, K. & Beck, I. M. Selective glucocorticoid receptor modulation: New directions with non-steroidal scaffolds. Pharmacol. Ther. 152, 28–41 (2015).
Surjit, M. et al. Widespread negative response elements mediate direct repression by agonist-liganded glucocorticoid receptor. Cell 145, 224–241 (2011).
Vandevyver, S., Dejager, L., Tuckermann, J. & Libert, C. New insights into the anti-inflammatory mechanisms of glucocorticoids: an emerging role for glucocorticoid-receptor-mediated transactivation. Endocrinology 154, 993–1007 (2013).
Dewint, P. et al. A plant-derived ligand favoring monomeric glucocorticoid receptor conformation with impaired transactivation potential attenuates collagen-induced arthritis. J. Immunol. 180, 2608–2615 (2008).
Ripa, L. et al. Discovery of a novel oral glucocorticoid receptor modulator (AZD9567) with improved side effect profile. J. Med. Chem. 61, 1785–1799 (2018).
Buttgereit, F. et al. Fosdagrocorat (PF-04171327) versus prednisone or placebo in rheumatoid arthritis: a randomised, double-blind, multicentre, phase IIb study. RMD Open. 5, e000889 (2019).
Safy, M. et al. Efficacy and safety of selective glucocorticoid receptor modulators in comparison to glucocorticoids in arthritis, a systematic review. PLoS One 12, e0188810 (2017).
Myrback, T. H. et al. Effects of a selective glucocorticoid receptor modulator (AZD9567) versus prednisolone in healthy volunteers: two phase 1, single-blind, randomised controlled trials. Lancet Rheumatol. 2, E31–E41 (2020).
US National Library of Medicine. ClinicalTrials.gov https://ClinicalTrials.gov/show/NCT04556760 (2020).
Masuzaki, H. et al. A transgenic model of visceral obesity and the metabolic syndrome. Science 294, 2166–2170 (2001).
Tomlinson, J. W. et al. Impaired glucose tolerance and insulin resistance are associated with increased adipose 11beta-hydroxysteroid dehydrogenase type 1 expression and elevated hepatic 5alpha-reductase activity. Diabetes 57, 2652–2660 (2008).
Peng, K. et al. 11β-Hydroxysteroid dehydrogenase type 1(11β-HSD1) mediates insulin resistance through JNK activation in adipocytes. Sci. Rep. 6, 37160 (2016).
Morgan, S. A. et al. 11β-HSD1 is the major regulator of the tissue-specific effects of circulating glucocorticoid excess. Proc. Natl Acad. Sci. USA 111, E2482–E2491 (2014).
Rosenstock, J. et al. The 11-beta-hydroxysteroid dehydrogenase type 1 inhibitor INCB13739 improves hyperglycemia in patients with type 2 diabetes inadequately controlled by metformin monotherapy. Diabetes Care 33, 1516–1522 (2010).
Heise, T. et al. Safety, efficacy and weight effect of two 11beta-HSD1 inhibitors in metformin-treated patients with type 2 diabetes. Diabetes Obes. Metab. 16, 1070–1077 (2014).
Feig, P. U. et al. Effects of an 11beta-hydroxysteroid dehydrogenase type 1 inhibitor, MK-0916, in patients with type 2 diabetes mellitus and metabolic syndrome. Diabetes Obes. Metab. 13, 498–504 (2011).
US National Library of Medicine. ClinicalTrials.gov https://ClinicalTrials.gov/show/NCT03111810 (2017).
Acknowledgements
The authors gratefully acknowledge the support of research programmes from the Canadian Institutes of Health Research (PJT-156194 and PJT-156159) and the Natural Science and Engineering Research Council of Canada (RGPIN-2020-07212). J.X.L. is a recipient of the Ontario Graduate Scholarship. The authors thank Dr Ahmed Aman (Ontario Institute for Cancer Research) for providing guidance on therapeutics currently under development.
Author information
Authors and Affiliations
Contributions
The authors contributed equally to all aspects of the article.
Corresponding author
Ethics declarations
Competing interests
C.L.C. is a co-inventor on US9428753 patent entitled “Use of LXR antagonists for treatment of side effects of elevated glucocorticoid levels”. J.X.L. declares no competing interests.
Peer review
Peer review information
Nature Reviews Endocrinology thanks the anonymous reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Li, JX., Cummins, C.L. Fresh insights into glucocorticoid-induced diabetes mellitus and new therapeutic directions. Nat Rev Endocrinol 18, 540–557 (2022). https://doi.org/10.1038/s41574-022-00683-6
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41574-022-00683-6
This article is cited by
-
VEGF and EGFR signaling pathways are involved in the baicalein attenuation of OVA-induced airway inflammation and airway remodeling in mice
Respiratory Research (2024)
-
Nuclear receptor corepressors non-canonically drive glucocorticoid receptor-dependent activation of hepatic gluconeogenesis
Nature Metabolism (2024)
-
Insights and implications of sexual dimorphism in osteoporosis
Bone Research (2024)
-
Safety of non-standard regimen of systemic steroid therapy in patients with Graves’ orbitopathy: a single-centre experience
Pharmacological Reports (2024)
-
Impact of subcutaneous belimumab on disease activity, patient satisfaction, and metabolic profile in long-lasting systemic lupus erythematosus
Clinical Rheumatology (2024)
