
Abstract
The utilization of winter-killed cover crops is increasing due to their benefits in intensive cropping systems based on summer cash crops. However, there is still a lack of knowledge about their management in temperate climates, where maize is planted early under conservation tillage techniques. For these conditions, here we document for the first time various agronomic effects of winter-killed cover crops under different management options. We evaluated the production, the nitrogen uptake, and the weed control of five pure winter-killed cover crop species, as well as the production of the subsequent maize and its nitrogen recovery, in four different sites. Several management options were compared (cover crop fertilization, sowing technique, and cover crop termination method). Legume cover crops (Trifolium alexandrinum L. and Vicia benghalensis L.) had a small above-ground biomass (on average 0.6 t DM ha−1 in November), while for non-legumes (Avena strigosa Schreb., Sinapis alba L., and Raphanus sativus L.) the production (on average 2.4 t DM ha−1, N uptake 89 kg N ha−1) was higher, as well as weed control and N uptake. The difference in the above-ground biomass between the two groups of cover crops was smaller at the end of winter (0.4 Mg DM ha−1), when in five out of eight site × year combinations, soil mineral N was significantly higher in a cover crop treatment compared to the no cover crop, presumably due to N release from cover crops. Cover crops did not increase maize production, and their residues did not hamper maize sowing and emergence. The recovery of cover crop N by maize was 86% for legumes and − 1% for non-legumes. We conclude that productivity, N uptake, and weed control of winter-killed cover crops (especially non-legume species) are encouraging, with no negative effects on maize yield.
Similar content being viewed by others

1 Introduction
Winter cover crops are defined as crops grown when a winter cash crop is normally absent (Justes 2017; Thorup-Kristensen et al. 2003). They are sown after harvest of a summer cash crop like maize (between August and October) and are grown during fall and winter, until the following cash crop is sown (Justes 2017). At the end of their cycle, they are normally left on the soil surface or incorporated in it. The benefits of cover crops are mainly the reduction of nitrate leaching (Constantin et al. 2010; Thapa et al. 2018a), weed control (Osipitan et al. 2018), and the improvement of soil fertility in terms of reduction of soil erosion, increase of soil organic matter, and improvement of soil structure (Poeplau and Don 2015). A disadvantage is their pre-emptive competition for nitrogen (Thorup-Kristensen et al. 2003). In addition, their management before planting the following cash crop is critical, to allow optimal cash crop emergence (Benincasa et al. 2010). Therefore, selecting cover crop species, sowing and termination date, termination methodology, and estimating their nitrogen (N) fertilizer value for the following crop are important decisions to make when cover crops are introduced into cultivation (Clark et al. 2007).
Species selection depends on climatic conditions, soil fertility, farm N availability, and frost resistance. For this reason, multicriteria decision analysis, based on the evaluation of different variables (e.g., crop biomass, N uptake, C/N ratio), has also been applied to test the performance of different species and cultivars according to their suitability to be used as cover crops depending on the requirements of the farming system (Ramírez-García et al. 2015). The most used cover crops belong to the families of grasses, brassicas, and legumes (Clark et al. 2007). Legume species are well suited when soil organic matter is low, in farms without livestock operations and in organic cultivation systems, where N cannot be provided with synthetic fertilizers (Clark et al. 2007). Grasses and brassicas are widely used in the other cases, mainly to reduce nitrate leaching (Thapa et al. 2018a; Tonitto et al. 2006). The sensitivity of winter cover crops to frost (Clark et al. 2007) is an important character that allows distinguishing species that in temperate climates are winter-killed from those that are winter-hardy. The classification of cover crop species in winter-killed and winter-hardy is somewhat subjective, because frost damage depends on many factors (among which sowing date—see below), and may not occur every year (e.g., Kaye et al. 2019). Winter-hardy cover crops (like hairy vetch, Vicia villosa Roth., and rye, Secale cereale L.) need to be terminated at the end of winter, either chemically or mechanically (Creamer and Dabney 2002). Winter-killed species (like white mustard, Sinapis alba L. and radish, Raphanus sativus L.) are preferred in conservation agriculture over winter-hardy species because their biomass (already partly decomposed at the end of winter) can be easily managed even with minimum tillage operations. For other species, like berseem clover (Trifolium alexandrinum L.), information about frost resistance is more limited, though there is evidence that it may winter kill (Clark et al. 2007; Ghaffarzadeh 1997; Toom et al. 2019a). Winter-hardy species can be terminated mechanically, but this can be successfully done only at an advanced development stage (e.g., after anthesis for grasses, Ashford and Reeves 2003), thus requiring to postpone the sowing of the summer cash crop. If termination needs to be done earlier, it shall be carried out chemically. In temperate climates, winter-killed species are also well suited when maize is sown early at the end of winter (from mid-March to mid-April), because the growth that a winter-hardy species can achieve between January and March is very limited (Hashemi et al. 2013). Instead, winter-hardy species can be used when the cash crop is sown later, as for tomato and soybean (end of April–early May, Singh et al. 2018). Despite the advantages of winter-killed species, winter-hardy species like hairy vetch and cereal rye are the most studied in temperate climates (Thapa et al. 2018b), while the literature about winter-killed species is much rarer. Indeed, under temperate climates, evidence about the behavior as a cover crop in maize rotations for black oat (Avena strigosa Schreb.) (Borkert et al. 2003), purple vetch (Vicia benghalensis L.) (Couëdel et al. 2018), berseem clover (Parr et al. 2011), and white mustard (Rüegg et al. 1998) is extremely scarce (Fig. 1).

Example of two cover crop species in autumn (November, on the left) and at the end of winter (March, on the right). White mustard (Sinapis alba L.) at the top and tillage radish (Raphanus sativus L.) at the bottom. Both of them, as a winter-killed species, are largely damaged by frost at the end of winter (all photographs by the authors).
Regarding cover crop sowing, this can be performed after seedbed preparation (minimum tillage), or in no-tillage conditions. Both techniques are interesting due to the cost reduction compared to conventional seedbed preparation techniques. However, experiments comparing the performance of cover crops under different tillage techniques are not frequent, as normally the study of tillage effects is focused on the cash crop (e.g., Crowley et al. 2018). Concerning sowing date, in temperate environments with sufficient water availability, as in the Po Valley (northern Italy), early cover crop sowing (end of August or beginning of September) allows taking advantage of autumn temperature and radiation, which are still high in this period. Consequently, the high biomass accumulation and the more advanced crop development stage make cover crops more frost-sensitive. However, excessive precipitation (in particular on fine soils or with a bad structure), or vice versa dry soil conditions, might require postponing sowing. In this case, late sowing would have two main negative effects: it would limit cover crop autumn growth (Thomsen and Hansen, 2014; Toom et al. 2019b) and it would make winter-killed cover crops less sensitive to frost damage. Cover crops in early vegetative stages are less sensitive to frost (Fowler et al. 2014), allowing spring growth that, as mentioned above, is not desirable in conservation farming with minimum tillage if the cash crop is sown at the end of winter.
The cover crop N made available for the subsequent cash crop depends on many factors, among which the total amount of N in cover crop biomass and its mineralization rate are the most important. The cover crop N fertilizing replacement value in general is low for non-legume species, reaching as net effect (difference between net N mineralization and pre-emptive competition of cover crops) between 0 and 25% of the cover crop N uptake (corresponding to a few kg N ha−1 or even to a null effect) (Thorup-Kristensen et al. 2003). Vice versa, the net effect for legume cover crops is higher, reaching 100 kg N ha−1 or even more (Thorup-Kristensen et al. 2003). However, these values are extremely variable since they depend on various factors, such as cover crop biomass, soil conditions, climate, and cash crop species and management. Moreover, such data in temperate climates for species that are winter-killed, like black oat, purple vetch, and white mustard, or potentially winter-killed, like berseem clover, are rather rare (e.g., Petroudi et al. 2011; Rüegg et al. 1998; Vicensi et al. 2020). We hypothesize that the mineralization rate of winter-killed cover crop N will be rather low after termination, due to low temperatures, and will increase in spring. Therefore, the N made available for the subsequent cash crop shall not differ from the order of magnitude already known in general and reported above, with small losses during the winter period.
In conclusion, there is a lack of information regarding cover crop agronomic management, especially in conservation agriculture, for intensive systems in temperate climates characterized by early maize sowing, where winter-killed species are particularly useful. Such an environment is found in the Po Valley, where intensive farming systems are characterized by irrigated maize as the predominant crop, fertilized with synthetic fertilizers and animal manures. Due to the high N load, the winter bare fallow period may lead to N leaching (Perego et al. 2012), and to soil structure deterioration. Even if winter cover crops are conceived to catch N remaining from the previous summer crop, farmers may need to apply part of the stored manure in the fall before cover crop sowing. Therefore, it is important to verify whether slurry can be valorized or not when applied before cover crops.
We made therefore these hypotheses: (1) by planting cover crops in September, good levels of biomass and N uptake can be obtained before the frost damage occurs, normally between December and January; (2) winter-killed species have similar N effects on cash crop as winter-hardy species; (3) the N effects from legume cover crops are lower compared to non-legume cover crops. Therefore, the objective of this work is to evaluate in four sites of the Po Valley: (1) production, N uptake, and weed control capacity of five monocultures of winter-killed winter cover crop species, grown during the autumn–winter period in substitution of fallow, and followed by a maize cash crop; (2) effects of management techniques (sowing method, slurry application before planting) on cover crop growth, N uptake, and weed control; (3) production of maize cultivated after cover crops and the recovery of cover crop N by maize.
2 Materials and methods
2.1 Experimental sites and design
The 2-year field experiments were located in four sites situated in a flat area in Lombardia (Northern Italy) comprised between 45°10′ N and 45°24′ N, and between 9°15′ E and 10°06′ E. The four locations were Landriano (PV), Orzinuovi (BS), Offlaga (BS), and Sesto Cremonese (CR), hereafter referred to as D-CT, D-MT, C-MT1, and C-MT2 respectively. The soils of the four sites are described in Table 1. Sites D-CT and D-MT were in dairy farms (D) adopting conventional (CT) and conservation agriculture with minimum tillage (MT), respectively, while C-MT1 and C-MT2 were cereal farms (C) using conservation agriculture.
Depending on the number of factors involved in each site, the experimental design with two replicates was a split-split plot in sites D-CT, D-MT, and C-MT1 (three factors), and a split plot with two factors and two replicates in C-MT2. In the four sites, plot area was 100 m2.
The factors studied were cover crop species, cover crop slurry fertilization, cover crop sowing technique, maize N fertilization, and cover crop termination/weed control method in maize. The type of factor varied among sites according to the soil characteristics and the agronomic management. The factors studied in each site and their hierarchy in the experimental design are given in supplemental material (Supplemental Material, Table S1).
Each of the factors reported above has a different number of levels. The cover crops were winter-killed species: a grass (black oat), two brassicas (white mustard and tillage radish, Raphanus sativus L. var. oleiformis), and two legumes (berseem clover and purple vetch). Non-legume species were cultivated in the dairy farms and in C-MT2, while legume species were cultivated only in the cereal farms. In each site, a level of the experimental factor “cover crop species” was a control treatment without cover crops during the autumn and winter, where weeds were not controlled. The cover crop management techniques differed according to farm type. We decided to test fertilization with cattle slurry (yes vs no) in the two dairy farms together with the sowing technique (minimum tillage vs direct seeding). In the two cereal farms, maize N fertilization (yes vs no) was a factor to evaluate the N fertilizer value of cover crops. In one of the cereal farms (C-MT1), we compared two techniques for cover crop termination and weed management: mechanical vs chemical.
2.2 Crop management
The experiment started in September 2017 and ended in August 2019. In every site, the rotation was winter cover crops–maize–winter cover crops–maize. Cover crop seed bed preparation was generally carried out with minimum tillage techniques (see Table S2, Supplemental Material); in D-CT and D-MT, a comparison was made with direct seeding. In two sites (D-CT, D-MT) cover crops received either no slurry or slurry application (40 m3 ha−1) a few days before planting. Cover crops were sown during September, on average 15 days after maize harvest, with a row distance of about 20 cm and a depth of 1.5 cm. The seed rate for cover crops was: 8 kg ha−1 (tillage radish), 15 kg ha−1 (white mustard), 50 kg ha−1 (black oat), 25 kg ha−1 (berseem clover), 50 kg ha−1 (purple vetch).
In 2018, in C-MT2 2.9 t ha−1 (MgCO3) was added to raise the pH value to neutrality.
All the cover crops were always terminated before their flowering stage according to the farmer needs. In spring, depending on the treatment, the cover crops were terminated with 2 L ha−1 of glyphosate (Roundup Platinum, glyphosate acid 480 g/L) or with a disk harrow.
Pre-planting maize fertilization was carried out with mineral fertilizers only in C-MT1 and C-MT2, while dairy slurry was used in D-CT and D-MT. Slurry applied pre-planting had an average total N concentration in the 2 years of 2.75 and 2.8 g N kg−1 fresh matter for D-CT and D-MT respectively, and NH4-N concentration of 1.2 and 1.1 N-NH4 g kg−1 fresh matter for D-CT and D-MT respectively. The N rate applied with slurry was therefore 110 kg N ha−1. The mineral N rate (see Supplemental Material, Table S2), on average 65 kg N ha−1 before planting and ranging from 40 to 160 kg N ha−1 at side dressing, was calculated based on the previous 5-year maize production and according to the Lombardy integrated nutrient management guidelines (Sistemi di produzione integrata nelle filiere agroalimentari, 2021). Maize was sown between the end of March and the end of April, with a row distance of 70 cm and an average planting density of 7.2 plants m−2, generally after minimum tillage seed bed preparation. Maize was always irrigated, generally with surface method. In all the sites, maize was harvested at R5 stage for silage utilization. Cover crop and maize sowing and harvest dates, and their management operations are described in detail in the supplemental material (Tables S2 and S3).
2.3 Meteorological conditions
During cover crop growth, the average temperatures were lower in 2017 compared to 2018, except for the site C-MT1 that showed the opposite situation. For sites D-CT, D-MT, C-MT1, and C-MT2, respectively, the accumulated thermal units (from cover crop sowing to the late autumn sampling) were 857, 730, 973, and 755 °C d in the first year, and 1107, 1303, 839, and 1249 °C d in the second year.
Both years, precipitation was scarce during the cover crop growing season, from September to November (180 mm in D-CT, 195 mm D-MT, 164 mm in C-MT1, and 167 mm in C-MT2). In 2017, some rain fell from late August to half of September, and at the beginning of November, but not during October. In 2018, after a dry September and most of October, intense rainfall occurred from the end of October until November. A daily water balance was calculated for the cover crop growing season in both years, separately for each site. We used the FAO methodology (Allen et al. 1998) representing the soil with a unique layer (0–30 cm), whose hydrological constants (field capacity and wilting point) were estimated based on soil texture, using Saxton’s pedotransfer function (Saxton et al. 1986). Table 2 reports, for the cover crop growing period, the sum of precipitation and the aggregated results of the water balance, including a crop water stress index, calculated based on the daily soil water content according to Allen et al. (1998), assuming the readily available water equal to 0.5. Despite the occurrence of water stress in both seasons, no irrigation was provided to cover crops due to lack of irrigation water availability. The monthly mean temperature and total precipitation of the 2 years are reported in the Supplemental material (Fig. S1).
2.4 Sample collection
Above ground biomass (AGB) of cover crops and weeds was sampled separately from the same area of 1 m2 plot−1; for tillage radish also the root biomass was collected. Cover crop samples were collected during late autumn (between November 14th and November 23rd) and at the end of winter (between March 1st and April 8th). Because of the difficulty in separating cover crop biomass from weeds, a mixed sample of AGB (i.e., made of cover crop + weeds) was collected at the end of winter. Tillage radish was not sampled at the end of winter because its biomass was mostly already decomposed. At the end of winter frost damage was evaluated visually.
Above ground biomass of maize was collected by sampling 15 plants (whole biomass) at the maize vegetative stage 6-leaf (V6) and 20 plants (whole biomass) at the maize reproductive growth stage R5 in three and four rows from each plot, respectively (5 plants row−1). Soil samples were collected from the 0–30-cm layer (mixing three samples per plot) in autumn, at the end of winter, and at maize V6 stage. At V3 maize stage, photos were taken to quantify cover crop regrowth and the presence of weeds in maize inter-rows. Images were collected with a Canon EOS 400D camera equipped with a Canon zoom lens EF-S 18–55 mm. We sampled 0.06 m2 plot−1 replicate−1 (three replicates). The camera exposure was standardized with a gray cardboard. All photos collected at maize V3 stage were analyzed with Canopeo App for Matlab V 2.0 (Patrignani and Ochsner, 2015) to assess the percentage of the green canopy (fractional green canopy cover).
2.5 Chemical analyses
Biomass samples were oven-dried at 105 °C until constant weight to determine the dry weight and the percentage of dry matter (% DM). When fresh samples were heavier than 1.5 kg, they were chopped, and a representative subsample was taken to determine % DM. Then, after the sample biomass grinding, a representative reduced dried sample of about 40 g was ground with a centrifugal mill, fitted out with a 0.2-mm mesh sieve (ZM 100, Retsch Gmbh & Co., Haan, Germany). The concentration of total nitrogen and carbon was determined by dry combustion using a ThermoQuest NA1500 elemental analyzer (Carlo Erba, Milano, Italy).
Nitrogen uptake was calculated by multiplying the AGB by its N concentration. For maize, the cover crop apparent N recovery (%) was calculated as 100 × (maize N uptake after cover crop − maize N uptake after no-cover crop) / cover crop N uptake.
Soil samples were analyzed according to Italian official methods for physical and chemical analyses of soil (MiPAAF, 1997). Soluble and exchangeable NH4+ and NO3– were extracted from 30 g of soil sieved at 5 mm, using 100 mL of a solution of 1 M KCl (extraction ratio 1:3). Suspensions were agitated by a rotating shaker for 1 h, then filtered through Whatman #2 filter paper (Whatman International Ltd, Maidstone, England), and stored at − 20 °C until analysis. Ammonium and nitrate concentrations, which reflect the soil mineral nitrogen (SMN) concentration in soil extracts, were determined by flow injection analysis and spectrometric detection according to ISO 11732 (1997) and ISO 13395 (1996) procedures, respectively.
2.6 Statistical analysis
Statistical analyses were performed with IBM® SPSS® Statistics 25. The assumptions of homogeneity of variances and residuals normality distribution were tested with the Levene test and Shapiro–Wilk test, respectively.
Analysis of variance (ANOVA) was initially performed separately for each site to test the effect of main factors and their interactions on the dependent variables (cover crop and maize biomass, weed biomass, SMN, and cover crop and maize N uptake) considering the split plot or the split-split plot design according to the site under analysis. Then, the Ryan-Einot-Gabriel-Welsh-Q or the Sidak post hoc tests (P < 0.05) were performed to separate means, in the case of significant main effect or interaction, respectively. The list of experimental factors (whole plot, subplot, sub-subplot factors) is given for each site in the Supplemental Material (Table S1). The results of the ANOVA by site (not shown in figures or tables) indicated that cover crop species and year were the only significant sources of variation for most of sites and dependent variables. Therefore, the average of each variable obtained for all management factors at each site is presented in the results.
For this reason, we carried out a second ANOVA across sites and years, using site, year, and cover crop species as factors. This analysis was carried out with a mixed model (P < 0.05), in which cover crop species was nested within the site or vice versa: variables that can be interpreted without reference to the no-cover crop control treatment (such as cover crop AGB) were analyzed by nesting sites within cover crop species, separately by year; on the contrary, variables that need to be presented with reference to the no-cover crop treatment (such as soil mineral nitrogen) were analyzed by nesting cover crop species within sites. These are the results shown in figures. Because the assumption of variance homogeneity was met, error bars are shown only once for each variable, not for individual treatments. The present mixed model allowed also to analyze the effects of the main factors (i.e., those in the main plot), thus obtaining the expected means from the average of nested factors.
3 Results and discussion
3.1 Effects of cover crop species on biomass/nitrogen accumulation and weed control capacity
3.1.1 Cover crop biomass
The results reported in Fig. 2 (cover crop biomass in November) indicate that cover crop sowing in September gave good results in terms of biomass accumulation in autumn, especially for non-legume cover crops, due to favorable weather and soil conditions. Therefore, early silage maize harvest provides a favorable cover crop growing period with a positive effect on production.

Cover crop (green) and weeds (red) above-ground biomass in the late autumn 2017 and 2018, split by cover crop species and site. D-CT conventional farm with dairy cows, D-MT minimum tillage farm with dairy cows, C-MT1 conservation cereal farm, C-MT2 conservation cereal farm; Nc no cover, As Avena strigosa, Sa Sinapis alba, Ta Trifolium alexandrinum, Tr Tillage radish, Vb Vicia benghalensis. Letters compare sites inside species; lowercase red letters compare weed biomass, and uppercase green letters compare cover crop biomass. Columns with different letters are significantly different (P < 0.05). Standard error bars are reported for each variable and year.
In general, non-legume cover crops (white mustard, black oat, and tillage radish) had considerable AGB production in autumn, with average values across all sites, years, and treatments of 2.7, 2.0, and 2.6 t DM ha−1, respectively for the three species. White mustard was the only species with biomass consistently higher than 2 t DM ha−1 across years and sites. Its biomass was not different between sites in the first year, while in the second year it was higher in D-MT than in C-MT2. White mustard production was comparable with the data range reported by the review of Haramoto and Gallandt (2004).
In the first year, black oat biomass was significantly higher in D-CT than in D-MT and C-MT2. Black oat and tillage radish AGB were similar to that normally found in the literature, namely: 1.8–3.1 t DM ha−1 in the case of black oat (Dial, 2014); 2.5–3.5 t DM ha−1 in the case of tillage radish (Dapaah and Vyn, 1998). Worth of mention is also the important root biomass of tillage radish (0.7 t DM ha−1 on average for the 2 years). The root biomass production is in agreement with that reported by Ruffatti et al. (2019) that ranged from 0.1 to 0.8 t DM ha−1. The duration of vegetative growth (computed counting days between cover crop sowing and the last sampling dates) and the Growing Degree Days (GDD, with base temperature of 0 °C) accumulation contribute to explain the significant different cover crop production between sites within the same year (Fig. 2). In 2017, the greater production of black oat in D-CT can be explained by the longer vegetative growth (+ 16 days) compared to other sites. As a consequence, in D-CT compared to D-MT and C-MT2 the equivalent accumulation of GDD (with a base temperature of 0 °C) was 857 °C d against 730 and 755 °C d in D-MT and C-MT2, respectively. The same situation occurred for white mustard in the second year when, in D-MT, it accumulated about 100 °C d more than in C-MT2.
In our study, legume cover crops (purple vetch and berseem clover) had the lowest and the less constant production during autumn compared to non-legume species (Fig. 2).
Berseem clover biomass was in general lower than 0.5 t DM ha−1, except for C-MT2 in the second year. A similar result was found for purple vetch, whose biomass was between 0.5 and 1.0 t DM ha−1, except for C-MT2 in the second year. Optimal clover biomass accumulation was found by Grüter et al. (2017) in similar climatic conditions, reaching more than 2 t DM ha−1. In the research conducted by Toom et al. (2019b), instead, berseem clover biomass never exceeded 1.5 t DM ha−1; their best result was obtained with an early sowing in the first week of August, while with later sowing biomass accumulation decreased until 0.3 t DM ha−1, not very different from our average of 0.46 t DM ha−1. The lack of purple vetch data from the literature makes it difficult any comparison with our results (average of 0.76 t DM ha−1), or with other vetch species that produce considerably more due to their frost resistance (Spargo et al. 2016). Reasons for the low performance of legume cover crops in our experiments might be a too late sowing date and weather for both sites (C-MT1 and C-MT2), poor soil structure in C-MT1, and low soil pH in C-MT2 (resolved in the second year).
In principle, between the end of autumn and the end of winter, cover crops can continue to grow depending on their winter hardening ability (Duiker, 2014), while weed biomass accumulation depends on the interactions between weather and seed bank.
For example, Richards et al. (1996) reported a cover crop uptake ranging from 0 to 38 kg N ha−1 during winter, while Baggs et al. (2000) obtained only 1–5 kg N ha−1. However, in our case, end-of-winter biomass (cover crop + weeds) for white mustard and black oat was lower compared to the autumn. As a general pattern, the biomass at the end of winter was higher for non-legume species compared to legume species, except for white mustard in the first year (Fig. 3). In fact, white mustard, at this time represented by standing stems, and tillage radish (degraded and lying on the soil surface) were always completely killed by frost, while black oat had a different behavior among sites and years, sometimes remaining alive despite the low temperatures, sometimes making a uniform dead mulch layer covering the soil. Black oat in the first year at C-MT2, probably due to the late sowing date compared to other sites and therefore to greater frost resistance, had greater biomass at the end of winter compared to the autumn. Comparing black oat biomass at the end of winter between sites, we found a significantly higher biomass accumulation in C-MT2 in the first year, while in the second year there were no differences among sites.

Cover crop (green) and weeds (red) above-ground biomass at the end of winter 2018 and 2019, split by cover crop species and site. Above-ground biomass refers to weeds in the control treatment (Nc), and to a mixture of cover crops and weeds in the other treatments. D-CT conventional farm with dairy cows, D-MT minimum tillage farm with dairy cows, C-MT1 conservation cereal farm, C-MT2 conservation cereal farm; Nc No cover, As Avena strigosa, Sa Sinapis alba, Ta Trifolium alexandrinum, Tr Tillage radish, Vb Vicia benghalensis. Letters compare sites inside species. Lowercase red letters compare weed biomass, and uppercase green letters compare cover crop biomass. Columns with different letters are significantly different (P < 0.05). Standard error bars are reported for each variable and year.
Differently to what was observed for non-legume species, in the first year, the biomass of mixes of legume cover crops and weeds was higher at the end of winter compared to November. In the second year, this was true only for C-MT1. However, this was not the result of cover crop growth, but rather of weed growth during winter, as demonstrated by high ratios of weed biomass/mix biomass in March (on average, 41 and 60% for berseem clover and purple vetch, respectively, measured in two blocks of C-MT1 and C-MT2 in March 2019). In addition, depending on the site and year, legume cover crops remained partially alive (as in the case of berseem clover in the second year in C-MT2), or were killed by frost (purple vetch in the first year in C-MT2). This indicates that legume cover crops were not capable of substantial weed control at the end of winter, as already demonstrated by Kaye et al. (2019) who highlighted the poor competition of clover against weeds in the cover crop season. Nevertheless, comparing legume biomass accumulation at the end of winter among sites, purple vetch and berseem clover produced significantly more in C-MT1 than in C-MT2 in the first year.
3.1.2 Cover crop nitrogen uptake
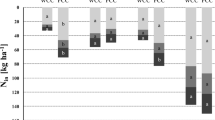
Figure 4 reports the amounts of mineral N in the soil and total N in the crop at the autumn sampling. The total height of the bars provides a measure of the total amount of N conserved in the system at the end of the autumn growing season of the cover crops.

Soil mineral nitrogen (darker blue) and above-ground nitrogen uptake (light blue) in the late autumn 2017 and 2018, split by site and cover crop species. Nitrogen uptake refers to weeds in the control treatment (Nc), and to cover crops in the other treatments. D-CT conventional farm with dairy cows, D-MT minimum tillage farm with dairy cows, C-MT1 conservation cereal farm, C-MT2 conservation cereal farm; Nc no cover, As Avena strigosa, Sa Sinapis alba, Ta Trifolium alexandrinum, Tr Tillage radish, Vb Vicia benghalensis. Letters compare cover crop species inside sites; lowercase light blue letters compare nitrogen uptake, and uppercase darker blue letters compare soil mineral nitrogen. Columns with different letters are significantly different (P < 0.05). Standard error bars are shown for cover crop and weed separately.
Despite the fact that they cannot fix atmospheric N, non-legume cover crops accumulated high quantities of N in their AGB (Fig. 4). Compared to legume species, they had a significantly (P < 0.001) higher concentration in 2017 (5.1%) and a significantly (P < 0.001) lower concentration in 2018 (2.5%). In the first year, black oat and white mustard reached uptake values of 120–130 kg N ha−1 (with N concentrations in the above ground biomass of 4.0–3.9% for the two species respectively, on average for the sites and the 2 years), while tillage radish reached 160 kg N ha−1 including the taproot. Non-legume cover crop N uptake in November was on average 62 kg N ha−1, a higher value than what reported by Tonitto et al. (2006) in their meta-analysis (37 kg N ha−1). This might be due to favorable N uptake conditions of the first year.
In fact, two distinct weather patterns were observed during the cover crop season in the first (2017) and in the second (2018) year, from early September to mid-November (Table 2). In 2017, October was very dry, with very low precipitation (2 mm on average), while in 2018 a dry period was experienced in September and the first half of October. Based also on the simplified water budget (Table 2), we suppose that in 2017 cover crops had a greater possibility to absorb N due to reduced drainage between the end of September and the beginning of November, while in 2018 intense leaching below the shallow rooting depth starting at the end of October might have reduced the N uptake. In addition, until the second decade of October, the growing season 2017 was slightly less affected by crop water stress compared to 2018. Both factors (reduced drainage and lower crop water stress) might have favored cover crop N uptake in 2017 compared to 2018. Weather patterns are recognized as a factor influencing cover crop growth, as already demonstrated by Benincasa et al. (2010), who stated that cover crop biomass and N accumulation varied for the same species between the experimental years. Moreover, cover crops were always more effective in catching and storing N than weeds in the control treatment, as demonstrated by their higher N uptake (Fig. 4). With high N availability conditions, as in 2017, non-legume cover crops AGB stored on average 74 kg N ha−1 more than weeds. This N uptake difference, even if smaller (18 kg N ha−1), persisted on average for all the sites also in 2018, despite the likely higher N leaching. This important role of cover crops was reviewed in several meta-analytic studies, who quantified the reduction of N leaching by non-legume species compared to bare soil as 56–70% (Thapa et al. 2018a; Tonitto et al. 2006; Valkama et al. 2015).
Nitrogen concentration of legume cover crops in November was rather high (3.7 and 4.5% for clover and vetch, respectively, on average for the two years and sites), consequence of its symbiosis with Rhizobium. In C-MT2 legume cover crops had a steady N concentration in the 2 years (on average for the two species: 4.1 and 3.9%, in 2017 and 2018, respectively). Due to very small biomass accumulation of legume cover crops in November (discussed above), their nitrogen uptake was very small in 2017 in both sites (25 and 14 kg N ha−1, respectively in C-MT1 and C-MT2), while in 2018 it increased in C-MT2 (60 kg N ha−1) and remained low in C-MT1 (19 kg N ha−1). Compared to the autumn, at the end of winter cover crop biomass N concentration was in general smaller (likely due to N dilution and to the presence of weeds), and differences among years and among species were reduced (data not shown). Nevertheless, in most cases, the N uptake of the cover crop treatments was significantly higher than that of the control treatment (Fig. 5).

Soil mineral nitrogen (darker blue) and above-ground nitrogen uptake (light blue) in winter 2018 and 2019, split by site and cover crop species. Nitrogen uptake refers to weeds in the control treatment (Nc), and to a mixture to cover crops and weeds in the other treatments. D-CT conventional farm with dairy cows, D-MT minimum tillage farm with dairy cows, C-MT1 conservation cereal farm, C-MT2 conservation cereal farm; Nc no cover, As Avena strigosa, Sa Sinapis alba, Ta Trifolium alexandrinum, Tr Tillage radish, Vb Vicia benghalensis. Letters compare cover crop species inside sites; lowercase light blue letters compare nitrogen uptake, and uppercase dark blue letters compare soil mineral nitrogen. Columns with different letters are significantly different (P < 0.05). Standard error bars are reported for each variable and year.
Legumes showed a higher N concentration than non-legumes in both years (on average 2.9 vs 2.4% in the first year, 2.8 vs 2.0% in the second year). In fact, N uptake at the end of winter (Fig. 5) was in general smaller than in autumn for non-legume cover crops, and vice versa for the legumes. The corresponding C/N ratio for each cover crop species across sites and years ranged between 13 and 23 for black oat, 14 and 37 for white mustard, 10 and 18 for berseem clover and 11 and 19 for purple vetch.
3.1.3 Soil mineral nitrogen
In autumn, soil mineral N in the 0–30-cm layer (Fig. 4) was between about 10 and 70 kg N ha−1 in most treatments in the 2 years, with higher values in dairy compared to cereal farms. The control treatment contained significantly more SMN than the cover crop treatments only in D-CT (both years). In the two dairy sites, SMN in November of the first year was not negligible (on average 50 kg N ha−1), even if we sampled cover crops at their hypothetical maximum biomass accumulation that usually occurs in November, in our climatic conditions. This may indicate that in the first year the supply of mineral N in these two sites was in excess compared to cover crop absorption capacity. As discussed above, legume and non-legume species responded differently to weather conditions in the 2 years. We attribute this effect to the higher soil N availability in the fall of 2017 (Fig. 4). In 2018, soil mineral N concentration was lower due to higher water drainage. For this reason, while non-legume species had a lower N concentration, legume cover crops were able to maintain their concentration independently from the soil N availability, due to N fixation as already demonstrated by other authors (Benincasa et al. 2010; Restovich et al. 2019).
At the end of winter, in the first year (2018), legumes showed a significantly lower amount of soil mineral N compared to non-legume species (23 vs. 42 kg ha−1). In the same way, in the second year (2019), legumes had the lowest N content in the soil compared to non-legumes and to the control treatment (25 vs. 53 and 33 kg ha−1). In the first year, SMN was significantly lower in the control treatment compared to at least one cover crop treatment in three sites (D-CT, D-MT, and C-MT1), and in two sites (D-CT and D-MT) in the second year (Fig. 5). In the second year, non-legume cover crop biomass degradation on the ground lead to a higher SMN at the end of winter (on average 59 kg N ha−1) compared to the autumn (on average 32 kg N ha−1). This effect was not found in the first year, probably because the N mineralized at the end of winter was leached due to abundant precipitation in December through March (199–265 mm depending on the location). Nitrogen release in the cover crop treatment at the end of winter is also reported by Kaye et al. (2019) who found increased SMN levels in the upper soil layer in late spring (May) when all cover crops had winter killed.
3.1.4 Weed control capacity of cover crops in autumn
In the first year (2017), weed biomass in November in the control treatment was significantly different between sites, while in the second year (2018) it was significantly higher only in D-MT (Fig. 2). Differences in weed growth in the control treatment between sites and years were likely due to differences in weed seed bank, weather conditions, soil organic matter, and seed bed preparation. Compared to the control treatment, the initial high crop growth rate of non-legumes allowed them to reduce or even eliminate the presence of weeds in November, as clearly shown in the two dairy farms (Fig. 2). Weed growth under the black oat treatment was important only in D-MT in both years; in the second year, this brought the total weed and cover crop biomass above 3 t DM ha−1. Weed biomass in mustard instead was always negligible, except in the second year in D-MT, where it reached 15% of the total AGB, still much lower than in the control treatment (2 t DM ha−1). The particular result in D-MT in the second year was likely due to a precipitation event of 78 mm (in the first two days of September) before cover crop sowing that likely promoted weed germination. Weed control in autumn by legume cover crops was difficult to evaluate due to the scarce presence of weeds in both years at C-MT1 and C-MT2 (always below 0.3 t DM ha−1 in the control treatment). The important role of cover crops for weed control, and of white mustard in particular, was confirmed by the meta-analysis of Osipitan et al. (2018) and by the review of Haramoto and Gallandt (2004). Baraibar et al. (2018) have found that legume cover crop species had the lowest ability to suppress weed growth compared to non-legumes (brassicas and grasses).
3.2 Effects of management practices on cover crop growth
Contrarily to cover crop species, the cover crop management factors tested in our experiment (i.e., slurry application before cover crop planting and cover crop seeding method) did not significantly affect the measured variables. One reason for this lack of effect is that in nested experimental designs (split-split plot and split plot) the precision for the measurement of the effects of the main-plot factor is sacrificed to improve that of the subplot factor (Gomez and Gomez 1984). This means that differences in the sub plot (or sub-sub plot) factor levels are easier to detect than those for the main factors. These designs were consistent with the purpose of our study, which was primarily to detect differences among cover crop species.
Slurry application before cover crop planting was tested only in the two sites of the dairy farms, where it did not affect biomass or N accumulation in any of the measurement dates: in D-CT, the average cover crop biomass in the 2 years was 1.76 and 2.18 t DM ha−1 with and without slurry application, respectively, and 2.01 and 1.89 t DM ha−1 in D-MT. One reason for this lack of effect could be the high potential N mineralization in the soils at the two sites (with 2.5 and 5.6% SOM; Table 1), as also demonstrated by the high SMN content in the first year (Fig. 4). Nitrogen was likely not limiting cover crop growth, and therefore no effects from the slurry application could be detected (as shown in the first experimental year by Cavalli et al. 2016 with a dairy slurry). Therefore, our data do not allow to recommend slurry application before cover crop sowing in such contexts. Its benefits could be greater in soils with lower SOM; further research is needed to evaluate this possibility.
The cover crop seeding method did not significantly affect cover crop biomass in autumn, in the two sites in which it was evaluated. Thus, under no-tillage cover crop biomass was not significantly lower compared to minimum tillage. The effectiveness of cover crop establishment under no-tillage was confirmed also by Fiorini et al. (2020) with rye. This finding is important to support techniques that minimize the costs associated to cover crop cultivation.
3.3 Effect of cover crop species on maize production and nitrogen recovery
At maize stage V3, a measurement of biomass grown in between maize rows was conducted (via the green canopy cover assessment; Fig. S2, Supplemental material), with the aim to detect possible cover crop regrowth and weed growth in the first maize phenological stages. The small presence of vegetation in between maize rows was entirely due to weeds, since there was no cover crop regrowth for the species tested (i.e., only weed biomass has been found in the assessment). Since no living cover crop biomass was detected, we conclude that the termination practice and frost control were effective at cover crop termination. Moreover, at C-MT1, no significant differences emerged between chemical and mechanical termination, so both techniques were equally effective in biomass destruction at the end of the cover crop vegetative cycle. The large differences of green canopy cover among years and sites were likely due to the interaction between the weed seed bank and the weather. For example, the higher precipitation (+ 180 mm, between March and May) in the first year in D-CT compared to 2019 probably led to greater weed germination.
Within each site, no significant differences of fractional green canopy cover were found between maize following different cover crop treatments, except for maize after tillage radish in the first year (2018) in D-CT, where a higher green fraction was recorded compared to maize after black oat. Therefore, cover crops did not provide additional weed control at V3 compared to the treatment without cover crops, as the green fraction canopy cover measured between rows was similar in the control and in the cover crop treatments. This finding is not in agreement with the results of the meta-analysis by Osipitan et al. (2018), who indicated that cover crops, compared to a bare control, on average significantly reduced weed biomass at cover crop termination and up to 7 weeks after cash crop sowing.
At V6 stage, in the first year (2018) maize AGB never exceeded 1.2 t DM ha−1 (considering all sites), while in the second year (2019) it reached 0.7 t DM ha−1 (see Supplemental Material Fig. S3). In each site, there were no significant effects of previous cover crop species in both years on V6 maize AGB, except for maize after purple vetch in the first year in C-MT1, whose biomass was significantly higher than maize after the other treatments (berseem clover and no cover crop). Similarly, at V6 we did not find significant effects of cover crop species on maize N uptake, SMN and their sum in both years (see Supplemental Material Fig. S4), again except for maize after purple vetch in C-MT1 in the first year that had a significantly higher N uptake than maize after the no cover crop control treatment. The lack of negative effects of cover crops biomass on maize growth at V6 suggests that, in our conditions, cover crop residues did not interfere with maize sowing, germination, and emergence. In addition, cover crop residue did not impact maize plant density (data not shown). The lack of positive effects at V6 is probably due to the strong pre-emptive competition generated by the depletion of soil N during cover crop growth and—in some cases—to the incorporation of biomass with high C/N ratio that in an early phase does not release N, but rather immobilizes soil N (Benincasa et al. 2010). End-of-winter C/N ratios higher than 20 were measured in the second year in D-CT (black oat), in D-MT (white mustard) and in C-MT2 (black oat and white mustard). The positive effect in C-MT1 after purple vetch in the first year might be related to the low C/N ratio of the cover crop biomass (12.1), with an above ground dry matter of 1.8 t ha−1. Similarly, to the green canopy cover at V3, also maize AGB at V6 and at R5 was not significantly affected by cover crop termination method at C-MT1.
Maize production at harvest at the R5 stage (Fig. 6) did not differ between previous cover crop species in every site in both years, except for maize after purple vetch in the first year, that produced significantly more than the control treatment in C-MT1.

Maize above-ground biomass at harvest in 2018 and 2019 grouped by site and cover crop species. D-CT conventional farm with dairy cows, D-MT minimum tillage farm with dairy cows, C-MT1 conservation cereal farm, C-MT2 conservation cereal farm; Nc no cover, As Avena strigosa, Sa Sinapis alba, Ta Trifolium alexandrinum, Tr Tillage radish, Vb Vicia benghalensis. Letters compare cover crop species inside sites; columns with different letters are significantly different (P < 0.05). Standard error bars are reported for each year.
We have two explanations for this lack of response of maize yield to cover crops. Firstly, in the two dairy sites, N was not limiting because weather factors may have constrained maize production (in particular in 2019 with 76 mm on average from April to August), and because N availability was already very high (masking the potential cover crop effect as also reported by Storr et al. 2020) due to historical manure applications that have likely accumulated organic matter pools providing easily mineralizable N. Secondly, in the two cereal farms, even if N was probably a limiting factor, the relatively small amount of N in the cover crops at the end of winter (ranging from 10 to 52 kg N ha−1) might not have been sufficient to determine a significant maize response. For example, if a cover crop contains 50 kg N ha−1, only 20–50% of it is made available during the maize growing season, depending on mean temperature and biomass C/N (Li et al. 2020; Sullivan and Andrews, 2012). In addition, Jensen et al. (2005) found that the initial phase with rapid mineralization or immobilization lasted for a maximum of 4 weeks at 15 °C, using 76 different crop residues. Therefore, in our case, with low cover crop C/N ratios that ranged from 13 to 37 at the end of winter, only a small fraction of maize N uptake (10–25 kg N ha−1) would derive from cover crops. Moreover, the rather high maize N uptake (in most cases higher than 150 kg N ha−1) makes this amount hardly detectable as a significant difference compared to the control treatment. The lack of response of maize yield to cover crops is further demonstrated by the fact that even the maize N fertilization factor (in C-MT1 and C-MT2 sites) did not lead to a greater yield response compared to the control, pointing the attention to limiting factors other than N (e.g., water).
No significant effects of cover crop species and year were found at R5 stage for maize N uptake within each site (data not shown). The apparent recovery of N from cover crops in maize was not significantly affected by cover crop species (between − 12 and 5% for black oat, and between − 14 and 4% for white mustard), except for legume cover crops that reported values between 80 and 130%.
The response ratio (maize total aboveground biomass after cover crop/maize total aboveground biomass after no-cover crop) was on average 1.01 for non-legume and 1.17 for legume cover crops. This finding is consistent with the literature for non-legume cover crops: in their meta-analysis, Marcillo and Miguez (2017) found a response ratio not significantly different than 1 for non-legume species; similar results were found in the meta-analysis by Tonitto et al. (2006), who reported cash crop yield on average of − 3% after cover crops. However, the literature provides evidence of significant positive effects of legume cover crops on the yield of the subsequent cash crop that we did not find in our study. For instance, Marcillo and Miguez (2017) reported significantly higher maize production after cover crop when maize received less than 200 kg N ha−1 with fertilizers (+ 33% for 0–99 kg N ha−1; + 9% for 100–199 kg N ha−1). Moreover, Benincasa et al. (2010) indicated that N release from pure legume biomass, even when termination was delayed, would occur before maize harvest and therefore would not reduce but rather would increase the final N uptake and recovery of supplied N.
4 Conclusion
In a 2-year experiment in four sites with pure winter cover crop species in rotation with silage maize, we have measured cover crop growth, N uptake and weed control capacity, and maize production. The results are original for the species tested (winter-killed or potentially winter-killed) and the environmental context (temperate climate, loamy or sandy loam soils, and cover crop planting after maize silage). Our results indicate that legume cover crops (purple vetch and berseem clover) had a lower and more variable biomass production than the non-legumes (tillage radish, black oat, and white mustard). We confirmed that the cover crop species tested were generally winter-killed, even if their susceptibility to frost damage varied by site, year, and sowing date.
Cover crop biomass at the end of winter did not hamper maize seed bed preparation. Accordingly, maize sowing operations were not delayed due to cover crops; this is a very important requirement in conservation agriculture. The efficacy of cover crops in controlling weeds during autumn and winter was good or even excellent in some cases (white mustard). Another positive aspect is that cover crops did not regrow during the maize growing season. However, there were no significant benefits of cover crops on weed control during the maize cropping season. Similarly, maize yield was not positively or negatively affected by cover crops; thus, no possibilities to reduce N fertilization of the subsequent maize emerged from this study, even if they are documented in the literature for legume cover crops. In addition, we tested for the first time some management factors (cover crop sowing technique, cover crop fertilization, and cover crop termination technique), but we did not detect significant effects on the measured variables.
As a future perspective, the following issues should be evaluated in this specific cropping system (maize-based rotations under conservation agriculture in temperate climates): (1) the effects of cover crop mixtures, to valorise also the benefits of the legume species which generally performed poorly when cultivated as pure crops; (2) the effects of winter cover crops on other summer cash crops like grain maize and soybean, exploring different cover crop sowing and termination dates; (3) slurry N use efficiency when applied in the autumn before cover crops sowing in farms with low organic matter content. These studies should be carried out in long-term experiments to quantify their effects over time.
Data availability
Not applicable.
Code availability
Not applicable.
References
Allen RG, Pereira LS, Raes D, Smith M (1998) Crop evapotranspiration—guidelines for computing crop water requirements—FAO Irrigation and drainage paper 56. FAO, Rome
Ashford DL, Reeves DW (2003) Use of a mechanical roller-crimper as an alternative kill method for cover crops. Am J Alternative Agr 18:37–45. https://doi.org/10.1079/AJAA2003037
Baggs EM, Watson CA, Rees RM (2000) The fate of nitrogen from incorporated cover crop and green manure residues. Nutr Cycl Agroecosys 56:153–163. https://doi.org/10.1023/A:1009825606341
Baraibar B, Hunter MC, Schipanski ME et al (2018) Weed suppression in cover crop monocultures and mixtures. Weed Sci 66:121–133. https://doi.org/10.1017/wsc.2017.59
Benincasa P, Tosti G, Tei F, Guiducci M (2010) Actual N availability from winter catch crops used for green manuring in maize cultivation. J Sustain Agr 34:705–723. https://doi.org/10.1080/10440046.2010.507452
Borkert CM, Gaudêncio CDA, Pereira JE, Pereira LR, De Oliveira JA (2003) Mineral nutrients in the shoot biomass of soil cover crops. Pesqui Agropecu Bras 38:143–153. https://doi.org/10.1590/S0100-204X2003000100019
Cavalli D, Cabassi G, Borrelli L et al (2016) Nitrogen fertilizer replacement value of undigested liquid cattle manure and digestates. Eur J Agron 73:34–41. https://doi.org/10.1016/j.eja.2015.10.007
Clark AJ, Meisinger JJ, Decker AM, Mulford FR (2007) Effects of a grass-selective herbicide in a vetch-rye cover crop system on nitrogen management. Agron J 99:36–42. https://doi.org/10.2134/agronj2005.0361
Constantin J, Mary B, Laurent F et al (2010) Effects of catch crops, no till and reduced nitrogen fertilization on nitrogen leaching and balance in three long-term experiments. Agr Ecosyst Environ 135:268–278. https://doi.org/10.1016/j.agee.2009.10.005
Couëdel A, Alletto L, Tribouillois H, Justes É (2018) Cover crop crucifer-legume mixtures provide effective nitrate catch crop and nitrogen green manure ecosystem services. Agr Ecosyst Environ 254:50–59. https://doi.org/10.1016/j.agee.2017.11.017
Creamer NG, Dabney SM (2002) Killing cover crops mechanically: review of recent literature and assessment of new research results. Am J Alternative Agr 17:32–40. https://doi.org/10.1079/AJAA20014
Crowley KA, Van Es HM, Gómez MI, Ryan MR (2018) Trade-offs in cereal rye management strategies prior to organically managed soybean. Agron J 110:1492–1504. https://doi.org/10.2134/agronj2017.10.0605
Dapaah HK, Vyn TJ (1998) Nitrogen fertilization and cover crop effects on soil structural stability and corn performance. Commun Soil Sci Plan 29:2557–2569. https://doi.org/10.1080/00103629809370134
Dial HL, (2014) Plant guide for black oat (Avena strigosa Schreb.). USDA—Natural Resources Conservation Service, Tucson Plant Materials Center, Tucson, AZ, 85705.
Duiker SW (2014) Establishment and termination dates affect fall-established cover crops. Agron J 106:670–678. https://doi.org/10.2134/agronj2013.0246
Fiorini A, Maris SC, Abalos D et al. (2020) Combining no-till with rye (Secale cereale L.) cover crop mitigates nitrous oxide emissions without decreasing yield. Soil Till Res 196 104442 https://doi.org/10.1016/j.still.2019.104442
Fowler DB, Byrns BM, Greer KJ (2014) Overwinter low-temperature responses of cereals: analyses and simulation. Crop Sci 54:2395–2405. https://doi.org/10.2135/cropsci2014.03.0196
Ghaffarzadeh M (1997) Economic and biological benefits of intercropping berseem clover with oat in corn-soybean-oat rotations. J Prod Agric 10:314–319. https://doi.org/10.2134/jpa1997.0314
Gomez KA, Gomez, AA (1984). Statistical procedures for agricultural research, 2nd edition, 704 Pages.
Grüter R, Costerousse B, Bertoni A et al (2017) Green manure and long-term fertilization effects on soil zinc and cadmium availability and uptake by wheat (Triticum aestivum L.) at different growth stages. Sci Total Environ 599–600:1330–1343. https://doi.org/10.1016/j.scitotenv.2017.05.070
Haramoto ER, Gallandt ER (2004) Brassica cover cropping for weed management: a review. Renew Agr Food Syst 19:187–198. https://doi.org/10.1079/RAFS200490
Hashemi M, Farsad A, Sadeghpour A et al (2013) Cover-crop seeding-date influence on fall nitrogen recovery. J Plant Nutr Soil Sci 176:69–75. https://doi.org/10.1002/jpln.201200062
Jensen LS, Salo T, Palmason F et al (2005) Influence of biochemical quality on C and N mineralisation from a broad variety of plant materials in soil. Plant Soil 273:307–326. https://doi.org/10.1007/s11104-004-8128-y
Justes E (2017) Cover crops for sustainable farming. Springer, Berlin Heidelberg, New York, NY
Kaye J, Finney D, White C et al (2019) Managing nitrogen through cover crop species selection in the U.S. mid-Atlantic. PLoS ONE 14:e0215448. https://doi.org/10.1371/journal.pone.0215448
Li F, Sørensen P, Li X, Olesen JE (2020) Carbon and nitrogen mineralization differ between incorporated shoots and roots of legume versus non-legume based cover crops. Plant Soil 446:243–257. https://doi.org/10.1007/s11104-019-04358-6
Marcillo GS, Miguez FE (2017) Corn yield response to winter cover crops: an updated meta-analysis. J Soil and Water Conserv 72:226–239. https://doi.org/10.2489/jswc.72.3.226
MiPAAF, Metodi ufficiali di analisi fisica del suolo, GU Serie Generale n.204 del 02–09–1997 - Suppl. Ordinario n. 173.
Osipitan OA, Dille JA, Assefa Y, Knezevic SZ (2018) Cover crop for early season weed suppression in crops: systematic review and meta-analysis. Agron J 110:2211–2221. https://doi.org/10.2134/agronj2017.12.0752
Parr M, Grossman JM, Reberg-Horton SC et al (2011) Nitrogen delivery from legume cover crops in no-till organic corn production. Agron J 103:1578–1590. https://doi.org/10.2134/agronj2011.0007
Patrignani A, Ochsner TE (2015) Canopeo: a powerful new tool for measuring fractional green canopy cover. Agron J 107:2312. https://doi.org/10.2134/agronj15.0150
Perego A, Basile A, Bonfante A et al (2012) Nitrate leaching under maize cropping systems in Po Valley (Italy). Agr Ecosyst Environ 147:57–65. https://doi.org/10.1016/j.agee.2011.06.014
Petroudi ER, Noormohammadi G, Mirhadi MJ et al (2011) Effects of nitrogen fertilization and rice harvest height on agronomic yield indices of ratoon rice–berseem clover intercropping system. AJCS 5(5):566–574
Poeplau C, Don A (2015) Carbon sequestration in agricultural soils via cultivation of cover crops—a meta-analysis. Agr Ecosyst Environ 200:33–41. https://doi.org/10.1016/j.agee.2014.10.024
Ramírez-García J, Carrillo JM, Ruiz M et al (2015) Multicriteria decision analysis applied to cover crop species and cultivars selection. Field Crop Res 175:106–115. https://doi.org/10.1016/j.fcr.2015.02.008
Restovich SB, Andriulo AE, Armas-Herrera CM et al (2019) Combining cover crops and low nitrogen fertilization improves soil supporting functions. Plant Soil 442:401–417. https://doi.org/10.1007/s11104-019-04205-8
Richards IR, Wallace PA, Turner IDS (1996) A comparison of six cover crop types in terms of nitrogen uptake and effect on response to nitrogen by a subsequent spring barley crop. J Agric Sci 127:441–449. https://doi.org/10.1017/S0021859600078667
Rüegg WT, Richner W, Stamp P, Feil B (1998) Accumulation of dry matter and nitrogen by minimum-tillage silage maize planted into winter cover crop residues. Eur J Agron 8:59–69. https://doi.org/10.1016/S1161-0301(97)00013-0
Ruffatti MD, Roth RT, Lacey CG, Armstrong SD (2019) Impacts of nitrogen application timing and cover crop inclusion on subsurface drainage water quality. Agr Water Manage 211:81–88. https://doi.org/10.1016/j.agwat.2018.09.016
Saxton KE, Rawls WJ, Romberger JS, Papendick RI (1986) Estimating generalized soil-water characteristics from texture. Soil Sci Soc Am J 50:1031–1036. https://doi.org/10.2136/sssaj1986.03615995005000040039x
Singh G, Williard KWJ, Schoonover JE (2018) Cover crops and tillage influence on nitrogen dynamics in plant-soil-water pools. Soil Sci Soc Am J 82:1572–1582. https://doi.org/10.2136/sssaj2018.03.0111
Sistemi di produzione integrata nelle filiere agroalimentari (2021). (https://www.regione.lombardia.it/wps/portal/istituzionale/HP/DettaglioRedazionale/servizi-e-informazioni/Imprese/Imprese-agricole/servizio-fitosanitario-regionale/uso-sostenibile-dei-prodotti-fitosanitari/disciplinari-produzione-integrata/02-disciplinari-produzione-integrata.2021)
Spargo JT, Cavigelli MA, Mirsky SB et al (2016) Organic supplemental nitrogen sources for field corn production after a hairy vetch cover crop. Agron J 108:1992–2002. https://doi.org/10.2134/agronj2015.0485
Storr T, Simmons RW, Hannam JA (2020) Using frost-sensitive cover crops for timely nitrogen mineralization and soil moisture management. Soil Use Manage 2020(00):19. https://doi.org/10.1111/sum.12619
Sullivan DM, Andrews ND (2012). Estimating plant-available nitrogen release from cover crops. Pacific Northwest Extension Publication, 636.
Thapa R, Mirsky SB, Tully KL (2018a) Cover crops reduce nitrate leaching in agroecosystems: a global meta-analysis. J Environ Qual 47:1400–1411. https://doi.org/10.2134/jeq2018.03.0107
Thapa R, Poffenbarger H, Tully KL et al (2018b) Biomass production and nitrogen accumulation by hairy vetch-cereal rye mixtures: a meta-analysis. Agron J 110:1197–1208. https://doi.org/10.2134/agronj2017.09.0544
Thomsen IK, Hansen EM (2014) Cover crop growth and impact on N leaching as affected by pre- and postharvest sowing and time of incorporation. Soil Use Manage 30:48–57. https://doi.org/10.1111/sum.12083
Thorup-Kristensen K, Magid J, Jensen LS (2003) Catch crops and green manures as biological tools in nitrogen management in temperate zones. Adv Agron. Academic Press, pp 227–302
Tonitto C, David MB, Drinkwater LE (2006) Replacing bare fallows with cover crops in fertilizer-intensive cropping systems: a meta-analysis of crop yield and N dynamics. Agr Ecosyst Environ 112:58–72. https://doi.org/10.1016/j.agee.2005.07.003
Toom M, Talgre L, Mäe A et al (2019a) Selecting winter cover crop species for northern climatic conditions. Biol Agric Hortic 35:263–274. https://doi.org/10.1080/01448765.2019.1627908
Toom M, Talgre L, Pechter P et al. (2019b) The effect of sowing date on cover crop biomass and nitrogen accumulation. Agr Research 17 4 1779 1787 https://doi.org/10.15159/AR.19.164
Valkama E, Lemola R, Känkänen H, Turtola E (2015) Meta-analysis of the effects of undersown catch crops on nitrogen leaching loss and grain yields in the Nordic countries. Agr Ecosyst Environ 203:93–101. https://doi.org/10.1016/j.agee.2015.01.023
Vicensi M, Lopes C, Koszalka V et al (2020) Soil fertility, root and aboveground growth of black oat under gypsum and urea rates in no till. J Soil Sci Plant Nutr 20:1271–1286. https://doi.org/10.1007/s42729-020-00211-3
Acknowledgements
This work is dedicated to the memory of Dr Mauro Agosti, former head of Condifesa Technical Service, who substantially contributed to develop the project proposal, but passed away before its start. The authors thank Regione Lombardia and the farmers involved in the project.
Funding
This work was supported by the project “BENCO—Demonstration of agronomic, economic and environmental benefits of cover crops in Lombardia,” co-funded by the European Fund for Rural Development under the Rural Development Program 2014–2020 of Regione Lombardia, Operation 1.2.01, Managing Authority: Regione Lombardia, https://sites.unimi.it/benco/.
Author information
Authors and Affiliations
Contributions
Conceptualization, T.T., L.B., P.M., and L.M.; methodology, L.B., P.M., and A.P.; investigation, T.T., L.B., P.M., E.P., L.M., and D.D.T.; writing—original draft, T.T., L.B., and E.P.; writing—review and editing, T.T, L.B., E.P., P.M., and A.P.; funding acquisition, L.B., P.M., and L.M.; supervision, L.B. and P.M.
Corresponding author
Ethics declarations
Ethics approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Conflict of interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
About this article

Cite this article
Tadiello, T., Potenza, E., Marino, P. et al. Growth, weed control, and nitrogen uptake of winter-killed cover crops, and their effects on maize in conservation agriculture. Agron. Sustain. Dev. 42, 18 (2022). https://doi.org/10.1007/s13593-021-00747-3
Accepted:
Published:
DOI: https://doi.org/10.1007/s13593-021-00747-3