
Phytopigment Alizarin Inhibits Multispecies Biofilm Development by Cutibacterium acnes, Staphylococcus aureus, and Candida albicans


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Chemicals
2.2. MIC Determinations and Planktonic Cell Growth Measurements
2.3. Biofilm Formation Suppression Assay
2.4. Biofilm Observations by Live Imaging Microscopy and SEM
2.5. Biofilm Observations by Confocal Microscopy
2.6. Assay of Multispecies Biofilm Development by C. acnes, S. aureus, and C. albicans
2.7. Cell Agglutination as Determined by EPS Production
2.8. Cell Surface Hydrophobicity
2.9. Lipase Production Assay
2.10. Cell Aggregation in Filamentous Growth Promoting Liquid Media
2.11. Transcriptomic Studies by Quantitative Reverse Transcriptase Real-Time PCR (qRT-PCR)
2.12. Biofilm Inhibition Analysis on Porcine Skin
2.13. Statistical Analysis
3. Results
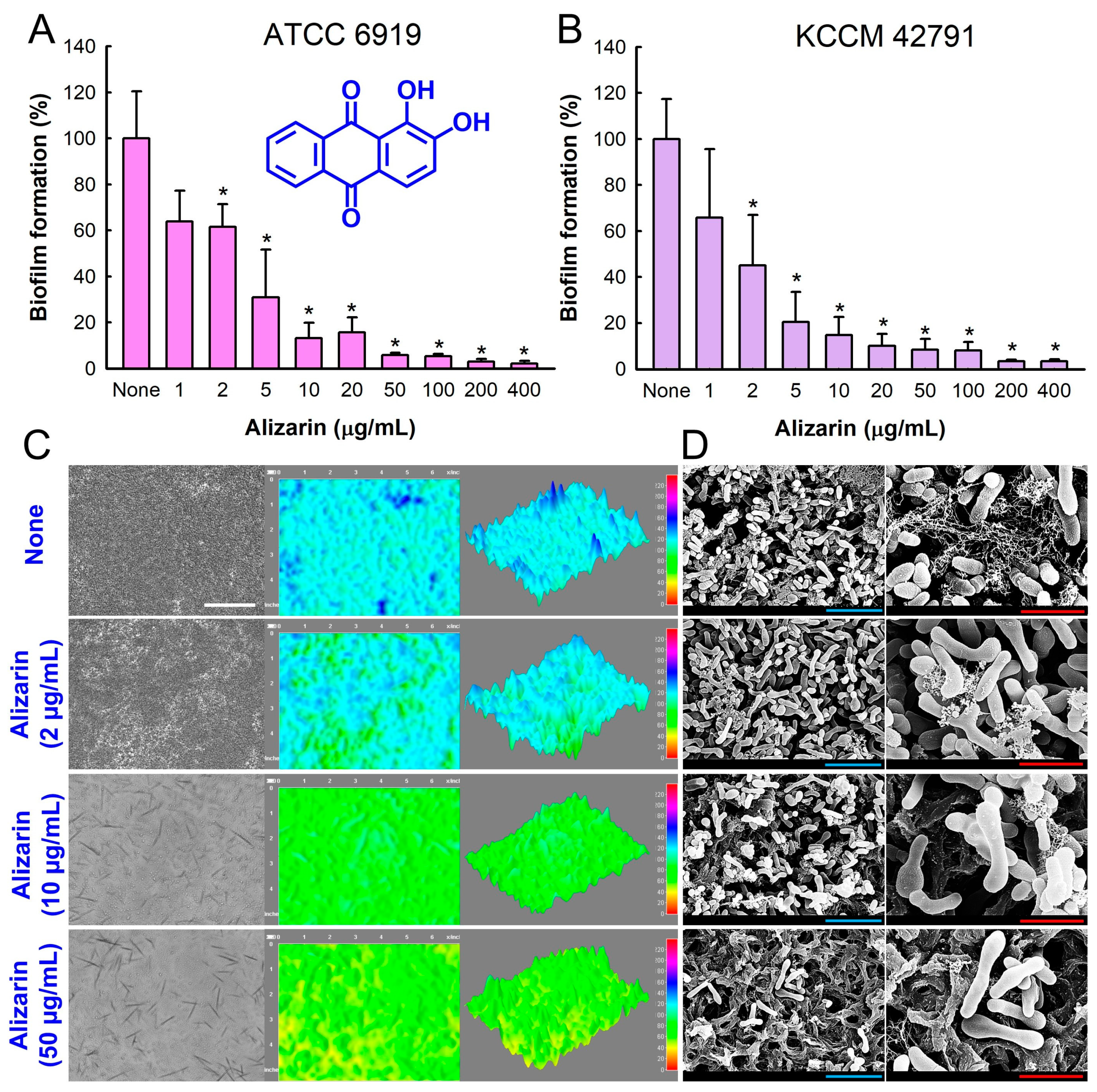
3.1. Antibiofilm Activity of Alizarin against C. acnes
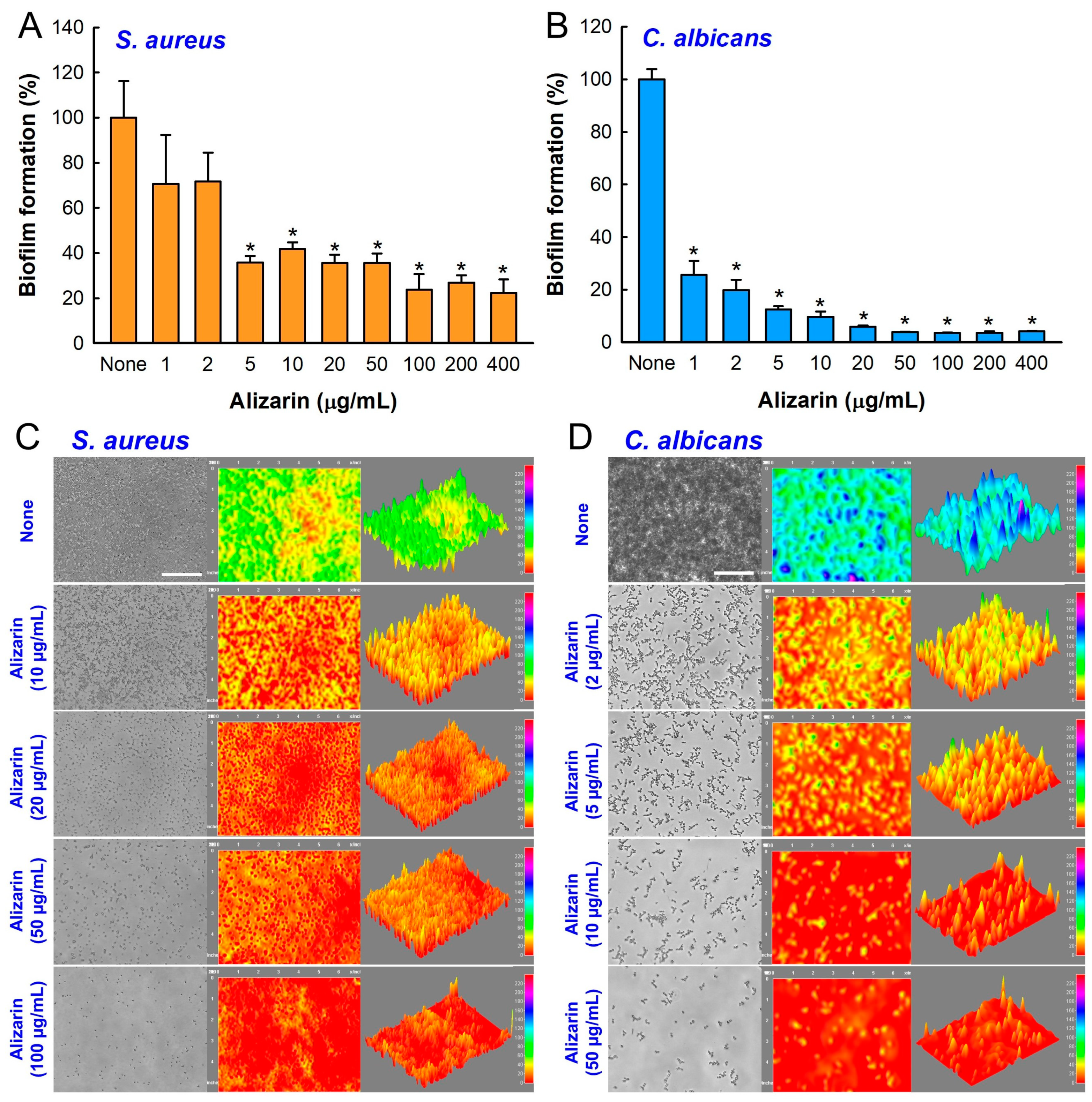
3.2. Alizarin Inhibited Biofilm Development by S. aureus and C. albicans under Anaerobic Conditions
3.3. Alizarin Prevented Multispecies Biofilm Development by C. acnes, S. aureus, and C. albicans
3.4. Alizarin Increased the Antibiofilm Activities of Antibiotics against C. acnes
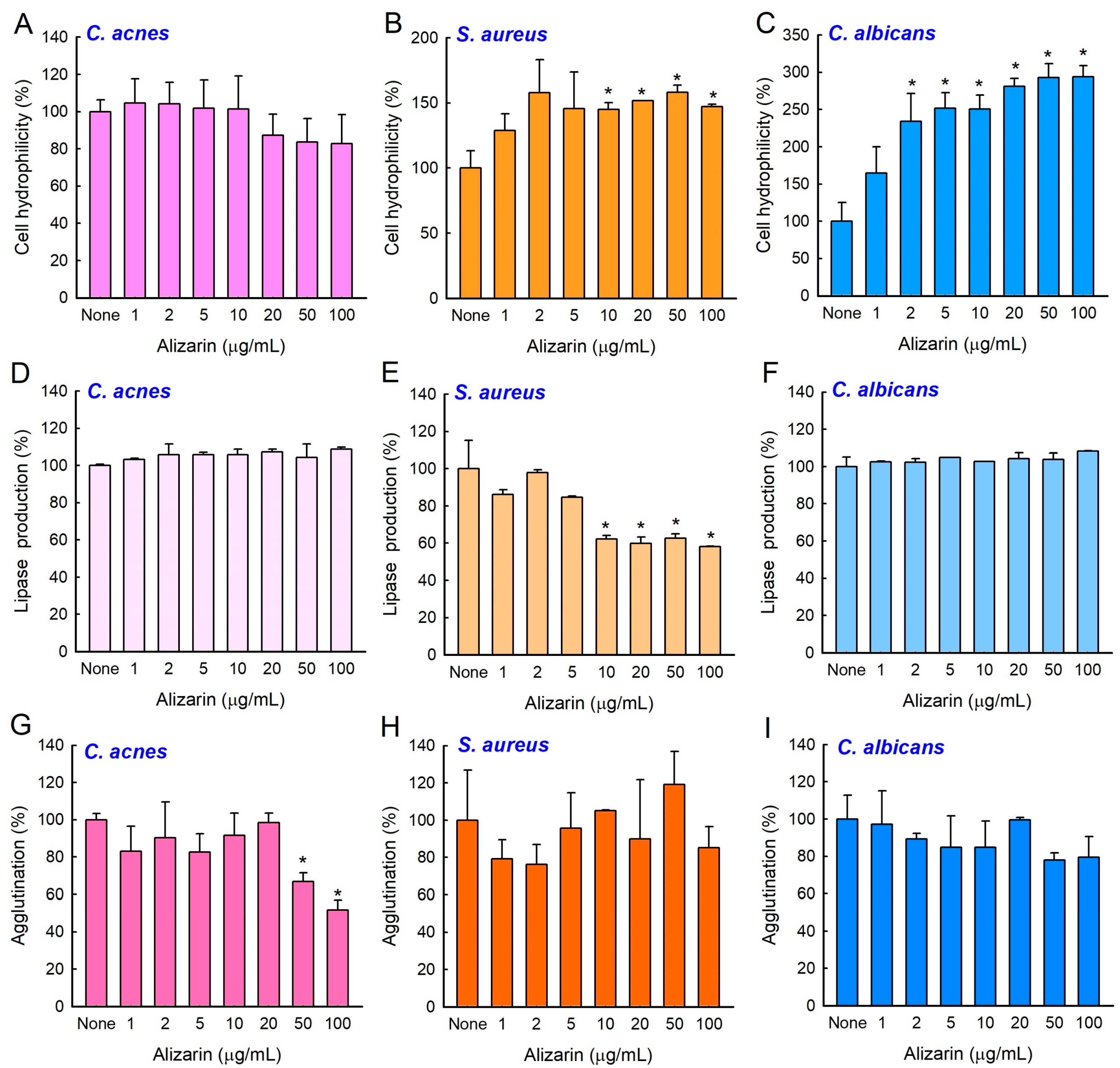
3.5. Effects of Alizarin on Cell Surface Hydrophilicity, Lipase Production, and Cell Agglutination
3.6. Alizarin Inhibited Cell Aggregation by C. albicans
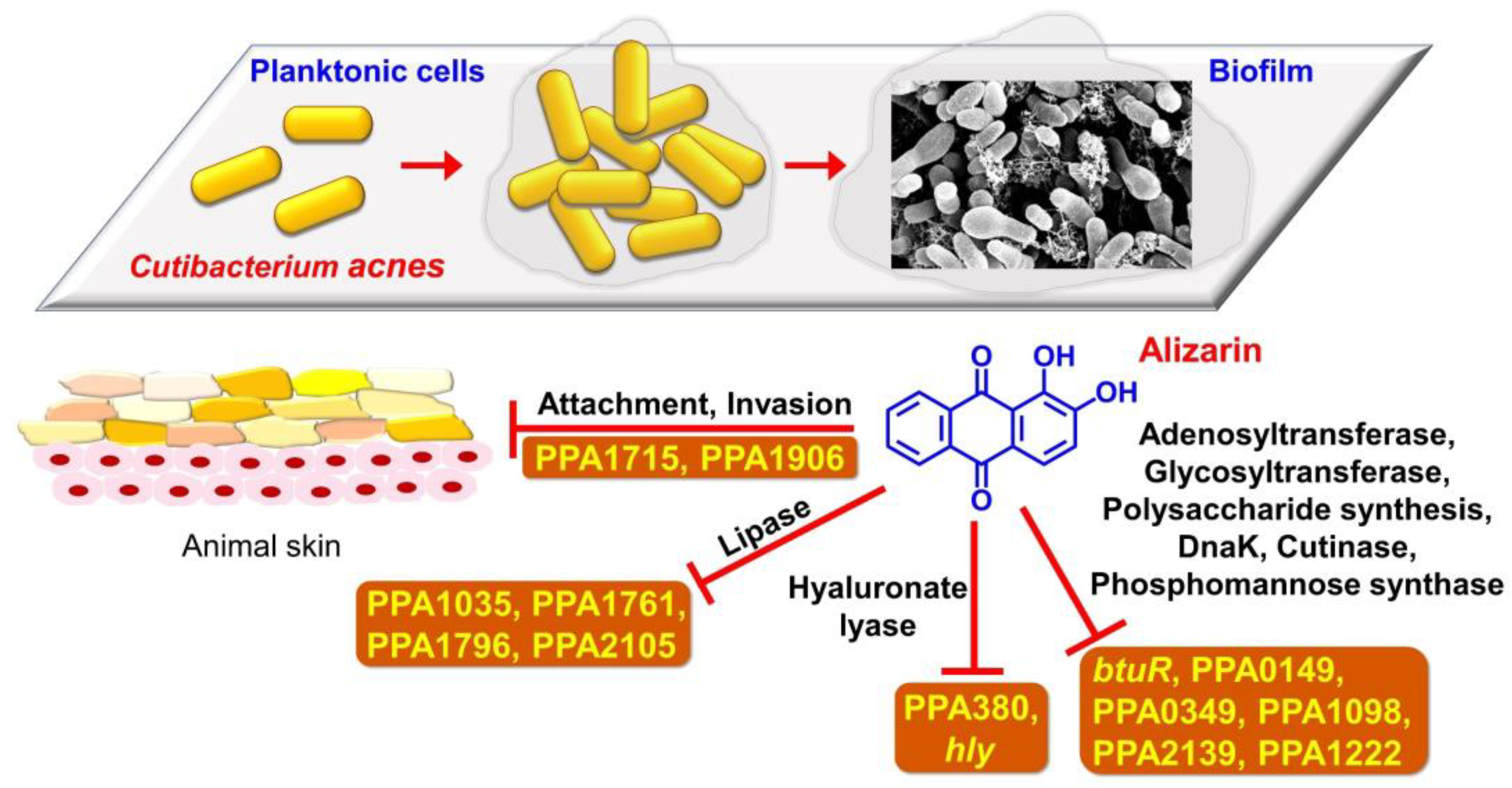
3.7. Alizarin Altered Gene Expressions in C. acnes
3.8. Alizarin Eradicated C. acnes on Porcine Skin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ATCC | American type culture collection |
| CFDA-SE | Carboxyfluorescein diacetate succinimidyl ester |
| CFU | Colony-forming unit |
| CLSM | Confocal laser scanning microscopy |
| CSH | Cell surface hydrophobicity |
| DMSO | Dimethyl sulfoxide |
| EPS | Extracellular polymeric substances |
| KCCM | Korean culture center of microorganisms |
| LUT | Lookup table |
| MIC | Minimum inhibitory concentration |
| LB | Lysogeny broth |
| PBS | Phosphate-buffered saline |
| PDB | Potato dextrose broth |
| qRT-PCR | Quantitative real-time reverse transcription polymerase chain reaction |
| RCM | Reinforced clostridium media |
| SEM | Scanning electron microscopy |
References
- Bruggemann, H.; Henne, A.; Hoster, F.; Liesegang, H.; Wiezer, A.; Strittmatter, A.; Hujer, S.; Durre, P.; Gottschalk, G. The complete genome sequence of Propionibacterium acnes, a commensal of human skin. Science 2004, 305, 671–673. [Google Scholar] [CrossRef] [PubMed]
- Dreno, B.; Pecastaings, S.; Corvec, S.; Veraldi, S.; Khammari, A.; Roques, C. Cutibacterium acnes (Propionibacterium acnes) and acne vulgaris: A brief look at the latest updates. J. Eur. Acad. Dermatol. 2018, 32, 5–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyner, H.; Patel, R. Propionibacterium acnes biofilm—A sanctuary for Staphylococcus aureus? Anaerobe 2016, 40, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Bernard, C.; Renaudeau, N.; Mollichella, M.L.; Quellard, N.; Girardot, M.; Imbert, C. Cutibacterium acnes protects Candida albicans from the effect of micafungin in biofilms. Int. J. Antimicrob. Agents 2018, 52, 942–946. [Google Scholar] [CrossRef]
- Seo, S.; Jung, J.Y.; Kim, C.Y.; Kang, H.; Lee, I.H. Antimicrobial peptides encounter resistance of aureolysin during their action on Staphylococcus aureus biofilm. Biotechnol. Bioproc. Eng. 2021, 26, 216–222. [Google Scholar] [CrossRef]
- Potera, C. Forging a link between biofilms and disease. Science 1999, 283, 1837–1839. [Google Scholar] [CrossRef]
- Lee, J.-H.; Kim, Y.-G.; Ryu, S.Y.; Lee, J. Calcium-chelating alizarin and other anthraquinones inhibit biofilm formation and the hemolytic activity of Staphylococcus aureus. Sci. Rep. 2016, 6, 19267. [Google Scholar] [CrossRef] [Green Version]
- Manoharan, R.K.; Lee, J.-H.; Kim, Y.-G.; Lee, J. Alizarin and chrysazin inhibit biofilm and hyphal formation by Candida albicans. Front. Cell. Infect. Microbiol. 2017, 7, 447. [Google Scholar] [CrossRef]
- Fotia, C.; Avnet, S.; Granchi, D.; Baldini, N. The natural compound alizarin as an osteotropic drug for the treatment of bone tumors. J. Orthop. Res. 2012, 30, 1486–1492. [Google Scholar] [CrossRef] [Green Version]
- Stana, A.; Vodnar, D.C.; Tamaian, R.; Pirnau, A.; Vlase, L.; Ionut, I.; Oniga, O.; Tiperciuc, B. Design, synthesis and antifungal activity evaluation of new thiazolin-4-ones as potential lanosterol 14a-demethylase inhibitors. Int. J. Mol. Sci. 2017, 18, 177. [Google Scholar] [CrossRef]
- Peeters, E.; Nelis, H.J.; Coenye, T. Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. J. Microbiol. Methods 2008, 72, 157–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.-G.; Lee, J.-H.; Gwon, G.; Kim, S.-I.; Park, J.G.; Lee, J. Essential oils and eugenols inhibit biofilm formation and the virulence of Escherichia coli O157:H7. Sci. Rep. 2016, 6, 36377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heydorn, A.; Nielsen, A.T.; Hentzer, M.; Sternberg, C.; Givskov, M.; Ersboll, B.K.; Molin, S. Quantification of biofilm structures by the novel computer program COMSTAT. Microbiology 2000, 146, 2395–2407. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-G.; Lee, J.-H.; Park, S.; Kim, S.; Lee, J. Inhibition of polymicrobial biofilm formation by saw palmetto oil, lauric acid and myristic acid. Microb. Biotechnol. 2022, 15, 590–602. [Google Scholar] [CrossRef] [PubMed]
- Sethupathy, S.; Sathiyamoorthi, E.; Kim, Y.-G.; Lee, J.-H.; Lee, J. Antibiofilm and antivirulence properties of indoles against Serratia marcescens. Front. Microbiol. 2020, 11, 584812. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, M.; Gutnick, D.; Rosenberg, E. Adherence of bacteria to hydrocarbons: A simple method for measuring cell-surface hydrophobicity. FEMS Microbiol. Lett. 1980, 9, 29–33. [Google Scholar] [CrossRef]
- Sivasankar, C.; Maruthupandiyan, S.; Balamurugan, K.; Bhaskar, J.P.; Krishnan, V.; Pandian, S.K. A combination of ellagic acid and tetracycline inhibits biofilm formation and the associated virulence of Propionibacterium acnes in vitro and in vivo. Biofouling 2016, 32, 397–410. [Google Scholar] [CrossRef]
- Lee, J.-H.; Kim, Y.-G.; Khadke, S.K.; Lee, J. Antibiofilm and antifungal activities of medium-chain fatty acids against Candida albicans via mimicking of the quorum-sensing molecule farnesol. Microb. Biotechnol. 2020, 14, 1353–1366. [Google Scholar] [CrossRef]
- Yang, Q.P.; Phillips, P.L.; Sampson, E.M.; Progulske-Fox, A.; Jin, S.G.; Antonelli, P.; Schultz, G.S. Development of a novel ex vivo porcine skin explant model for the assessment of mature bacterial biofilms. Wound Repair Regen. 2013, 21, 704–714. [Google Scholar] [CrossRef]
- Kim, Y.-G.; Lee, J.-H.; Lee, J. Antibiofilm activities of fatty acids including myristoleic acid against Cutibacterium acnes via reduced cell hydrophobicity. Phytomedicine 2021, 91, 153710. [Google Scholar] [CrossRef]
- Tribedi, P.; Sil, A.K. Cell surface hydrophobicity: A key component in the degradation of polyethylene succinate by Pseudomonas sp. AKS2. J. Appl. Microbiol. 2014, 116, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, R.; Santhakumari, S.; Poonguzhali, P.; Geetha, M.; Dyavaiah, M.; Lin, X.M. Bacterial biofilm inhibition: A focused review on recent therapeutic strategies for combating the biofilm mediated infections. Front. Microbiol. 2021, 12, 676458. [Google Scholar] [CrossRef] [PubMed]
- Chandra, J.; Kuhn, D.M.; Mukherjee, P.K.; Hoyer, L.L.; McCormick, T.; Ghannoum, M.A. Biofilm formation by the fungal pathogen Candida albicans: Development, architecture, and drug resistance. J. Bacteriol. 2001, 183, 5385–5394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achermann, Y.; Goldstein, E.J.; Coenye, T.; Shirtliff, M.E. Propionibacterium acnes: From commensal to opportunistic biofilm-associated implant pathogen. Clin. Microbiol. Rev. 2014, 27, 419–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portillo, M.E.; Corvec, S.; Borens, O.; Trampuz, A. Propionibacterium acnes: An underestimated pathogen in implant-associated infections. Biomed Res. Int. 2013, 2013, 804391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usai, M.; Marchetti, M. Anthraquinone distribution in the hypogeal apparatus of Rubia peregrina L. growing wild in Sardinia. Nat. Prod. Res. 2010, 24, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Wang, P.L.; Wang, L.; Liu, C.M.; Xu, S.X.; Cheng, Y.T.; Wang, Y.H.; Li, Q.; Lei, H.M. Quinone derivatives from the genus Rubia and their bioactivities. Chem. Biodivers. 2014, 11, 341–363. [Google Scholar] [CrossRef]
- Malmir, M.; Serrano, R.; Silva, O. Anthraquinones as potential antimicrobial agents—A review. In Antimicrobial Research: Novel Bioknowledge and Educational Programs; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2017; pp. 55–61. [Google Scholar]
- Coenye, T.; Honraet, K.; Rigole, P.; Nadal Jimenez, P.; Nelis, H.J. In vitro inhibition of Streptococcus mutans biofilm formation on hydroxyapatite by subinhibitory concentrations of anthraquinones. Antimicrob. Agents Chemother. 2007, 51, 1541–1544. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.M.; Zhang, J.L.; Zhou, K.; Yue, L.M.; Zhang, Y.; Wang, C.Y.; Wang, K.L.; Xu, Y. Anthraquinones as potential antibiofilm agents against methicillin-resistant Staphylococcus aureus. Front. Microbiol. 2021, 12, 709826. [Google Scholar] [CrossRef]
- Nobile, C.J.; Mitchell, A.P. Regulation of cell-surface genes and biofilm formation by the C. albicans transcription factor Bcr1p. Curr. Biol. 2005, 15, 1150–1155. [Google Scholar] [CrossRef] [Green Version]
- An, Y.H.; Friedman, R.J. Concise review of mechanisms of bacterial adhesion to biomaterial surfaces. J. Biomed. Mater. Res. 1998, 43, 338–348. [Google Scholar] [CrossRef]
- Dunne, W.M., Jr. Bacterial adhesion: Seen any good biofilms lately? Clin. Microbiol. Rev. 2002, 15, 155–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faleye, O.S.; Sathiyamoorthi, E.; Lee, J.-H.; Lee, J. Inhibitory effects of cinnamaldehyde derivatives on biofilm formation and virulence factors in Vibrio Species. Pharmaceutics 2021, 13, 2176. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Kim, Y.-G.; Lee, K.; Kim, S.C.; Lee, J. Temperature-dependent control of Staphylococcus aureus biofilms and virulence by thermoresponsive oligo(N-vinylcaprolactam). Biotechnol. Bioeng. 2015, 112, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Coenye, T.; Peeters, E.; Nelis, H.J. Biofilm formation by Propionibacterium acnes is associated with increased resistance to antimicrobial agents and increased production of putative virulence factors. Res. Microbiol. 2007, 158, 386–392. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-G.; Lee, J.-H.; Park, S.; Lee, J. The anticancer agent 3,3’-diindolylmethane inhibits multispecies biofilm formation by acne-causing bacteria and Candida albicans. Microbiol. Spect. 2022, 10, e0205621. [Google Scholar] [CrossRef] [PubMed]
- Holland, C.; Mak, T.N.; Zimny-Arndt, U.; Schmid, M.; Meyer, T.F.; Jungblut, P.R.; Bruggemann, H. Proteomic identification of secreted proteins of Propionibacterium acnes. BMC Microbiol. 2010, 10, 230. [Google Scholar] [CrossRef] [Green Version]
- Allhorn, M.; Arve, S.; Bruggemann, H.; Lood, R. A novel enzyme with antioxidant capacity produced by the ubiquitous skin colonizer Propionibacterium acnes. Sci. Rep. 2016, 6, 36412. [Google Scholar] [CrossRef] [Green Version]
- Raj, V.; Kim, Y.; Kim, Y.-G.; Lee, J.-H.; Lee, J. Chitosan-gum arabic embedded alizarin nanocarriers inhibit biofilm formation of multispecies microorganisms. Carbohydr. Polym. 2022, 284, 118959. [Google Scholar] [CrossRef]
- Choi, Y.; Jang, J.; Koo, H.J.; Tanaka, M.; Lee, K.H.; Choi, J. Alginate-chitosan hydrogel patch with beta-glucan nanoemulsion for antibacterial applications. Biotechnol. Bioproc. Eng. 2021, 26, 71–77. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-H.; Kim, Y.-G.; Park, S.; Hu, L.; Lee, J. Phytopigment Alizarin Inhibits Multispecies Biofilm Development by Cutibacterium acnes, Staphylococcus aureus, and Candida albicans. Pharmaceutics 2022, 14, 1047. https://doi.org/10.3390/pharmaceutics14051047
Lee J-H, Kim Y-G, Park S, Hu L, Lee J. Phytopigment Alizarin Inhibits Multispecies Biofilm Development by Cutibacterium acnes, Staphylococcus aureus, and Candida albicans. Pharmaceutics. 2022; 14(5):1047. https://doi.org/10.3390/pharmaceutics14051047
Chicago/Turabian StyleLee, Jin-Hyung, Yong-Guy Kim, Sunyoung Park, Liangbin Hu, and Jintae Lee. 2022. "Phytopigment Alizarin Inhibits Multispecies Biofilm Development by Cutibacterium acnes, Staphylococcus aureus, and Candida albicans" Pharmaceutics 14, no. 5: 1047. https://doi.org/10.3390/pharmaceutics14051047