
Abstract
Climatic changes and their impact on animals and plants at the margins of their distributions are evident in terrestrial, limnic, and marine environments. I review and summarise the literature related to the direct physiological effects of climate change on marginal populations in all realms and categories. I summarise literature showing physiological and ecological emphasis but do not include studies on modelling, genetic or range shift. I found a great discrepancy in representation of each category, with the majority of studies and the greatest variety of species represented in the marine environment and the least research conducted in limnic environments. The least represented in all environments and marginalities was the plant kingdom, including algae. I identified an uneven distribution of studies around the globe with a major focus on Europe and North America. Literature results show an overarching negative impact on organismal physiology independent of a realm and/or geographic location, potentially leading to species and hence biodiversity loss, and tightly coupled with this, a loss of ecosystem functions at the respective sites. All these points may lead to unstable ecosystems and socio-economic implications at the respective sites.
Similar content being viewed by others

Introduction
A central assumption of biogeography is the extraordinary control over the natural distribution of species by climate. The abiotic and biotic environment changes continuously. These changes challenge organisms to maintain homeostasis, forcing individuals, populations and species to acclimatise and evolve both physiologically and structurally. Past climatic changes profoundly influenced species range distribution (Woodward 1987). With significant climatic changes occurring in the next 50–100 years due to anthropogenic emissions (IPCC 2021), many organisms will likely be unable to exist within their current natural distribution.
It is commonly thought that challenges caused by current and future climatic changes may be greater for populations living at the edge of their range. Different hypotheses attempt to describe the cause of this perception looking at variance in distribution, density, genetic diversity, environmental conditions or a combination of these. Lawton (1993), for instance, postulates that, within their natural range, species have several different patterns of distribution and abundance with an overall decline in the proportion of occupied sites and the average population densities from the centre to the margin of a species’ range. According to the “centre-periphery” hypothesis (Mayr 1963), the lower genetic diversity of marginal populations limits their adaptive ability compared to central populations (Gaston 2003; Reed 2004; see also review Hampe and Petit 2005), which could play an important role when facing future climatic changes. A third theory, the “abundant-centre” theory (Brown 1984; Sagarin and Gaines 2002), hypothesizes that the geographic centre of a species distribution holds the most favourable environmental conditions and that, therefore, the population density is highest in the centre (Whittaker 1956; Brussard 1984). Towards the margins of the distribution, environmental variables are thought to become less favourable due to greater abiotic stress and increased interspecific competition (Aitken et al. 2008), initiating a decrease in population densities and lower relative fertility (Case and Taper 2000). The abundant centre distribution has widely been used as the basis for hypotheses about ecological and evolutionary processes, however, many factors may confound the abundant centre hypothesis, such as latitude, ecological environmental gradients, phylogeographic history, etc. Other studies show complex demographic patterns in which peaks of abundances are shifted towards one end of the species' marginal populations or show no pattern (reviewed by Sagarin and Gaines (2006) and Sagarin et al. (2006)). Ultimately, it is likely that a combination of some or all of the above-named hypotheses directly influences the physiological response of central and marginal populations to future climatic changes, depending on the species and environment, and it is difficult to determine the exact cause or hypothesis without in-depth and meticulous research.
While characterizing a species geographical distribution, there are two established types of marginality. Ecological marginality (also called continuous peripheral populations), focussing on the species’ optimum conditions and the ecology of the environment it would be able to survive in, and geographical marginality (also called disjunct peripheral populations), described as the limit of the species’ natural, geographical distribution (Soulé 1973; Hoffmann and Blows 1994; Lesica and Allendorf 1995). Marginal populations are, therefore, populations, often but not necessarily, living on the limit of environmental tolerance, mostly isolated from central and continuous populations (Bridle and Vines 2007; Bertocci et al. 2011) and with greater temporal variability when environmental fluctuations exceed the species’ tolerance (Sexton et al. 2009). Additionally, they tend to be more vulnerable to stochastic or catastrophic events, particularly to extinction (estimated to be up to 21–32%; Thomas et al. 2004) due to their smaller sizes, compared to central populations (Soulé 1973; Brussard 1984; Lawton 1993; Lesica and Allendorf 1995; Vucetich and Waite 2003; Nicastro et al. 2013).
Two visions of the future of marginal populations currently coexist, ranging between predictions of complete disappearance, based on modelling (Hampe and Jump 2011), and more optimistic expectations based on past persistence. The authors of some of these studies suggest it would be interesting to test if climate change affected marginal population performance primarily through direct abiotic limitations, such as drought in terrestrial or water temperature in aquatic systems (Hampe and Petit 2005). Research on marginal populations is still very limited and little is known about the direct effects of climate change on them (predominantly low and high latitude distribution range margins) across different ecosystems. With respect to the genetic variation across species’ geographic ranges, more than 40% of studies focussed on the northern limit of species in the temperate zone of the northern hemisphere (reviewed by Hampe and Petit (2005) and Eckert et al. (2008)). None involved an exclusively tropical species. Their results suggest that northern peripheral populations may exhibit a limited evolutionary response to warming conditions (Etterson and Shaw 2001). To avoid extinction, marginal populations can respond to changing climate by migrating to track temperature changes, resulting in range shifts, or by adapting to novel local climates.
Comparing central to marginal populations of species provides an understanding of determinants of species ranges and of how spatial variation in fitness translates to population-level differences in abundance (Sexton et al. 2009; Araújo et al. 2014; Guo 2014). Marginal populations may represent an important source for speciation events in the future, triggered by e.g. climatic changes, subsequently developing unique eco-physiological local adaptations and genetic structure (Araújo et al. 2009). Thus, range limits can aid as testing grounds to understand the conditions by which populations can adapt—or fail to adapt—to novel conditions (Helmuth et al. 2002; Thornton et al. 2018). This will enable us to predict how these species and subsequently the ecosystems they are living in might change.
Scope of this review
This review focuses on studies of marginal populations (Fig. 1) of wild species, independently of ecosystem and/or different types of marginality or life stages. It includes only direct and observed, positive and negative effects of anthropogenic climate changes, excluding studies on range shift, genetic and modelling, since these have been addressed in previous reviews (Hampe and Petit 2005; Sagarin and Gaines 2006; Eckert et al. 2008). Methodology or conservation implications are not discussed within this work. Rather, studies are outlined and summarised to illustrate the complexity and variability of climatic changes on marginal populations. There is large uncertainty within the scientific community as to how terrestrial, marine and limnic populations would react to climatic changes with the limitations forced on them through anthropogenic influences. This review elucidates most of the above-named aspects and summarises them naming examples for each mentioned broader ecosystem.

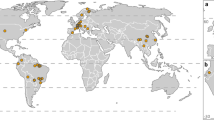
World map including areas of investigation (line along continents) and number of studies per continent (hexagons)
Materials and methods
We systematically searched for literature focussing on the effect of climate changes on marginal and peripheral populations. We conducted multiple searches in the ISI Web of Science, BIOSIS and SciELO databases between March and September 2019 and June 2020. Full key words in these searches were as follows: ([species (range or distribution) climate change* (margin* OR limit* OR edge* OR peripheral* OR limit* OR border*) AND (marine* OR terrestrial* OR freshwater*)). The second part of the literature research used the following full key work search: ([species (range or distribution) (margin* OR limit* OR edge* OR peripheral* OR limit* OR border*) AND (marine* OR terrestrial* OR freshwater*) AND climate change* (cause* OR effect*)]). Merging results of these searches resulted in 10,225 unique records. After reading full abstracts, studies unrelated to climate change impacts on marginal populations were discarded. Studies on range shift, modelled effects on organisms, solely central populations and those looking at genetics were excluded in line with the scope of this review. Review papers within this search were double-checked for potentially missed literature and after examination, were excluded to avoid double recording. The remaining 93 studies were reviewed in depth. Following the example of other studies, we did not attempt to further re-analyse or verify the results of these studies. Based on the conclusions of the authors of each study, we examined if each factor of interest (marginality, climate change conditions, direct effects) was supported.
To facilitate the review process, we separated the studies into terrestrial, limnic and marine sections and thereafter, split these into groups including either flora or fauna. Within each category, a few studies were investigating multiple species and were accounted for depending on the number of species as analyses were summarised on a species level within each group. Species represented multiple times in different studies, were counted individually for each appearance.
Terrestrial populations
Research on terrestrial animal and plant species are relatively evenly distributed (Table 1, Fig. 2A). However, the majority of studies focussed on adult individuals or did not further characterise the life stages of the tested organisms. In animals, the majority of research focused on the northern margin and high latitude populations introducing a bias towards the interpretation of ecological and physiological data throughout a latitudinal and altitudinal distribution gradient.

Positive (dark grey bars) and negative (light grey bars) direct effects of climate change on multiple physiological traits of terrestrial (A), limnic (B) and intertidal and marine (C) animals, plants and algae found in the reviewed studies. Literature did not report neutral or no direct effects of climate change on physiological traits. Studies are divided into geographical locations (north, centre and south)
Animals
Negative effects on animals
In terrestrial populations, an increase in temperature seems to be one of the major drivers of changes in all animal life stages. Current literature showed a dependency between temperature increase, life stages and the species-specific temperature threshold, and a correlation of those factors in relation with positive or negative effects on marginal populations. For example, the air temperature has a direct negative effect on early life-cycle stages of butterfly species Aporia crataegi and Parnassius apollo within their oviposition, egg survival, habitat use and survival of caterpillars in Spanish mountain ranges, at their elevational and local marginality (Merrill et al. 2008; Ashton et al. 2009). For instance, egg survival of A. crataegi decreased with elevated temperatures (Merrill et al. 2008) and P. apollo showed a shift in microhabitat usage of larvae towards more shaded and colder conditions favouring different host plans than at colder temperatures (Ashton et al. 2009).
Other butterfly taxa show negative relationships between temperature changes and fecundity and fitness, which declined at the north range margins, and was expressed through e.g. a reduction in abdominal mass and wing load (insect mass/wing area) or a decline in immune function. This reduction in body shape and size and decline in immune function has detrimental effects on insect flight speed and manoeuvrability of the family of skipper butterflies and the butterfly Polygonia c-album, consequently leading to increased vulnerability towards predation and survival of changing environmental factors (Braschler and Hill 2007; Hill et al. 2011 and references therein).
The temperature in mammals, particularly in rodents and endotherms, imposes an immense pressure on maximum thermoregulatory capacity (reviewed by Bozinovic et al. (2011)). Humphries et al. (2002) demonstrated restrictions in hibernation energetics, like winter energy requirements and expenditure, by increasing temperature in its southern distribution, resulting in a northward expansion of the American southern and northern marginal populations of the bat Myotis lucifugus.
Adverse weather conditions, such as cool and rainy days, are also shown to result in reduced feeding activity in Swiss alp bird Upupa epops higher and cooler altitudinal peripheral populations leading to a decrease in biomass of an individual and therefore to a decrease of its productivity (Arlettaz et al. 2010). Around hatching time is when the impact on survival and productivity on this species is observed to be particularly negative, in contrast, warm conditions boosted its productivity.
A temperature and location connected impact of climate change is aridity. Davies et al. (2013) showed that the populations of the koala species Phascolarctos cinereus at their arid margin of distribution in southwestern Queensland excrete higher cortisol metabolite concentrations in faecal samples, as a measure of stress. This area is characterised by low precipitation levels, inducing increasingly higher physiological stress levels in P. cinereus when the water content is low. In the light of climate change, increasing draught possibilities, increasing temperatures and number of heatwaves, P. cinereus populations at the arid and semi-arid, inland margin of their geographical distribution will fail to survive supported by these findings.
In the class Insecta, an effect regarding temperature-induced alteration of life cycles has also been detected, similarly to the one in Aves. The mosquito Wyeomyia smithii had developed a much shorter critical photoperiod in relation to a longer growing season. Northern populations now use a shorter day-length signal to start winter diapause later in the year than 24 years ago (Bradshaw and Holzapfel 2001). It is thought that warming, although relatively small in magnitude, has a higher negative effect on insects inhabiting the tropics compared to organisms living in temperate regions. This is due to already living close to their optimal thermal limit compared to species living at higher latitudes and with a broader thermal tolerance since these populations are living cooler than their thermal optimum and could withstand greater temperature changes (Deutsch et al. 2008).
In butterflies, elevated temperatures showed different responses regarding their physiology. Ashton et al. (2009) showed that P. apollo changed larval preferences for microhabitats changing from bare grounds and dead grasses at colder temperatures to shrubs in warmer temperature conditions, ultimately changing the distribution and elevation of that species at the warm range margin since shrubs grow in higher elevation and will therefore limit the distribution and reproduction success of this species. Crozier (2004) pointed out that population growth rates in the centre as well as marginal populations (South and North) in Atalopedes campestris are positively correlated to monthly and annual mean temperatures over a 2-year period, however, larval survivorship in transplant experiments was low at both sites and significantly lower at the range margin, potentially due to lethal temperatures frequent occurrence. As a combination of these contradicting findings, Crozier (2004) suggests that temperature affects persistence in northern margin populations of this species directly and warmer temperatures in the winter aid range expansions. This is also supported by finding that individuals of the family of skipper butterflies were killed by either a single, short exposure to extremely low temperatures or repeated exposures to lower temperatures indicating that winter temperatures dictate the northern range margin (Crozier 2003, 2004).
Along with the host species, pest species will also be experiencing direct and indirect effects through climatic changes. A parasite nematode Umingmakstrongylus pallikuukensis increased in abundance due to shortened life cycles in response to warming climates. Its explosion in abundance at their north-eastern margin of the range has been linked to negative impacts on range populations of its host, the wild ox Ovibos moschatus, causing a decreased survival rate and fecundity (Kutz et al. 2002, 2005).
Marginal populations of terrestrial animals may often demonstrate higher fitness and adaptation under unfavourable current marginal conditions, however, perform poorly under changing future marginal conditions (Hoffmann and Blows 1994). A geographic pattern is revealed in which species in colder conditions show higher metabolic rates than species in warmer conditions due to an increased energy expenditure to withstand colder conditions which subsequently enables the species to become wider distributed. This means that most taxa need to be considered at a population level when trying to predict the effects of climate change since they might respond differently due to genetically based, distinctive ways to adapt to changing conditions (O’Neil et al. 2014).
Positive effects on animals
In comparison to the named negative effects on terrestrial animals, some studies showed positive feedback to a changing climate. The larvae of the frog Rana arvalis from higher latitudes range margins are developing faster than the ones from the southern range margin. In addition, larvae from the northern margin populations were able to compensate for physiological effects caused by most colder temperatures and a shorter growing season with higher growth and development rates. However, they failed to adapt to the lowest chosen temperature and were only able to adapt to the two warmer temperature treatments in an experiment conducted by Orizaola et al. (2010). Larvae from the northern margin populations grew and developed faster and reached a similar size at metamorphosis in a shorter period than populations from the southern locations of the study area. Only a few studies found positive effects on marginal populations of terrestrial mammals. Northern populations of the red deer Cervus elaphus in Norway showed a very plastic response to climatic changes. Their body size is coupled with the yearly variability of winter temperatures. Warmer winters cause young males to grow larger and females to develop smaller (Post et al. 1999). Males, therefore, increase their access to females who prefer larger mating partners and females invest in an early sexual maturity at the cost of body size. Consequently, this change in sexual dimorphism in this species is a result of gradual winter warming and may lead to higher population densities.
When investigating the butterfly Aricia agestis, populations near the northern margin of the species distribution had specialised on one host genus Helianthemum which grows in hot microclimates and can therefore support larval growth, when adapting to cool conditions. Specimens further towards the centre of the butterfly distribution, however, showed a greater tendency for other host plants, for instance, Geraniaceae (Thomas et al. 2001). A temperature rise could therefore lead to a greater abundance of this species due to the increasing existence of suitable host plants.
Field and laboratory experiments on individuals of the pine processionary moth Thaumetopoea pityocampa comparing north and southern populations of its geographic distribution in Europe revealed an association between feeding behaviour and survival to minimum night temperatures and linked its expansion to warmer winters (Battisti et al. 2005). Across its distribution, this moth showed higher feeding rates at elevated temperatures, resulting in increased survival with increased temperatures.
Plants
Plants are sessile organisms, therefore, it is harder for them to escape stress factors, such as climatic changes. To cope with environmental changes, plants show morphological (e.g. cushion growth, lignotubers, deep root system) and physiological adaptations (mechanisms enhancing the production of stress hormones and stomatal closure) (reviewed by Hampe and Jump (2011)). Most analyses of the impact of abiotic factors to date concentrate on primary and secondary productivity and geographic range shifts. Very few studies investigate the potential effects of climatic changes on the physiological functions of plants, which mirrors the real method on how to be able to predict future changes in individuals and their fitness (Bozinovic et al. 2011). Stevens (1992) reported maintenance of range boundaries due to physiological tolerances between species and showed that species living at higher altitudes showed broader distribution ranges than species living at lower elevations (Janzen 1967). This also means that if such plant species cannot fully avoid climatic changes but change elevation instead, latitudinal range boundaries would still get thinner and these species might disappear from lower elevations at the same time. In all studies performed with both, north and south range margin populations, there is a clear climatic and anthropogenic effect on especially southern populations.
Negative effects on plants
Both monkeyflower species (Mimulus cardinalis and Mimulus lewisii) and the orchid Himantoglossum hircinum showed a negative effect on both marginal populations (Northern and Southern) with a reduction in fecundity, flower production per stem and overall plant size (Angert 2006; Pfeifer et al. 2010). However, the growth rates of the two populations of the monkeyflower species showed contradicting patterns with a higher rate in the centre than at both margins for M. lewisii but the opposite for M. cardinalis. The authors linked this finding to the influence of other dominant external factors. In one of the more recent studies focussing on Calluna vulgaris, populations at the southern and eastern margin of the species distribution showed lower biomass production, shoot–root ratios and tissue δ13C values, indicating a decreased compatibility for soil nutrients and draught susceptibility than in central populations after 1-year exposure in a non-tempered greenhouse experiment. After a 2-year exposure, this was reversed, however, an increase in nitrogen can attenuate the higher draught resistance in marginal populations and the resultant induced shoot–root ratios, making these populations more vulnerable to anthropogenically induced eutrophication along with climate change again (Meyer-Grünefeldt et al. 2016). In polar plant species, a review by Lesica and McCune (2004) revealed that 31–65% of the studies examined showed a decline in abundance of 4 out of 7 marginal arctic-alpine indicator species, none increased. This includes the species reviewed by Robinson et al. (2003 and references therein), who have concluded that the Antarctic maritime moss Sanionia uncinata depends on lower temperatures for positive net carbon balance, showing a low photosynthesis rate and a steady increase in dark respiration rates in a range of increased temperatures (Nakatsubo 2002). For other arctic plant species like Colobanthus quitensis and Deschampsia antarctica (Robinson et al. 2003 and references therein) major impacts were registered for cell length, leaves length and amount per shoot, increased leaves thickness and less branching all leading to reduced size and biomass of the plant trying to fight UV-B damage by increasing leaves thickness and sacrificing growth and branching (Xiong and Day 2001; Xiong et al. 2002). An increase in plant growth due to increased water availability and temperature in polar regions, would lead to higher demand for nutrients and lead to a nutrient-depleted ecosystem. Trees such as Cedrus libani produce more stress indicator metabolites in seedlings (e.g. H2O2, proline) in northern marginal habitats. In response to, especially, frost these metabolites accumulate to build stress resistances or prevent oxidative stress in seedlings and indicate low enzyme activity due to stress conditions (Ayan et al. 2018). A reduction in seedling establishment with increasing draught and temperature was observed in central populations of Fagus sylvatica and has been predicted to occur in marginal populations as well, maybe even in higher manifestation (Jump et al. 2006). Limited growth of seedlings and mature trees has been observed at the southern population of this species geographical distribution relating to more frequent and longer periods of draughts and elevated temperatures. In combination with reduced precipitation, there is not enough water supply to relieve negative effects on tree growth (Jump et al. 2006). Different studies on F. sylvatica and Pinus contorta stated that growth and productivity in marginal populations are generally reduced along a temperature gradient in comparison to central populations, increasing susceptibility to stress, aridity, cold, and soil salinity (Rehfeldt et al. 1999; Kreyling et al. 2014).
Positive effects on plants
CO2 concentrations can also have a positive impact on marginal plant populations in the Northern Hemisphere and could lead to elevated growth through a combination of increased photosynthesis, water use efficiency and a lengthening of vegetative growing (Morison and Gifford 1984; Poorter and Navas 2003; reviewed by Parmesan (2006)). Therefore, plants threatened by drought stress could benefit from rising CO2 levels.
A high phenotypic and genetic variability and resistance to the mentioned water stress has already been observed in both marginal populations of Hordeum spontaneum (Volis et al. 1998). The central-marginal hypothesis, which supports the idea of centre populations being better equipped to sustain stress, is generally thought to be accurate for the majority of species. However, this hypothesis is starting to be opposed by recent research, for instance in the boxthorn Liriodendron chinense (Yang et al. 2016). Other factors such as orientation, latitudinal and ecological environmental gradients may influence central-marginal patterns. In the Antarctic ecosystem, increased ambient CO2 levels might intensify CO2 limitations in some populations but is very likely to cause an overall significant increase in productivity due to increased temperature optima for photosynthesis and water availability (reviewed by Robinson et al. (2003)). Increased temperature in Antarctica is changing community dynamics causing increased photosynthetic rates in three continental moss species and two maritime vascular plants (Lewis Smith 1999; Xiong et al. 2000). In trees, there is a very limited number of studies looking at positive effects induced by climate change. There is evidence for a local adaption to winter and late spring frost in F. sylvatica, showing stronger signs of local adaptation in marginal than central populations giving them the ability to cope with climate change due to higher survival rates in the temperature elevated treatment of those populations originating from initially colder conditions (Kreyling et al. 2014). Similarly, Pinus contorta showed elevated growth in warmer conditions in their marginal (cooler) populations which leads to the assumption that they would cope better with climatic changes than central populations which are having to withstand higher temperatures and increased aridity. However, this aridity may counteract the positive effects of increased annual temperatures as trees are more sensitive to inter-annual climate fluctuations, leaving those in warmer central locations to be more robust to climatic changes after all. The potential impact of elevated atmospheric CO2 and water efficiency was not investigated in this study. These contradicting findings by McLane et al. (2011) show how difficult it is to find an overall conclusion on the effect of climate change on terrestrial plants even on species levels.
Limnic populations
A lack of studies focussing on the eastern, western and altitudinal studies of limnic animals and plants (Table 1, Fig. 2B) may cause a wide-reaching misinterpretation of data sets investigating the direct effects of climate change on those populations inhabiting these critical yet understudied populations. A disproportionally high number of studies researched animal populations but not plant or algae representatives. This shows the need for enhanced research on missing habitats and species representatives of the Plant Kingdom.
Animals
Negative effects on animals
The IPCC (2014) already concluded that freshwater fish populations in streams and rivers on the margin of their geographic distribution will likely be the first populations to respond to climatic induced effects due to the high rate of air–water heat transfer in their ecosystems. Multiple negative effects of climatic changes have been recorded for Limnic, or freshwater, animals. For example, a consistent relationship between abundance and temperature has been found in the fish species Salvelinus confluentus within the United States, throughout their southern margin with less fish in warmer waters. (Dunham et al. 2003).
Additional negative effects have been reported for the freshwater dependent damselfly Coenagrion scitulum, which showed a 31% increase in larval activity and higher growth rates in marginal northern compared to central populations (Therry et al. 2014a). Larger larvae had a higher mortality rate compared to smaller larvae. This leads to a higher energy demand due to higher activity and growth rates, and subsequently to higher mortality and predation rates in the northern margin compared to central populations (Therry et al. 2014a). Therry et al. (2014b) investigated the immune functions, body condition and flight-related morphology in C. scitulum at the northern front of expansion and found that females had a lower body condition, indicating more stressful conditions at the marginal population compared to the centre populations. They determined a higher flight muscle ratio at the northern expansion front, indicating a strong selection for dispersal ability and therefore range expansion, despite the counteracting effect of a shorter growth season.
Loulida et al. (2019) revealed a limitation in metabolic, osmo- and iono-regulation capacities on the southern-most distribution margin of the freshwater turtle Mauremys leprosa saharica in relation to drought and water salinization. This causes a decline in the effectiveness of organismal processes, leading to reduced feeding rates, body weight and therefore body condition in specimens exposed to increased salinity and drought conditions compared to control conditions. This ultimately leads to an increased vulnerability of southern marginal populations to predicted future environmental conditions caused by anthropogenic activities and climate change.
Positive effects on animals
Some freshwater animals have evolved to cope with climatic changes. Metabolic rates were positively correlated to temperature in southern populations of the South American freshwater fish Galaxias platei suggesting that this species adaptation to climatic changes is more dependent on its prior thermal history than its thermal tolerance. This species could be better at maintaining homeostasis at elevated temperatures leaving them with more energy available to spend on growth (Barrantes et al. 2017). Since a decline in marginal populations won’t be consistent across all freshwater bodies, it is important to conserve a minimum amount of populations across the peripheral range. After all, larger populations in higher quality habitats and more heterogeneous streams are likely to withstand climate change induced effects such as drought, flooding and elevated temperatures (Haak et al. 2010).
Plants
Negative effects on plants
In plants associated with freshwater systems, including wetland and swamp plants, populations of Decodon verticillatus at the northern margin of their species distribution, produced none or little amounts of seeds while most central populations produced abundant seeds. This low seed production in peripheral populations has been reported for multiple plant species (e.g. Sculthorpe 1967; Hutchinson 1975), however, comparisons with central populations or establishments of connections with ecological or genetic factors have rarely been performed in these studies. Dorken and Eckert (2001) found a strong association between low seed production and limiting sexual recruitment of D. verticillatus at its northern range has been found. The authors suggest that temperature may likely be a major determinant of sexual reproduction in northern populations of this species.
Positive effects on plants
No studies reporting positive effects of climate change on limnic plants on their range margin were found during this literature research.
Intertidal and marine populations
A lack of studies focussing on the eastern, western and depth margins of intertidal and marine plants and algae (Table 1, Fig. 2C) may cause an unsatisfactory interpretation of data sets. Out of the three major habitat classifications (terrestrial, limnic, intertidal and marine), marine studies present the most comprehensive information and number of studies and representation of a wide range of species. However, in all aquatic environments, the amount of research undertaken on plants and algae are far behind what is produced for animal populations. Studies mainly focused on adult individuals with some exceptions researching the effect on echinoderm and crustacean larvae. No studies on marine algae or plants focussed on eastern or western marginal populations or depth gradients.
Intertidal species inhabit the shores along coastlines where frequent increases and decreases of water levels due to tidal influence prevail. Therefore as part of the daily cycle, these organisms are exposed to additional abiotic and biotic stress, such as exposure to air, direct sunlight, elevated temperatures, elevated salinity, strongly fluctuating pH and oxygen levels, wind forces, wave action and many different predators, e.g. birds (reviewed by Hampe and Petit (2005)). Due to the additional stress, microclimatic variations in the intertidal zone are immense, in addition to predicted climatic changes, this may influence intertidal organisms in their physiology, structure and therefore, their distribution (Helmuth et al. 2002). Rock pools represent another extraordinary variable habitat within intertidal environments. Experiencing these conditions might make rock pool organisms more resilient to changing abiotic environmental conditions due to a potentially better adaptation and acclimatisation capacity (Brodie et al. 2013). The coastal shores, therefore, entail highly climatically affected, exposed and influenced ecosystems, especially for marginal populations.
Animals
Negative effects on animals
Within the studies conducted, negative impacts were among the most dominant findings. Sagarin et al. (2006) found that populations near any range margin might show both, relatively low abundance and depressed organismal conditions, reflected in negative growth, stress resilience and reproduction success. They concluded that the distribution of species appears to be more complicated and interesting than previously assumed. Negative effects on populations at the northern extent of sea turtles, like the sea turtle Caretta caretta, were described in Hawkes et al. (2007), here temperature-induced negative effects on physical limits as well as the sex ratio of these animals that is determined by the temperature experienced by eggs during incubation. For the turtle Dermochelys coriacea a critical isotherm line was determined of which this species is hindered to spread further south and is getting encapsulated. This isotherm moved northern wards forcing the sea turtles to follow, at the same time these turtles cannot withstand colder temperatures for a long time due to metabolic effects on the organism, hindering their expansion into the subpolar and Polar Regions and ultimately thinning the species range (McMahon and Hays 2006).
Additional negative effects have been observed for intertidal crabs along their gradient of distribution from the tropic to the southern end of the southern temperate zone (resembling colder conditions). The crab Uca urvillei showed little changes throughout their distributional range, however, the mangrove crab Perisesarma gattatum showed adaptation to local conditions. This suggests that both central and southern marginal populations of U. urvillei in the southern hemisphere seemed not to be impaired by elevated temperatures, which translates into increased oxygen consumption, during aerial respiration. Additionally, this species displays a highly efficient gas exchange between air and blood, making their gills a successful terrestrial adaptation, similar to lungs (Fusi et al. 2015). This is the same case for the central populations of P. guttatum, even though the southern marginal populations of this species can sustain this response as well, they can only do so with difficulties and at a high cost of metabolic expenditure (Fusi et al. 2015). Another fiddler crab species, Uca pugnax—which cold-temperature adapted larvae were cultured in multiple temperature conditions, including colder conditions than experienced at their present northern range margin—were not able to complete development at the lower thermal limit for development (Sanford et al. 2006), indicating potential resilience but only if temperature thresholds are met. In the barnacle Semibalanus balanoides southern margin populations death rates were heavily affected by elevated temperature which even overrides the negative effect of decreased pH (Findlay et al. 2010a) and increased mortality in intertidal zones when exceeding temperatures above 10 °C (Wethey et al. 2011; Jones et al. 2012). In the early life stages of S. balanoides, this effect accumulates to a reduction in survival of ~ 25%, significantly altering the population level at the southern limit (Findlay et al. 2010a). As for the northern margin of S. balanoides, the growth and development of larvae were negatively impacted by pH but not temperature compared to larvae of central populations. Individuals exposed to low pH were smaller and grew slower compared to control conditions (Findlay et al. 2010b). In another intertidal species, southern margin populations of the sea urchin Centrostephanus rodgersii in the southern hemisphere, displayed reduced larva survival in elevated temperature conditions, indicating a threshold temperature above which growth, productivity and reproduction of these urchins are severely limited with indications (Ling et al. 2008; Pecorino et al. 2014). In two other sea urchin species, Loxechinus albus and Pseudoechinus magellanicus, a combined negative effect of elevated temperature and reduced food availability due to changing environments in southern margin populations in the southern hemisphere was found (Détrée et al. 2020). The authors conclude that this may ultimately lead to an inability to acclimate to reduced food availability in future climate conditions and an effect on populations fitness of both species resulting in ecological and economic consequences.
The same negative relationship between recruitment success and elevated temperature as in the sea urchin C. rodgersii has also been shown for the gastropod Turbo torquatus (Wernberg et al. 2008, 2011). Other negative responses have been reported in mollusc species such as various limpets and Marcoma baltica, Mytilus ssp. and Mytilus galloprovincialis showing negative metabolic effects in these species not only on their marginal populations, due to a gradual reduction in conditioning index, starvation, adult mortality and recruitment failure on both margins (Wethey et al. 2011), but also on the individuals living in the centre of their distribution (Gilman 2006; Jansen et al. 2007a, b; Jones et al. 2010). The gastropod Littorina littorea exhibit greater shell dissolution and the inability to upregulate their metabolism after individuals of both, northern and southern marginal populations had been exposed to ocean acidification treatments making them more sensitive to predation and mechanical forces than central populations in the same study and treatment (Calosi et al. 2017). Another species of gastropod, Littorina obtusata, displayed a decreased antioxidant response in individuals of marginal populations, potentially leading these species to increased cellular damage (Cardoso et al. 2017). A combination of stressors on marine grazer-seaweed interactions between A. nodosum and L. littorea and L. obtusata in marginal habitats demonstrated how increased temperature and pCO2 in the seawater negatively influences the grazer-seaweed interaction due to an unbalance between the periwinkle mortality and consumption rates (Cardoso et al. 2017). In addition to the studies on molluscs above, it has been shown for the endangered abalone Haliotis kamtschatkana that in the southern part of the species range, studies suggest that increasing climate change results in population decline due to undersaturated aragonite saturation states (Crim et al. 2011). The limpet Nacella concinna and clam Laternula elliptica showed lower critical thermal and lethal limits in their northern populations in the Southern Ocean when originated from warmer, more northern, compared to colder, more southern, environments suggesting lower temperature tolerances in future, warmer conditions when already living in a warmer environment today (Morley et al. 2009). Additionally, experiments have shown that these two species with a survivable temperature envelope between 5 and 12 °C suffer from a 50 to 100% decrease in essential biological activity when exposed to 9–10 °C and 5 °C conditions, respectively, for a few days (Peck et al. 2004). It is also mentioned that the Antarctic scallop Adamussium colbecki loses its ability to swim if temperatures increase by 3 °C or more. This means that for polar marginal populations this failure of activity might cause species extinction.
Further negative effects have been found for southern margin populations of the Crustacea Chthamalus montagui and Chthamalus stellatus, the Mollusc Patella rustica and the Polychaeta Diopatra spp. For these species, Wethey et al. (2011) discovered adult mortality and recruitment failure after an extraordinarily cold winter in the Bay of Biscay.
Concerning fish, Valiente et al. (2010) forecasted the rapid decline in southern margin salmon populations leaving them extremely endangered or even becoming extinct. Species of bottom-dwelling fish in the North Sea revealed vertical instead of a horizontal, geographical shift in response to changing environmental conditions (Dulvy et al. 2008) which can be seen as the analogue to elevation, upward shifts in terrestrial species. In northern marginal populations of Platichthys flesus a decrease in size and condition if specimens were observed not only due to climatic changes but also as a result of anthropogenic impacts such as eutrophication, homogenisation of habitats, hypoxia etc. (Jokinen et al. 2015). Similar results have been shown by Pédron et al. (2017a) where, in common garden experiments between central and southern populations, the southern margin population did not grow under cold conditions, exhibited a low activity in producing defence metabolites to reduce e.g. oxidative damage and showed reduced muscle activity compared to the centre population. This suggests that southern margin populations experience a lower capacity to cope with environmental changes. Pédron et al. (2017b) examined the proteomic response of this species and found that liver enzyme regulation of southern population individuals could lead to a higher capacity to cope with future climate change like elevated temperature and hypoxia relative to central populations. These two contradicting findings represent the complexity to predict how future climatic changes influence the physiology of marine organisms.
In an extreme environment, the Arctic polar bears Ursus Maritimus have already suffered alarming declines at both latitudinal boundaries of their geographic distribution. For instance, at their southern boundary (Hudson Bay), they are steadily declining in numbers and body weight (Stirling et al. 1999) due to the reduction in the duration of sea-ice cover. This is causing starvation and living off of their fat reserves since an ice cover is necessary for hunting and feeding. In Antarctic penguins, a long life span inhibits microevolution and potential adaptation to rapid warming consequently leading to dispersal rather than adaptation responses to climatic changes (Forcada and Trathan 2009). Species showing low adaptability to changing environments, for instance, ice-obligates, may be more affected by the need to disperse rather than adapt to climate change and may struggle to find new suitable habitats This already led to a decline in breeding pairs by 50–90%, depending on the species of penguin (Smith et al. 1999; Croxall et al. 2002; Ainley et al. 2003; Forcada and Trathan 2009). This pattern of abandoning breeding grounds and dispersal can be tracked back with paleoecological records as a common response to climatic changes (e.g. Emslie et al. 1998; reviewed by Smith et al. (1999)) implying that the best option for understanding adaption in penguins is the study of phenological change (Berteaux et al. 2004; Visser 2008).
Research around seabirds is emphasizing the need to consider climate change along with other human-related threats. One example is Cepphus grylle of the auk family, at the southern margin of its global distribution occurring in the Baltic Sea region. It is already listed as ‘vulnerable’ to ‘near threatened’ at their southern margin (IUCN Red List Committee 2013) due to additional stress from oil spills, marine pollution and bycatch (Buchadas and Hof 2017 and references therein). The shift in newly arriving predators and prey populations’ abundance and distribution, such as the Pholis gunnellus and the Ammodytes tobianus, which are correlated to SST (surface seawater temperature) and plankton abundance, have an additional negative effect on this birds’ population sizes (Qasim 1956; Cramp et al. 1985; Gaston and Jones 1998; Arnott and Ruxton 2002). Direct effects of temperature and precipitation increases on C. grylle was reflected in their choice of breeding site (Buchadas et al. 2017) similarly, a different species within the auk family, Uria lomvia, has advanced its egg-laying date only at its southern boundary with no changes at its northern boundary with increasing temperature and subsequently a reduction in sea-ice cover (Gaston et al. 2005).
Positive effects on animals
Only very few positive effects of climatic changes on marine animals have been found to date. Almost nothing is known about larval connections along gradients, in particular hermaphroditic corals in small marginal populations. It is thought that they may show increased reliance on self-fertilisation and asexual reproduction like fragmentation due to the lack of external recruitments (Ayre and Hughes 2004; Baums et al. 2014). It is also suggested that warming waters in higher latitudes may favour the range expansion for corals since warmer temperatures could lead to settlement in suitable habitats (Laffoley and Baxter 2016). Contrary to the sea urchin C. rodgersii which displayed reduced larva survival in elevated temperature conditions, another sea urchin Strongylocentrotus purpuratus showed no distinct negative pattern in reproduction at the southern boundary of the species distribution when exposed to elevated temperature. This could mean that this species reproductive success might be influenced by food and nutrient supply rather than climatic changes (Lester et al. 2007). Other studies on C. rodgersii looking at the newly inhabited region of Tasmania and New Zealand as the new southern boundary, confirmed findings that elevated temperature in old southern margin populations reduces reproductive success and that conditions in the new habitats in colder waters are environmentally more favourable for this species (Ling et al. 2008; Pecorino et al. 2014). Changes in pH values, however, did not show an impact on fertilisation in both, old and new southern margins, and only a slight decrease in succeeding developmental stages in both (Ling et al. 2008; Pecorino et al. 2014). Additionally, limiting the effects of temperature in southern marginal populations of the limpet Patella vulgata can likely be traced back to microtopography rather than being related to latitude. Drastic environmental changes may influence this species indirectly by the availability of these microrefugia (Lima et al. 2016).
Additional positive effects regarding elevated recruitment rates and decreased mortality were found in northern margin populations of the Polichaeta Arenicola marina (Wethey et al. 2011), potentially expanding this species southern range margin.
In birds, Munilla et al. (2007) pointed out that a population decline of Uria aalge should be related to adult survival, associated with a decline in prey fish, instead of reproduction difficulties, suggesting conserving populations regarding anthropogenic threats that may cause the decline in adult survival. The southern-margin population of this species is somewhat prone to extinction as a consequence of additional impacts of climate change. Munilla et al. (2007) illustrate that linking this species decline at its southern rear margins with climate change could be misleading as it is most likely caused by anthropogenic, careless, particularly human impacts.
Other positive and at the same time highly variable adaptions were verified in the saline water bug Sigara selecta, which showed higher metabolic rates, higher reproduction and positive oviposition within the marginal compared to the central populations. However, the egg size was 27% smaller in northern marginal compared to central populations and also smaller in warmer and lower saline conditions. Due to stress, an increase in metabolic rates was found in this bug which is positively correlated with salinity and temperature increase in northern marginal populations but not central populations (Carbonell et al. 2017). This shows again that peripheral populations may be better adapted to prevailing unfavourable conditions but perform poorly when exposed to other conditions, such as increased temperature or acidification, associated with climate change (Hoffmann and Blows 1994).
Plants and algae
Negative effects on plants and algae
In addition to the negative impacts of climatic changes on intertidal and marine animals, even more negative impacts are found in algae. Due to climate change, intertidal benthic algae communities are predicted to change and disappear in the southern part of the northern hemisphere by 2200 (Kuffner et al. 2008; Jueterbock et al. 2014). The resulting habitat loss could be extenuated if algae can physiologically acclimatize to elevated temperatures. Responses of intertidal organisms can be considered as an early warning (Pearson et al. 2009) due to species already living at their maximum thermal tolerance and therefore low potential in responding to increased climatic stressors over the next century (Somero 2010; Tomanek 2010). Influences of climatic changes as well as anthropogenic impacts, such as trampling, on Ascophyllum nodosum were found to be detrimental in the southern range of this species distribution. It was unable to re-cover the locations and return to the pre-disturbed state (Araújo et al. 2009). Araújo et al. (2015) revealed that southern marginal populations of A. nodosum showed increased investment in reproduction (i.e. the number of receptacles per thallus) compared to relatively higher investments in defence (i.e. phlorotannin content) in central populations signifying that reproduction and defence have different fitness values for this species. The authors suggest a possible shift in the fitness values due to climatic changes leaving southern marginal populations with increased investment into defence rather than reproduction causing negative long-term consequences in population growth. In the southern populations of other brown algae: Fucus serratus, Fucus vesiculosus and Fucus spiralis, a maladaptation to desiccation and heat stress caused the southern margin populations to be less resilient compared to central populations. Marginal populations of these species also showed a reduced capacity to recover back to pre-stress levels of Fv/Fm, a chlorophyll fluorescence measuring parameter used to test stress and effects on the photosystem II (PSII). This may be either due to damage during the stress or less efficient or impaired PSII repair (Kim et al. 1993; Pearson et al. 2009; Ferreira et al. 2014). These results suggest that ongoing climate change and the stress it forces on Fucus southern marginal population may threaten small, fragmented marginal populations due to reduced fitness and lower adaptive capacity compared to central populations (Pearson et al. 2009). For F. vesiculosus, a climate-correlated diversity loss has been revealed by Nicastro et al. (2013), predicting a southern range contraction in the marine realm. Such elimination of F. vesiculosus is likely to cause other immediate ecological effects. Thus, its large-scale disappearance may modify and decrease ecosystem complexity, reducing the diversity and abundance of associated species with potential effects on all trophic levels (Nicastro et al. 2013). A comparison between F. vesiculosus and the seagrass Zostera marina by Mota et al. (2018) and their southern and northern margin populations showed higher thermal resistance and recovery in southern margin populations of F. vesiculosus and an opposing finding for northern and southern margin populations of Z. marina. This confirms that differentiation of thermal stress responses can arise between both marginal populations.
The macroalgae and indicator species F. serratus has created some adaptive responses to climatic changes, however, reproduction capacity at the southern margin of its species distribution got severely diminished compared to central populations due to unfavourable abiotic conditions linked to either climate change or short-term extreme environmental fluctuations (Viejo et al. 2011). Compared to northern margin populations, populations on the southern range margin like the ones of the seaweed F. serratus, have only recently gained the interest of the research community (review by Hampe and Petit (2005), Araújo et al. (2011), Ferreira et al. (2014) and Zardi et al. (2015)). In the brown algae Hormosira banksii specimens from northern margin populations in the southern hemisphere showed increased thermal sensitivity indicated by decreased photosynthetic yields of the PSII and delays in recovery after desiccation stress, contrasting studies of Ferreira et al. (2014), Saada et al. (2016) and Clark et al. (2018). Negative effects have also been found for southern populations of the red algae Corallina officinalis in the NE Atlantic, downregulating metabolic processes and failing to adapt to changing temperature conditions (Kolzenburg et al. 2019).
High irradiances of photosynthetic active radiation (PAR) and ultraviolet radiation (UVR) during low tide can cause photoinhibition and is relatively common in macroalgae in intertidal zones (Martínez et al. 2012). The negative effect also showed an increasing trend under elevated temperatures in intertidal central fucoids (Altamirano et al. 2003). This is not only shown for central populations, but a study by Martínez et al. (2012) suggests that the three additive stressors (air and ocean temperatures and solar radiation) are highly relevant to predict F. serratus range shifts on southern margins.
In addition to elevated temperature, which most of the studies analysed in this review focussed on, ocean acidification (OA) is a second key factor caused by anthropogenic climate change. It directly affects calcareous organisms due to the lowering of the calcium carbonate saturation state (Ω = [Ca2+] × [CO32−]/Ksp) in the seawater (Orr et al. 2005; Doney et al. 2009). This translates into a decrease in the Mg content of calcite hard parts in most marine environments. Additionally, the proportion of calcifiers depositing low-Mg-calcite will increase and the average Mg content of organic carbonate sediments is decreasing due to dissolution (Andersson et al. 2008). A study looking at natural CO2 seeps and acidified seawater in today’s oceans revealed that, generally, calcifying macroalgae like coralline algae will be severely impacted and are the first calcifying organism to disappear in these areas (Kleypas et al. 2006; Hall-Spencer et al. 2008; Kroeker et al. 2013). The physiological response to OA, especially regarding calcification, is very variable amongst species (Ries et al. 2009). OA is proved to cause even more severe effects on whole benthic community structures, especially in shallow warm-water carbonate ecosystems, such as the tropics and sub-tropics (Kuffner et al. 2008). Antarctic and tropical macroalgae in coastal waters appear to be the most vulnerable to warming, (IPCC 2014) resulting in a negative change in the total oceanic production, of which coastal macrophytes coverage supplies 2–5%. Species able to tolerate high fluctuations in pCO2 levels might therefore be resilient to OA (Egilsdottir et al. 2013). An increase of 3 °C in SST is likely to cause an increase in Mg content in C. officinalis and Corallina caespitosa around 1.1 mol% Mg-calcite, which is corresponding to ~ 32% of the seasonal variability of Mg content in these species in the NE Atlantic (Williamson et al. 2014) and will make them more susceptible to OA.
Positive effects on algae
In algal photosynthesis, both CO2 and bicarbonate can be used as a substrate for photosynthesis, reducing the effects of OA (Raven and Beardall 2003; Galbraith et al. 2002). Additional studies on non-marginal populations have shown how a doubling in pCO2 concentration in seawater can have a positive effect on red temperate, non-calcifying (Kübler et al. 1999) and calcifying algae species, such as Corallina pilulifera (Gao et al. 1993). Primary producers relying exclusively on CO2 are expected to enhance photosynthesis rates with elevated CO2 availability (Kübler et al. 1999) and may outcompete algae that cannot downregulate their carbon concentration mechanisms (Hepburn et al. 2011; Cornwall et al. 2012; Raven et al. 2012). A southern marginal population of A. nodosum showed a higher reproductive output in another study conducted by Araújo et al. (2011) and showed similar mortality, however, specimens were smaller and populations denser due to increased energy costs channelled into reproduction or defence. In the same study, F. serratus showed a distribution into narrower vertical habitat ranges. For both species at the southern margin of their distribution in the NE Atlantic, they found different patterns of potential adaptations to environmental changes (Araújo et al. 2011). In a comparison between two Fucus species, F. vesiculosus and Fucus radicans, in the Baltic Sea, featuring their northern margin of the species distribution and limit of salinity tolerance, prevention of receptacle formation was exhibited in both Fucus species under climate change scenario, suggesting that Fucus failed to reproduce sexually, whereas photosynthesis and growth were not affected. This suggests that these marginal populations may be resilient to future conditions, including reduced salinity and elevated temperatures, but only if they achieve to reproduce asexually (Rothäusler et al. 2018; Rugiu et al. 2018). Another study on F. vesiculosus suggests that southern range populations may be more resilient to heat stress than their northern counterparts, whereas growth rates in this study did not change significantly in both populations (Saada et al. 2016). Similarly, a better adaptation and tolerance to temperature shocks was also found in southern marginal populations of the kelp Laminaria digitata (King et al. 2019). This was suggested due to a greater upregulation of the heat shock protein hsp70 in central compared to southern marginal populations which leads the authors to suggest the existence of distinct thermal ecotypes of this species and a potentially positive and more complex response to future water temperatures.
The little understanding of how stressors interact, and which influence they might have on organisms is limiting the capacity of scientists to predict the effects of climate change (Martínez et al. 2012) and requests for more physiological studies.
Conclusions
A great shortage of research relating to the effect of climate change on marginal populations was found. Most studies were conducted in Europe, followed by North America. An even greater, astonishing lack of knowledge has been uncovered for the rest of the continents (Fig. 1). A summary of all relevant studies reviewed herein and a division in habitat, kingdom and classes can be found in Fig. 3. The total number of studies found, and species investigated herein are summarised in Fig. 4 and Table A1 in the Supplementary online content. We present the finding that significantly less literature is focussing on limnic (4.9%) compared to terrestrial (32.5%) or marine (62.6%) environments. This highlights the urgent need for more research on the direct effects of climate change on particularly limnic organisms living on the margin of their species distribution.

Summary of studies on marginal populations. Habitats (top: terrestrial, bottom: aquatic) are divided into kingdoms (left: fauna, right: flora (top: plants, bottom: plant and protists)) and species classes. For each species class percentages of the total number are given in the corresponding section. Legends begin at the top of each chart and continue clockwise

Total number of species (A) and studies (B) focussing on direct effects of climate change on marginal populations
Having laid out the complexity and details in the introduction, when researching species responses, it is crucial to compare marginal and central populations in order to not over-estimate the margins of the species distribution and have precise forecasts on how these species could be influenced in the future. In addition to one or a combination of the hypotheses presented in the introduction, looking at variances in distribution, density, genetic diversity or environmental conditions, we agree with Munilla et al. (2007) who clearly state that linking species declines, especially at their southernmost margins of distribution, to climate change, could potentially be misleading. This is because it is most likely caused not only by climatic impacts but also by anthropogenic, careless, particularly human impacts, adding an additional stressor to the list of influential factors on all but especially marginal populations.
Another aspect that is leading to a more and more alarming and dramatic picture of the direct effects of climatic changes on marginal populations results from publishing bias. There is limited published information of neutral or no direct effects on these populations.
An aspect that was tried to be included but proved difficult was the absence of a clear north-to-south gradient in polar species. Due to this fact, it is also difficult for polar species to shift their distribution in the future if necessary, leading to a thinning of their range distribution and a contraction in east/west instead of north/south directions to other areas.
Summary
We analysed 93 studies on terrestrial (n = 30), limnic (n = 7) and marine (n = 56) marginal populations including a total of 123 species of animalia (n = 89), plantae (n = 25) and algae (n = 9) across the globe (Figs. 2A and 3). Most of the studies focused on the marine, followed by the terrestrial environment. Only one publication exists on marginal populations of a purely aquatic plant. Several experiments are dealing with elevated temperature or CO2 in algae. However, only one paper on marginal populations focussed on red algae, none on green algae and the remaining studies researched brown algae. Within the aquatic animals, a good distribution between vertebrate and invertebrate organisms was found resulting in a decent overview of the direct effects of climatic changes on marine animals.
Little evidence of the effects of climatic changes on freshwater organisms was found. Only four studies were conducted in freshwater and no studies on marginal populations of invertebrates.
Within the category of terrestrial plants, mostly species of the clade angiosperms and only three studies on two species of gymnosperms were conducted. Further two studies on bryophytes within the terrestrial environment were performed. Within the category of terrestrial fauna, studies are split evenly between vertebrates and invertebrates. Studies on terrestrial flora presented multiple aspects, this included altitude and an east–west rather than a north–south marginal approach for studies on the Antarctic peninsula.
Marine studies on seaweeds originate from a limited number of institutes, presenting a bias to a certain extent.
Additional bias is introduced by the availability and accessibility of organisms, this results in an uneven distribution of studies representing flora and fauna independent of the ecosystem but dependant on accessibility, mobility and throughput (e.g. trees: extended and long-term studies). More studies focus on the northern than the southern margin of a species distribution; and only a few studies have focussed on both range margins and the centre populations in the same study (n = 13, Fig. 5). We imperatively suggest studies should focus on all populations, margins and centres. However, if a choice needs to be made due to restrictions, investigations should focus on the southern margin as it is predicted to be at the forefront of direct impacts of climatic changes (Pinsky et al. 2019) with especially elevated temperature having a greater effect than ocean acidification.

Summary and number of studies investigating the northern and southern range margins and the centre of the respective species latitudinal distribution in the same study for the same species
Overall, our results indicate that direct positive and negative effects related to climate change are already extensive, although levels of climate change thus far are fairly small relative to those predicted for the future. Especially the negative effects are potentially becoming much more frequent in correlation with global climate change.
Gaps out of summary points
During our research, we discovered some important gaps in the literature and the writing of publications. We found a great need to clearly specify which geographically marginal or peripheral populations were researched and a brief statement of the current boundary of the species distribution would aid understanding of the location. Additionally, we found a disproportion in numbers of taxonomic groups, depending on for which funding was available, resulting in a greater representation of e.g. fish than insect species. Regarding direct effects, more elevated temperature than CO2 studies were conducted, potentially due to convenience. This makes it challenging to predict future synergetic effects on marginal populations as a combination of both interrelated key factors of climate change.
Availability of data and material
A detailed table including all analysed studies and their classifications can be found at: https://doi.org/10.6084/m9.figshare.14485059.
Code availability
Not applicable.
References
Ainley DG, Ballard G, Emslie SD, Fraser WR, Wilson PR, Woehler EJ (2003) Adelie penguins and environmental change. Science 300(5618):429–430
Aitken SN, Yeaman S, Holliday JA, Wang T, Curtis-McLane S (2008) Adaptation, migration or extirpation: climate change outcomes for tree populations. Evol Appl 1(1):95–111
Altamirano M, Flores-Moya A, Figueroa FL (2003) Effects of UV radiation and temperature on growth of germlings of three species of Fucus (Phaeophyceae). Aquat Bot 75(1):9–20
Andersson AJ, Mackenzie FT, Bates NR (2008) Life on the margin: implications of ocean acidification on Mg-calcite, high latitude and cold-water marine calcifiers. Mar Ecol Prog Ser 373:265–273
Angert AL (2006) Demography of central and marginal populations of monkeyflowers (Mimulus cardinalis and M. lewisii). Ecology 87(8):2014–2025
Araújo RM, Vaselli S, Almeida M, Serrão E, Sousa-Pinto I (2009) Effects of disturbance on marginal populations: human trampling on Ascophyllum nodosum assemblages at its southern distribution limit. Mar Ecol Prog Ser 378:81–92
Araújo R, Serrão EA, Sousa-Pinto I, Åberg P (2011) Phenotypic differentiation at southern limit borders: the case study of two fucoid macroalgal species with different life-history traits 1. J Phycol 47(3):451–462
Araújo RM, Serrao EA, Sousa-Pinto I, Aberg P (2014) Spatial and temporal dynamics of fucoid populations (Ascophyllum nodosum and Fucus serratus): a comparison between central and range edge populations. PLoS ONE. https://doi.org/10.1371/journal.pone.0092177
Araújo R, Serrão EA, Sousa-Pinto I, Arenas F, Monteiro CA, Toth G et al (2015) Trade-offs between life-history traits at range-edge and central locations. J Phycol 51(4):808–818
Arlettaz R, Schaad M, Reichlin TS, Schaub M (2010) Impact of weather and climate variation on Hoopoe reproductive ecology and population growth. J Ornithol 151(4):889–899
Arnott SA, Ruxton GD (2002) Sandeel recruitment in the North Sea: demographic, climatic and trophic effects. Mar Ecol Prog Ser 238:199–210
Ashton S, Gutierrez D, Wilson RJ (2009) Effects of temperature and elevation on habitat use by a rare mountain butterfly: implications for species responses to climate change. Ecol Entomol 34(4):437–446
Ayan S, Turfan N, Yer EN, Šeho M, Barş Özel H, Ducci F (2018) Antioxidant variability of the seeds in central and marginal populations of taurus cedar (Cedrus libani A. Rich). Šumarski List 142(11–12):600
Ayre DJ, Hughes TP (2004) Climate change, genotypic diversity and gene flow in reef-building corals. Ecol Lett 7(4):273–278
Barrantes ME, Lattuca ME, Vanella FA, Fernández DA (2017) Thermal ecology of Galaxias platei (Pisces, Galaxiidae) in South Patagonia: perspectives under a climate change scenario. Hydrobiologia 802(1):255–267
Battisti A, Stastny M, Netherer S, Robinet C, Schopf A, Roques A, Larsson S (2005) Expansion of geographic range in the pine processionary moth caused by increased winter temperatures. Ecol Appl 15(6):2084–2096
Baums IB, Devlin-Durante M, Laing BAA, Feingold J, Smith T, Bruckner A, Monteiro J (2014) Marginal coral populations: the densest known aggregation of Pocillopora in the Galápagos Archipelago is of asexual origin. Front Mar Sci 1:59
Berteaux D, Réale D, McAdam AG, Boutin S (2004) Keeping pace with fast climate change: can arctic life count on evolution? Integr Comp Biol 44(2):140–151
Bertocci I, Araújo RM, Vaselli S, Sousa-Pinto I (2011) Marginal populations under pressure: spatial and temporal heterogeneity of Ascophyllum nodosum and associated assemblages affected by human trampling in Portugal. Mar Ecol Prog Ser 439:73–82
Bozinovic F, Calosi P, Spicer JI (2011) Physiological correlates of geographic range in animals. Annu Rev Ecol Evol Syst 42:155–179
Bradshaw WE, Holzapfel CM (2001) Genetic shift in photoperiodic response correlated with global warming. Proc Natl Acad Sci 98(25):14509–14511
Braschler B, Hill JK (2007) Role of larval host plants in the climate-driven range expansion of the butterfly Polygonia c-album. J Anim Ecol 76(3):415–423
Bridle JR, Vines TH (2007) Limits to evolution at range margins: when and why does adaptation fail? Trends Ecol Evol 22(3):140–147
Brodie J, Walker RH, Williamson CJ, Irvine LM (2013) Epitypification and redescription of Corallina officinalis L., the type of the g, and C. elongata Ellis et Solander (Corallinales, Rhodophyta). Cryptogam Algol 34(1):49–56. https://doi.org/10.7872/crya.v34.iss1.2013.49
Brown JH (1984) On the relationship between abundance and distribution of species. Am Nat 124(2):255–279
Brussard PF (1984) Geographic patterns and environmental gradients: the central-marginal model in Drosophila revisited. Annu Rev Ecol Syst 15(1):25–64
Buchadas ARC, Hof AR (2017) Future breeding and foraging sites of a southern edge population of the locally endangered Black Guillemot Cepphus grylle. Bird Study 64(3):306–316
Calosi P, Melatunan S, Turner LM, Artioli Y, Davidson RL, Byrne JJ et al (2017) Regional adaptation defines sensitivity to future ocean acidification. Nat Commun 8:13994. https://doi.org/10.1038/ncomms13994
Carbonell JA, Bilton DT, Calosi P, Millán A, Stewart A, Velasco J (2017) Metabolic and reproductive plasticity of central and marginal populations of the eurythermic saline water bug Sigara selecta (Hemiptera: Corixidae) in a climate change context. J Insect Physiol 98:59–66
Cardoso PG, Grilo TF, Dionísio G, Aurélio M, Lopes AR, Pereira R et al (2017) Short-term effects of increased temperature and lowered pH on a temperate grazer-seaweed interaction (Littorina obtusata/Ascophyllum nodosum). Estuar Coast Shelf Sci 197:35–44
Case TJ, Taper ML (2000) Interspecific competition, environmental gradients, gene flow, and the coevolution of species’ borders. Am Nat 155(5):583–605
Clark JS, Poore AGB, Doblin MA (2018) Shaping up for stress: physiological flexibility is key to survivorship in a habitat-forming macroalga. J Plant Physiol 231:346–355
Cornwall CE, Hepburn CD, Pritchard D, Currie KI, McGraw CM, Hunter KA, Hurd CL (2012) Carbon-use strategies in macroalgae: differential responses to lowered ph and implications for ocean acidification. J Phycol 48(1):137–144. https://doi.org/10.1111/j.1529-8817.2011.01085.x
Cramp S, Brooks DJ, Dunn E, Gillmor R, Hall-Craggs J, Hollom PAD et al (1985) The birds of Western Palearctic, vol 4. Oxford University Press, Oxford
Crim RN, Sunday JM, Harley CDG (2011) Elevated seawater CO2 concentrations impair larval development and reduce larval survival in endangered northern abalone (Haliotis kamtschatkana). J Exp Mar Biol Ecol 400(1–2):272–277
Croxall JP, Trathan PN, Murphy EJ (2002) Environmental change and Antarctic seabird populations. Science 297(5586):1510–1514
Crozier L (2003) Winter warming facilitates range expansion: cold tolerance of the butterfly Atalopedes campestris. Oecologia 135(4):648–656
Crozier L (2004) Warmer winters drive butterfly range expansion by increasing survivorship. Ecology 85(1):231–241
Davies NA, Gramotnev G, McAlpine C, Seabrook L, Baxter G, Lunney D et al (2013) Physiological stress in Koala populations near the arid edge of their distribution. PLoS ONE 8(11):e79136. https://doi.org/10.1371/journal.pone.0079136
Détrée C, Navarro JM, Font A, Gonzalez M (2020) Species vulnerability under climate change: study of two sea urchins at their distribution margin. Sci Total Environ 728:138850
Deutsch CA, Tewksbury JJ, Huey RB, Sheldon KS, Ghalambor CK, Haak DC, Martin PR (2008) Impacts of climate warming on terrestrial ectotherms across latitude. Proc Natl Acad Sci 105(18):6668–6672
Doney SC, Fabry VJ, Feely RA, Kleypas JA (2009) Ocean acidification: the other CO2 problem. Ann Rev Mar Sci 1(1):169–192. https://doi.org/10.1146/annurev.marine.010908.163834
Dorken ME, Eckert CG (2001) Severely reduced sexual reproduction in northern populations of a clonal plant, Decodon verticillatus (Lythraceae). J Ecol 89(3):339–350
Dulvy NK, Rogers SI, Jennings S, Stelzenmüller V, Dye SR, Skjoldal HR (2008) Climate change and deepening of the North Sea fish assemblage: a biotic indicator of warming seas. J Appl Ecol 45(4):1029–1039
Dunham J, Rieman B, Chandler G (2003) Influences of temperature and environmental variables on the distribution of bull trout within streams at the southern margin of its range. N Am J Fish Manag 23(3):894–904
Eckert CG, Samis KE, Lougheed SC (2008) Genetic variation across species’ geographical ranges: the central–marginal hypothesis and beyond. Mol Ecol 17(5):1170–1188
Egilsdottir H, Noisette F, Noël LM-LJ, Olafsson J, Martin S (2013) Effects of pCO2 on physiology and skeletal mineralogy in a tidal pool coralline alga Corallina elongata. Mar Biol 160(8):2103–2112. https://doi.org/10.1007/s00227-012-2090-7
Emslie SD, Fraser W, Smith RC, Walker W (1998) Abandoned penguin colonies and environmental change in the Palmer Station area, Anvers Island, Antarctic Peninsula. Antarct Sci 10(3):257–268
Etterson JR, Shaw RG (2001) Constraint to adaptive evolution in response to global warming. Science 294(5540):151–154
Ferreira JG, Arenas F, Martínez B, Hawkins SJ, Jenkins SR (2014) Physiological response of fucoid algae to environmental stress: comparing range centre and southern populations. New Phytol 202(4):1157–1172. https://doi.org/10.1111/nph.12749
Findlay HS, Burrows MT, Kendall MA, Spicer JI, Widdicombe S (2010a) Can ocean acidification affect population dynamics of the barnacle Semibalanus balanoides at its southern range edge? Ecology 91(10):2931–2940
Findlay HS, Kendall MA, Spicer JI, Widdicombe S (2010b) Relative influences of ocean acidification and temperature on intertidal barnacle post-larvae at the northern edge of their geographic distribution. Estuar Coast Shelf Sci 86(4):675–682
Forcada J, Trathan PN (2009) Penguin responses to climate change in the Southern Ocean. Glob Change Biol 15(7):1618–1630
Fusi M, Giomi F, Babbini S, Daffonchio D, McQuaid CD, Porri F, Cannicci S (2015) Thermal specialization across large geographical scales predicts the resilience of mangrove crab populations to global warming. Oikos 124(6):784–795
Galbraith H, Jones R, Park R, Clough J, Herrod-Julius S, Harrington B, Page G (2002) Global climate change and sea level rise: potential losses of intertidal habitat for shorebirds. Waterbirds Inter J Waterbird Biol 25(2):173–183
Gao K, Aruga Y, Asada K, Ishihara T, Akano T, Kiyohara M (1993) Calcification in the articulated coralline alga Corallina pilulifera, with special reference to the effect of elevated CO2 concentration. Mar Biol 117(1):129–132. https://doi.org/10.1007/BF00346434
Gaston K (2003) The structure and dynamics of geographic ranges. Oxford University Press, Oxford
Gaston AJ, Jones IL (1998) Bird families of the world: the auks. Oxford University Press, New York
Gaston AJ, Gilchrist HG, Hipfner JM (2005) Climate change, ice conditions and reproduction in an Arctic nesting marine bird: Brunnich’s guillemot (Uria lomvia L.). J Anim Ecol 74(5):832–841
Gilman SE (2006) The northern geographic range limit of the intertidal limpet Collisella scabra: a test of performance, recruitment, and temperature hypotheses. Ecography. https://doi.org/10.1111/j.0906-7590.2006.04572.x
Guo Q (2014) Central-marginal population dynamics in species invasions. Front Ecol Evol 2:23
Haak AL, Williams JE, Neville HM, Dauwalter DC, Colyer WT (2010) Conserving peripheral trout populations: the values and risks of life on the edge. Fisheries 35(11):530–549
Hall-Spencer JM, Rodolfo-Metalpa R, Martin S, Ransome E, Fine M, Turner SM et al (2008) Volcanic carbon dioxide vents show ecosystem effects of ocean acidification. Nature 454(7200):96
Hampe A, Jump AS (2011) Climate relicts: past, present, future. Annu Rev Ecol Evol Syst 42(1):313–333. https://doi.org/10.1146/annurev-ecolsys-102710-145015
Hampe A, Petit RJ (2005) Conserving biodiversity under climate change: the rear edge matters. Ecol Lett. https://doi.org/10.1111/j.1461-0248.2005.00739.x
Hawkes LA, Broderick AC, Godfrey MH, Godley BJ (2007) Investigating the potential impacts of climate change on a marine turtle population. Glob Change Biol 13(5):923–932
Helmuth B, Harley CDG, Halpin PM, O’Donnell M, Hofmann GE, Blanchette CA (2002) Climate change and latitudinal patterns of intertidal thermal stress. Science 298(5595):1015–1017
Hepburn CD, Pritchard DW, Cornwall CE, McLeod RJ, Beardall J, Raven JA, Hurd CL (2011) Diversity of carbon use strategies in a kelp forest community: implications for a high CO2 ocean. Glob Change Biol 17(7):2488–2497. https://doi.org/10.1111/j.1365-2486.2011.02411.x
Hill JK, Griffiths HM, Thomas CD (2011) Climate change and evolutionary adaptations at species’ range margins. Annu Rev Entomol 56:143–159. https://doi.org/10.1146/annurev-ento-120709-144746
Hoffmann AA, Blows MW (1994) Species borders: ecological and evolutionary perspectives. Trends Ecol Evol 9(6):223–227
Humphries MM, Thomas DW, Speakman JR (2002) Climate-mediated energetic constraints on the distribution of hibernating mammals. Nature 418(6895):313
Hutchinson GE (1975) A treatise of limnology. 3: Limnological botany. Wiley, New York
IPCC (2014) Climate change 2014: synthesis report. In: Core Writing Team, Pachauri RK, Meyer LA (eds) Contribution of working groups I, II and III to the fifth assessment report of the intergovernmental panel on climate change. IPCC, Geneva, Switzerland, p 151
IPCC (2021) Climate change 2021: the physical science basis. In: Masson-Delmotte V, Zhai P, Pirani A, Connors SL, Péan C, Berger S, Caud N, Chen Y, Goldfarb L, Gomis MI, Huang M, Leitzell K, Lonnoy E, Matthews JBR, Maycock TK, Waterfield T, Yelekçi O, Yu R, Zhou B (eds) Contribution of working group I to the sixth assessment report of the intergovernmental panel on climate change. Cambridge University Press, In Press
IUCN Red List Committee (2013) The IUCN Red List of Threatened Species™ Strategic Plan 2013–2020. Version 1.0. Prepared by the IUCN Red List Committee
Jansen JM, Pronker AE, Bonga SW, Hummel H (2007a) Macoma balthica in Spain, a few decades back in climate history. J Exp Mar Biol Ecol 344(2):161–169
Jansen JM, Pronker AE, Kube S, Sokolowski A, Sola JC, Marquiegui MA et al (2007b) Geographic and seasonal patterns and limits on the adaptive response to temperature of European Mytilus spp. and Macoma balthica populations. Oecologia 154(1):23–34. https://doi.org/10.1007/s00442-007-0808-x
Janzen DH (1967) Why mountain passes are higher in the tropics. Am Nat 101(919):233–249
Jokinen H, Wennhage H, Lappalainen A, Ådjers K, Rask M, Norkko A (2015) Decline of flounder (Platichthys flesus (L.)) at the margin of the species’ distribution range. J Sea Res 105:1–9
Jones SJ, Lima FP, Wethey DS (2010) Rising environmental temperatures and biogeography: poleward range contraction of the blue mussel, Mytilus edulis L., in the western Atlantic. J Biogeogr 37(12):2243–2259
Jones SJ, Southward AJ, Wethey DS (2012) Climate change and historical biogeography of the barnacle Semibalanus balanoides. Glob Ecol Biogeogr 21(7):716–724
Jueterbock A, Kollias S, Smolina I, Fernandes JMO, Coyer JA, Olsen JL, Hoarau G (2014) Thermal stress resistance of the brown alga Fucus serratus along the North-Atlantic coast: acclimatization potential to climate change. Mar Genom. https://doi.org/10.1016/j.margen.2013.12.008
Jump AS, Hunt JM, Penuelas J (2006) Rapid climate change-related growth decline at the southern range edge of Fagus sylvatica. Glob Change Biol 12(11):2163–2174
Kim JH, Nemson JA, Melis A (1993) Photosystem II reaction center damage and repair in Dunaliella salina (green alga) (analysis under physiological and irradiance-stress conditions). Plant Physiol 103(1):181–189
King NG, McKeown NJ, Smale DA, Wilcockson DC, Hoelters L, Groves EA, Stamp T, Moore PJ (2019) Evidence for different thermal ecotypes in range centre and trailing edge kelp populations. J Exp Mar Biol Ecol 514:10–17
Kleypas JA, Feely RA, Fabry VJ, Langdon C, Sabine CL, Robbins LL (2006) Impacts of ocean acidification on coral reefs and other marine calcifiers: a guide for future research, report of a workshop held 18–20 April 2005, St. Petersburg, FL, sponsored by NSF, NOAA, and the U.S. Geological Survey, p 88
Kolzenburg R, Nicastro KR, McCoy SJ, Ford AT, Zardi GI, Ragazzola F (2019) Understanding the margin squeeze: differentiation in fitness-related traits between central and trailing edge populations of Corallina officinalis. Ecol Evol 9(10):5787–5801
Kreyling J, Buhk C, Backhaus S, Hallinger M, Huber G, Huber L et al (2014) Local adaptations to frost in marginal and central populations of the dominant forest tree Fagus sylvatica L. as affected by temperature and extreme drought in common garden experiments. Ecol Evol 4(5):594–605
Kroeker KJ, Micheli F, Gambi MC (2013) Ocean acidification causes ecosystem shifts via altered competitive interactions. Nat Clim Chang 3(2):156–159. https://doi.org/10.1038/nclimate1680
Kübler JE, Johnston AM, Raven JA (1999) The effects of reduced and elevated CO2 and O2 on the seaweed Lomentaria articulata. Plant Cell Environ 22(10):1303–1310. https://doi.org/10.1046/j.1365-3040.1999.00492.x
Kuffner IB, Andersson AJ, Jokiel PL, Rodgers KS, Mackenzie FT (2008) Decreased abundance of crustose coralline algae due to ocean acidification. Nat Geosci 1(2):114–117. https://doi.org/10.1038/ngeo100
Kutz SJ, Hoberg EP, Nishi J, Polley L (2002) Development of the muskox lungworm, Umingmakstrongylus pallikuukensis (Protostrongylidae), in gastropods in the Arctic. Can J Zool 80(11):1977–1985
Kutz SJ, Hoberg EP, Polley L, Jenkins EJ (2005) Global warming is changing the dynamics of Arctic host–parasite systems. Proc R Soc B Biol Sci 272(1581):2571–2576
Laffoley DD, Baxter JM (2016) Explaining ocean warming: causes, scale, effects and consequences. IUCN Gland, Switzerland
Lawton JH (1993) Range, population abundance and conservation. Trends Ecol Evol 8(11):409–413
Lesica P, Allendorf FW (1995) When are peripheral populations valuable for conservation? Conserv Biol 9(4):753–760. https://doi.org/10.1046/j.1523-1739.1995.09040753.x
Lesica P, McCune B (2004) Decline of arctic-alpine plants at the southern margin of their range following a decade of climatic warming. J Veg Sci 15(5):679–690
Lester SE, Gaines SD, Kinlan BP (2007) Reproduction on the edge: large-scale patterns of individual performance in a marine invertebrate. Ecology. https://doi.org/10.1890/06-1784.1
Lewis Smith RI (1999) Biological and environmental characteristics of three cosmopolitan mosses dominant in continental Antarctica. J Veg Sci 10(2):231–242
Lima FP, Gomes F, Seabra R, Wethey DS, Seabra MI, Cruz T, Santos AM, Hilbish TJ (2016) Loss of thermal refugia near equatorial range limits. Glob Change Biol 22(1):254–263
Ling SD, Johnson CR, Frusher S, King CK (2008) Reproductive potential of a marine ecosystem engineer at the edge of a newly expanded range. Glob Change Biol 14(4):907–915
Loulida S, Znari M, Naimi M, Bendami S (2019) Tolerance to salinity and dehydration in the Sahara Desert blue-eyed turtle, Mauremys leprosa saharica (Testudines: Geoemydidae) from a brackish pond in the Lower Draa basin, southern Morocco. Afr J Herpetol 68(1):58–76
Martínez B, Arenas F, Rubal M, Burgués S, Esteban R, García-Plazaola I et al (2012) Physical factors driving intertidal macroalgae distribution: physiological stress of a dominant fucoid at its southern limit. Oecologia 170(2):341–353
APA Harvard Chicago Vancouver Mayr E (1963) Animal species and evolution. Harvard University Press, Cambridge, MA and London, England. https://doi.org/10.4159/harvard.9780674865327
McLane SC, Daniels LD, Aitken SN (2011) Climate impacts on lodgepole pine (Pinus contorta) radial growth in a provenance experiment. For Ecol Manag 262(2):115–123
McMahon CR, Hays GC (2006) Thermal niche, large-scale movements and implications of climate change for a critically endangered marine vertebrate. Glob Change Biol 12(7):1330–1338
Merrill RM, Gutiérrez D, Lewis OT, Gutiérrez J, Díez SB, Wilson RJ (2008) Combined effects of climate and biotic interactions on the elevational range of a phytophagous insect. J Anim Ecol 77(1):145–155
Meyer-Grünefeldt M, Belz K, Calvo L, Marcos E, von Oheimb G, Härdtle W (2016) Marginal Calluna populations are more resistant to climate change, but not under high-nitrogen loads. Plant Ecol 217(1):111–122
Morison JIL, Gifford RM (1984) Plant growth and water use with limited water supply in high CO2 concentrations. II. Plant dry weight, partitioning and water use efficiency. Aust J Plant Physiol 11:361–374
Morley SA, Hirse T, Pörtner H-O, Peck LS (2009) Geographical variation in thermal tolerance within Southern Ocean marine ectotherms. Comp Biochem Physiol Part A Mol Integr Physiol 153(2):154–161. https://doi.org/10.1016/j.cbpa.2009.02.001
Mota CF, Engelen AH, Serrao EA, Coelho MAG, Marbà N, Krause-Jensen D, Pearson GA (2018) Differentiation in fitness-related traits in response to elevated temperatures between leading and trailing edge populations of marine macrophytes. PLoS ONE 13(9):e0203666. https://doi.org/10.1371/journal.pone.0203666
Munilla I, Díez C, Velando A (2007) Are edge bird populations doomed to extinction? A retrospective analysis of the common guillemot collapse in Iberia. Biol Cons 137(3):359–371
Nakatsubo T (2002) Predicting the impact of climatic warming on the carbon balance of the moss Sanionia uncinata on a maritime Antarctic island. J Plant Res 115(2):99–106
Nicastro KR, Zardi GI, Teixeira S, Neiva J, Serrão EA, Pearson GA (2013) Shift happens: trailing edge contraction associated with recent warming trends threatens a distinct genetic lineage in the marine macroalga Fucus vesiculosus. BMC Biol 11(1):1
O’Neil ST, Dzurisin JDK, Williams CM, Lobo NF, Higgins JK, Deines JM et al (2014) Gene expression in closely related species mirrors local adaptation: consequences for responses to a warming world. Mol Ecol 23(11):2686–2698
Orizaola G, Quintela M, Laurila A (2010) Climatic adaptation in an isolated and genetically impoverished amphibian population. Ecography 33(4):730–737
Orr JC, Fabry VJ, Aumont O, Bopp L, Doney SC, Feely RA et al (2005) Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature 437(7059):681–686. https://doi.org/10.1038/nature04095
Parmesan C (2006) Ecological and evolutionary responses to recent climate change. Annu Ecol Evol Syst 37(2006):637–669. https://doi.org/10.2307/annurev.ecolsys.37.091305.30000024
Pearson GA, Lago-Leston A, Mota C (2009) Frayed at the edges: selective pressure and adaptive response to abiotic stressors are mismatched in low diversity edge populations. J Ecol 97(3):450–462. https://doi.org/10.1111/j.1365-2745.2009.01481.x
Peck LS, Webb KE, Bailey DM (2004) Extreme sensitivity of biological function to temperature in Antarctic marine species. Funct Ecol 18(5):625–630
Pecorino D, Barker MF, Dworjanyn SA, Byrne M, Lamare MD (2014) Impacts of near future sea surface pH and temperature conditions on fertilisation and embryonic development in Centrostephanus rodgersii from northern New Zealand and northern New South Wales, Australia. Mar Biol 161(1):101–110
Pédron N, Artigaud S, Infante J-LZ, Le Bayon N, Charrier G, Pichereau V, Laroche J (2017a) Proteomic responses of European flounder to temperature and hypoxia as interacting stressors: differential sensitivities of populations. Sci Total Environ 586:890–899
Pédron N, Le Du J, Charrier G, Zambonino-Infante J-L, Le Bayon N, Vasconcelos RP et al (2017b) Contrasting patterns of energy metabolism in northern vs southern peripheral European flounder populations exposed to temperature rising and hypoxia. Mar Environ Res 129:258–267
Pfeifer M, Passalacqua NG, Bartram S, Schatz B, Croce A, Carey PD et al (2010) Conservation priorities differ at opposing species borders of a European orchid. Biol Cons 143(9):2207–2220
Pinsky ML, Eikeset AM, McCauley DJ, Payne JL, Sunday JM (2019) Greater vulnerability to warming of marine versus terrestrial ectotherms. Nature 569(7754):108–111
Poorter H, Navas ML (2003) Plant growth and competition at elevated CO2: on winners, losers and functional groups. New Phytol 157:175–198
Post E, Langvatn R, Forchhammer MC, Stenseth NC (1999) Environmental variation shapes sexual dimorphism in red deer. Proc Natl Acad Sci 96(8):4467–4471
Qasim SZ (1956) Time and duration of the spawning season in some marine teleosts in relation to their distribution. Extrait Du Journal Du Conseil International Pour L’exploration De La Mer 21(2):141–155
Raven JA, Beardall J (2003) Carbon acquisition mechanisms of algae: carbon dioxide diffusion and carbon dioxide concentrating mechanisms. In: Tollefsbol TO (ed) Photosynthesis in algae. Springer, pp 225–244. https://doi.org/10.1007/978-94-007-1038-2_11
Raven JA, Giordano M, Beardall J, Maberly SC (2012) Algal evolution in relation to atmospheric CO2: carboxylases, carbon-concentrating mechanisms and carbon oxidation cycles. Philos Trans R Soc B Biol Sci 367(1588):493–507. https://doi.org/10.1098/rstb.2011.0212
Reed HR (2004) Relationship between population size and fitness. Conserv Biol 19:563–568
Rehfeldt GE, Ying CC, Spittlehouse DL, Hamilton DA Jr (1999) Genetic responses to climate in Pinus contorta: niche breadth, climate change, and reforestation. Ecol Monogr 69(3):375–407
Ries JB, Cohen AL, McCorkle DC (2009) Marine calcifiers exhibit mixed responses to CO2-induced ocean acidification. Geology 37(12):1131–1134
Robinson SA, Wasley J, Tobin AK (2003) Living on the edge–plants and global change in continental and maritime Antarctica. Glob Change Biol 9(12):1681–1717
Rothäusler E, Rugiu L, Jormalainen V (2018) Forecast climate change conditions sustain growth and physiology but hamper reproduction in range-margin populations of a foundation rockweed species. Mar Environ Res 141:205–213
Rugiu L, Manninen I, Rothäusler E, Jormalainen V (2018) Tolerance and potential for adaptation of a Baltic Sea rockweed under predicted climate change conditions. Mar Environ Res 134:76–84
Saada G, Nicastro KR, Jacinto R, McQuaid CD, Serrão EA, Pearson GA, Zardi GI (2016) Taking the heat: distinct vulnerability to thermal stress of central and threatened peripheral lineages of a marine macroalga. Divers Distrib 22(10):1060–1068
Sagarin RD, Gaines SD (2002) The ‘abundant centre ‘distribution: to what extent is it a biogeographical rule? Ecol Lett 5(1):137–147
Sagarin RD, Gaines SD (2006) Recent studies improve understanding of population dynamics across species ranges. Oikos 115(2):386–388
Sagarin RD, Gaines SD, Gaylord B (2006) Moving beyond assumptions to understand abundance distributions across the ranges of species. Trends Ecol Evol 21(9):524–530
Sanford E, Holzman SB, Haney RA, Rand DM, Bertness MD (2006) Larval tolerance, gene flow, and the northern geographic range limit of fiddler crabs. Ecology 87(11):2882–2894
Sculthorpe CD (1967) Biology of aquatic vascular plants. Edward Arnold, London
Sexton JP, McIntyre PJ, Angert AL, Rice KJ (2009) Evolution and ecology of species range limits. Annu Rev Ecol Evol Syst. https://doi.org/10.1146/annurev.ecolsys.110308.120317
Smith RC, Ainley D, Baker K, Domack E, Emslie S, Fraser B et al (1999) Marine ecosystem sensitivity to climate change: historical observations and paleoecological records reveal ecological transitions in the Antarctic Peninsula region. Bioscience 49(5):393–404
Somero GN (2010) The physiology of climate change: how potentials for acclimatization and genetic adaptation will determine ‘winners’ and ‘losers.’ J Exp Biol 213(6):912–920
Soulé M (1973) A theory of marginal populations. Annu Rev Ecol Syst 4:165–187
Stevens GC (1992) The elevational gradient in altitudinal range: an extension of Rapoport’s latitudinal rule to altitude. Am Nat 140(6):893–911
Stirling I, Lunn NJ, Iacozza J (1999) Long-term trends in the population ecology of polar bears in western Hudson Bay in relation to climatic change. Arctic 52:294–306
Therry L, Lefevre E, Bonte D, Stoks R (2014a) Increased activity and growth rate in the non-dispersive aquatic larval stage of a damselfly at an expanding range edge. Freshw Biol 59(6):1266–1277
Therry L, Zawal A, Bonte D, Stoks R (2014b) What factors shape female phenotypes of a poleward-moving damselfly at the edge of its range? Biol J Lin Soc 112(3):556–568
Thomas CD, Bodsworth EJ, Wilson RJ, Simmons AD, Davies ZG, Musche M, Conradt L (2001) Ecological and evolutionary processes at expanding range margins. Nature 411(6837):577
Thomas CD, Cameron A, Green RE, Bakkenes M, Beaumont LJ, Collingham YC et al (2004) Extinction risk from climate change. Nature 427(6970):145–148. https://doi.org/10.1038/nature02121
Thornton DH, Wirsing AJ, Lopez-Gonzalez C, Squires JR, Fisher S, Larsen KW et al (2018) Asymmetric cross-border protection of peripheral transboundary species. Conserv Lett 11(3):e12430
Tomanek L (2010) Variation in the heat shock response and its implication for predicting the effect of global climate change on species’ biogeographical distribution ranges and metabolic costs. J Exp Biol 213(6):971–979
Valiente AG, Beall E, Garcia-Vazquez E (2010) Population genetics of south European Atlantic salmon under global change. Glob Change Biol 16(1):36–47
Viejo RM, Martínez B, Arrontes J, Astudillo C, Hernández L (2011) Reproductive patterns in central and marginal populations of a large brown seaweed: drastic changes at the southern range limit. Ecography 34(1):75–84. https://doi.org/10.1111/j.1600-0587.2010.06365.x
Visser ME (2008) Keeping up with a warming world; assessing the rate of adaptation to climate change. Proc R Soc B Biol Sci 275(1635):649–659
Volis S, Mendlinger S, Olsvig-Whittaker L, Safriel UN, Orlovsky N (1998) Phenotypic variation and stress resistance in central and peripheral populations of Hordeum spontaneum. Biodivers Conserv 7(6):799–813
Vucetich JA, Waite TA (2003) Spatial patterns of demography and genetic processes across the species’ range: null hypotheses for landscape conservation genetics. Conserv Genet 4(5):639–645
Wernberg T, White M, Vanderklift MA (2008) Population structure of turbinid gastropods on wave-exposed subtidal reefs: effects of density, body size and algae on grazing behaviour. Mar Ecol Prog Ser 362:169–179
Wernberg T, Russell BD, Moore PJ, Ling SD, Smale DA, Campbell A et al (2011) Impacts of climate change in a global hotspot for temperate marine biodiversity and ocean warming. J Exp Mar Biol Ecol. https://doi.org/10.1016/j.jembe.2011.02.021
Wethey DS, Woodin SA, Hilbish TJ, Jones SJ, Lima FP, Brannock PM (2011) Response of intertidal populations to climate: effects of extreme events versus long term change. J Exp Mar Biol Ecol 400(1–2):132–144
Whittaker RH (1956) Vegetation of the Great Smoky Mountains. Ecol Monogr 26:1–80
Williamson CJ, Brodie J, Goss B, Yallop M, Lee S, Perkins R (2014) Corallina and Ellisolandia (Corallinales, Rhodophyta) photophysiology over daylight tidal emersion: Interactions with irradiance, temperature and carbonate chemistry. Mar Biol. https://doi.org/10.1007/s00227-014-2485-8
Woodward FI (1987) Climate and plant distribution. Cambridge University Press
Xiong FS, Day TA (2001) Effect of solar ultraviolet-B radiation during springtime ozone depletion on photosynthesis and biomass production of Antarctic vascular plants. Plant Physiol 125(2):738–751
Xiong FS, Mueller EC, Day TA (2000) Photosynthetic and respiratory acclimation and growth response of Antarctic vascular plants to contrasting temperature regimes. Am J Bot 87(5):700–710
Xiong FS, Ruhland CT, Day TA (2002) Effect of springtime solar ultraviolet-B radiation on growth of Colobanthus quitensis at Palmer Station, Antarctica. Global Change Biol 8(11):1146–1155
Yang A, Dick CW, Yao X, Huang H (2016) Impacts of biogeographic history and marginal population genetics on species range limits: a case study of Liriodendron chinense. Sci Rep 6:25632. https://doi.org/10.1038/srep25632
Zardi GI, Nicastro KR, Serrão EA, Jacinto R, Monteiro CA, Pearson GA (2015) Closer to the rear edge: ecology and genetic diversity down the central-edge gradient of a marine macroalga. Ecosphere 6(2):1–25
Acknowledgements
I thank Dr Federica Ragazzola from the Department of Integrative Marine Ecology, Ischia Marine Centre, Stazione Zoologica Anton Dohrn, Italy, for her valuable input and constructive comments and discussions of this manuscript, and her continuous support over the years.
Funding
This review was prepared with funding from the University of Portsmouth.
Author information
Authors and Affiliations
Contributions
RK: literature research, analyses, writing of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kolzenburg, R. The direct influence of climate change on marginal populations: a review. Aquat Sci 84, 24 (2022). https://doi.org/10.1007/s00027-022-00856-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00027-022-00856-5