
Abstract
Sexual reproduction is the basic way to form high genetic diversity and it is beneficial in evolution and speciation of fungi. The global diversity of teleomorphic species in Ascomycota has not been estimated. This paper estimates the species number for sexual ascomycetes based on five different estimation approaches, viz. by numbers of described fungi, by fungus:substrate ratio, by ecological distribution, by meta-DNA barcoding or culture-independent studies and by previous estimates of species in Ascomycota. The assumptions were made with the currently most accepted, “2.2–3.8 million” species estimate and results of previous studies concluding that 90% of the described ascomycetes reproduce sexually. The Catalogue of Life, Species Fungorum and published research were used for data procurement. The average value of teleomorphic species in Ascomycota from all methods is 1.86 million, ranging from 1.37 to 2.56 million. However, only around 83,000 teleomorphic species have been described in Ascomycota and deposited in data repositories. The ratio between described teleomorphic ascomycetes to predicted teleomorphic ascomycetes is 1:22. Therefore, where are the undiscovered teleomorphic ascomycetes? The undescribed species are no doubt to be found in biodiversity hot spots, poorly-studied areas and species complexes. Other poorly studied niches include extremophiles, lichenicolous fungi, human pathogens, marine fungi, and fungicolous fungi. Undescribed species are present in unexamined collections in specimen repositories or incompletely described earlier species. Nomenclatural issues, such as the use of separate names for teleomorph and anamorphs, synonyms, conspecific names, illegitimate and invalid names also affect the number of described species. Interspecies introgression results in new species, while species numbers are reduced by extinctions.
Similar content being viewed by others


Introduction
Ascomycota Caval-Sm. is the largest fungal phylum comprising around 93,000 extant species and are generally known as “sac fungi” (Bennett and Turgeon 2017; Clark et al. 2018; Catalog of Life 2021). Members of Ascomycota are ubiquitously spread in various terrestrial and fresh or marine ecosystems (Naranjo-Ortiz and Gabaldón 2019). Most ascomycetes are saprobes while some are soil or dung inhabitants (Richardson 2019). Some are animal, human and plant pathogens or parasites such as epiphytes or fungicolous fungi (Wu et al. 2011), while others are symbionts as endophytes, lichenicolous and mycorrhizae (Lawrey and Diederich 2003; Chomnunti et al. 2014; Kim et al. 2017; Sun et al. 2019; Hyde et al. 2020b). The sexual reproduction in ascomycetes often occurs as a response to adverse environmental conditions (Nieuwenhuis and James 2016) and it results high genetic diversity between species (Lee et al. 2010). Sexual reproduction helps to purge deleterious mutations and also selects beneficial mutations to adapt to a fluctuating environment (Otto and Lenormand 2002). Ascospores are more resistant to environmental stress and more widely dispersed than the conidia (Kirschner 2019).
Sexual reproduction in ascomycetes comprising the mating-type loci (MAT) which encodes key transcription factor genes that govern speciation (Paoletti et al. 2005). Two compatible partners fuse their genetic materials by recombination or crossing-over and also meiosis and mitosis to produce genetically diversified offspring (O'Gorman et al. 2009). Sexual reproduction occurs in the same mycelium (homothallic/self-fertile) or two different mycelia (heterothallic) and they produce spores in a sac-like structure called an ascus (Fig. 1). The sexual structures such as asci and ascospores are contained in fruiting bodies. Released ascospores from fruiting bodies germinate when contact with a suitable substrate and produce the mating type male (+) and female (−) mycelia (Taylor et al. 2006). The male mycelium produces an antheridium and the female produces an ascogonium, which are different reproductive organs.

Sexual reproduction of filamentous ascomycetes. a Reproduction cycle. b Ascus formation within fertilized ascogonium (drawn from Peraza-Reyes and Berteaux-Lecellier 2013)
Sexual reproduction in Ascomycota includes three stages as plasmogamy, karyogamy and meiosis (Wallen and Michae 2018). During the first step (plasmogamy), two haploid cells are fused and lead to a dikaryotic stage where two haploid nuclei coexist in a single cell. During the second step (karyogamy), the haploid nuclei of + and − mycelia fuse to form a diploid zygote nucleus. The diploid zygote nucleus produces unique haploid nuclei by meiosis and these haploid nuclei form haploid spores with mitosis and cell division within the sac-like structures called ascus. These ascospores are then released and germinate to form new hyphae in new environments. The production of these highly specialized sexual tissues is initiated and controlled by gene, protein, and secondary metabolite networks and those proteins regulate the expression of primary MAT genes (MAT1-1-1 and MAT1-2-1) (Dyer 2007). Both MAT genes are typically essential for successful fertilization and ascomatal development (Ferreira et al. 1998). Additionally, the MAT1-1-1 gene is also critical for ascospore production in some species (Debuchy et al. 2010) and alteration of both MAT genes results in failure to form even immature sexual structures (Lee et al. 2003).
Some anamorphic ascomycetes shuffle their genetic material by parasexuality. Parasexual reproduction results in recombination of genes from different individuals but does not involve meiosis and formation of a zygote by fertilization as in sexual reproduction. Parasexuality generates both genotypic and phenotypic diversity in species (Hirakawa et al. 2017). The variations in genotypes create by shuffling of different chromosome homologs, recombination between homologs and the generation of cells in various ploidy states with one or more supernumerary chromosomes (Forche et al. 2008). The mutations in genetic materials and haploidization occur inside the heterokaryotic hyphae prior to conidial formation (Becker and de Castro-Prado 2006). However, the mixing-up genetic materials without forming sexual structures does not discuss here and this study only estimates species formed by true sexual reproduction.
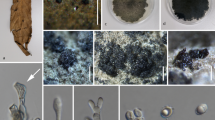
Teleomorphic ascomycetes are morphologically diversified (Fig. 2) and ubiquitous taxa that can survive in various ecological habitats in both terrestrial and aquatic ecosystems (Gould 2008; Schoch et al. 2009). Teleomorphic species are reported from only 17 classes in Ascomycota including all classes in subphylum Pezizomycotina and Neolectomycetes in subphylum Taphrinomycotina.

Diversity of fruitting bodies in teleomorphic Ascomycota. a, j, k, p, t, w, x, z Leotiomycetes, d Neolectomycetes, b, h, m, y Dothideomycetes, n, r Eurotiomycetes, e Geoglossomycetes, o Orbiliomycetes, q, v Pezizomycetes, c, f, g, i, l, s, u Sordariomycetes
Why should we estimate the global species richness of teleomorphic ascomycetes?
In mycology, “species” is simply defined as a diagnosable cluster of individuals within a parental pattern of lineage displays a pattern of phylogenetic ancestry and descent among units and hence, it is valuable to give it a species name (Brown 2002; Aldhebiani 2018; Maharachchikumbura et al. 2021). Finalizing the global fungal inventory is a challenge due to their morphological, ecological and physiological diversity (Purvis and Hector 2000). Estimates of the total number of teleomorphic ascomycetes have major inferences for systematics, resource management and classification (Hawksworth 1991) as they play key roles in ecosystems as decomposers, mutualists and pathogens individually and with the interactions of each other (Schmit and Mueller 2007) and some of them in the plant rhizosphere protect root systems from pathogens (Mendes et al. 2013). Mutualistic ascomycetes are associated with their host without causing harm. It is a beneficial relationship for both fungi and the host (Volk 2013). Ascomycetes associate with algae or cyanobacteria to form lichens (Weber and Büdel 2011) and Arthoniomycetes, Dothideomycetes, Eurotiomycetes, Lecanoromycetes, Lichinomycetes and Sordariomycetes comprise the lichenized species (Grube and Winka 2002; Andersen and Ekman 2005). More than 40% of lichenized fungi are species in Ascomycota (Brodo et al. 2001; Schoch et al. 2009). Mycorrhizae are symbiotically associated with plant roots while endophytes are associated with living plants (Volk 2013).
There are numerous of plant pathogenic teleomorphic ascomycetes causing various diseases of economic crops and forest trees (Lu et al. 2003). The increasing number of virulent fungal infectious diseases is regarded as a worldwide threat to food security (Hyde et al. 2018). An unprecedented number of diseases caused by fungi including teleomorphic ascomycetes have resulted in some of the most severe diebacks in economic crops and wild species (Fisher et al. 2012; Hyde et al. 2019). Many species can be harmless endophytes in some plants, however cause severe damages in others (Hardoim et al. 2015; Terhonen et al. 2019; Song et al. 2021). Therefore, description and cataloging of teleomorphic ascomycetes helps to identify fungal pathogens and prevent future disasters.
The nutritional sources of the teleomorphic ascomycetes vary from dead organic matter to synthesized compounds by other organisms and they decompose litter, maintain the nutrient cycles and improve soil quality (Gams 2007; Gould 2008; Frąc et al. 2018; Senanayake et al. 2020a). However, the mycota involved in decomposition incompletely known and there may be many species interactions (Frey-Klett et al. 2011; Volk 2013). Hyaloscyphaceae, Melanommataceae, Mytilinidiaceae and Savoryellaceae are some ascomycetous families which have many saprobic teleomorphic species (Hernández-Restrepo et al. 2017). Identifying and describing the teleomorphic ascomycetes involved in litter degradation is important in organic farming and fertilizer production (Peyvast et al. 2008). Isolation of those fungi and application to soil as a microbial assortment to enhance plant growth and yield is required (Khalid et al. 2017). Additionally, antibiotics, anticancer, anti-inflammatory and some medically important chemicals are extracted from teleomorphic ascomycete cultures (Rajamanikyam et al. 2018; Al-Fakih and Almaqtri 2019; Wu et al. 2019b). Many organic acids and enzymes such as citric acid, gluconic acid, amylases and proteases are produced by teleomorphic ascomycetes. Morels, truffles, Hypomyces lactifluorum are edible ascomycetes (Acton and Sandler 2008; Splivallo et al. 2010). Therefore, revealing the undescribed teleomorphic ascomycetes, estimating the species number and exploring their chemical and biological properties are important (Yang et al. 2018).
How many teleomorphic species in Ascomycota based on different estimation methods?
Estimated number of teleomorphic ascomycetes based on numbers of described fungi
Traditionally, taxonomic studies of teleomorphic ascomycetes were based mostly on morphological characters, subcellular arrangement, bio-chemical, physiological and ecological studies (Yang 2011; Maharachchikumbura et al. 2021). During 1960–2000, phenotypic taxonomic studies were improved by microscopy and in vitro culturing (Klopfstein 2016). Many groups of teleomorphic ascomycetes have been intensively studied (Todd et al. 2014) and simultaneously significant taxonomic monographs were published (Barr 1978, 1987; Kohlmeyer and Kohlmeyer 1979; Schmit and Lodge 2005; Senanayake et al. 2017, 2018). In the previous 20 years, molecular methods have modernized and studies are based on biogeography, phylogeny, population genetics, systematics and taxonomy (Yang 2011).
Since 1982, there has been a periodic update in the classification of taxa in Ascomycota especially in the Journal Systema Ascomycetum. The Species Fungorum database (http://www.speciesfungorum.org) has recorded the number of new species described each year. The taxonomy of the phylum Ascomycota has been updated at a fast pace over the last few years (Hyde et al. 2013, 2020a, b, c; Jaklitsch et al. 2016; Maharachchikumbura et al. 2016; Liu et al. 2017; Ekanayaka et al. 2018). Accepted families with descriptions and list of genera in the Ascomycota were provided by Jaklitsch et al. (2016). Currently there are 22 classes in the phylum Ascomycota as Archaeorhizomycetes, Arthoniomycetes, Candelariomycetes, Coniocybomycetes, Dothideomycetes, Eurotiomycetes, Geoglossomycetes, Laboulbeniomycetes, Lecanoromycetes, Leotiomycetes, Lichinomycetes, Neolectomycetes, Orbiliomycetes, Pezizomycetes, Pneumocystidomycetes, Saccharomycetes, Sareomycetes, Schizosaccharomycetes, Sordariomycetes, Taphrinomycetes, Xylobotryomycetes and Xylonomycetes (Lumbsch and Huhndorf 2010; Voglmayr et al. 2019; Beimforde et al. 2020).
The “2.2–3.8 million” species estimate (Hawksworth and Lücking 2017) is considered as the most rational estimate by many mycologysts (Hyde et al. 2020b). There are approximately 150,000 extant fungal species (Roskov et al. 2019; Species Fungorum 2021), however this is only 15–26% of the estimated species (Hyde et al. 2020b). Early mycologists believed that only half of ascomycetes are meiosporic fungi which obligatory sexually reproduced and do not produce asexual spores (Reynolds and Taylor 1993). However, the rest are probably obligatory mitosporic or facultative mitosporic fungi with undetected teleomorphs (Nieuwenhuis and James 2016). Reynolds and Taylor (1993) showed that about 5% of obligatory anamorphic ascomycetes are known to be pleomorphic and thus discretionary sexually reproduce. However, the most accepted value is that may be the 90% of the described ascomycetes sexually reproduce (Judson and Normark 1996; Normark et al. 2003).
There are 92,725 described species in Ascomycota (Catalog of Life 2021; Species Fungorum 2021). Wijayawardene et al. (2017) listed 8897 species in Ascomycota with undetermined teleomorphs. There are 523 anamorphic ascomycetes with undetermined teleomorph have been introduced from 2018 to 2020 (31 December 2020) (Species Fungorum 2021). Hence, 9420 ascomycetous species are obligatory anamorphic species and therefore around 83,305 teleomorphic ascomycetes have been described. The reproduction arrangements appear to be similar across the phylum Ascomycota even though the life cycles among the major groups are different (Nieuwenhuis and James 2016). Hence, there should be 1.25–2.17 million teleomorphic ascomycetes based on described number of species considering the 2.2–3.8 million species estimate.
Estimated number of teleomorphic ascomycetes based on fungus:substrate or host ratio
The ratio of fungal species to each plant species was one of the key elements in estimating global species richness as 1.5 million (Hawksworth 1991) and this estimate was assumed from independent data sets which shown that the number of fungi in all environments was six times higher than the vascular plants present, inferred on a global scale. The “1.5 million estimate” was considered too low because, the number of plant species and the fungus: plant ratios were too conservative and many were collected from other substrata such as insects (Hawksworth 2001). The number of plant species has increased from 250,000 to 390,000 (Pimm and Joppa 2015) signifying that the estimated number of fungal species must rise to 2.4–3 million. Some authors also suggested that the ratio of fungi: plants is about 10:1, those found in soil, insects or lichen were excluded (O’Brien et al. 2005; Blackwell 2011). However, there are several complications in the fungus: plant ratio concept. The total inventory of species described in a particular area increases gradually, while the number of plant species remains more or less unchanged (Hawksworth and Lücking 2017). Therefore, the fungus: plant ratio decreases gradually with the description of new species.
Some studies based on meta-DNA sequencing of decaying litter samples showed that the fungus: plant ratio is 13:1 (Hawksworth and Lücking 2017). Therefore, the actual ratio in a particular area may be significantly higher than the ratio indicated by traditional inventory techniques. Further, the whole planet has not been screened evenly and known species number in some places is higher than in others. For example, fungal diversity in North America, Europe and Japan is well-studied compared to South Asia and Africa (Hawksworth 2001; Větrovský et al. 2019). O’Brien et al. (2005) noted that the fungus: plant ratios of two forests in North Carolina gave 19:1 and 13:1 suggesting that there may be 3.5 to 5.1 million species. Further, this ratio changes according to the substrata and Taylor et al. (2010) showed that fungus: plant ratio is 7.5:1 in forest soils in Alaska. Tedersoo et al. (2014) analyzed soil samples using meta-barcoding molecular methods and concluded that the number of species had been overestimated by 1.5 to 2.5 times from data based on plant: fungus species ratios.
Therefore, the estimates based on studies of fungus: plant/insect/lichen/plant OTUs in soil ratios in a site, obtained by field survey and molecular approaches, have generated lower ranges from 0.42 to 3.5 million to (O’Brien et al. 2005; Tedersoo et al. 2014) an upper range from 0.6 to 5.1 million (O’Brien et al. 2005; Piepenbring et al. 2012) (Table 1). Considering the average of the upper and lower range of previous estimates, we estimated species number is 1.96–2.85 million based on fungus: host ratio. There are around 150,000 described species and 92,725 are ascomycetes, which is around 63.4%. Therefore, there should be about 1.11–1.62 million estimated teleomorphic ascomycetes (Table 1), excluding 10% obligatory anamorphic species (Normark et al. 2003).
The patterns of introducing new species are biased with more described from economically important plants (Cannon and Hawksworth 1995). Most early described species were collected from temperate floral communities and host specificity in tropical plants are not well-reported and new host records are not published (Tedersoo et al. 2010; Piepenbring et al. 2011). Therefore, fungus: plant/insect/substrate ratio is not an ideal method to estimate species numbers because of uneven exploration of global species in habitats.
Estimated number of teleomorphic ascomycetes based on ecological distribution
The traditional quantification approaches of teleomorphic ascomycetes are established on the opportunistic collections of specimens based on host, substrate, area and transects (Schmit and Lodge 2005). Opportunistic collecting requires highly trained collectors who can recognize taxa in the field without a bias. Some collectors only perceive favored particular groups of teleomorphic ascomycetes. Further, conspicuous species and more common species are often overlooked (Lodge et al. 2004). Teleomorphic ascomycetes produce fruit bodies in different types of substrata (Lodge 1996; Huhndorf and Lodge 1997; Schmit and Lodge 2005; Sainz et al. 2018). Some ascomycetes sexually reproduce rather dependably while others do so only occasionally, and therefore require long periods to be recorded from a particular area (Straatsma et al. 2001). Further, fruiting patterns, abundance and dispersion of ascomycetes differs among substrata (Lodge et al. 2004).
Some teleomorphic ascomycetes show a wide range of host and substrate variation and also different modes of life. Daldinia eschscholtzii, one of the common endophytes in plants (Stadler et al. 2014; Helaly et al. 2018) and marine algae (Tarman et al. 2012), has been reported as an endosymbiont of a mantis gut (Zhang et al. 2011), and a human pathogen (Chan et al. 2015). Further, Diaporthe sojae, a known pathogen of soybean, was also isolated from infected skin of an immunocompromised patient after kidney transplantation (Garcia-Reyne et al. 2011). Diaporthe toxica is a plant endophyte and occasionally a plant pathogen (Williamson et al. 1991) and produces secondary metabolites that result in toxicoses of animals such as liver disease known as lupinosis of sheep (Gardiner 1975; Allen and Wood 1979; Williamson et al. 1994). Therefore, it is necessary to understand the ecology and life strategies of teleomorphic ascomycetes before estimating the species number. Further, many endophytes do not sporulate in culture (Sun and Guo 2012) and some ascomycetes change colony morphology while growing and sub-culturing on different media (Senanayake et al. 2017). Some ascomycetes do not sexually reproduce or need specific conditions for sexual reproduction (Sun and Heitman 2011). Direct morphological examination of fruiting structures on substrata or media only is therefore biased in estimating number of teleomorphic ascomycetes (Guo et al. 2001; Promputtha et al. 2004).
Case studies from marine ascomycetes
Marine ascomycetes are recovered repeatedly from marine habitats, able to grow and/or sporulate on substrata in marine environments, form symbiotic relationships with other marine organisms, adapt and evolve at the genetic level or be metabolically active in marine environments (Pang et al. 2016). They are observed in a range of marine substrates, including mangrove plant wood and leaves, driftwood, saltmarsh plants, algae, dead coral, and sand grains on beaches (Gonçalves et al. 2021; Walker and Robicheau 2021), along with severe marine ecosystems such as deep-sea trenches, hydrothermal vents, deep-sea subsurfaces, cold methane seeps and hypersaline, anoxic, and suboxic waters (Raghukumar and Ravindram 2012; Xu et al. 2018). Marine ascomycetes colonize a variety of substrata based on their ability to degrade complex substrata such as lignocellulose, keratin, chitin and calcareous structures and ascomycetes are the major decomposers in marine ecosystems (Kohlmeyer and Volkmann-Kohlmeyer 2001; Walker and Campbell 2010). Marine ascomycetes are also known as symbionts and pathogens of marine algae and marine fauna (Hyde et al. 1998).
The accessibility and the nature of substrate for colonization, competition, pH, temperature, and saltiness of water affect the diversity of marine ascomycetes (Jones 2000, 2011). Most marine fungi are recognized to have a cosmopolitan distribution (Pugh and Jones 1986). However, basic biogeographic diversity data are lacking for marine ascomycetes in most parts of the world (Walker and Robicheau 2021). Some marine fungi such as Aniptodera chesapeakensis Shearer & M.A. Mill., Ceriosporopsis Halima Linder, Corollospora maritima Werderm., Lignincola laevis Höhnk, Savoryella lignicola E.B.G. Jones & R.A. Eaton, and Torpedospora radiata Meyers have diverse geographic dispersion which is classified as tropical to subtropical while Lulwoana uniseptata (Nakagiri) Kohlm. et al. is reported from temperate habitats only (Torta et al. 2015; Tibell et al. 2020).
Mora et al. (2011) presented an approach to estimate species numbers on earth and ocean and predicted that 0.005 million species are marine. However, 91% of species in the ocean await description and increasing the sampling intensity is required to characterize the underexplored species of marine biodiversity (Walker and Robicheau 2021). It is estimated that more than 10,000 marine fungal species exist globally (Jones 2011; Walker et al. 2017) and only around 1000 have been described (Jones et al. 2015; Pang et al. 2016). Jones et al. (2019) listed 1257 marine species belonging to 539 genera and 943 of them are ascomycetes (Jones et al. 2009, 2015; Abdel-Wahab et al. 2010; Pang et al. 2010; Abdel-Wahab and Nagahama 2011; Dayarathne et al. 2016, 2019).
The number of species is estimated as 2.2–3.8 million (Hawksworth and Lücking 2017) while only around 150,000 species have been described (Species Fungorum 2021). If there are 943 described marine ascomycetous species, then it is predicted that 13,831–23,889 marine ascomycetes should be in oceans. However, considering only 90% of described ascomycetes are sexually reproduced (Judson and Normark 1996; Normark et al. 2003), then there are 12,448–21,500 marine, teleomorphic ascomycetes.
Case studies from freshwater ascomycetes
Freshwater ascomycetes are an ecological assortment rather than a taxonomic group and they reproduce sexually or asexually residing on sunken or partially submerged woody substrata in freshwater environments (Tsui et al. 2016; Calabon et al. 2021). In spite of their importance as decomposers and food sources in freshwater food webs, there has been little research on their global distribution, community structure and species diversity (Shearer et al. 2015). Freshwater ascomycetes occur on submerged or partially submerged substrata in lotic and lentic aquatic habitats. The teleomorphic ascomycetes are more dominant on submerged wood, while the anamorphic ascomycetes occur on submerged leaf litter (El-Elimat et al. 2021).
Phylogenetically, freshwater ascomycetes are grouped mostly throughout the class Dothideomycetes, Leotiomycetes and Sordariomycetes in Ascomycota (Shearer et al. 2009, 2014). They have soft fruiting bodies during teleomorphic stage with appendage baring ascospores (Hyde et al. 1998). Asci developed in ascomata possess pathways for efficient spore discharge and ascospores are frequently appendaged or have sheaths. The appendages facilitate spore dissemination and bonding to the substrata (Hyde and Goh 2003).
Freshwater ascomycetes are observed across the both lentic and lotic ecosystems, and they are commonly associated as endophytes and parasites on algae and aquatic macrophytes along with the saprobes on the dead plant matter (Lu et al. 2018). Many freshwater ascomycetes are believed to have evolved from terrestrial ancestors through a wide range of evolutionary pathways (Vijaykrishna et al. 2006; Grossart et al. 2019).
The number of aquatic species has been estimated as 0.5–10 million based on molecular data (Bass and Richards 2011; Blackwell 2011; Mora et al. 2011). A significant number of freshwater species are teleomorphic ascomycetes (Shearer et al. 2007; Hu et al. 2013; Shearer and Raja 2021). About 738 species of freshwater ascomycetes are known from their teleomorph, belonging to approximately 170 genera (El-Elimat et al. 2021; Shearer and Raja 2021). There are around 83,305 described teleomorphic ascomycetes (Species Fungorum 2021) and if there are 738 described freshwater teleomorphic ascomycetes, it is estimated that there are 19,490–33,664 aquatic teleomorphic ascomycetes based on “2.2–3.8 figure”.
Case studies from insect-associated ascomycetes
Insects are an extremely diversified group of organisms in all ecosystems (Stork 1988) and include dragonflies, mayflies, grasshoppers, cockroaches, termites, stoneflies, true bugs, flies, beetles, butterflies, moths, ants, bees, and wasps (Stork et al. 2015, 2018). Insects and fungi share a long history of relationship in the similar habitats and (Bourtzis and Miller 2003) those interactions can be mutualistic or harmful. Insects involved in associations with fungi include members of the Coleoptera, Diptera, Homoptera, Hymenoptera, and Isoptera.
Fungal biotrophic parasites of insects are rare, except for the very successful associations of Laboulbeniomycetes (Blackwell et al. 2020; Haelewaters et al. 2021). Vega and Dowd (2005) highlighted the role of yeast-insect endosymbionts in supporting the digestion and detoxification of plant materials ingested by insects and discovered an enormous number of species of Saccharomycetes. Some fungi also interact with insects by providing nutritional supplements (Vega and Blackwell 2005). Suh et al. (2001) described around 200 new yeast species from the gut of beetles. It is suspected that these yeasts also might provide nutritional supplements.
Insect associated fungi were estimated to be 1.5 million (Hywel-Jones 1993) and Stork (2018) updated this to 5.5 million, while 1–2% of them may be cryptic species. Therefore, including the cryptic species, the consensus estimate of insect associated species ranges from 5.505 to 5.511 million. However, this is more deviated from currently estimated species numbers. The diversity of insect associated ascomycetes has been extensively studied (Aung et al. 2008; Mora et al. 2011; Hyde et al. 2018) and they are taxonomically distributed in Clavicipitaceae, Cordycipitaceae, and Ophiocordycipitaceae in Hypocreales, ambrosia fungi (e.g., Ceratocystis, Ophiostoma) in the Ophiostomatales, all families in Laboulbeniomycetes and some species in Saccharomycetes (Sung et al. 2007; Vega et al. 2012; Araújo and Hughes 2016; Maharachchikumbura et al. 2016; Wijayawardene et al. 2018). Mueller and Schmit (2007) estimated around 50,000 insect associated species, when there are 750 described species. However, currently, there are more than 4000 insect-associated species described (Species Fungorum 2021). Hence, there should be 52,800–91,200 insect associated teleomorphic ascomycetes according to “2.2–3.8 species estimate” and considering generally 90% of ascomycetes reproduce sexually (Judson and Normark 1996; Normark et al. 2003).
Case studies from coprophilous ascomycetes
Coprophilous fungi grow, sporulate and germinate on herbivore dung (Tretter et al. 2014; Lazarus et al. 2017) and they are specialized to survive in the harsh environment of the gastrointestinal tract of animals (Richardson 2001b; Bell 2005; Kirschner et al. 2015; De Souza et al. 2017; Lavrinienko et al. 2021). Coprophilous fungi recycle the nutrients in animal dung and release nutrients to the soil (Basumatary and McDonald 2017; Florenzano 2019).
Species richness and composition of coprophilous ascomycetes differ with abiotic and biotic factors. Intra- and inter-specific interactions in a dung pile affect fungal succession and species composition (Maynard et al. 2018; Lavrinienko et al. 2021). Many coprophilous ascomycetes are most common on only one or a few dung types (Lundqvist 1972) and dung from animals that live together generally show a similar species composition (Richardson 2001a).
However, the fungal community varied more between animal dung types than between the various grassland habitats (Angel and Wicklow 1983). Coprophilous ascomycetes can be found more frequently on dung of herbivores than carnivores (Lundqvist 1972; Richardson 2001a). In addition, they have seldom been reported on reptile or amphibian dung, indicating that coprophily in fungi developed among the warm-blooded animals (Webster 1970). Some ascomycetes are strictly coprophilous and they have a distinct life-cycle restricted to dung pile, plant surface and animal gut (Wicklow 1992). However, some spores disperse in soil. The spores of coprophilous ascomycetes are highly pigmented, with thick walls and are protected against the harmful ultraviolet sunlight (Ingold and Hudson 1993). Therefore, spores remain in soil alive and those species have been reported as soil fungi.
Kruys (2005) reported that many coprophilous ascomycetes belong to order Pleosporales in Dothideomycetes and three of the families are solely or mostly coprophilous, viz. Delitschiaceae, Phaeotrichaceae and Sporormiaceae. Calaça et al. (2015) listed 143 coprophilous ascomycetous species recorded from Brazil. Melo et al. (2020) studied the diversity and species richness of coprophilous fungi in Brazil. A total of 271 species are reported from dung substrata and among them, 70% of recorded species are ascomycetes. Most species are included in Sordariales, Hypocreales and Microascales and 9% of recorded species are anamorphic ascomycetes (Saumell et al. 1999, 2000; Saumell and Padilha 2000).
Calaça et al. (2020) listed the coprophilous species recorded in Brazil during 1900–2013 and 117 from 210 coprophilous species are ascomycetes. They were collected from 12 states of Brazil and total area of these 12 states is 303146 km2. Therefore, one coprophilous ascomycetous species was collected from each 2591 km2. If this value inferred for land area of earth, there should be 196,874 coprophilous ascomycetes. If 9% of recorded species are anamorphic species (Saumell et al. 1999, 2000; Saumell and Padilha 2000), then there should be 177,187 teleomorphic coprophilous ascomycetes. However, this value is 443,061 according to Melo et al. (2020). Therefore, it is assumed that there should be around 177,000–443,000 teleomorphic coprophilous ascomycetes.
Case studies from soil ascomycetes
Fungi occur in the soil or soil-associated environments at least for some stage in their life-cycle known as soil fungi (Bridge and Spooner 2001). They are active, freely growing fungi closely associated with other organisms or inactive dormant propagules (Rämä and Quandt 2021). The role of soil fungi are an extremely complex and are fundamental to the soil ecosystem (Hawksworth et al. 1995). Soil fungi carry out many different functions in soils such as the degradation of dead organic matter, binding soil particles to improve the aeration, water penetration, destroy soil pathogens, and improve soil health by formation of propagules (Zin and Badaluddin 2020; Jayaraman et al. 2021).
Soil fungi can only be consistently identified if they produce fruiting bodies (Hibbett et al. 2016) and conventional techniques are unable to reliably identify the species that are assumed to be present in any given soil sample due to the fastidious nature of the great majority of species (Wardle and Lindahl 2014). Fungal communities in soil can be extremely species rich and patchy at small spatial scales (Taylor and Sinsabaugh 2015). High throughput sequencing of soil fungi in boreal forest sites revealed around 300 taxa in 0.25 g soil and the dominant taxa in the sites were quite distinct from each other (Taylor and Sinsabaugh 2015).
Around 80% of all soil-inhabiting taxa cannot be identified to species and 20% cannot be reliably assigned to known orders (Hawksworth 2001; Vartoukian et al. 2010; Tedersoo et al. 2014, 2017). The number of soil fungal species is considerably greater than the described amount and studies with the integration of molecular, genetic and ecological factors may reveal more species. The number of species identified by traditional culture dependent methods doubles when the same soil samples are analysed by culture-independent methods (Lord et al. 2002; Arenz et al. 2006; Malosso et al. 2006; Smith and Jaffee 2009; Zachow et al. 2009; Hirsch et al. 2013; Rodolfi et al. 2016).
There are no significant estimates of the number of species made for soil fungi. Gilman (1957) included around 700 fungal species which grew on only non-selective media by soil dilution method and many species were later included (Barron 1968; Domsch et al. 1993). Watanabe (1994) suggested that at least 1200 species have been isolated from soil. Pugh (1969) showed that only 17% of soil fungi can be readily grown in culture media. Therefore, Hawksworth (1991) estimated that around 7000 species could be considered as soil fungi based on Watanabe (1994). However, there are more than 80,000 fungal species so far named and described, and they are likely to occur in the soil environment at some stage in their life-cycle (Bridge and Spooner 2001). These species are mainly distributed in the subphylum Taphrinomycotina as the fission yeasts, animal and plant pathogens, the root-associated, sporocarp-forming, filamentous fungi (Schoch et al. 2009), while the Saccharomycotina includes the budding yeasts. Pezizomycotina contains lichen-forming fungi, mycorrhizal fungi, dark-septate endophytes, pathogens and saprotrophs. Further, Bridge and Spooner (2001) proposed that at least 10% of the described fungal species are obligatory soil fungi and around 75% of them are ascomycetes (Taylor and Sinsabaugh 2015). If this applies to 2.2–3.8 estimate considering 90% of them are teleomorphs (Normark et al. 2003), there should be 148,500–256,500 teleomorphic ascomycetes in soil.
Case studies from lichenized ascomycetes
Lichens are stable self-supporting associations of a mycobiont and a photobiont (Maria et al. 2021). They produce many secondary metabolites such as phenolic compounds, dibenzofurans, depsides, depsidones, depsones, lactones, quinones and pulvinic acid derivatives which are accumulated externally on the hyphae rather within the cells (Tehler and Irestedt 2007). These compounds are unique to each species and can be used as food, fodder, dyes, and pharmaceuticals. The lichens are the best bio-indicators of air pollution (Garty 2001). The mycobiont is usually an ascomycete but in a few cases it is a basidiomycete. The photosynthetic partners are generally green algae or cyanobacteria (Richardson 2002). The relationship between fungi and lichens can be endolichenic and lichenicolous (Tripathi and Joshi 2019).
Most of the endolichenic fungi and other accessory fungi reported from inside the lichen thalli are phylogenetically distinct from lichenicolous fungi (Miadlikowska et al. 2004) and more closely related to endophytic ascomycetes in vascular plants (Miadlikowska and Lutzoni 2004). Generally, most of the lichenized fungi belong to Ascomycota or rarely to Basidiomycota. Further, the ascolichens mainly belong to Sordariomycetes, Lecanoromycetes and Eurotiomycetes. The Lecanoromycetes is almost an entirely lichenized class comprising the remarkable population of lichen-forming species (Nash 2008).
A study of the diversity and distribution of the fungal communities that were associated with seven lichens in the Ny-Ålesund Region (Svalbard, High Arctic) using Roche 454 pyrosequencing method reported 370 OTUs of which 294 belonged to Ascomycota (Zhang et al. 2015). Among these, Leotiomycetes, Dothideomycetes, and Eurotiomycetes were the major classes, with Helotiales, Capnodiales, and Chaetothyriales as the dominant orders. Further, Wang et al. (2016) studied fungal diversity associated with a common lichen Hypogymnia hypotrypa in China and 28 were ascomycetes from 50 species. It is assumed roughly that lichen: fungi ratio as 1:45 (Fernández-Mendoza et al. 2011; Muggia and Grube 2018).
There are around 20,000 described lichen species (Çobanoğlu et al. 2010) and about 98% have an ascomycetous mycobiont (François et al. 2001). Therefore, there should be 19,600 endolichenic ascomycetous species. Further, about 40% of species in the Ascomycota are lichenized or lichenicolous fungi (Kirk et al. 2008). Currently, there are around 93,000 described species in Ascomycota and 40% is 37,200. However, excluding 10% obligatory anamorphic species, there are around 17,640–33,480 teleomorphic lichenized or lichenicolous species in Ascomycota.
Case study from ligninolytic ascomycetes
Fungi play a vital role in plant litter decomposition in ecosystems (Boddy et al. 2008; Watkinson et al. 2015; Baldrian 2017). They can degrade different types of organic compounds in the litter (Baldrian and Lindahl 2011), which other organisms are unable to degrade (de Boer et al. 2005). Ligninolytic fungi produce several kinds of extracellular enzymes that help to degrade cellulose and other organic compounds in litter and helps nutrient turnover (Sinsabaugh et al. 2002; Romaní et al. 2006). Most common extracellular enzymes produced by ligninolytic fungi are α-glucosidase, β-glucosidase, cellobiosidase, xylosidase, polyphenol oxidase, N-acetyl-polyphenol oxidase, N-acetyl-β-glucosaminidase and acid phosphatase (Marx et al. 2001; De-Forest 2009).
The majority of ligninolytic fungi are ascomycetes (Seena et al. 2019), that colonize during the early stages of decomposition (Aneja et al. 2006; Voříšková and Baldrian 2013; Prakash et al. 2015). It was proposed that ascomycetes dominate during the initial stages of litter decay presumably due to a superior ability to degrade cellulose (Weber et al. 2011) and decreases during the process of degradation as they are gradually replaced by other non-ascomycetous saprobes (Frankland 1998; Osono 2007). The classes Dothideomycetes, Eurotiomycetes, Saccharomycetes and Taphrinomycetes are the most ligninolytic species abundant classes in Ascomycota (Zhang et al. 2018). A few studies have denoted that endophytes living in plants shift their lifestyle to saprotrophs when the substrates die and they play a key role in early stage decomposition (Purahong and Hyde 2011; Fesel and Zuccaro 2015; Purahong et al. 2016; Szink et al. 2016). However, NGS data may partly reflect the fungal succession from ascomycetes to other fungi in early to later stages of litter decomposition and it does not clearly provide an idea about the species richness and abundance (Amend et al. 2010; Peršoh 2015).
Many studies have been revealed the species composition in decaying litter. However, the number of ligninolytic fungal species has not been estimated. Haňáčková et al. (2015) analyzed fungal species involved in decomposition of pine needle litter through culture dependent and culture independent methods. This study proved that the ratio of species recognition of culture dependent method to culture independent method is 1:2. Purahong et al. (2016) sampled leaf litter 473 times to study decomposing fungi and showed that the percentage of detection frequency of ascomycetes was 66–82%. Zhang et al. (2018) studied the ligninolytic fungal diversity in China and revealed 2621 fungal OTUs which mainly belong to Ascomycota, Basidiomycota and Zygomycota. Further, 75% are ascomycetes. Meanwhile, Seena et al. (2019) studied the ligninolytic fungal diversity in 19 globally distributed streams and the total number of fungal OTUs revealed in this study was 1311 with 79.7% being ascomycetes. Voříšková and Baldrian (2013) did a similar study and revealed that 71% of species are ascomycetes. Osono (2019) obtained 127 fungal species from 1133 leaf litter isolates and 95 are ascomycetes.
Based on the above studies, it is assumed that around 70–80% of ligninolytic fungi are ascomycetes and therefore, around 1045–1966 ligninolytic ascomycetes have been recorded in culture dependent and culture independent studies (Zhang et al. 2018; Seena et al. 2019). Further, Dashtban et al. (2010) reported that more than 14,000 fungal species produce ligninolytic enzymes and all litter degrading fungi must produce ligninolytic enzymes (Kumar and Chandra 2020). Assuming that 70–80% of above ligninolytic enzyme producing fungi are ligninolytic ascomycetes, there are around 10,500 described ligninolytic ascomycetes. If this applies to 2.2–3.8 estimate, considering 90% of them are teleomorphs (Normark et al. 2003), it is assumed that there are around 138,600–239,400 teleomorphic species of ligninolytic ascomycetes.
Case studies from endophytic ascomycetes
Endophytes are mutualists that colonize asymptomatically inside of any tissues of living plants at least in any phase in their life cycle (Singh and Dubey 2015). Bills (1996) proposed that some type of mycorrhizae such as ericoid mycorrhizae and pseudomycorrhizae can be endophytes. Endophytic colonization generally does not cause any damage to its host and does not produce any structures emerging from the external plant (Azevedo and Araújo 2007). Some endophytes can grow invitro in culture media.
Endophytes are ubiquitous and occur within a broad range of host plants, such as mosses, ferns, grasses, shrubs, deciduous and coniferous trees and lichens (Guo et al. 2008; Albrectsen et al. 2010; Mohamed et al. 2010; Su et al. 2010; Sun et al. 2011). Endophytes are an important component in natural ecosystems and they produce various bioactive chemicals, promote host growth, improve resistance to environmental stress and decompose litter (Aly et al. 2010; Saikkonen et al. 2010; Xu et al. 2010; Purahong and Hyde 2011; Tejesvi et al. 2011; Gouda et al. 2016).
Endophytic fungi have not been seriously considered in the estimation of fungal numbers (Hawksworth 1991). However, there could be more than 1 million endophytic fungal species based on ratios of vascular plants to fungal species of 1:4 (Petrini 1991). Dreyfuss and Chapela (1994) proposed that there should be 1.3 million endophytic fungal species. Most culturable plant endophytes are ascomycetes belonging to orders Amphisphaeriales, Capnodiales, Diaporthales, Hypocreales, Pleosporales, Sordariales, Trichosphaeriales and Xylariales (Guo et al. 2001; Crozier et al. 2006; He et al. 2012; Koukol et al. 2012).
Hamzah et al. (2018) revealed that the ratio of endophytic Ascomycota: Basidiomycota is around 25:1 and there are around 30,000 described, endophytic basidiomycetes species (Anke 1989; Anke and Steglich 1988). Hence, there should be 750,000 endophytic ascomycetes. Further, any vascular plant species can host somewhere 4–5 different endophytic fungal species (Sun and Guo 2012). There are 372,383 species of vascular plants and therefore, there could be 1.49 million endophytic fungal species. Hence, it is estimated that there are 675,000–1,341,000 endophytic teleomorphic ascomycetes excluding 10% obligatory anamorphs (Normark et al. 2003).
Case studies from epiphytic ascomycetes
Epiphytic fungi reside either permanently or casually on the surface of plants (Langvad 1980). They can multiply and grow on the surface of healthy leaves without any adverse effect to the host, while casual epiphytes land on the healthy leaf surface in the form of spores or mycelia but cannot grow like residents (Kharwar et al. 2010. The coexistence of epiphytic and endophytic microorganisms may play an important role for plant health and plant protection (Andrews and Harris 2000) as well as contributing to microbial biodiversity (Hawksworth and Rossman 1997).
Epiphytic fungi are dominant in Ascomycota and Basidiomycota with very few in other phyla and Sordariomycetes, Dothideomycetes and Eurotiomycetes are the most frequent among the all classes in Ascomycota (Dong et al. 2021). Among epiphytic fungi in the phyllosphere, 70–98% is ascomycetes while the rhizosphere comprises 73% of epiphytic species in Ascomycota (Oliveira et al. 2017). A comparative study of endophytic and epiphytic fungal association in leaves of Eucalyptus citriodora Hook. revealed 279 epiphytes out of 478 fungal isolates. This means number of epiphytic fungi is 1.4 times higher than endophytes. Further, Dong et al. (2021) analyzed the epiphytic and endophytic fungal communities of tomato plants and revealed 161 epiphytic fungal OTUs and 119 endophytic fungal OTUs. This suggested that the number of epiphytic fungi is around 1.4 higher than the endophytic fungi (Kharwar et al. 2010). In this study, we concluded that there should be 335,000–675,000 teleomorphic, endophytic ascomycetes and therefore, we suggest that there should be 469,000–945,000 teleomorphic epiphytic ascomycetes.
Based on the above case studies, it is estimated that there are around 1,710,000–3,405,000 teleomorphic ascomycetes in different ecological habitats and the average is around 2,558,000 species. However, all the predictions are based on the available data and some ecological groups are well-studied while others are poorly examined.
Estimated number of teleomorphic ascomycetes based on meta-DNA and culture-independent studies
The identification of some teleomorphic ascomycetes such as fungal symbionts, endophytes, marine species associated with plants and green algae, and parasites is challenging due to their unculturable nature (Blackwell 2011). However, advanced molecular techniques facilitate the discovery of undescribed species from unculturable samples (Zhang et al. 2010; Blackwell 2011). The fungal diversity estimate increases with the advent of more uncultured fungi and fungi from environmental samples. Environmental DNA (meta-DNA) can be genetic material acquired directly from environmental samples, such as soil, sediment, water and others devoid of any clear signs of biological material is an effective, safe and quick standardized sampling method (Prosser and Hedgpeth 2018). The development of advanced molecular techniques such as high-throughput sequencing has greatly contributed in identification of undescribed species (Barnes and Turner 2015). More fungal species were identified by culture-independent approaches than by culture-dependent methods (Zhang et al. 2010) and the fungal species detected by one method is really different from other method, even for the dominant fungal species (Wu et al. 2019a).
Environmental DNA is a powerful tool to explore the hidden teleomorphic ascomycetes and it challenges understanding of global biodiversity (Venter et al. 2004). It was estimated that the number of fungal species on earth ranged between 3.5 and 5.1 million when considering the species recorded from environmental samples (Blackwell 2011). The class Archaeorhizomycetes in the sub-phylum Taphrinomycotina was introduced based on only environmental DNA, even its precise ecological niches and life cycle is unknown (Rosling 2011). Further, an unknown, basal clade of phylum Ascomycota which is characterized by unicellular zoospores with a single, non-chitin or non-cellulose-walled micro-tubular flagellum was described as Cryptomycota based on meta-DNA sequences (Jones et al. 2011).
Even though there is an argument as to use environmental DNA for nomenclature, Hawksworth and Rossman (1997) proposed to use this technique to explore the fungi existing in un-examined niches as well as known habitats. Therefore, the fungal species number could be much higher than the current reliable estimates of 2.2–3.8 million.
In a study based on fungal DNA assemblages and their spatial structure in river water using environmental DNA metabarcoding targeting of ITS locus revealed 985 fungal OTUs with 97% sequence similarity (Matsuoka et al. 2019). Totally, 770 OTUs were assigned as Ascomycota and it is 78.2% of total fungal OTUs. However, when there are 150,000 described fungal species, only 92,725 are ascomycetes and it is 61.8% in total. Therefore, environmental DNA metabarcoding method provides 16.4% additional amount of species and hence number of species in Ascomycota should be 117,325. If this applies to 2.2–3.8 estimate, considering 90% of them are teleomorphs (Normark et al. 2003), it is assumed that there are 1,548,690–2,675,010 species.
A study based on high-throughput sequencing of fungus: plant ratios revealed that the number of fungal species may be around 3.5–5.1 million species (O’Brien et al. 2005). Around 62% of described fungi are ascomycetes (Species Fungorum 2021) and if this applies to 3.5–5.1 estimate (O’Brien et al. 2005), considering 90% of them are teleomorphs (Normark et al. 2003), there should be 1,947,225–2,836,620 species. Here, we estimate around 1,747,958–2,755,815 teleomorphic species based on O’Brien et al. (2005), Hawksworth and Lücking (2017) and Matsuoka et al. (2019). Wu et al. (2019a) suggested that the range of species numbers based on environmental DNA is 8.8 times higher than the traditional culture dependent methods and this gives 11–19 million species for our estimate 1.25–2.17 million species based on described species in the data bases. This is quite large value and it is significantly different from other estimates in this study.
Estimated number of teleomorphic ascomycetes based on previous estimates of Ascomycota
De Meeûs and Renaud (2002) studied the phylogenetic relationship between the parasites and the eukaryotes. This study estimated that there should be 60,000 species in Ascomycota. Further, Aptroot (2001) studied fungal diversity of Elaeocarpus sp. and estimated that there should be 40,000–70,000 species of ascomycetes. However, these estimates were done two decades ago and more species have been introduced in last two decades. About 1900 fungal species were described per year over the past two decades (Hawksworth and Lücking 2017) and there should be around 38,000 more described species. Therefore, the updated estimate in Aptroot (2001) is 0.078–0.108 million. If 90% of described ascomycetes are teleomorphic species (Normark et al. 2003), it ranges for teleomorphic ascomycetes from 0.070 to 0.097 million with 0.084 million average.
Mueller and Schmit (2007) studied several groups of ascomycetes and estimated the species number. This study was based on Rossman (1994), Hawksworth et al. (1995) and data of the Dictionary of Fungi. They estimated species number for several groups in Ascomycota including Endomycetales, Helotiales, Hypocreales, insect-associated fungi, macrolichens, non-dematiaceous hyphomycetes and coelomycetes, other perithecioid ascomycetes, Pezizales and Xylariales. There were 40,706 described species in above groups and it was predicted as 694,000 species (Mueller and Schmit 2007) with the ratio of described species: estimated species as 1:17. If this ratio applys for the average of updated estimate in Aptroot (2001) (0.084 million), there should be 1,190,000–1,649,000 teleomorphic ascomycetous species.
Updated estimation of teleomorphic species number in Ascomycota
We evaluated species number for teleomorphic ascomycetes based on five approaches; number of described species in databases, fungus:substrate ratio, ecological distribution (Table 2), meta-DNA and culture-independent methods, and previous estimates by other authors (Table 3). The average of each method was used to propose the updated value for species number of teleomorphic ascomycetes and it is 1.86 million. The species number of teleomorphic ascomycetes ranges 1.37–2.56 million and the ratio between described teleomorphic ascomycetes to predicted teleomorphic ascomycetes is 1:22. Further, there should be 3.3 million fungi when 150,000 species has been described.
The estimates based on ecological distributions and environmental DNA analysis provide large numbers. It is suggested that one species can occur in several different habitats or in different life modes and counted several times as the different species. Additionally, environmental DNA reveal the hidden diversity in habitats and most unculturable species or species rarely produce teleomorph can be trace. Therefore, the species number in these two methods are higher than others (Fig. 3).

Estimated numbers of teleomorphic species in Ascomycota based on five different approaches. The range of each approach and their mean values are marked in black dots and blue diamonds, respectively. The red dashed-line shows the average value for estimated species number of the five approaches
Limitations in estimation methods: Why estimating the species number of teleomorphic ascomycetes is challenging
The number of existing teleomorphic species in Ascomycota is not well-predicted and this estimation depends on the known number of species. So, what are the difficulties in knowing or describing teleomorphic ascomycetes? The diversity of teleomorphic ascomycetes is much higher due to their easily adaptable capability to different ecological conditions and therefore, in any given community or ecosystem, teleomorphic ascomycetes can be abundantly discovered (Berbee and Taylor 1992). A high level of reproductive plasticity and different life cycles with exciting teleomorphic and anamorphic reproduction mechanisms can be observed in ascomycetes due to their diversity (Wilson et al. 2019). These variations and diversity have led to a high level of species richness across different ecological niches.
There are three distinct phases in species introduction: an ascending phase in 1750s to 1860s, a steep phase in the 1870s to 1880s and a relatively constant phase from the 1890s to the present day based on publications (Hawksworth and Lücking 2017). However, the number of described species may be greater than this in ascending and steep phases. The internet was not previously available and most mycologists worked independently. Conferences, scientific meetings and societies were limited, and funding to attend was often unavailable. Most mycologists were unable to directly share their knowledge and experiences with others (Agerer et al. 2000). Further, scientific research was not done for commercial purposes. Most described species could not be published or published papers were destroyed during world wars. Additionally, studies in mycology and description of new fungal species decreased during the Second World War period (Hawksworth and Lücking 2017). Further, many books, notes, experimental observations and fungal specimens were destroyed and most mycologists had to move to other places or retired.
Recent intensive studies based on comprehensive inventories of ascomycetous genera and families have neglected morphology and are mostly based on molecular data (Senanayake et al. 2018). Over 90% of the collected specimens may constitute undescribed species (Hyde et al. 2018). The young mycologists and students are willing to describe new collections as novel taxa rather than assign them to existing species (Hawksworth and Lücking 2017). Further, it is required to combine the data on biogeographic distributions, levels of endemism and host specificity into the described species list when estimating the number of teleomorphic ascomycetes (Mueller and Schmit 2007). However, the number of described species has increased due to application of molecular techniques for species delimitation. There are up and downs in the number of described species after 2010. This may reflect the resolve of cryptic species, synonymize and link teleomorphs and anamorphs, rather than introducing new collections as new species.
The grid map-based method for predicting species richness introduced in Lücking et al. (2014) has been used to predict species richness in the lichenized family Graphidaceae in Ascomycota (Aptroot and Cáceres 2016; Cáceres et al. 2017; Mendonça et al. 2020). This method uses known occurrence records to provide a prediction and a precise estimate of species richness. Thus, to predict the species richness of a large group of teleomorphic species in Ascomycota, the accuracy of the used records is important. Errors during species introduction, incorrect nomenclature, misidentification of cryptic species or species complexes, fungi from understudied fungal habitats and hosts can be problematic. The correct estimates of teleomorphic species richness in Ascomycota can be a difficult task (Hyde et al. 2020b). There are several limitations of the species estimates as below.
One name for one fungus (1N1F) for pleomorphic species
The “One name = One fungus” system used nowadays has been effective in establishing standards for naming fungi in the scientific community. Before the 1N1F system came into effect, teleomorphs and anamorphs were given separate names depending on the circumstances from which they were discovered. Some ascomycetes produce an anamorph in their life cycle (Seifert and Samuels 2000) and the anamorph sometimes becomes the prominent, commonly available morph in nature (Li et al. 2020).
With the use of DNA sequence data for species identification, the accuracy in identification and linking teleomorphs and anamorphs of a species are important. Limitations in available DNA sequence data could however lead to erroneous identification. Recent studies have been resolved errors made when selecting one name for pleomorphic fungal genera (Hawksworth 2012, 2015; Réblová et al. 2016; May 2017; Taylor et al. 2016). There are plenty of teleomorphic species epithets in the species catalogs without linking to anamorphic species due to a lack of molecular data from ex-type and other authentic cultures or poor morphological descriptions (Seifert and Samuels 2000; Hawksworth et al. 2013). Most of the early introduced species do not have molecular data and species introduction was based on morphology (Vellinga et al. 2015; Koukol and Delgado 2021). Further, cultural studies not performed often and morphology of anamorph or teleomorph derived from pure cultures was not recorded. Therefore, recollection of earlier species or use fungarium materials to obtain DNA is required (Seifert and Rossman 2010; Aime et al. 2021).
Even after the implementation of 1N1F species system, certain research areas still predominantly use the older names in studies where the main focus is not related to taxonomy and nomenclature. The teleomorph of fungi are rarely encountered in plant pathology, thus plant pathologists tend to name the pathogenic species related to their anamorph, but the link between the two states was rarely established (Wingfield et al. 2012). These types of basic errors in species naming would hinder the possibility of accurate species estimates. Article F.8 for pleomorphic fungi in Shenzhen code states names proposed simultaneously for separate morphs (anamorph and teleomorph) of a taxon of non-lichen-forming Ascomycota and Basidiomycota are necessarily heterotypic and are not therefore alternative names (Turland et al. 2018). This code facilitates both teleomorphic and anamorphic names in the legitimate state and those legitimate names are treated equally when establishing priority to conserve the accepted name regardless of the life-history or stage of the type.
Rossman et al. (2015a, b) and Réblová et al. (2016) have provided recommendations for conservation or use of pleomorphic generic names in Dothideomycetes and Sordariomycetes. However, linking teleomorph and anamorph of species is challenging. Pure cultures obtained from single germinating ascospores often sporulate and anamorphs are formed (Senanayake et al. 2020a). Sometime, strains of pleomorphic species obtained from different specimens cluster together with strong support in phylogenetic trees (Karunarathna et al. 2017; Wanasinghe et al. 2018). The colony characters and nucleotide identity of molecular sequences should be checked even if cultures do not sporulate. Further, some unculturable ascomycetes that are only known from their teleomorph cannot be linked to the anamorph.
However, some other classes such as Orbiliomycetes, the generic names are much more complicated because of the names based on single morphological differences without molecular data (Baral et al. 2018). In Orbiliomycetes, a narrow concept has used for the demarcating the generic boundaries of the anamorphs, while a broad concept relies on the teleomorphs. Therefore, more generic names have been established for the anamorphs. Hence, it should be avoided to adopt a certain generic concept prematurely, as this may imply a lot of unnecessary name changes (Baral et al. 2018). Therefore, linking teleomorph and anamorph of species is an important practice in nomenclature and it can affect species number.
Phenotypic plasticity
Phenotypic plasticity in fungi denotes that changes in morphology, behavior and physiology in response to the environmental variation (Price et al. 2003). Phenotypic plasticity allows teleomorphic ascomycetes to respond to climatic changes within their lifetime (Williams et al. 2008). This is important for species to survive as evolutionary responses for climatic changes by natural selection takes time to make any adaptation.
More than 40% of ascomycetes live in symbiosis as lichens (Kirk et al. 2008). Lichens show high phenotypic plasticity together with geographical distributions (Divakar et al. 2013; Muggia et al. 2014). Parmeliaceae is a hyper- lichenized fungal family mainly distributed in the tropics (Kraichak et al. 2015). The type genus Parmelia includes several distinct species by phenotypic plasticity (Valladares et al. 2007). Parmelia discordans and P. omphalodes were described based on morphological differences. However, molecular data showed that these two species are conspecific and phenotypic variations are made according to environmental changes (Divakar and Upreti 2005). Nipponoparmelia pseudolaevior and N. laevior show phenotypic plasticity in this family (Molina-Montenegro et al. 2016).
Phenotypic stasis
Phenotypic stasis is explained by natural selection and genetic drift, or by constraints imposed by mutation and recombination of standing genetic variation (Mallard et al. 2019). This is a basic method in speciation and genetic variations (Chethana et al. 2020). Gene variations formed by phenotypic stasis can completely disappear to reduce the genetic variation or initially rare alleles become much more frequent to dominant the gene variations (Mallard et al. 2019). However, the morphological variations formed by phenotypic stasis are retained in a population if only individuals survive and reproduce. Phenotypic stasis forms species morphologically similar, but genetically different. Therefore, species estimates must include these species.
Homoplasy
Homoplasy is a trait that has been gained or lost independently in separate lineages with evolution (Torres-Montúfar et al. 2018) and it can arise by selection pressures or genetic drift (Stearns and Hoekstra 2005; Hall and Colegrave 2008). Homoplasy mostly appears in similarity of morphological characters, but also in molecular sequences (Reece et al. 2011), life cycle (Silberfeld et al. 2010) and behavior (de Queiroz and Wimberger 1993).
Jiang et al. (2020) showed in a phylogenetic study of foliicolous lichens that a new lineage sister with Strigulaceae (Dothideomycetes) was formed, however morphologically similar to Porina (Lecanoromycetes). This new clade represents a monogeneric family Tenuitholiascaceae which is typified by Tenuitholiascus with a single species T. porinoides. This species is morphologically similar to the genus Porina in external morphology, ascospore type, the thin-walled asci and unbranched paraphyses. Further, Schmitt (2011) showed that homoplasy affects the evolution of fruiting body type and ascus at the class level within the phylum Ascomycota. This may increase the species number described if only based on phenotypic characters.
Synonyms and conspecific species
Synonym is a scientific name currently applies to a taxon that goes by a different scientific nameand synonyms form strong, monophyletic clades with currently applied taxon in phylogenetic trees. Therefore, these synonyms are known as conspecific species (Rossman et al. 2015a). Hence, there is often more than one scientific names for a single species and the morphs had been described in different genera (McNeill et al. 2006). Two or more names for different morphs of the same species are not accepted according to the Melbourne Code (McNeill et al. 2012). Hence, Wijayawardene et al. (2012), Rossman et al. (2015a,b), Réblová et al. (2016) have proposed recommendations to determine which name to conserve. Proposals were based on excluding synonymy, giving priority to basionyms, commonly used names or the commonly occurring morph in nature.
Conspecific species being identified as distinct species through morphological data, but with molecular data providing evidence for them being identical, has also led to incorrect species identification. Some sexually compatible conspecific fungal species can also produce new pathogens via interspecific hybridization and reproductive interference (Giordano et al. 2019). The genus Diatrype is typified by D. disciformis (Fries 1849). Libertella betulina, the type species of Libertella, is the anamorph of Diatrype stigma (Grove 1937; Kutorga et al. 2006), while L. disciformis is the anamorph of D. disciformis. Diatrype disciformis and D. stigma are conspecific (Trouillas et al. 2010). Further, Libertella is the older name as it was erected in 1830 while Diatrype was only erected in 1842, thus Diatrype and Libertella are synonyms. However, Diatrype has a great number of species including important plant pathogens. Hence, Diatrype was recommended for protection over Libertella (Réblová et al. 2016).
There are many recommendations proposed for taxa of Xylariaceae (Réblová et al. 2016). The genus Daldinia is typified by D. concentrica (Stadler et al. 2014). The monotypic genera Annellosporium which is typified by A. nemorosa and Versiomyces typified by V. cahuchucosus Whalley & Watling, have been synonymized under Daldinia as D. nemorosa based on the phylogeny and D. cahuchucosa based on morphology and chemotaxonomic evidences (Stadler et al. 2014). Daldinia is common with many species and has been recommended for use. Therefore, recognizing excluded synonyms is essential to estimate the actual number of fungi.
Illegitimate and invalid names
Published taxonomic names may be illegitimate and invalid. This means the species exist, but are nomenclaturally incorrect due to contravening some of the articles laid down by the nomenclature codes. If a published species name is not accepted as a proven valid species, then it can be superfluous as a synonym of a known species, non-compliant with nomenclature codes thus considered a “bad” name and doubtful name with insufficient study (Wang et al. 2019). The number of accepted names, synonyms, invalid or illegitimate names, and unstudied names has been compared by Wang et al. (2019). They found that accepted names increased markedly over time and increased significantly after the 1900s. The number of synonyms, invalid or illegitimate names increased slowly and it is evident that the quality of fungal taxonomic work has improved with the application of molecular techniques. The International Code of Nomenclature for Algae, Fungi, and Plants is updated every four years and proposes and regulates all the articles related to nomenclature (Turland et al. 2018).
Introgression and natural hybridization
Introgression means transfer and incorporates alleles from one species into the gene pool of another species by hybridization and backcrossing (Schardl and Craven 2003; Stukenbrock 2013, 2016; Restrepo et al. 2014). There are many occasions when genetic information can be transfered between closely related species and thus gene flow between cryptic species has frequently been found (Hawksworth 2001; Bickford et al. 2007; Hawksworth and Lücking 2017). Therefore, species boundaries in morphologically indistinct species and species complexes may be doubtful (Barton and Hewitt 1985; Barton and Gale 1993). If a portion of the introgressed gene pool of each of the hybridizing taxa remains constant and uncontaminated then different distinct gene pools can be recognized as new species.
Beneficial alleles tend to introgress easily for habitat adaptation or reproduction (Barton 1979). Thus, patterns of differential introgression across hybrid zones in genes or genome regions are important for habitat adaptation and speciation (Payseur 2010; Shaw and Mullen 2011; Nachman and Payseur 2012). Sometimes, the gene flow between species is limited or prevented in nature by a set of basic barriers. These limits control transfer of the genetic material which affects phenotypic variations between species and determines if species reproduce individually (Bouck et al. 2005; Lemmon et al. 2007; Roe and Sperling 2007; Wagner et al. 2013). However, the question is how many species evolves presently by introgression and hybridization? The answer is unpredictable. Therefore, this should be considered when estimate the teleomorphic species in Ascomycota.
“Man-made” or domesticated species
Adaptive hybridization is used to obtain industrially important species additionally to natural hybridization (Burgarella et al. 2019). Genetic materials are changed during domestication of wild species and new species or varities may form (Shibata et al. 2015). Industrial cultivation of some teleomorphic ascomycetes such as Cordyceps, morels, truffles requires hybrid varities to obtain high yield. However, the obtained number of hybrid species in a particular period is undetermined. Additionally, the behavior of hybrid species with the wild species gene pool is not well studied. Therefore, an idea of the number of domesticated species is needed to estimate the number of species in Ascomycota.
Polyphyletic genera and species
The polyphyletic nature of fungal genera derived from more than one common evolutionary ancestor or ancestral group cannot taxonomically be in the same genus. Sometime morphologically similar species cluster in different sub-clades in phylogenetic trees representing several distinct genera (Phookamsak et al. 2015, 2017; Konta et al. 2020). This affects the number of described species. Some studies have used slightly different morphological characters of taxa along with the phylogenetic analyses to introduce new genera (Hyde et al. 2020a).
Xylariaceae comprises several polyphyletic genera which are phylogenetically distantly related to each other (Peršoh et al. 2009; Senanayake et al. 2015). Xylaria is typified by X. hypoxylon (Schrank 1789; Greville 1824) and species in Xylaria are saprobes or endophytes (Thomas et al. 2016). Lumping and splitting of Xylaria species in phylogenetic trees forming unresolved lineages has occurred over time. Most phylogenetic analyses have shown that Xylaria species do not form a monophyletic clade and are scattered within Xylariaceae (Hsieh et al. 2010; Senanayake et al. 2015; Maharachchikumbura et al. 2016; Wendt et al. 2018). Few Xylaria species are clustered with Amphirosellinia, Astrocystis, and Collodiscula without strong statistical support (Wendt et al. 2018; Konta et al. 2020).
Generic polyphyly does not markedly change the number of existing species, but species polyphyly changes the number of species. The “special status” of a species comprises the unique, observable morphological characters (Queiroz and Donoghue 1988). It is implicitly assumed that species are monophyletic or at least paraphyletic. However, hybrid speciation arguably leads to polyphyletic species (Hörandl and Stuessy 2010). Hybrid species are a common phenomenon that allows for rapid speciation (Linder and Risenberg 2004) and polyphyletic species develop into different species later.
Extinct or endangered species
The evolution of fungi begun around 1.5 billion years ago (Wang et al. 1999; Brundrett 2002). There is evidence that fungal communities in Ascomycota were present in the Devonian period, 416–359 million years ago (Strullu-Derrien et al. 2018). Ascomycetes diversified rapidly in terrestrial environments and therefore, they occupied numerous ecological niches. However, teleomorphic ascomycetes are recently threatened by habitat loss, loss of symbiotic hosts, pollution, over exploitation of the animal and plants, destruction of ecosystems and climate change and they are also becoming extinct (Wang et al. 1999). However, the vast majority of teleomorphic ascomycetes have not been assessed. The IUCN has listed 280 threaten fungal species under several catogaries, such as critically endangered (CR, 15 species), endangered (EN, 59 species), vulnerable (VU, 90 species), near threatened (NT, 40 species), least concern (LC, 54 species) and data deficient (DD, 22 species). The IUCN Red List contains 46 threaten teleomorphic ascomycetes (Table 4) (IUCN 2021). The objective of the global IUCN red list of threatened fungal species is to determine conservation issues to the public and policy makers and help the international community decrease species decline and extinction (Lughadha et al. 2020). The IUCN Red List is the most comprehensive, objective global approach for evaluating the conservation status of fungal species. The largest number of threatened species is from Europe (IUCN 2021). IUCN organized three workshops in Chile, and the UK in 2020 and this effort will assess the conservation status of endemic species of three regions, South America, Europe and Southeast Asia.
However, the number of threaten teleomorphic ascomycetes is very low compared with the described number of teleomorphic species in Ascomycota due to the difficulties in screening fungal populations. Most teleomorphic ascomycetes are only visible when they produce fruiting bodies and may not be found in the same place every year (Senanayake et al. 2020a). Some early evolved teleomorphic ascomycetes have become extinct and there is no documentation for them.
Biodiversity hotspots
Only about 93,000 Ascomycota species have been introduced and documented even more are estimated (Roskov et al. 2019). Studying the fungi in biodiversity hotspots is important to determine the undescribed taxa (Hawksworth and Lücking 2017). Biodiversity hot spots and geographic and ecological habitats which are poorly or under-studied are major localities for these undescribed species. Biodiversity hot spots are designated by IUCN and they are conserved by government in located country (Marchese 2015). Biodiversity hotspots occupy approximately 1.4% of earth’s land area, but 60% of Earth’s biodiversity is gathered there (Possingham and Wilson 2005; Marchese 2015).
The global biodiversity hotspots are referred to as areas featuring exceptional concentrations of endemic species and experiencing exceptional loss of habitat (Myers et al. 2000). Most ascomycetous type collections were obtained from North America, Europe (France, Germany, Italy, Spain, Sweden, UK), Japan, India, China, Thailand, Philippines, Australia and Brazil (Species Fungorum 2021). Most of these countries are located in biodiversity hotspots, but the whole country has not been preserved as biodiversity hotspots (Matutea and Sepúlvedab 2019). Although many species have been collected from countries which are located in biodiversity hotspots, it does not mean that fungal diversity is restricted to the biodiversity hotspots (Marchese 2015). Generally, rules and regulations for entrance and utilization of reso urces are strictly controlled in these areas, even for scientific studies. This is one reason that mycologists could not estimate exact number of teleomorphic ascomycetes in hotspots. However, it is predicted that biodiversity hot spots have extremely favorable conditions for ascomycetes and thus production of sexual, thick-walled spores is not prominent. Therefore, species in Ascomycota may occur in vegetative phase or reproduce asexually.
Hidden species in niches
Morphologically, teleomorphs of ascomycetous fungi are only recognized when they produce sexual reproductive organs. Some ascomycetes occur in niches without forming any visible, distinct fruiting structures and are only obtained as hyphae (Schardl and Craven 2003). These species are ecologically cryptic and difficult to screen by traditional phenotypic approaches. Environmental DNA analyses can reveal such species (Wu et al. 2019a). Additionally, fungal succession may help to obtain those species separate. This has been defined as the sequential occupation of different ascomycetes or different associations of ascomycetes on the same substrate or site (Challacombe et al. 2019). This happens because of a sequence of sporulating fungi on a substrate by mycelium. However, replacement of one ascomycetous species by another is not necessary and some ascomycetes sporulate together on a substrate (Hyde and Jones 2002). Sometimes, incubation of fresh specimens is necessary to obtain maximum fungal diversity, especially rare or slow growing species.
Poorly-studied fields
Many terrestrial teleomorphic ascomycetes have been described, but fewer are known from aquatic habitats. Natural groundwater limestone aquifers are challenging and unexplored fungal habitats (Krauss et al. 2003; Lategan et al. 2012; Risse-Buhl et al. 2013). Mangrove and wetland associated teleomorphic ascomycetes are poorly known as compared to terrestrial taxa (Lee et al. 2019). Therefore, it is necessary to fully screen marine and aquatic habitats to explore the fungal diversity. Research on aquatic fungi in recent years have incorporated molecular techniques to achieve a better identification of taxa and many new species have been introduced (Zhang et al. 2017; Hyde et al. 2020a; Wei et al. 2020). Most of the earlier studies are based on observation of morphological characters and culture-dependent techniques (Luo et al. 2019). There are unculturable freshwater ascomycetes and hence, culture-independent techniques need to be followed to understand the diversity among teleomorphic, freshwater ascomycetes (Hyde et al. 2020b). Clinical mycology is another field which has not been studied much with few human and animal pathogens reported (Homei 2006; Pihet et al. 2009; Köhler et al. 2015; Sullivan et al. 2015).
Less-studied sites in developing countries
Biodiversity hotspots in Asia (except Japan) and South America are generally poorly studied and not well-known (Hyde et al. 2020c). Most countries in these two continents are economically impoverished developing countries and lack resources and enough funding for fungal research (Subramanian 1986; Jones et al. 2007). Most developing countries are located in tropical and subtropical regions and fungi thrive in these countries due to the favorable habitat conditions. However, some species rich, biodiversity hotspots in these countries lack studies on species diversity and richness (Marchese 2015). In fact, the whole planet is not being evenly screened and therefore, many important teleomorphic ascomycetes may become extinct prior to description and documentation.
In 2017, 35% of the world’s described fungal species were from Asia (Willis 2018). This may have increased within the last two years (Hyde et al. 2020c). Africa is very different from South America or Asia in biodiversity having dry climates and different geographical lands, while there are many rainforests and wet ecological sites in South America. Therefore, less-studied areas should be examined well to explore undescribed teleomorphic species in Ascomycota.
Unexamined collections in specimen repositories and missing specimen records
Traditionally, teleomorphic ascomycetes have been distinguished by different approaches and concepts such as morphology, physiology, biochemistry or reactions to chemical tests. Morphology was commonly used to introduce teleomorphic ascomycetes and it was improved with the innovation of the microscope. However, there are many poorly identified or superficially examined collections in fungaria (Senanayake et al. 2020a). Many reference specimens in fungaria and living cultures in fungal culture collections are not named to the species level, and in some cases not even to the generic level (Hawksworth and Lücking 2017).
Additionally, some collections and isolates are connected to existing species names without comprehensive specimen examination and sometimes it is possible to find different fungi in the same specimen if those specimens are examined carefully. It has been suggested that morethan 20,000 teleomorphic ascomycetes have been collected worldwide but not described yet (Hawksworth and Rossman 1997). Mycologists often have a large collection of material waiting for formal introduction and description. Some taxonomic records, descriptions and specimens have been lost in various situations before description (Senanayake et al. 2016). Therefore, re-visiting early-introduced ascomycetes which are only based on morphology is important (Ariyawansa et al. 2014).
Some monographic work and some journal issues that focus on exploration of global fungal biodiversity such as Mycosphere Notes (ZUAE, Guangzhou, China), Fungal Diversity Notes (Springer Nature, Switzerland) or Fungal planet description sheets (CBS, The Netherlands) describe hundreds of novel teleomorphic ascomycetes annually. However, some species are introduced only based on morphology and Mega-blast similarities in GenBank without phylogenetic analysis. Most of the new collections are described without re-visiting morphologically similar earlier species or genera. If a collection is from known host, then it is necessary to check previous species records on the host and locality (country or continent) prior to describing a new species (Senanayake et al. 2020b). Careless practices will result in proliferation of synonyms or invalid names leading to confusion and instability in nomenclature. Many described teleomorphic ascomycetes in the pre-molecular era are only based on morphology and there are older collections in repositories that need to be examined and described.
Incomplete descriptions of earlier described species
In earlier studies, specimens were examined by light microscope and 40 was the maximum magnification (Senanayake et al. 2020a). Lenses of light microscopes were calibrated manually (Zhang et al. 2016). One unit of the ocular micrometer disc is needed to calibrate against a known length and measurements were subjective from person to person. Newer technologies facilitate measurements directly when photographing and software is already calibrated. Hence, errors in measurements have been less in recent descriptions. Earlier prologues based on the limited characters and some important, inconspicuous characters were not examined or not mentioned (Senanayake et al. 2018). Therefore, new collections of existing species are mistakenly described as new species. A prologue should include information of examined materials (Seifert and Rossman 2010). However, some specimens lack these details or are incomplete. Earlier prologues usually do not contain culture characters. Therefore, incomplete descriptions of earlier introduced species make complications in species identification.
Synanamorphism
Synanamorphism is a teleomorphic species produces several morphologically distinct anamorphs and these synanamorphs have been often reported from cultural studies (Crous et al. 2009; Réblová et al. 2021). Before molecular data was available for taxonomic studies, species delimitation was based on the morphology of specimens and cultures. Pure cultures were used to obtain teleomorphs or anamorphs and then to describe the holomorph. However, some teleomorphic species produce several anamorphs and those anamorphs are described as different species. Molecular phylogenies of some studies proved that some teleomorphic species produce two or more anamorphs (Fan et al. 2018; Réblová and Štěpánek 2018). Therefore, it is assumed that some synanamorphic species might have been considered as separate species dueto their distinct morphologies and they should avoid from the unmber of known species.
Species in Cytosporina Sacc., Libertella Desm., and Naemospora Sacc. have been reported as anamorphs of diatrypaceous species (Glawe and Rogers 1984). However, the morphology of the anamorph is not useful when differentiating taxa in Diatrypaceae either at the genus or species level. There is indistinguishable morphology and many species produce different types of anamorphs in the same culture (Rappaz 1987). Additionally, different types of conidiogenesis have been reported in the same Diatrype strain (Glawe and Rogers 1982). Eutypella parasitica produces both pycnidia and acervuli on both natural substrata and cultures (Glawe 1983). Further, the anamorph of Hapalocystis (Sydowiellaceae, Diaporthales) is reported as Stilbospora, Hendersonia or Dothiorella (Wehmeyer 1941; Castlebury et al. 2002), or as stilbospora-like taxa (Barr 1978). A phoma-like anamorph is reported for H. berkeleyi in culture (Glawe 1985; Liu et al. 2015; Senanayake et al. 2016).
Absence of molecular data for described species
Before the 1990s, fungal species were introduced based on morphology and there are no molecular data for many of those described species. Classification of those species is challenging without molecular data (Huang et al. 2021). Cryptic species, phenotypic plasticity, phenotypic stasis, pleomorphism, and homoplasy between teleomorphic ascomycetes are difficult to reveal without molecular data (Maharachchikumbura et al. 2014). Hence, the estimated number of teleomorphic species in Ascomycota should include those hidden species.
Addition to the phylogenetic relationships, molecular data reveals the character evolution of teleomorphic species in Ascomycota. Character evolution deals with the process of evolution of a trait along the branches over a period of time from a common ancestry (Hongsanan et al. 2018). This explains the history of life, the relationships among extant species and character states for each species (Vijaykrishna et al. 2006). Most character evolution studies were carried out after the 1990s (Liu and Hall 2004; Li et al. 2005; Schoch et al. 2009; Schmitt 2011; Kumar et al. 2012) and these studies have not been obviously conducted on teleomorphic ascomycetes which lack sequence data. The unavailability of complete sets of sequence data is the major issue for absence of character evolution studies. These studies concur or argue against species delimitation. Hence, these studies influence the estimate number of teleomorphic species in Ascomycota.
Apothecia are the primitive fruiting body type of Pezizomycotina. However, the formation of perithecia and cleistothecia is still unclear (Hongsanan et al. 2018). The amyloid reaction of stromatic tissues occurs in some species in Sordariomycetes such as Hypocreaceae in Hypocreomycetidae, Xylariaceae in Xylariomycetidae and Cryponectriaceae in Diaporthomycetidae. However, the reason for the appearance and disappearance of the same character during evolution and when, how and why these characters evolved are unclear. If there is a complete set of molecular data, character evolution of teleomorphic species in Ascomycota can be predicted clearly and it provides additional taxonomic value for a species.
Molecular data unavailability for both teleomorph and anamorph of ascomycetes
Phenotypic species were previously introduced based on morphological characters and later, sequence data obtained for some of earlier described species. However, many species lack sequence data. If a culture of a teleomorphic Ascomycota sporulates then, the anamorph can be obtained and identified. These teleomorphs and anamorphs can be linked if they are derived from a pure culture. If a specimen contains both ascomata and conidiomata in close to each other, then cultures obtained from both ascospores and conidia should be further analysed for molecular and colony morphology. If the colony morphology in the same media and at the same maturity is similar and sequence data are identical, then the conidiomata are anamorph of teleomorphic fungi and it should be named as one species. However, colony characters and sequence similarity can be used if teleomorphs and anamorphs are described from different specimens. The sequence quality should be high and both forward and reverse directions are needed.
Senanayake et al. (2015) introduced the genus Ciferriascosea Senan., Bhat, Camporesi & K.D. Hyde with two teleomorphic species C. fluctuatimura Senan., Bhat, Camporesi & K.D. Hyde and C. rectimura Senan., Bhat, Camporesi & K.D. Hyde and they were accommodated in family Phlogicylindriaceae (Xylariales). The anamorph was not reported for this genus. Phlogicylindriaceae is typified by anamorphic genus Phlogicylindrium Crous, Summerb. & Summerell and teleomorph is not reported for this genus. Therefore, species in Ciferriascosea and Phlogicylindrium cannot be morphologically compared. Phylogenetically, species in these two genera clustered together with high bootstrap support. However, ITS loci of the type species of Ciferriascosea, C. rectimura showed 10.4% base pair differences from ITS loci of the type species of Phlogicylindrium, P. eucalypti (Jeewon and Hyde 2016). Further, 85 base pairs are missing in the ITS loci of Phlogicylindrium eucalypti. Additionally, Ciferriascosea rectimura was isolated from twigs of Spartium junceum L., while Phlogicylindrium eucalypti was collected from leaves of Eucalyptus globulus Labill. Therefore, Senanayake et al. (2015) proposed a new genus Ciferriascosea for this teleomorphic taxon instead of proposing it as the teleomorph of Phlogicylindrium.
Camarosporidiella caraganicola was introduced based on a strain derived from the anamorph that was described by Liu et al. (2015). Wanasinghe et al. (2017) has examined several specimens of the teleomorph of Camarosporidiella caraganicola. Phylogenetically, these strains clustered together with strong support. Both teleomorph and anamorph were collected from the same host (Caragana frutex (L.) K.Koch) in the Rostov Region, Russia. Therefore, Wanasinghe et al. (2017) linked these teleomorph and anamorph together as the holomorph of Camarosporidiella caraganicola by considering the host similarity and statistical support in phylogeny.
A teleomorph of an astragalicola-like taxon was collected and obtained in culture (MFLUCC 17˗0832). The multi-gene sequence analyses showed that this isolate clusters with Astragalicola amorpha with strong support (Wanasinghe et al. 2018). However, A. amorpha is known from its anamorph and thus, it is not possible to compare their morphologies. A comparison of the ITS loci of these two strains revealed 2.12% nucleotide differences, which justifies these two isolates as two distinct taxa (Jeewon and Hyde 2016). Therefore, the new collection was introduced as Astragalicola vasilyevae.
The anamorph of Wojnowiciella dactylidis was illustrated in Liu et al. (2015) and molecular data was provided. Karunarathna et al. (2017) illustrated the teleomorph for this species. Both teleomorph and anamorph were collected from Italy on different host plants but colony characters of both strains are similar. Phylogenetically, both strains clustered together with strong support. The ITS sequences obtained from the teleomorphic strain was identical to anamorphic strain with 100% base pair similarity. Therefore, Karunarathna et al. (2017) linked these two strains as the holomorph of Wojnowiciella dactylidis.
Teleomorphic ascomycetes described based on only Internal Transcribed Spacer (ITS) region
The nuclear ribosomal internal transcribed spacer (ITS) region is located between the sequences encoding the small (SSU) and large (LSU) subunits of the ribosomal operon (White et al. 1990). The ITS region is the formal primary fungal barcode with the highest probability of correct species identification of a broad group of fungi (Horton and Bruns 2001; Bridge et al. 2005; Martin and Rygiewicz 2005; Seifert 2009; Bellemain et al. 2010; Schoch et al. 2012). More than 100,000 ITS sequences of teleomorphic ascomycetes are deposited in international nucleotide sequence databases (Baturo-Cieśniewska et al. 2020). Even though, many sequences of correctly identified species are deposited in databases, there are some sequences with technical errors, incorrectly named species, atypical chimeric ITS sequences, and sequences verified only at the generic level or above which makes reliable problems in species identification (Nilsson et al. 2009, 2014; Hongsanan et al. 2018). The ITS locus in species of some genera has minimal molecular variation (Andrew et al. 2009) and cannot resolve species (de Hoog and Horré 2002).
Further, the ITS region has additions and deletions in some groups of fungi and is not equally variable in all taxa (Nilsson et al. 2008). Most databases are improving sequence availability. The lack of sufficient ITS sequences especially in species-rich, morphologically indistinct genera, can therefore be problematic (Seifert 2009). A high-fidelity, universal primer pairs for amplification of the EEF1A1 gene has been developed as secondary DNA barcodes for the fungi (Stielow et al. 2015). However, the availability of deposited reference sequences of EEF1A1 gene in databases is poor (Meyer et al. 2019). Further, mycologists who have deposited sequences in the NCBI database determine the species name by themselves. The introduction of incorrectly identified sequences creates in errors because users adopt incorrect species names (Ko et al. 2011; Nilsson et al. 2012, 2019). In most species, the nucleotide composition of the ITS sequences in the single species is identical or slightly varied. The ITS locus has certain interspecific variations (Nilsson et al. 2008; Talgø et al. 2010). These interspecific variations in ITS sequences may be higher than 1% due to technical errors, may be evidence to be a new species (Jeewon and Hyde 2016).
As an example, 245 ITS sequences of Diaporthe eres obtained from GenBank and those sequences were aligned using default settings of MAFFT v.7 with Diaporthella corylina (CBS 121124) as the out group. Maximum likelihood analysis was performed after excluding 105 identical sequences and then resulted in 14 different clades (Fig. 4) representing 14 types of sequence patterns. The ex-epitype strain of Diaporthe eres (AR5193) clustered in clade A, while clades B-E showed around 1% base-pair difference with sequence pattern of clade A (A-B:1.1%, A-C: 1.3%, A-D:1.1% A-E:1.3%). However, clades F-N showed more than around 1% base pair variation than clades A (A-F:4.0%, A-G:3.7%, A-H:3.9%, A-I:4.4%, A-J:4.4%, A-K:4.6%, A-L:5.2%, A-M:5.0%, A-N:5.4%). According to the guidence of Jeewon and Hyde (2016), 1% position ambiguities in ITS with 450 base pairs can be a new species. In our analysis, B-E types of sequences show fewer base-pair differences than other patterns of sequence types showing certain interspecific variations (Nilsson et al. 2008; Talgø et al. 2010). However, clades F-N show more base-pair differences than the epitype strain due to technical errors of sequencing or misidentification of fungi. Therefore, newly obtained strains should always be compared with the sequences of ex-type strains and ex-type strains should be included in phylogenetic trees.


Phylogram generated from maximum likelihood analysis based on ITS sequence data of Diaporthe eres. The tree is rooted with Diaporthella corylina (CBS 121124) and ex-epitype strain of D. eres is in red bold. Subclades which are representing the different type of sequences are labeled A to N
Further, ITS based species identification may not be suitable when interspecies nucleotide variations are less (Baturo-Cieśniewska et al. 2020). Sclerotinia sclerotiorum and S. trifoliorum are causative agents of clover rot disease and the symptoms are identical (Vleugels et al. 2012). ITS sequences of Sclerotinia species are almost identical (Freeman et al. 2002). Sclerotinia sclerotiorum differs from S. trifoliorum by one nucleotide, guanine or thymine in position 120 (Njambere et al. 2008; Baturo-Cieśniewska et al. 2017). Therefore, ITS based species identification is not suitable for this situation. Interspecific and intraspecific variation of ITS sequences is often low for some species. Phacidium fennicum differs from P. lacerum by two base pairs while others share identical ITS sequences but are morphologically distinct (Crous et al. 2014; Tanney and Seifert 2018). Therefore, use of ITS sequences together with morphology and at least a protein coding gene should be emphasized (Pryor and Michailides 2002; Schubert et al. 2007; Samson et al. 2014; Robbertse et al. 2017; Bensch et al. 2018).
Next generation sequencing (NGS) and meta-genomic DNA
NGS or high-throughput sequencing methods are the modern sequencing technologies (van der Heijden et al. 2008). These approaches have several advantages in that an initial knowledge of the genome or genomic features are not required, single-nucleotide resolution facilitates detection of related genes, alternatively spliced transcripts, allelic gene variants and requires less DNA/RNA as input and has higher reproducibility (Peršoh 2015). Diversity and distribution patterns of teleomorphic ascomycetes in ecosystems give important evidence on species number, ecosystem functions and stability (Schmit and Lodge 2005). Microscopic observations and culture-dependent approaches cannot efficiently calculate the number of teleomorphic species of Ascomycota in environmental samples such as soil, water, and air as the majority of them is generally complex, unculturable and not visible without a microscope (Mitchell and Zuccaro 2006; Stewart 2012; Hongsanan et al. 2018).
Authentic physical specimens do not exist for these species and they are therefore known as “dark taxa” (Tedersoo and Smith 2017; Ryberg and Nilsson 2018). There are no given any formal names to genus and species level, and hence types are not designated (Taberlet et al. 2012; Herder et al. 2014; Hawksworth et al. 2016). Article 40 (Valid publication of names; section 2: Names of new taxa) in Shenzhen Code states that a new taxon at the rank of genus or below published on or after 1st January 1958 is valid only when the type specimen of the name is indicated (Turland et al. 2018). Therefore, high-throughput sequencing approaches are needed to reasonably characterize and estimate those fungal communities.
However, there are some limitations in meta-DNA sequencing and many mycologists disagree with validating names. Concerns of the mycological community against the premature introduction of DNA-only based nomenclature have been presented (Hongsanan et al. 2018; Thines et al. 2018; Zamora et al. 2018). However, DNA meta-barcoding has the potential to provide a much better understanding the species diversity and richness in fungal communities (Heeger et al. 2018) and meta-genomic DNA has provided clues as to where and which teleomorphic ascomycetes can be found and this is applicable in estimation of the species number, but it cannot be used for classification.
Environmental samples are poorly linked to species-based databases (Hibbett et al. 2011). Therefore, errors in communication have occurred from one publication to another when using the system databases (Hibbett et al. 2011; Ryberg and Nilsson 2018). Thus, metagenomics DNA-based nomenclature can be accepted at least for species number estimations and fungal diversity studies (Hawksworth et al. 2016; Ryberg and Nilsson 2018). There is uncertainty when naming a species, if the morphology is indistinct or DNA sequences fail to resolve species relationships. Eventhough the species lack morphology, genomic and ecological significance supported this introduction. Khan et al. (2020) introduced two new species, Archaeorhizomyces victor and A. secundus (Archaeorhizomycetes, Taphrinomycotina, Ascomycota) based on the distinct base pairs comparison of the internal transcribed spacer region ITS1 and ITS2 with similar taxa obtained from environmental DNA.
Buèe et al. (2009) found 1000 operational taxonomic units (OTUs) in 4 g of soil. A study on litter decomposition in temperate forests based on meta-DNA confirmed that Ascomycota species have highest relative abundances in the later stages of decomposition (Voříšková and Baldrian 2013). Therefore, high-throughput sequencing methods can be used for fungal diversity estimates. Further, species estimates from different studies may not be compared directly due to differing laboratory standards, protocols and data processing methods (Lindahl et al. 2013; Purahong et al. 2017). However, determination of a common NGS platform, primer pairs and re-analyzation of missing or non-matching gene regions may allow comparison of the results of NGS across different studies and biomes (Nilsson et al. 2011).
How to avoid errors in species identification and estimation of teleomorphic ascomycetes
Species identification and description gives fundamental data for prediction of species number. Hence, documentation of species names in data bases without errors is important. Here we discuss some guidelines to improve this.
Documentation and personal errors
DNA sequences in public databases are annotated by the submitting authors and mostly, further validation by the curators in databases is not often. This approach creates erroneous taxonomic sequence labels (Kozlov et al. 2016). The novel sequences are annotated based on existing ones and mislabeled sequences induce downstream errors. Some sequences in databases are confusing because the same species are classified with different names (Ashelford et al. 2005). This may increase synonyms which are hidden under previous classifications (Bidartondo 2008; Nilsson et al. 2008; Robbertse et al. 2017). Therefore, the taxonomic credibility in sequence databases is with errors (Binder et al. 2005; Bridge et al. 2005; Nilsson et al. 2006). Blast similarities of sequences sometimes give uncultured or unverified sequences and names not identified to species level (Baturo-Cieśniewska et al. 2020). The major reason for these unidentified or insufficiently identified sequences in public databases is that most international journals require all sequences used in the manuscript to be deposited (Ryberg et al. 2008). Errors in protein coding gene sequences give wrongly translated protein sequences which are sometime named as “unverified”. Further, all species in a genus necessary to form a well-supported, monophyletic clade with the type strain of the type species in the genus and the newly introduce species must be in this monophyletic clade without distantly cluster from generic type.
Senanayake et al. (2016) studied the taxonomy and phylogeny of phomatospora-like taxa and showed that Paramicrothyrium chinensis H.X. Wu & K.D. Hyde has 99% similarity to Phomatospora biseriata. Wu et al. (2011) introduced Paramicrothyrium based on P. chinensis using morphology and molecular data. However, the combined LSU and SSU analysis (Wu et al. 2011) showed Paramicrothyrium chinensis as morphologically close to Microthyrium, but phylogenetically distant from Microthyrium. Further, Singtripop et al. (2016) showed that Paramicrothyrium chinensis (IFRDCC 2258) clusters with Chaetothyrina mangiferae (Micropeltidaceae) with high support. Hence, these sequences might have some errors (Senanayake et al. 2016).
Voglmayr et al. (2018) revealed problems in some xylarialean sequences. The LSU part included in the ITS sequences of KP297406 and KP297396 which were deposited as Anthostomella helicofissa and A. forlicesenica respectively, in NCBI searches show obvious variations in the sequence alignment. Further, the LSU sequence of KP340547 was revealed as xylarialean by BLAST searches; however, it has 60 nucleotides difference in LSU sequences with other Anthostomella species. A BLAST search of LSU and RPB2 sequence of KP340538 and KP340524 from MFLUCC 14-0007 (A. forlicesenica) revealed various pleosporalean taxa as the closest matches. Therefore, most scientific journals have recently made it a prerequisite that the sequence data and the alignments of novel taxa must be provided for review to have better quality control.
There is a 1% intra-species variation within Aspergillus niger (Henry et al. 2000). The ITS sequence of Aspergillus niger (MK461010) is identical to ITS sequences of Alternaria alternata (Baturo-Cieśniewska et al. 2020). Further, the ITS sequences of Aspergillus niger (MK461010) shows 10% inter-species variation of ITS sequences of other A. niger strains. This probably suggests a misnomer of the MK461010 sequence. This happens because the person who deposits sequences in the NCBI database determines the species affiliation. The deposition of incorrectly identified sequences creates errors and mistakes because users obtain incorrect species names (Ko et al. 2011; Nilsson et al. 2012). About 65% of the GenBank entries are mislabeled sequences or from poorly characterized vouchers (Leray et al. 2019).
Common, standard data base for taxonomic information of fungi
Recently, most scientific data are documented in websites (Hyde et al. 2020b). Websites gives quick and easy access, sorting and filtering facilities and safeness for data. Currently, there are several databases for molecular, taxonomic and ecological information of teleomorphic ascomycetes (Rossman 1994). There is no standardized protocol for introducing new teleomorphic species in Ascomycota (Seifert and Rossman 2010; Senanayake et al. 2020a). However, there are some formal requirements for proposing names that are imposed by the Nomenclatural Codes (Turland et al. 2018). All the taxonomic information must be available in public for future researchers including taxonomic characters, descriptions, prologues, sequence data, phylogenetic analysis, type specimens, ex-type cultures and ecological data. However, the mycologists have their own preferences and different policies when selecting databases to deposit taxonomic information. Therefore, taxonomic data are scattered in various databanks and this makes difficulties when comparing phenotypic characters. An introduction of “combined platform for taxonomic information of teleomorphic ascomycetes” with a unique identity number for each species is essential. Then, the entire phenotypic, genotypic, chemotaxonomic, ecological, evolutionary, biological and classification data of a teleomorphic species in Ascomycota can be obtained and compared easily. All the data of species can be updated with the newly obtain information. This kind of universal database would minimize errors due to poor updating and also personal arguments. Hibbett et al. (2016) listed commonly used databases and tools for fungal classification and identification.
The number of described, and accepted species in Ascomycota has been counted since 1943 through Ainsworth and Bisby’s Dictionary of the Fungi and currently, the Catalogue of Life web site (https://www.catalogueoflife.org/index?taxonKey=F), which is updated annually through the Species Fungorum, The Integrated Taxonomic Information System and The Global Information System for Lichenized and non-lichenized Ascomycetes outputs.
A standard approach to demarcate species boundaries and describe teleomorphic species in Ascomycota
As there are different approaches to designate a species, mycologysts use different methods according to their requrements and those approaches demarcate species boundaries based on different criteria. Hence, the species numbers vary from one to another and this results complications in species identification and descriptions in Ascomycota (Maharachchikumbura et al. 2021). Therefore, it is necessary to designate a common criterion for species introduction and description (Pažoutová et al. 2013; Kamil et al. 2018). The number of described teleomorphic ascomycetes has increased with the use of molecular techniques for species delimitation. Most teleomorphic ascomycetes introduced earlier than the 1990s lack molecular data from their type collections (Wu et al. 2019a). Hence, it is impossible to compare genotypic characters of fresh collections with genotypic characters of available materials. Therefore, mycologists may misinterpret those collections as new species. Introduction of previously described teleomorphic ascomycetes as new species with molecular data has increased the number of teleomorphic species in the Ascomycota. Further, this value has accelerated with the ending of the separate naming of teleomorph and anamorph of the same species. Therefore, it is necessary to follow well-defined, sets of criteria when introducing a new species and obtain molecular data from the new collection revealing the genetically close taxa. Sequence data should be obtained from both conserved and variable gene regions and all the genes or loci are blasted with GenBank for base pair similarities. Morphological characters could be compared with all phylogenetically similar taxa to ensure that it is not an existing species and also it should be compare with all species in the genus which molecular data unavailable. Ecological data, colony characters, and secondary metabolites provide additional information. Sporulation or conidiation of cultures is important. Describing both teleomorph and anamorph characters and linked them as one is required.
Discussion and conclusion
Assessment of the actual number of teleomorphic species in Ascomycota and their diversity is important for systematics, resource utilization, industrial production and environmental management (Maharachchikumbura et al. 2021). Traditionally, species estimations were based on the numbers of fungi recorded on particular plants and insects. There are several estimations such as around 0.1 million by Bisby and Ainsworth (1943) to 2.2–3.8 million by Hawksworth and Lücking (2017). Traditional species estimation approaches have several drawbacks. The major limitation of these estimates is that they only focus on the fungi that either produces fruiting bodies, which can be identified using a microscope, or those that can be easily cultured on artificial media (Duong et al. 2006). Many ascomycetous endophytes do not sporulate in culture (White and Cole 1986), while some hostspecific species need host tissues to sporulate. Therefore, it is a bias estimate of teleomorphic ascomycetes based on microscopic examination of fruiting structures on substrata or media (Guo et al. 2001; Promputtha et al. 2004). Several studies have shown that most species are singletons found only once (Anslan et al. 2016) or found in only a few locations (Wu et al. 2013) however with plants and animals, some teleomorphic ascomycetes are endemic species. Even though fungi are ubiquitous, individual fungal species are restricted to a specific niche. Therefore, the distribution of teleomorphic ascomycetes is distinct and probably the fungal species number could be much higher than the current estimates (O’Brien et al. 2005; Blackwell 2011).
The development of molecular techniques, such as next generation sequencing has helped identify the previously undescribed diversity in the teleomorphic ascomycetes (Raja et al. 2017; Wilson et al. 2019). Obtaining molecular sequence data from environmental samples from a wide range of localities and substrata has generated a new source of data for estimating species numbers (Wu et al. 2013). The numerous sequences obtained through high-throughput sequencing do not have any close matches with described fungi in sequence databases (Lindahl et al. 2013) and this shows that the current estimated species number is too low. The occurrence of more uncultured teleomorphic ascomycetes also supports that the diversity of fungal species is in underestimation (Hawksworth 1991). However, the main problem in estimates is merging data obtained from traditional culture-based approaches with data obtained from high-throughput sequencing methods (Buée et al. 2009; Kubartová et al. 2012; Dissanayake et al. 2018). Several studies disagree to use sequences derived from high-throughput sequencing methods because of various reasons such as NGS detects only the predominant fungi in a sample instead of all available species (Dissanayake et al. 2018), specific PCR primers need to be developed for some ascomycetes, and the ITS locus is longer than the NGS sequences (Kruse et al. 2017). Further, many ascomycetous species are not well-separated into well-resolved species with a single gene region. Therefore, NGS are suitable to separate fungi only at the genus level (Purahong et al. 2017).
OTUs are also detected by other molecular approaches based on DNA sequence data such as TGGE (thermal gradient gel electrophoresis), DGGE (denaturing gradient gel electrophoresis), SSCP (single-strand conformation polymorphism), RFLP (restriction fragment length polymorphism), TRFLP (terminal restriction fragment length polymorphism), ARDRA (amplified ribosomal DNA restriction analysis), 454 pyrosequencing and illumina MiSeq sequencing can be used to establish a better estimate of species numbers (Hawksworth and Lücking 2017). These methods provide species-specific sequences and therefore, the OTUs generated in these methods reveal an enormous, unprecedented magnitude of fungal diversity.
Wu et al. (2019a, b) estimated that the total fungal diversity is about 12 million species based on meta-DNA and culture-independent methods. In culture-dependent methods, teleomorphic species in Ascomycota are mostly collected and isolated based on their conspicuous fruiting bodies. However, more fungal species are detected by culture-independent approaches and the fungal species detected by culture-independent and culture-dependent approaches often do not overlap (Zhang et al. 2010). Therefore, culture-independent methods are important in estimates of the teleomorphic species in Ascomycota. Hence, neither culture-dependent nor culture-independent method can accurately determinethe structure of a given community. Because of the intrinsic selectivity of each method, the probability of a given species being detected often differs with the method (Zhang et al. 2010).
The above-mentioned problems could be avoided with appropriate morpho-molecular phylogenetic approaches and proper taxon sampling. Furthermore, polyphasic taxonomy should be the preferred approach when introducing new teleomorphic species in Ascomycota, as the conclusions of one method may be eliminated or justified with evidence from the other methods (Maharachchikumbura et al. 2021). Furthermore, molecular data of more extype strains should be included in the phylogenetic analyses as these would provide more clarification where misidentified pleomorphic fungi could be correctly placed. This has not been followed universally by mycologists, thus leading to misidentifications of many fungal species. The best approach to avoid errors in future taxonomic studies of teleomorphic ascomycetes is to focus on increasing missing sequence data and linking those data to digital voucher specimens (Lücking et al. 2020; Jayawardene et al. 2021).
Change history
01 March 2022
A Correction to this paper has been published: https://doi.org/10.1007/s13225-022-00504-1
References
Abdel-Wahab MA, Nagahama T (2011) Gesasha (Halosphaeriaceae, Ascomycota), a new genus with three new species from the Gesashi mangroves in Japan. Nova Hedwigia 92:497–512
Abdel-Wahab MA, Pang KL, Nagahama T, Abdel-Aziz F, Jones EBG (2010) Phylogenetic evaluation of anamorphic species of Cirrenalia and Cumulospora with the description of eight new genera and four new species. Mycol Prog 9:537–558
Acton J, Sandler N (2008) Mushroom. Kyle Cathie, London, p 160
Agerer R, Ammirati JF, Blanz P, Courtecuisse R, Desjardin DE, Gams W, Hallenberg N, Halling R, Hawksworth DL, Horak E, Korf RP, Mueller GM, Oberwinkler F, Rambold G, Summerbell RC, Triebel D, Watling R (2000) Always deposit vouchers. Mycol Res 104(6):642–644
Aime MC, Miller AN, Aoki T, Bensch K, Cai L, Crous PW, Hawksworth DL, Hyde KD, Kirk PM, Lücking R, May TW, Malosso E, Redhead SA, Rossman AY, Stadler M, Thines M, Yurkov AM, Zhang N, Schoch CL (2021) How to publish a new fungal species, or name, version 3.0. IMA Fungus 12:11. https://doi.org/10.1186/s43008-021-00063-1
Albrectsen BR, Bjorken L, Varad A, Hagner A, Wedin M, Karlsson J, Jansson S (2010) Endophytic fungi in European aspen (Populus tremula) leaves-diversity, detection, and a suggested correlation with herbivory resistance. Fungal Divers 41:17–28
Aldhebiani AY (2018) Species concept and speciation. Saudi J Biol Sci 25(3):437–440
Al-Fakih AA, Almaqtri W (2019) Overview on antibacterial metabolites from terrestrial Aspergillus spp. Mycology 10(4):191–209. https://doi.org/10.1080/21501203.2019.1604576
Allen JG, Wood PM (1979) The prevention of lupinosis by making lupin hay. Aust Vet J 1:38–39
Aly AH, Debbab A, Kjer J, Chaidir C (2010) Fungal endophytes from higher plants: a prolific source of phytochemicals and other bioactive natural products. Fungal Divers 41(1):1–16
Amend AS, Seifert KA, Bruns TD (2010) Quantifying microbial communities with 454 pyrosequencing: does read abundance count? Mol Ecol 19:5555–5565
Andersen HL, Ekman S (2005) Disintegration of the Micareaceae (lichenized Ascomycota): a molecular phylogeny based on mitochondrial rDNA sequences. Mycol Res 109(1):21–30
Andrews JH, Harris RF (2000) The ecology and biogeography of microorganisms on plant surfaces. Annu Rev Phytopathol 38(1):145–180
Aneja MK, Sharma S, Fleischmann F, Stich S, Heller W, Bahnweg G, Munch JC, Schloter M (2006) Microbial colonization of beech and spruce litter—influence of decomposition site and plant litter species on the diversity of microbial community. Microb Ecol 52:127–135
Angel K, Wicklow DT (1983) Coprohilous fungal communities in semiarid to mesic grasslands. Can J Bot 61:594–602
Anke T, Steglich W (1988) Forum Mikrobiologie 2:21–25
Anke T (1989) Bioactive metabolites from microorganisms. In: Bushell ME, Gräfe U (eds) Progress in Industrial Microbiology, vol 7. Elsevier, Amsterdam, pp 51–66
Aptroot A (2001) Lichenized and saprobic fungal biodiversity of a single Elaeocarpus tree in Papua New Guinea, with the report of 200 species of ascomycetes associated with one tree. Fungal Divers 6:1–11
Aptroot A, Cáceres MES (2016) First inventory of lichens from the Brazilian Amazon in Amapá State. Bryologist 119:250–265
Araújo JPM, Hughes DP (2016) Diversity of entomopathogenic fungi: which groups conquered the insect body? Adv Genet 94:1–39
Arenz BE, Held BW, Jurgens JA, Farrell RL, Blanchette RA (2006) Fungal diversity in soils and historic wood from the Ross Sea Region of Antarctica. Soil Biol Biochem 38:3057–3064
Ariyawansa HA, Hawksworth DL, Hyde KD, Jones EBG, Maharachchikumbura SSN, Manamgoda DS, Thambugala KM, Udayanga D, Camporesi E, Daranagama A, Jayawardena R, Liu JK, McKenzie EHC, Phookamsak R, Senanayake IC, Shivas RG, Tian Q, Xu JC (2014) Epitypification and neotypification: guidelines with appropriate and inappropriate examples. Fungal Divers 69:57–91
Ashelford KE, Chuzhanova NA, Fry JC, Jones AJ, Weightman AJ (2005) At least 1 in 20 16S rRNA sequence records currently held in public repositories is estimated to contain substantial anomalies. Appl Environ Microbiol 71:7724–7736
Aung OM, Soytong K, Hyde KD (2008) Diversity of entomopathogenic fungi in rainforests of Chiang Mai Province. Thailand Fungal Divers 30:15–22
Azevedo JL, Araújo WL (2007) Diversity and applications of endophytic fungi isolated from tropical plants. In: Ganguli BN, Deshmukh SK (eds) Fungi: multifaceted microbes. CRC Press, Boca Raton, pp 189–207
Baldrian P (2017) Forest microbiome: diversity, complexity and dynamics. FEMS Microbiol Rev 41(2):109–130
Baldrian P, Lindahl BD (2011) Decomposition in forest ecosystems: after decades of research still novel findings. Fungal Ecol 4(6):359–361
Baral HO, Weber E, Gams W, Hagedorn WG, Liu B, Liu X, Marson G, Marvanová L, Stadler M, Weiß M (2018) Generic names in the Orbiliaceae (Orbiliomycetes) and recommendations on which names should be protected or suppressed. Mycol Prog 17:5–31. https://doi.org/10.1007/s11557-017-1300-6
Barnes M, Turner C (2015) The ecology of environmental DNA and implications for conservation genetics. Conserv Genet. https://doi.org/10.1007/s10592-015-0775-4
Barr ME (1978) The Diaporthales in North America: with emphasis on Gnomonia and its segregates. Mycol Mem 7:1–232
Barr ME (1987) New taxa and combinations in the Loculoascomycetes. Mycotaxon 29:501–505
Barron GL (1968) The Genera of Hyphomycetes from Soil. Williams and Wilkins, Baltimore, MD, p 364
Barton NH (1979) Gene flow past a cline. Heredity 43:333–339
Barton NH, Gale KS (1993) Genetic analysis of hybrid zones. In: Harrison RG (ed) Hybrid zones and the evolutionary process. Oxford University Press, Oxford, pp 13–45
Barton NH, Hewitt GM (1985) Analysis of hybrid zones. Annu Rev Ecol Syst 16:113–148
Bass D, Richards TA (2011) Three reasons to re-evaluate fungal diversity ‘on Earth and in the ocean.’ Fungal Biol Rev 25:159–164
Basumatary S, McDonald H (2017) Coprophilous fungi from dung of the Greater One-Horned Rhino in Kaziranga National Park, India and its implication to paleoherbivory and paleoecology. Quat Res 88:14–22
Baturo-Cieśniewska A, Groves CL, Albrecht KA, Grau CR, Willis DK, Smith DL (2017) Molecular identification of Sclerotinia trifoliorum and Sclerotinia sclerotiorum isolates from the United States and Poland. Plant Dis 101(1):192–199
Baturo-Cieśniewska A, Pusz W, Patejuk K (2020) Problems, limitations, and challenges in species identification of Ascomycota members on the basis of ITS regions. Acta Mycol 55(1):5512. https://doi.org/10.5586/am.5512
Becker TCA, de Castro-Prado MAA (2006) Parasexuality in asexual development mutants of Aspergillus nidulans. Biol Res 39(2):297–305
Beimforde C, Schmidt AR, Rikkinen J, Mitchell JK (2020) Sareomycetes cl. nov.: a new proposal for placement of the resinicolous genus Sarea (Ascomycota, Pezizomycotina). Fungal Syst Evol 6:25–37
Bell A (2005) An illustrated guide to the coprophilous ascomycetes of Australia. CBS Biodiversity Series, Utrecht
Bellemain E, Carlsen T, Brochmann C, Coissac E, Taberlet P, Kauserud H (2010) ITS as an environmental DNA barcode for fungi: an in silico approach reveals potential PCR biases. BMC Microbiol 10(1):189. https://doi.org/10.1186/1471-2180-10-189
Bennett RJ, Turgeon BG (2017) Fungal sex: the Ascomycota. In: Heitman J, Howlett BJ, Crous PW, Stukenbrock EH, James TY, Gow NAR (eds) The fungal kingdom. ASM Press, Washington, pp 117–145
Bensch K, Groenewald JZ, Meijer M, Dijksterhuis J, Jurjević Ž, Andersen B, Houbraken J, Crous PW, Samson RA (2018) Cladosporium species in indoor environments. Stud Mycol 89:177–301
Berbee ML, Taylor JW (1992) Two ascomycete classes based on fruiting-body characters and ribosomal DNA sequence. Mol Biol Evol 9:278–284
Bickford D, Lohman DJ, Sodhi NS, Ng PKL, Meier R, Winker K, Ingram KK, Das I (2007) Cryptic species as a window on diversity and conservation. Trends Ecol Evol 22(3):148–155
Bidartondo MI (2008) Preserving accuracy in GenBank. Science 319(5870):1616
Bills GF (1996) Isolation and analysis of endophytic fungal communities from woody plants. In: Redlin SC, Carris LM (eds) Endophytic fungi in grasses and woody plants: systematic, ecology and evolution. APS Press, St. Paul, pp 31–65
Binder M, Hibbett DS, Larsson KH, Larsson E, Langer E, Langer G (2005) The phylogenetic distribution of resupinate forms across the major clades of mushroom forming fungi (Homobasidiomycetes). Syst Biodivers 3(2):113–157
Bisby GR, Ainsworth GC (1943) The numbers of fungi. T Brit Mycol Soc 26:16–19
Blackwell M (2011) The fungi: 5.1 million species? Am J Bot 98:426–438
Blackwell M, Haelewaters D, Pfister DH (2020) Laboulbeniomycetes: Evolution, natural history, and Thaxter’s final word. Mycologia 112(6):1048–1059
Boddy L, Frankland JC, Van West P (2008) Ecology of saprotrophic basidiomycetes. British Mycological Society Symposia Series (vol 28). Elsevier, London
Bouck AC, Peeler R, Arnold ML, Wessler SR (2005) Genetic mapping of species boundaries in Louisiana irises using IRRE retrotransposon display markers. Genetics 171:1289–1303
Bourtzis K, Miller TA (2003) Insect symbiosis, 1st edn. CRC Press, Florida, USA, p 368
Bridge P, Spooner B (2001) Soil fungi: Diversity and detection. Plant Soil 232(1):147–154
Bridge PD, Spooner BM, Roberts PJ (2005) The impact of molecular data in fungal systematics. Adv Bot Res 42:33–67
Brodo IM, Sharnoff SD, Sharnoff S (2001) Lichens of North America. Yale University Press, London, p 828
Brown TA (2002) Genomes, 2nd edition. Oxford Wiley-Liss, United Kingdom, 600pp.
Brundrett MC (2002) Coevolution of roots and mycorrhizas of land plants. New Phytol 154(2):275–304
Buèe M, Reich M, Murat C, Morin E, Nilsson RH, Uroz S, Martin F (2009) 454 Pyrosequencing analyses of forest soils reveal an unexpectedly high fungal diversity. New Phytol 184:449–456
Burgarella C, Barnaud A, Kane NA, Jankowski F, Scarcelli N, Billot C, Vigouroux Y, Berthouly-Salazar C (2019) Adaptive introgression: an untapped evolutionary mechanism for crop adaptation. Front Plant Sci 10:4. https://doi.org/10.3389/fpls.2019.00004
Cáceres MES, Aptroot A, Lücking R (2017) Lichen fungi in the Atlantic rain forest of Northeast Brazil: the relationship of species richness with habitat diversity and conservation status. Braz J Bot 40:145–156
Calabon MS, Jones EBG, Boonmee S, Doilom M, Lumyong S, Hyde KD (2021) Five novel freshwater ascomycetes indicate high undiscovered diversity in lotic habitats in Thailand. J Fungi 7:117. https://doi.org/10.3390/jof7020117
Calaça FJS, Delpont M, Xavier-Santos S (2015) Delitschia gigaspora var. pescanii: a new variety of coprophilous fungi from Brazil. Mycosphere 6(1):122–126
Calaça FJS, Cortez VG, Santos SX (2020) Dung fungi from Brazil: Agrocybe pediades (Fr.) Fayod (Basidiomycota) in Cerrado. Scientia Plena 16(6):1–6
Cannon PF, Hawksworth DL (1995) The diversity of fungi associated with vascular plants: the known, the unknown and the need to bridge the knowledge gap. Adv Plant Pathol 11:277–302
Castlebury LA, Rossman AY, Jaklitsch WM, Vasilyeva LN (2002) A preliminary overview of the Diaporthales based on large subunit nuclear ribosomal DNA sequences. Mycologia 94:1017–1031
Catalogue of Life (2021) Available at http://www.catalogueoflife.org/annual-checklist/. Accessed 30 Sep 2021
Challacombe JF, Hesse CN, Bramer LM, McCue LA, Lipton M, Purvine S, Nicora C, Gallegos-Graves LV, Porras-Alfaro A, Kuske CR (2019) Genomes and secretomes of Ascomycota fungi reveal diverse functions in plant biomass decomposition and pathogenesis. BMC Genomics 20:976. https://doi.org/10.1186/s12864-019-6358-x
Chan CL, Yew SM, Ngeow YF, Na SL, Lee KW, Hoh CC, Yee WY, Ng KP (2015) Genome analysis of Daldinia eschscholtzii strains UM 1400 and UM 1020, wood-decaying fungi isolated from human hosts. BMC Genomics 16(1):1–14
Chethana KWT, Jayawardena RS, Hyde KD (2020) Hurdles in fungal taxonomy: effectiveness of recent methods in discriminating taxa. Megataxa 1(2):114–122
Chomnunti P, Hongsanan S, Aguirre-Hudson B, Tian Q, Peršoh D, Dhami MK, Alias AS, Xu JC, Liu XZ, Stadler M, Hyde KD (2014) The Sooty Moulds. Fungal Divers 66:1–36
Clark MA, Douglas M, Choi J (2018) Biology, 2nd edn. OpenStax, Houston, p 1578
Çobanoğlu G, Sesal C, Gökmen B, Çakar S (2010) Evaluation of the antimicrobial properties of some lichens. South-West J Hortic Biol Environ 1(2):153–158
Crous PW, Schoch CL, Hyde KD, Wood AR, Gueidan C, de Hoog GS, Groenewald JZ (2009) Phylogenetic lineages in the Capnodiales. Stud Mycol 64:17–47
Crous PW, Quaedvlieg W, Hansen K, Hawksworth DL, Groenewald JZ (2014) Phacidium and Ceuthospora (Phacidiaceae) are congeneric: taxonomic and nomenclatural implications. IMA Fungus 5(2):173–193
Crozier J, Thomas SE, Aime MC, Evans HC, Holmes KA (2006) Molecular characterization of fungal endophytic morphospecies isolated from stems and pods of Theobroma cacao. Plant Pathol 55(6):783–791
Dashtban N, Schraft H, Syed TA, Qin W (2010) Fungal biodegradation and enzymatic modification of lignin. Int J Biochem Mol Biol 1:36–50
Dayarathne MC, Boonmee S, Braun U, Crous PW, Daranagama DA, Dissanayake AJ, Ekanayaka H, Jayawardena R, Jones EBG, Maharachchikumbura SSN, Perera RH, Phillips AJL, Stadler M, Thambugala KM, Wanasinghe DN, Zhao Q, Hyde KD, Jeewon R (2016) Taxonomic utility of old names in current fungal classification and nomenclature conflicts, confusion & clarifications. Mycosphere 7(11):1622–1648
Dayarathne MC, Maharachchikumbura SSN, Jones EBG, Dong W, Devadatha B, Yang J, Ekanayaka AH, De Silva W, Sarma VV, Al-Sadi AM, Khongphinitbunjong K, Hyde KD, Zhao RL (2019) Phylogenetic revision of Savoryellaceae and evidence for its ranking as a subclass. Front Microbiol 10:840. https://doi.org/10.3389/fmicb.2019.00840
De Hoog GS, Horré R (2002) Molecular taxonomy of the Alternaria and Ulocladium species from humans and their identification in the routine laboratory. Mycoses 45(7–8):259–276
De Meeûs T, Renaud F (2002) Parasites within the new phylogeny of eukaryotes. Trends Parasitol 18:247–251
De Queiroz A, Wimberger PH (1993) The usefulness of behaviour for phylogeny estimation: levels of homoplasy in behavioural and morphological characters. Int J Org Evol 47(1):46–60
De Boer W, Folman LB, Summerbell RC, Boddy L (2005) Living in a fungal world: impact of fungi on soil bacterial niche development. FEMS Microbiol Rev 29:795–811
De Souza CA, Lima DX, Gurgel LM, Santiago AL (2017) Coprophilous Mucorales (ex Zygomycota) from three areas in the semi-arid of Pernambuco. Brazil Braz J Microbiol 48(1):79–86
Debuchy R, Berteaux-Lecellier V, Silar P (2010) Mating systems and sexual morphogenesis in ascomycetes. In: Borkovich KA, Ebbole DJ (eds) Cellular and molecular biology of filamentous fungi. ASM Press, Washington, pp 501–535
De-Forest J (2009) The influence of time, storage temperature, and substrate age on potential soil enzyme activity in acidic forest soils using MUB-linked substrates and L-DOPA. Soil Biol Biochem 41(6):1180–1186
Dissanayake AJ, Purahong W, Wubet T, Hyde KD, Zhang W, Xu H, Zhang G, Fu C, Liu M, Xing Q, Li X, Yan J (2018) Direct comparison of culture-dependent and culture-independent molecular approaches reveal the diversity of fungal endophytic communities in stems of grapevine Vitis vinifera. Fungal Divers 90:85–107
Divakar PK, Upreti DK (2005) Parmelioid lichens in India (A revisionary study). Bishen Singh Mahendra Pal Singh, Dehra Dun, p 488
Divakar PK, Kauff F, Crespo A, Leavitt SD, Lumbsch HT (2013) Understanding Phenotypical Character Evolution in Parmelioid Lichenized Fungi (Parmeliaceae, Ascomycota). PLoS ONE 8(11):e83115
Domsch KH, Gams W, Anderson TH (1993) Compendium of soil fungi. IHW, Eching
Dong C, Wang L, Li Q, Shang Q (2021) Epiphytic and endophytic fungal communities of tomato plants. Hortic Plant J 7(1):38–48
Dreyfuss MM, Chapela IH (1994) Potential of fungi in the discovery of novel, low-molecular weight pharmaceuticals. In: Gullo V (ed) The discovery of natural products with therapeutic potential. Butterworth Heinemann, London, pp 49–80
Duong LM, Jeewon R, Lumyong S, Hyde KD (2006) DGGE coupled with ribosomal DNA gene phylogenies reveal uncharacterized fungal phylotypes. Fungal Divers 23:121–138
Dyer PS (2007) Sexual reproduction and the significance of MAT in the Aspergilli. In: Heitman J, Kronstad JW, Taylor JW, Casselton LA (eds) Sex in fungi: molecular determination and evolutionary implications. American Society of Microbiology Press, Washington, pp 123–142
Ekanayaka AH, Hyde KD, Jones EBG, Zhao Q (2018) Taxonomy and phylogeny of operculate discomycetes: Pezizomycetes. Fungal Divers 90:161–243
El-Elimat RHA, Figueroa M, Al Sharie AH, Bunch RL, Oberlies NH (2021) Freshwater fungi as a source of chemical diversity: a review. J Nat Prod 84:898–916
Fan K, Weisenhorn P, Gilbert JA, Chu H (2018) Wheat rhizosphere harbors a less complex and more stable microbial co-occurrence pattern than bulk soil. Soil Biol Biochem 125:251–260
Fernández-Mendoza F, Domaschke S, García MA, Jordan P, Martín MP, Printzen C (2011) Population structure of mycobionts and photobionts of the widespread lichen Cetraria aculeata. Mol Ecol 20:1208–1232
Ferreira AVB, An Z, Metzenberg RL, Glass NL (1998) Characterization of matA-2, matA-3 and ΔmatA Mating-Type Mutants of Neurospora crassa. Genetics 148:1069–1079
Fesel PH, Zuccaro A (2015) Beta-glucan: Crucial component of the fungal cell wall and elusive MAMP in plants. Fungal Genet Biol 90:53–60
Fisher MC, Henk DA, Briggs CJ, Brownstein JS, Madoff LC, McCraw SL, Gurr SJ (2012) Emerging fungal threats to animal, plant and ecosystem health. Nature 484(7393):186–194
Florenzano A (2019) The history of pastoral activities in italy inferred from palynology: a long-term perspective to support biodiversity awareness. Sustainability 11(2):404. https://doi.org/10.3390/su11020404
Forche A, Alby K, Schaefer D, Johnson AD, Berman J, Bennett RJ (2008) The parasexual cycle in Candida albicans provides an alternative pathway to meiosis for the formation of recombinant strains. PLoS Biol 6:e110
Frąc M, Hannula SE, Bełka M, Jędryczka M (2018) Fungal biodiversity and their role in soil health. Front Microbiol 9:707. https://doi.org/10.3389/fmicb.2018.00707
François L, Mark P, Valérie R (2001) Major fungal lineages are derived from lichen symbiotic ancestors. Nature 411(6840):937–940
Frankland JC (1998) Fungal succession—unraveling the unpredictable. Mycol Res 102:1–15
Freeman J, Ward E, Calderon C, McCartney A (2002) A polymerase chain reaction (PCR) assay for the detection of inoculum of Sclerotinia sclerotiorum. Eur J Plant Pathol 108(9):877–886
Frey-Klett P, Burlinson P, Deveau A, Barret M, Tarkka M, Sarniguet A (2011) Bacterial-fungal interactions: hyphens between agricultural, clinical, environmental, and food microbiologists. Microbiol Mol Biol Rev 75(4):583–609
Fries EM (1849) A summary of the plants of Scandinavia. The latter section. Typographia Academica, Uppsala, Stockholm, pp 259–572
Species Fungorum (2021) http://www.speciesfungorum.org Accessed 24 Sep 2021
Gams W (2007) Biodiversity of soil-inhabiting fungi. Biodivers Conserv 16(1):69–72
Garcia-Reyne A, López-Medrano F, Morales JM, Esteban CG, Martín I, Eraña I, Meije Y, Lalueza A, Alastruey-Izquierdo A, Rodríguez-Tudela JL, Aguado JM (2011) Cutaneous infection by Phomopsis longicolla in a renal transplant recipient from Guinea: first report of human infection by this fungus. Transpl Infect Dis 13:204–207
Gardiner MR (1975) Lupinosis. J Agric Western Australia 16:26–30
Garty J (2001) Biomonitoring atmospheric heavy metals with lichens: theory and application. Crit Rev Plant Sci 20:309–371
Gilman JC (1957) A manual of soil fungi, 2nd edn. Iowa State University Press Inc., Iowa
Giordano L, Gonthier P, Lione G, Garbelotto M (2019) The invasiveness of a non-native fungal forest pathogen is boosted by the presence of a congeneric native species. Forestry 92(2):157–166
Glawe DA (1983) Observations on the Anamorph of Eutypella parasitica. Mycologia 75(4):742–743
Glawe DA (1985) Hapalocystis berkeleyi and Macrodiplodiopsis desmazieresii in artificial culture. Mycologia 77(6):880–886
Glawe DA, Rogers JD (1982) Observations on the anamorphs of six species of Eutypa and Eutypella (Fungi, structure, descriptions). Mycotaxon 14(1):334–346
Glawe DA, Rogers JD (1984) Diatrypaceae in the Pacific Northwest. Mycotaxon 20:401–460
Gonçalves MFM, Abreu AC, Hilário S, Alves A (2021) Diversity of marine fungi associated with wood baits in the estuary Ria de Aveiro, with descriptions of Paralulworthia halima, comb. Nov., Remispora submersa, sp. nov., and Zalerion pseudomaritima, sp. nov. Mycologia 113(3):664–683
Gouda S, Das G, Sen SK, Shin HS, Patra JK (2016) Endophytes: a treasure house of bioactive compounds of medicinal importance. Front Microbiol 7:1538. https://doi.org/10.3389/fmicb.2016.01538
Gould AB (2008) Fungi: plant pathogenic. In: Trigiano RJ, Windham MT, Windham AS (eds) Plant pathology concepts and laboratory exercises, 2nd edn. CRC Press, Boca Raton, pp 457–477
Greville RK (1824) Flora Edinensis: or a description of plants growing near Edinburgh, arranged according to the Linnean system. Blackwood, Edinburgh, p 478
Grossart HP, Van den Wyngaert S, Kagami M, Wurzbacher C, Cunliffe M, Rojas-Jimenez K (2019) Fungi in aquatic ecosystems. Nat Rev Microbiol 17:339–354
Grove WB (1937) British stem- and leaf-fungi (Coelomycetes), vol 2. Cambridge University Press, Cambridge, p 405
Grube M, Winka K (2002) Progress in understanding the evolution and classification of lichenized ascomycetes. Mycologist 16(2):67–76
Guo LD, Hyde KD, Liew ECY (2001) Detection and taxonomic placement of endophytic fungi within frond tissues of Livistona chinensis based on rDNA sequences. Mol Phylogenet Evol 20:1–13
Guo B, Wang Y, Sun X, Tang K (2008) Bioactive natural products from endophytes: a review. Appl Biochem Microbiol 44:136–142
Haelewaters D, Blackwell M, Pfister DH (2021) Laboulbeniomycetes: intimate fungal associates of Arthropods. Annu Rev Entomol 66:257–276
Hall AR, Colegrave N (2008) Decay of unused characters by selection and drift. J Evol Biol 21(2):610–617
Hamzah TNT, Lee SY, Hidayat A, Terhem R, Faridah-Hanum I, Mohamed R (2018) Diversity and characterization of endophytic fungi isolated from the tropical mangrove species, Rhizophora mucronata, and identification of potential antagonists against the soil-borne fungus Fusarium solani. Front Microbiol 9:1707. https://doi.org/10.3389/fmicb.2018.01707
Haňáčková Z, Koukol O, Štursová M, Kolařík M, Baldrian P (2015) Fungal succession in the needle litter of a montane Picea abies forest investigated through strain isolation and molecular fingerprinting. Fungal Ecol 13:157–166
Hardoim PR, van Overbeek LS, Berg G, Pirttilä AM, Compant S, Campisano A, Döring M, Sessitsch A (2015) The hidden world within plants: ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol Mol Biol Rev 79(3):293–320
Hawksworth DL (1991) The fungal dimension of biodiversity: magnitude, significance, and conservation. Mycol Res 95:641–655
Hawksworth DL (2001) The magnitude of fungal diversity: the 1±5 million species estimate revisited. Mycol Res 105(12):1422–1432
Hawksworth DL (2012) Managing and coping with names of pleomorphic fungi in a period of transition. IMA Fungus 3(1):15–24
Hawksworth DL (2015) Proposals to clarify and enhance the naming of fungi under the International Code of Nomenclature for algae, fungi, and plants. IMA Fungus 6:199–205. https://doi.org/10.5598/imafungus.2015.06.01.12
Hawksworth DL, Rossman AY (1997) Where are all the undescribed fungi? Phytopathology 87:888–891
Hawksworth DL, Kirk PM, Sutton BC, Pegler DN (1995) Ainsworth and Bisby’s Dictionary of the Fungi. CAB International, Wallingford, p 424
Hawksworth DL, McNeill J, de Beer ZW, Wingfield MJ (2013) Names of fungal species with the same epithet applied to different morphs: how to treat them. IMA Fungus 4(1):53–56
Hawksworth DL, Hibbett DS, Kirk PM, Lücking R (2016) (308–310) Proposals to permit DNA sequence data to serve as types of names of fungi. Taxon 65:899–900
Hawksworth DL, Lücking R (2017) Fungal diversity revisited: 22 to 38 million species. Microbiol Spectr 5(4):79–95
He X, Han G, Lin Y, Tian X, Xiang C, Tian Q, He Z (2012) Diversity and decomposition potential of endophytes in leaves of a Cinnamomum camphora plantation in China. Ecol Res 27(2):273–284
Heeger F, Bourne EC, Baschien C, Yurkov A, Bunk B, Spröer C, Overmann J, Mazzoni CJ, Monaghan MT (2018) Long-read DNA metabarcoding of ribosomal rRNA in the analysis of fungi from aquatic environments. bioRxiv. https://doi.org/10.1101/283127
Helaly SE, Thongbai B, Stadler M (2018) Diversity of biologically active secondary metabolites from endophytic and saprotrophic fungi of the ascomycete order Xylariales. Nat Prod Rep 35:992–1014
Herder J, Valentini A, Bellemain E, Dejean T, Delft J, Thomsen P, Taberlet P (2014) Environmental DNA—a review of the possible applications for the detection of (invasive) species. Stichting RAVON, Nijmegen. Report 2013, 104
Hernández-Restrepo M, Gené J, Castañeda-Ruiz RF, Mena-Portales J, Crous PW, Guarro J (2017) Phylogeny of saprobic microfungi from Southern Europe. Stud Mycol 86:53–97
Hibbett DS, Ohman A, Glotzer D, Nuhn M, Kirk P, Nilsson RH (2011) Progress in molecular and morphological taxon discovery in fungi and options for formal classification of environmental sequences. Fungal Biol Rev 25:38–47
Hibbett DS, Abarenkov K, Kõljalg U, Opik M, Chai BR, Cole’s J, Geiser D (2016) Sequence-based classification and identification of fungi. Mycologia 108:1049–1068
Hirakawa MP, Chyou DE, Huang D, Slan AR, Bennett RJ (2017) Parasex generates phenotypic diversity de novo and impacts drug resistance and virulence in Candida albicans. Genetics 207:1195–1211
Hirsch J, Galidevara S, Strohmeier S, Devi KU, Reineke A (2013) Effects on diversity of soil fungal community and fate of an artificially applied Beauveria bassiana strain assessed through 454 pyrosequencing. Microb Ecol 66:608–620
Homei A (2006) Medical mycology development and epidemiology in the USA, UK and Japan. Med Mycol 44:S39–S54
Hongsanan S, Jeewon R, Purahong W, Xie N, Liu JK, Jayawardena RS, Ekanayaka AH, Dissanayake A, Raspé O, Hyde KD, Stadler M, Peršoh D (2018) Can we use environmental DNA as holotypes? Fungal Divers 92:1–30
Hörandl E, Stuessy TF (2010) Paraphyletic groups as natural units of biological classification. Taxon 59(6):1641–1653
Horton TR, Bruns TD (2001) The molecular revolution in ectomycorrhizal ecology: peeking into the black-box. Mol Ecol 10(8):1855–1871
Hsieh HM, Lin CR, Fang MJ, Rogers JD, Fournier J, Lechat C, Ju YM (2010) Phylogenetic status of Xylaria subgenus Pseudoxylaria among taxa of the subfamily Xylarioidea (Xylariaceae) and phylogeny of the taxa involved in the subfamily. Mol Phylogen Evol 54:957–969
Hu DM, Liu F, Cai L (2013) Biodiversity of aquatic fungi in China. Mycology 4:125–168
Huang SK, Hyde KD, Mapook A, Maharachchikumbura SSN, Bhat DJ, McKenzie EHC, Jeewon R, Wen TC (2021) Taxonomic studies of some often over-looked diaporthomycetidae and sordariomycetidae. Fungal Divers. https://doi.org/10.21203/rs.3.rs-517350/v1
Huhndorf SM, Lodge DJ (1997) Host specificity among wood-inhabiting pyrenomycetes (Fungi, Ascomycetes) in a wet tropical forest in Puerto Rico. Trop Ecol 38(2):307–315
Huhndrof SM, Lodge DJ (1997) Host specificity amoung wood-inhabiting pyrenomycetes (Fungi, Ascomycetes) in a wet tropical forest in Puerto Rico. Trop Ecol 38:307–315
Hyde KD, Goh TK (2003) Adaption of dispersal in filmentous freshwater fungi. Fungal Diverse Res Ser 10:231–258
Hyde KD, Jones EBG, Leano E, Pointing SB, Poonyth AD, Vrijmoed LLP (1998) Role of fungi in marine ecosystems. Biodivers Conserv 7:1147–1161
Hyde KD, Jones EBG, Liu JK, Ariyawansa H, Boehm E, Boonmee S, Braun U, Chomnunti P, Crous PW, Dai DQ, Diederich P, Dissanayake A, Doilom M, Doveri F, Hongsanan S, Jayawardena R, Lawrey JD, Li YM, Liu YX, Lücking R, Monkai J, Muggia L, Nelsen MP, Pang KL, Phookamsak R, Senanayake IC, Shearer CA, Suetrong S, Tanaka K, Thambugala KM, Wijayawardene NN, Wikee S, Wu HX, Zhang Y, Aguirre-Hudson B, Alias SA, Aptroot A, Bahkali AH, Bezerra JL, Bhat DJ, Camporesi E, Chukeatirote E, Gueidan C, Hawksworth DL, Hirayama K, Hoog SD, Kang JC, Knudsen K, Li WJ, Li XH, Liu ZY, Mapook A, McKenzie EHC, Miller AN, Mortimer PE, Phillips AJL, Raja HA, Scheuer C, Schumm F, Taylor JE, Tian Q, Tibpromma S, Wanasinghe DN, Wang Y, Xu JC, Yacharoen S, Yan JY, Zhang M (2013) Families of Dothideomycetes. Fungal Divers 63:1–313
Hyde KD, Al-Hatmi A, Andersen B, Boekhout T, Buzina W, Dawson JTL, Eastwood DC, Gareth Jones EB, de Hoog S, Kang Y, Longcore JE, McKenzie EHC, Meis JF, Pinson-Gadais L, Rathnayaka AR, Richard-Forget F, Stadler M, Theelen B, Thongbai B, Tsui CKM (2018) The world’s ten most feared fungi. Fungal Divers 93:161–194
Hyde KD, Xu J, Rapior S, Jeewon R, Lumyong S, Niego AGT, Abeywickrama PD, Aluthmuhandiram JV, Brahamanage RS, Brooks S, Chaiyasen A, Chethana KWT, Chomnunti P, Chepkirui C, Chuankid B, de Silva NI, Doilom M, Faulds C, Gentekaki E, Gopalan V, Kakumyan P, Harishchandra D, Hemachandran H, Hongsanan S, Karunarathna A, Karunarathna SC, Khan S, Kumla J, Jayawardena RS, Liu JK, Liu N, Luangharn T, Macabeo APG, Marasinghe DS, Meeks D, Mortimer PE, Mueller P, Nadir S, Nataraja KN, Nontachaiyapoom S, O’Brien M, Penkhrue W, Phukhamsakda C, Ramanan US, Rathnayaka AR, Sadaba RB, Sandargo B, Samarakoon BC, Tennakoon DS, Siva R, Sriprom W, Suryanarayanan TS, Sujarit K, Suwannarach N, Suwunwong T, Thongbai B, Thongklang N, Wei D, Wijesinghe SN, Winiski J, Yan J, Yasanthika E, Stadler M (2019) The amazing potential of fungi: 50 ways we can exploit fungi industrially. Fungal Divers 97:1–136
Hyde KD, Chethana KWT, Jayawardena RS, Luangharn T, Calabon MS, Jones EBG, Hongsanan S, Lumyongi S (2020a) The rise of mycology in Asia. Sci Asia 46S:1–11
Hyde KD, Jeewon R, Chen YJ, Bhunjun C, Calabon MS, Jiang HB, Lin CG, Norphanphoun C, Sysouphanthong P, Pem D, Tibpromma S, Zhang Q, Doilom M, Jayawardena RS, Liu JK, Maharachchikumbura SSN, Phukhamsakda C, Phookamsak R, Al-Sadi AM, Thongklang N, Wang Y, Gafforov Y, Jones EBG, Lumyong S (2020b) The numbers of fungi, is the descriptive curve flattening? Fungal Divers 103:219–271
Hyde KD, Norphanphoun C, Maharachchikumbura SSN, Bhat DJ, Jones EBG, Bundhun D, Chen YJ, Bao DF, Boonmee S, Calabon MS, Chaiwan N, Chethana KWT, Dai DQ, Dayarathne MC, Devadatha B, Dissanayake AJ, Dissanayake LS, Doilom M, Dong W, Fan XL, Goonasekara ID, Hongsanan S, Huang SK, Jayawardena RS, Jeewon R, Karunarathna A, Konta S, Kumar V, Lin CG, Liu JK, Liu NG, Luangsa-ard J, Lumyong S, Luo ZL, Marasinghe DS, McKenzie EHC, Niego AGT, Niranjan M, Perera RH, Phukhamsakda C, Rathnayaka AR, Samarakoon MC, Samarakoon SMBC, Sarma VV, Senanayake IC, Shang QJ, Stadler M, Tibpromma S, Wanasinghe DN, Wei DP, Wijayawardene NN, Xiao YP, Yang J, Zeng XY, Zhang SN, Xiang MM (2020c) Refined families of sordariomycetes. Mycosphere 11(1):305–1059
Hyde KD, Jones EBG (2002) Introduction to fungal succession. Fungal Divers. 10:1–4
Hywel-Jones N (1993) A systematic survey of insect fungi from natural tropical forest in Thailand. In: Isaac S, Frankland JC, Watling R, Whalley AJS (eds) Aspects of tropical mycology. Cambridge University Press, Cambridge, pp 300–301
Ingold CT, Hudson HJ (1993) Ecology of saprotrophic fungi. The biology of fungi. Springer, Dordrecht, pp 145–157
IUCN (2021) Red list of threatened species. International Union for Conservation of Nature and Natural Resources, Version 2019–3, ISSN 2307–8235. Retrieved on 20/9/2021
Jaklitsch WM, Baral HO, Lücking R, Lumbsch HT, Frey W (2016) Syllabus of plant families—A. Engler’s Syllabus der Pflanzenfamilien Part 1/2: Ascomycota, 13th edn. Borntraeger, Stuttgart, p 322
Jayaraman S, Naorem A, Lal R, Dalal RC, Sinha NK, Patra AK, Chaudhari SK (2021) Disease-suppressive soils—beyond food production: a critical review. J Soil Sci Plant Nutr 21:1437–1465
Jayawardena RS, Bhunjun CS, Hyde KD, Gentekaki E, Itthayakorn P (2021) Colletotrichum: lifestyles, biology, morpho-species, species complexes and accepted species. Mycosphere 12(1):519–669
Jeewon R, Hyde KD (2016) Establishing species boundaries and new taxa among fungi: recommendations to resolve taxonomic ambiguities. Mycosphere 7:1669–1677
Jiang N, Liang YM, Tian CM (2020) A novel bambusicolous fungus from China, Arthrinium chinense (Xylariales). Sydowia 72:77–83
Jones EBG (2000) Marine fungi: some factors influencing biodiversity. Fungal Divers 4:53–73
Jones EBG (2011) Are there more marine fungi to be described? Bot Mar 54:343–354
Jones EBG, Hyde KD, Vikineswary S (2007) Malaysian Fungal Diversity, Mushroom Research Centre, University of Malaya and Ministry of Natural Resources and Environment, Malaysia
Jones EBG, Sakayaroj J, Suetrong S, Somrithipol S, Pang KL (2009) Classification of marine Ascomycota, anamorphic taxa and Basidiomycota. Fungal Divers 35:1–187
Jones MDM, Richards TA, Hawksworth DL, Bass D (2011) Validation and justification of the phylum name Cryptomycota phyl. IMA Fungus 2:173–175
Jones EBG, Suetrong S, Sakayaroj J, Bahkali AH, Abdel-Wahab MA, Boekhout T, Pang KL (2015) Classification of marine Ascomycota, Basidiomycota, Blastocladiomycota and Chytridiomycota. Fungal Divers 73:1–72
Jones EBG, Pang KL, Abdel-Wahab MA, Scholz B, Hyde KD, Boekhout T, Ebel R, Rateb ME, Henderson L, Sakayaroj J, Suetrong S, Dayarathne MC, Kumar V, Raghukumar S, Sridhar KR, Bahkali AHA, Gleason FH, Norphanphoun C (2019) An online resource for marine fungi. Fungal Divers 96:347–433
Judson OP, Normark BB (1996) Ancient Asexual Scandals. Trends Ecol Evol 11:41–46
Kamil FH, Saeed EE, Tarabily KAEI, AbuQamar S (2018) Biological control of mango dieback disease caused by Lasiodiplodia theobromae using streptomycete and non-streptomycete Actinobacteria in the United Arab Emirates. Front Microbiol 9:829. https://doi.org/10.3389/fmicb.2018.00829
Karunarathna A, Papizadeh M, Senanayake IC, Jeewon R, Phookamsak R, Goonasekara ID, Wanasinghe DN, Wijayawardene NN, Amoozegar MA, Shahzadeh Fazeli SA, Camporesi E, Hyde KD, Weerahewa HLD, Lumyong S, McKenzie EHC (2017) Novel fungal species of Phaeosphaeriaceae with an asexual/sexual morph connection. Mycosphere 8(10):1818–1834
Khalid M, Hassani D, Bilal M, Asad F, Huang D (2017) Influence of bio-fertilizer containing beneficial fungi and rhizospheric bacteria on health promoting compounds and antioxidant activity of Spinacia oleracea L. Bot Stud 58(1):35. https://doi.org/10.1186/s40529-017-0189-3
Khan FK, Kluting K, Tångrot J, Urbina H, Ammunet T, Sahraei SE, Rydén M, Ryberg M, Rosling A (2020) Naming the untouchable–environmental sequences and niche partitioning as taxonomical evidence in fungi. IMA Fungus 11:23. https://doi.org/10.1186/s43008-020-00045-9
Kharwar RN, Gond S, Anuj K, Mishra A (2010) A comparative study of endophytic and epiphytic fungal association with leaf of Eucalyptus citriodora Hook, and their antimicrobial activity. World J Microbiol Biotechnol 26(11):1941–1948
Kim JI, Nam SW, So JE, Hong SG, Choi HG, Shin W (2017) Asterochloris sejongensis sp. nov. (Trebouxiophyceae, Chlorophyta) from King George Island, Antarctica. Phytotaxa 295:60–70
Kirk PM, Cannon PF, Minter DW, Stalpers JA (2008) Ainsworth & Bisby’s dictionary of the fungi, 10th edn. CAB International, Wallingford
Kirschner R (2019) Sex does not sell: the argument for using the terms “anamorph” and “teleomorph” for fungi. Mycol Progr 18:305–312
Kirschner R, Hsu T, Tuan NN, Chen CL, Huang SL (2015) Characterization of fungal and bacterial components in gut/fecal microbiome. Curr Drug Metab 16(4):272–283
Klopfstein S (2016) Revising Australian Pristomerus (Hymenoptera, Ichneumonidae, Cremastinae): Species with a tooth on the hind femur. Zootaxa 4168(2):201–238
Ko TWK, Stephenson SL, Bahkali AH, Hyde KD (2011) From morphology to molecular biology: Can we use sequence data to identify fungal endophytes? Fungal Divers. https://doi.org/10.1007/s13225-011-0130-0
Köhler JR, Casadevall A, Perfect J (2015) The spectrum of fungi that infects humans. Cold Spring Harb Perspect Med 5(1):a019273. https://doi.org/10.1101/cshperspect.a019273
Kohlmeyer J, Kohlmeyer E (1979) Marine mycology—the higher fungi. Academic Press, New York, p 704
Kohlmeyer J, Volkmann-Kohlmeyer B (2001) The biodiversity of fungi on Juncus roemerianus. Mycol Res 105:1409–1412
Konta S, Hyde KD, Phookamsak R, Xu JC, Maharachchikumbura SSN, Daranagama DA, McKenzie EHC, Boonmee S, Tibpromma S, Eungwanichayapant PD, Samarakoon MC, Lu YZ (2020) Polyphyletic genera in Xylariaceae (Xylariales): Neoxylaria gen. nov. and Stilbohypoxylon. Mycosphere 11(1):2629–2651
Koukol O, Delgado G (2021) Why morphology matters: the negative consequences of hasty descriptions of putative novelties in asexual ascomycetes. IMA Fungus 12:26. https://doi.org/10.1186/s43008-021-00073-z
Koukol O, Kolarík M, Kolářová Z, Baldrian P (2012) Diversity of foliar endophytes in wind-fallen Picea abies trees. Fungal Divers 54:69–77
Kozlov AM, Zhang J, Yilmaz P, Glöckner FO, Stamatakis A (2016) Phylogeny-aware identification and correction of taxonomically mislabeled sequences. Nucleic Acids Res 44(11):5022–5033
Kraichak E, Divakar P, Crespo A, Leavitt SD, Nelsen MP, Lücking R, Lumbsch HT (2015) A Tale of Two Hyper-diversities: diversification dynamics of the two largest families of lichenized fungi. Sci Rep 5:10028. https://doi.org/10.1038/srep10028
Krauss G, Sridhar KR, Jung K, Wennrich R, Ehrman JM, Bärlocher F (2003) Aquatic hyphomycetes in polluted groundwater habitats of Central Germany. Microbial Ecol 45:329–339
Kruse J, Choi YJ, Thines M (2017) New smut-specific primers for the ITS barcoding of Ustilaginomycotina. Mycol pro 16:213–221
Kruys ÅN (2005) Phylogenetic relationships and species richness of coprophilous ascomycetes. Department of Ecology and Environmental Science, Umeå University, Umeå
Kubartová A, Ottosson E, Dahlberg A, Stenlid J (2012) Patterns of fungal communities among and within decaying logs, revealed by 454 sequencing. Mol Ecol 21(18):4514–4532
Kumar TKA, Healy R, Spatafora JW, Blackwell M, McLaughlin DJ (2012) Orbilia ultrastructure, character evolution and phylogeny of Pezizomycotina. Mycologia 104:462–476
Kumar A, Chandra R (2020) Ligninolytic enzymes and its mechanisms for degradation of lignocellulosic waste in environment. Heliyon 6 (2): p. e03170
Kutorga E, Rukđënienë J, Treigienë A (2006) Microscopic fungi on Carpinus betulus in Lithuania 1. Teleomorphs and Associated Anamorphs. Bot Lith 12:233–241
Langvad F (1980) A simple and rapid method for qualitative and quantitative study of the fungal flora of leaves. Can J Bot 26:666–670
Lategan MJ, Torpy FR, Newby S, Stephenson S, Hose GC (2012) Fungal diversity of shallow aquifers in Southeastern Australia. Geomicrobiol J 29:352–361
Lavrinienko A, Scholier T, Bates ST, Miller AN, Watts PC (2021) Defining gut mycobiota for wild animals: a need for caution in assigning authentic resident fungal taxa. Anim Microbiome 3(1):75. https://doi.org/10.1186/s42523-021-00134-z
Lawrey JD, Diederich P (2003) Lichenicolous fungi: interactions, evolution, and biodiversity. Bryologist 106(1):80–120
Lazarus KL, Benny GL, Ho HM, Smith ME (2017) Phylogenetic systematics of Syncephalis (Zoopagales, Zoopagomycotina), a genus of ubiquitous mycoparasites. Mycologia 109:333–349
Lee J, Lee T, Lee YW, Yun SH, Turgeon BG (2003) Shifting fungal reproductive mode by manipulation of mating type genes: obligatory heterothallism of Gibberella zeae. Mol Microbiol 50:145–152
Lee NLY, Huang D, Quek ZBR, Lee JN, Wainwright BJ (2019) Mangrove-associated fungal communities are differentiated by geographic location and host structure. Front Microbiol 10: 2456. https://www.frontiersin.org/article/10.3389/fmicb.2019.02456
Lemmon EM, Lemmon AR, Collins JT, Lee-Yaw JA, Cannatella DC (2007) Phylogeny-based delimitation of species boundaries and contact zones in the trilling chorus frogs (Pseudacris). Mol Phylogenet Evol 44:1068–1082
Leray M, Knowlton N, Ho S-L, Machida RJ (2019) GenBank is a reliable resource for 21st century biodiversity research. PNAS 116(45):22651–22656
Li Y, Hyde KD, Jeewon R, Cai L, Vijaykrishna D, Zhang K (2005) Phylogenetics and evolution of nematode-trapping fungi (Orbiliales) estimated from nuclear and protein coding genes. Mycologia 97:1034–1046
Li WJ, McKenzie EHC, Liu JK, Bhat DJ, Dai DQ, Camporesi E, Tian Q, Maharachchikumbura SSN, Luo ZL, Shang QJ, Zhang JF, Tangthirasunun N, Karunarathna SC, Xu JC, Hyde KD (2020) Taxonomy and phylogeny of hyaline-spored coelomycetes. Fungal Divers 100:279–801
Lindahl BD, Nilsson RH, Tedersoo L, Abarenkov K, Carlsen T, Kjøller R, Köljalg U, Pennanen T, Rosendahl S, Stenlid J, Kauserud H (2013) Fungal community analysis by highthroughput sequencing of amplified markers—a user’s guide. New Phytol 199:288–299
Linder CR, Risenberg LH (2004) Reconstructing patterns of reticulate evolution in plants. Am J Bot 91(10):1700–1708
Liu YJ, Hall BD (2004) Body plan evolution of ascomycetes, as inferred from an RNA polymerase II, phylogeny. Proc Natl Acad Sci USA 101:4507–4512
Liu JK, Hyde KD, Jones EBG, Ariyawansa HA, Bhat JD, Boonmee S, Maharachchikumbura SSN, Mckenzie EHC, Phookamsak R, Phukhamsakda C, Shenoy BD, Abdel-Wahab MA, Buyck B, Chen J, Chethana KWT, Singtripop C, Dai DQ, Dai YC, Daranagama DA, Dissanayake AJ, Doilom M, D’souza MJ, Fan XL, Goonasekara ID, Hirayama K, Hongsanan S, Jayasiri SC, Jayawardena RS, Karunarathana SC, Li WJ, Mapook A, Norphanphoun C, Pang KL, Perera RH, Peršoh D, Pinruan U, Senanayake IC, Somrithipol S, Suetrong S, Tanaka K, Thambugala KM, Tian Q, Tibpromma S, Udayanga D, Wuayawardene NN, Wanasinghe D, Wisitrassameewong K, Zeng XY, AbdelAziz FA, Adamčík S, Bahkali AH, Boonyuen N, Bulgakov T, Callac P, Chomnunti P, Greiner K, Hashimoto A, Hofstetter V, Kang JC, Lewis D, Li XL, Liu XX, Liu ZY, Matsumura M, Mortimer PE, Rambold G, Randrianjohany E, Sato G, Sriindrasutdhi V, Tian CM, Verbeken A, Von Brackel W, Wang Y, Wen TC, Xu JC, Yan JY, Zhao RL, Camporesi E (2015) Fungal diversity notes 1–110: taxonomic and phylogenetic contributions to fungal species. Fungal Divers 72:1–197
Liu JK, Hyde KD, Jeewon R, Phillips AJL, Maharachchikumbura SSN, Ryberg M, Liu ZY, Zhao Q (2017) Ranking higher taxa using divergence times: a case study in Dothideomycetes. Fungal Divers 84:75–99
Lodge DJ (1996) Microorganisms. In: Regan DP, Waide RB (eds) The food web of a tropical forest. University of Chicago Press, Chicago, pp 53–108
Lodge DJ, Ammirati J, O’Dell TE, Mueller GM (2004) Collecting and describing macrofungi. In: Mueller GM, Bills GF, Foster MS (eds) Biodiversity of fungi: inventory and monitoring methods. Academic Press, New York, pp 123–168
Lord NS, Kaplan CW, Shank P, Kitts CL, Elrod SL (2002) Assessment of fungal diversity using terminal restriction fragment (TRF) pattern analysis: comparison of 18S and ITS ribosomal regions. FEMS Microbiol Ecol 42:327–337
Lu SW, Kroken S, Lee BN, Robbertse B, Churchill ACL, Yoder OC, Turgeon BG (2003) A novel class of gene controlling virulence in plant pathogenic ascomycete fungi. PNAS 100(10):5980–5985
Lu YZ, Liu JK, Hyde KD, Jeewon R, Kang JC, Fan C, Boonmee S, Bhat DJ, Luo ZL, Lin CG, Eungwanichayapant PD (2018) A taxonomic reassessment of Tubeufiales based on multi-locus phylogeny and morphology. Fungal Divers 92:131–344
Lücking R, Johnston MK, Aptroot A, Kraichak E, Lendemer JC, Boonpragob K, Cáceres MES, Ertz D, Ferraro LI, Jia ZF, Kalb K, Mangold A, Manoch L, Mercado-Díaz JA, Moncada B, Mongkolsuk P, Papong KB, Parnmen S, Peláez RN, Poengsungnoen V, Plata ER, Saipunkaew W, Sipman HJM, Sutjaritturakan J, Broeck DVD, Konrat MV, Weerakoon G, Lumbsch HT (2014) One hundred and seventy-five new species of Graphidaceae: closing the gap or a drop in the bucket? Phytotaxa 189:7–38
Lücking R, Aime MC, Robbertse B, Miller AN, Ariyawansa HA, Aoki T, Schoch CL, Cardinali G, Crous PW, Druzhinina IS, Geiser DM, Hawksworth DL, Hyde KD, Irinyi L, Jeewon R, Johnston PR, Kirk PM, Malosso E, May TW, Meyer W, Öpik M, Robert V, Stadler M, Thines M, Vu D, Yurkov AM, Zhang N (2020) Unambiguous identification of fungi: where do we stand and how accurate and precise is fungal DNA barcoding? IMA Fungus 11(1):14. https://doi.org/10.1186/s43008-020-00033-z
Lughadha EN, Bachman SP, Leão TCC, Forest F, Halley JM, Moat J, Acedo C, Bacon KL, Brewer RFA, Gâteblé G, Gonçalves SC, Govaerts R, Hollingsworth PM, Krisai-Greilhuber I, de Lirio EJ, Moore PGP, Negrão R, Onana JM, Rajaovelona LR, Razanajatovo H, Reich PB, Richards SL, Rivers MC, Cooper A, Iganci J, Lewis GP, Smidt EC, Antonelli A, Mueller GM, Walker BE (2020) Extinction risk and threats to plants and fungi. Plant People Planet 2(5):389–408
Lumbsch HT, Huhndorf SM (2010) Myconet volume 14. Part One Outline of Ascomycota—2009. Fieldiana Life Earth Sci 1:1–64
Lundqvist N (1972) Nordic Sordariaceae s. lat. Symbolae Botanicae Upsalienses, Uppsala
Luo ZL, Hyde KD, Liu JK, Maharachchikumbura SSN, Jeewon R, Bao DF, Bhat DJ, Lin CG, LiWL YJ, Liu NG, Lu YZ, Jayawardena RS, Li JF, Su HY (2019) Freshwater Sordariomycetes. Fungal Divers 99:451–660
Maharachchikumbura SSN, Hyde KD, Groenewald JZ, Xu J, Crous PW (2014) Pestalotiopsis revisited. Stud Mycol 79:121–186
Maharachchikumbura SSN, Hyde KD, Jones EBG, McKenzie EHC, Bhat DJ, Dayarathne MC, Huang SK, Norphanphoun C, Senanayake IC, Perera RH, Shang QJ, Xiao YP, D’souza MJ, Hongsanan S, Jayawardena RS, Daranagama DA, Konta S, Goonasekara ID, Zhuang WY, Jeewon R, Phillips AJL, AbdelWahab MA, Al-Sadi AM, Bahkali AH, Boonmee S, Boonyuen N, Cheewangkoon R, Dissanayake AJ, Kang JC, Li QR, Liu JK, Liu XZ, Liu ZY, Luangsa-ard JJ, Pang KL, Phookamsak R, Promputtha I, Suetrong S, Stadler M, Wen TC, Wijayawardene NN (2016) Families of Sordariomycetes. Fungal Divers 79:1–317
Maharachchikumbura SSN, Chen Y, Ariyawansa HA, Hyde KD, Haelewaters D, Perera RH, Samarakoon MC, Wanasinghe DN, Bustamante DE, Liu JK, Lawrence DP, Cheewangkoon R, Stadler M (2021) Integrative approaches for species delimitation in Ascomycota. Fungal Divers. https://doi.org/10.1007/s13225-021-00486-6
Mallard F, Noble L, Guzella T, Afonso B, Baer CF, Teotónio H (2019) Selection and drift determine phenotypic stasis despite genetic divergence. bioRxiv 778282
Malosso E, Waite IS, English L, Hopkins DW, O’Donnell AG (2006) Fungal diversity in maritime Antarctic soils determined using a combination of culture isolation, molecular fingerprinting and cloning techniques. Polar Biol 29:552–561
Marchese C (2015) Biodiversity hotspots: a shortcut for a more complicated concept. Glob Ecol Conserv 3:297–309
Maria G, Martin G, Ulf S, Daniela Z, Jörg B, Katharina R (2021) The Lichens’ microbiota, still a mystery? Front Microbiol 12:714. https://doi.org/10.3389/fmicb.2021.623839
Martin KJ, Rygiewicz PT (2005) Fungal-specific PCR primers developed for analysis of the ITS region of environmental DNA extracts. BMC Microbiol. https://doi.org/10.1186/1471-2180-5-28
Marx MC, Wood M, Jarvis SC (2001) A microplate fluorimetric assay for the study of enzyme diversity in soils. Soil Biol Biochem 33:1633–1640
Matsuoka S, Sugiyama Y, Sato H, Katano I, Harada K, Doi H (2019) Spatial structure of fungal DNA assemblages revealed with eDNA metabarcoding in a forest river network in western Japan. MBMG 3:37–47
Matutea DR, Sepúlvedab VE (2019) Fungal species boundaries in the genomics era. Fungal Genet Biol 131:103–249
May TW (2017) Report of the Nomenclature Committee for Fungi—21: lists from working groups. IMA Fungus 8(1):205–210. https://doi.org/10.5598/imafungus.2017.08.01.13
Maynard DS, Covey KR, Crowther TW, Sokol NW, Morrison EW, Frey SD, Van Diepen LTA, Bradford MA (2018) Species associations overwhelm abiotic conditions to dictate the structure and function of wood-decay fungal communities. Ecology 99(4):801–811
McNeill J, Barrie FR, Brundet HM, Demoulin V, Hawksworth DJ, Marhold K, Nicolson DH, Prado J, Silva PC, Skog JE, Wiersema JH, Turland NJ (2006) International code of botanical nomenclature (Vienna Code) adopted by the Seventh International Botanical Congress Vienna, Austria, July 2005. A.R.G. Gantner Velag, Ruggell, 568pp
McNeill J, Barrie FR, Buck WR, Demoulin V, Greuter W, Hawksworth DL, Herendeen PS, Knapp S, Marhold K, Prado J, Reine PV, Smith GF, Wiersema JH, Turland NJ (2012) International Code of Nomenclature for algae, fungi and plants (Melbourne Code) adopted by the 18th International Botanical Congress Melbourne, Australia, July 2011. Publ. 2012. Regnum Vegetabile 154. Koeltz Scientific Books. ISBN 978-3-87429-425-6
Melo RFR, Gondim NHDB, Cabral AL, Santiago MA, Maia LC, Miller AN (2020) Coprophilous fungi from Brazil: updated identification keys to all recorded species. Phytotaxa 436(2):104–124
Mendes R, Garbeva P, Raaijmakers JM (2013) The rhizosphere microbiome: significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol Rev 37(5):634–663
Mendonça CO, Aptroot A, Lücking R, Cáceres MES (2020) Global species richness prediction for Pyrenulaceae (Ascomycota: Pyrenulales), the last of the “big three” most speciose tropical microlichen families. Biodivers Conserv 29(3):1059–1079
Meyer W, Irinyi L, Hoang MTV, Robert V, Garcia-Hermoso D, Desnos-Ollivier M, Yurayart C, Tsang CC, Lee CY, Woo PC, Pchelin IM, Uhrlaß S, Nenoff P, Chindamporn A, Chen S, Hebert PD, Sorrell TC (2019) Database establishment for the secondary fungal DNA barcode translational elongation factor 1α (TEF1α). Genome 62(3):160–169
Miadlikowska J, Lutzoni F (2004) Phylogenetic classification of peltigeralean fungi (Peltigerales, Ascomycota) based on ribosomal RNA small and large subunits. Am J Bot 91:449–464
Miadlikowska J, Arnold AE, Lutzoni F (2004) High diversity of cryptic fungi inhabiting healthy lichen thalli in a temperate and tropical forest. Ecol Soc Am Annu Meet 89:349–350
Mitchell JI, Zuccaro A (2006) Sequences, the environment and fungi. Mycologist 20:62–74
Mohamed R, Jong PL, Zali MS (2010) Fungal diversity in wounded stems of Aquilaria malaccensis. Fungal Divers 43:67–74
Molina-Montenegro MA, Galleguillos C, Oses R, Acuña-Rodríguez IS, Lavín P, Gallardo-Cerda J, TorresDíaz C, Diez B, Pizarro GE, Atala C (2016) Adaptive phenotypic plasticity and competitive ability deployed under a climate change scenario may promote the invasion of Poa annua in Antarctica. Biol Invasions 18:603–618
Mora C, Tittensor DP, Adl S, Simpson AGB, Worm B (2011) How many species are there on Earth and in the ocean? PLoS Biol 9:e1001127.
Mueller GM, Schmit JP (2007) Fungal biodiversity: what do we know? What can we predict? Biodiversity Conserv 16:1–5
Muggia L, Grube M (2018) Fungal diversity in lichens: from extremotolerance to interaction with algae. Life 8:15. https://doi.org/10.3390/life8020015
Muggia L, Pérez-Ortega S, Fryday A, Spribille T, Grube M (2014) Global assessment of genetic variation and phenotypic plasticity in the lichen-forming species Tephromela atra. Fungal Divers 64(1):233–251
Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GAB, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858
Nachman MW, Payseur BA (2012) Recombination rate variation and speciation: theoretical predictions and empirical results from rabbits and mice. Philos Trans R Soc Lond B Biol Sci 367:409–421
Naranjo-Ortiz MA, Gabaldón T (2019) Fungal evolution: major ecological adaptations and evolutionary transitions. Biol Rev 94(4):1443–1476
Nash HT (2008) Lichen biology, 2nd edn. Cambridge University Press, Cambridge
Nieuwenhuis BP, James TY (2016) The frequency of sex in fungi. Philos Trans R Soc Lond Ser B Biol Sci 371(1706):20150540
Nilsson RH, Kristiansson E, Ryberg M, Hallenberg N, Larsson KH (2008) Intraspecific ITS variability in the kingdom Fungi as expressed in the international sequence databases and its implications for molecular species identification. Evol Bioinf Online 4:193
Nilsson RH, Ryberg M, Abarenkov K, Sjökvist E, Kristiansson E (2009) The ITS region as a target for characterization of fungal communities using emerging sequencing technologies. FEMS Microbiol Lett 296(1):97–101
Nilsson RH, Tedersoo L, Lindahl BD, Kjøller R, Carlsen T, Quince C, Abarenkov K, Pennanen T, Stenlid J, Bruns T, Larsson KH, Kõljalg U, Kauserud H (2011) Towards standardization of the description and publication of next-generation sequencing datasets of fungal communities. New Phytol 191:314–318
Nilsson RH, Tedersoo L, Abarenkov K, Ryberg M, Kristiansson E, Hartmann M, Schoch CL, Nylander JAA, Bergsten J, Porter TM, Jumpponen A, Vaishampayan P, Ovaskainen O, Hallenberg N, Bengtsson-Palme J, Eriksson KM, Larsson KH, Larsson E, Kõljalg U (2012) Five simple guidelines for establishing basic authenticity and reliability of newly generated fungal ITS sequences. MycoKeys 4:37–63
Nilsson RH, Hyde KD, Pawłowska J, Ryberg M, Tedersoo L, Aas AB, Alias SA, Alves A, Anderson CL, Antonelli A, Arnold AE, Bahnmann B, Bahram M, Bengtsson-Palme J, Berlin A, Branco S, Chomnunti P, Dissanayake A, Drenkhan R, Abarenkov K (2014) Improving ITS sequence data for identification of plant pathogenic fungi. Fungal Divers 67(1):11–19
Nilsson RH, Larsson KH, Taylor AFS, Bengtsson-Palme J, Jeppesen TS, Schigel D, Kennedy P, Picard K, Glöckner FO, Tedersoo L, Saar I, Kõljalg U, Abarenkov K (2019) The UNITE database for molecular identification of fungi: handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res 47:259–264
Nilsson RH, Ryberg M, Kristiansson E, Abarenkov K, Larsson KH, Kõljalg U (2006) Taxonomic reliability of DNA sequences in public sequence databases: A fungal perspective. PLoS One 1(1): Article e59. https://doi.org/10.1371/journal.pone.0000059
Njambere EN, Chen W, Frate C, Wu BM, Temple SR, Muehlbauer FJ (2008) Stem and crown rot of chickpea in California caused by Sclerotinia trifoliorum. Plant Dis 92(6):917–922
Normark BB, Judson OP, Moran NA (2003) Genomic signatures of ancient asexual lineages. Biol J Linn Soc 79:69–84
O’Brien BL, Parrent JL, Jackson JA, Moncalvo JM, Vilgalys R (2005) Fungal community analysis by large-scale sequencing of environmental samples. Appl Environ Microbiol 71:5544–5550
O’Gorman CM, Fuller HT, Dyer PS (2009) Discovery of a sexual cycle in the opportunistic fungal pathogen Aspergillus fumigatus. Nature 457:471–474
Oliveira M, Arenas M, Lage O, Cunha M, Amorim MI (2017) Epiphytic fungal community in Vitis vinifera of the Portuguese wine regions. Lett Appl Microbiol 66:93–102
Osono T (2007) Ecology of ligninolytic fungi associated with leaf litter decomposition. Ecol Res 22:955–974
Osono T (2019) Functional diversity of ligninolytic fungi associated with leaf litter decomposition. Ecol Res 35(3):1–14
Otto SP, Lenormand T (2002) Resolving the paradox of sex and recombination. Nat Rev Genet 3:252–261
Pang KL, Alias SA, Chiang MWL, Vrijmoed LLP, Jones EBG (2010) Sedecimiella taiwanensis gen. et sp. nov., a marine mangrove fungus in the Hypocreales (Hypocreomycetidae, Ascomycota). Bot Mar 53:493–498
Pang KL, Overy DP, Jones EBG, Calado ML, Burgaud G, Walker AK, Johnson JA, Kerr RG, Cha HJ, Bills GF (2016) ‘Marine fungi’ and ‘marine-derived fungi’ in natural product chemistry research: toward a new consensual definition. Fungal Biol Rev 30:163–175
Paoletti M, Rydholm C, Schwier EU, Anderson MJ, Szakacs G, Lutzoni F, Debeaupuis JP, Latgé JP, Denning DW, Dyer PS (2005) Evidence for sexuality in the opportunistic fungal pathogen Aspergillus fumigatus. Curr Biol 15(13):1242–1248
Payseur BA (2010) Using differential introgression in hybrid zones to identify genomic regions involved in speciation. Mol Ecol Resour 10(5):806–820
Pažoutová S, Follert S, Bitzer J, Keck M, Surup F, Šrůtka P, Holuša J, Stadler M (2013) A new endophytic insect-associated Daldinia species, recognised from a comparison of secondary metabolite profiles and molecular phylogeny. Fungal Divers 60:107–123
Peraza-Reyes L, Berteaux-Lecellier V (2013) Peroxisomes and sexual development in fungi. Front Physiol 4:244. https://doi.org/10.3389/fphys.2013.00244
Peršoh D (2015) Plant-associated fungal communities in the light of meta’omics. Fungal Divers 75:1–25
Peršoh D, Melcher M, Graf K, Fournier J, Stadler M, Rambold G (2009) Molecular and morphological evidence for the delimitation of Xylaria hypoxylon. Mycologia 101:256–268
Petrini O (1991) Fungal endophytes of tree leaves. In: Andrews JH, Hirano SS (eds) Microbial ecology of leaves. Springer, New York, pp 179–197
Peyvast GH, Olfati JA, Madeni S, Forghani A (2008) Effect of vermicompost on the growth and yield of spinach (Spinacia oleracea L.). J. Food Agric. Environ. 16(1):110–113
Phookamsak R, Norphanphoun C, Tanaka K, Dai DQ, Luo ZL, Liu JK, Su HY, Bhat DJ, Bahkali AH, Mortimer PE, Xu JC, Hyde KD (2015) Towards a natural classification of Astrosphaeriella-like species; introducing Astrophaeriellaceae and Pseudoastrosphaeriellaceae fam. nov. and Astrosphaeriellopsis, gen. nov. Fungal Divers 74:143–197
Phookamsak R, Wanasinghe DN, Hongsanan S, Phukhamsakda C, Huang SK, Tennakoon DS, Norphanphoun C, Camporesi E, Bulgakov TS, Promputtha I, Mortimer PE, Xu JC, Hyde KD (2017) Towards a natural classification of Ophiobolus and ophiobolus-like taxa; introducing three novel genera Ophiobolopsis, Paraophiobolus and Pseudoophiobolus in Phaeoshaeriaceae (Pleosporales). Fungal Divers 87:299–339
Piepenbring M, Hofmann TA, Kirschner R, Mangelsdorff R, Perdomo O, Rodríguez Justavino D, Trampe T (2011) Diversity patterns of neotropical plant parasitic microfungi. Ecotropica 17:27–40
Piepenbring M, Hofmann TA, Unterseher M, Kost G (2012) Species richness of plants and fungi in western Panama: towards a fungal inventory in the tropics. Biodivers Conserv. https://doi.org/10.1007/s10531-011-0213-y
Pihet M, Carrere J, Cimon B, Chabasse D, Delhaes L, Symoens F, Bouchara JP (2009) Occurrence and relevance of filamentous fungi in respiratory secretions of patients with cystic fibrosis—a review. Med Mycol 47:387–397
Pimm SL, Joppa LN (2015) How many plant species are there, where are they, and at what rate are they going extinct? Ann Mo Bot Gard 100(3):170–176
Possingham HP, Wilson KA (2005) Biodiversity: turning up the heat on hotspots. Nature 436:919–920
Prakash CP, Thirumalai E, Rajulu G, Thirunavukkarasu N, Suryanarayanan TS, Griffith GW (2015) Ecology and diversity of leaf litter fungi during early-stage decomposition in a seasonally dry tropical forest. Fungal Ecol 17:103–113
Price TD, Qvarnström A, Irwin DE (2003) The role of phenotypic plasticity in driving genetic evolution. Proc Biol Sci 270(1523):1433–1440
Promputtha I, Lumyong S, Lumyong P, Ehc M, Hyde KD (2004) Fungal saprobes on dead leaves of Magnolia liliifera (Magnoliaceae) in Thailand. Cryptogamie Mycol 25:315–321
Prosser CM, Hedgpeth BM (2018) Effects of bioturbation on environmental DNA migration through soil media. PLoS ONE 13(4):e0196430. https://doi.org/10.1371/journal.pone.0196430
Pryor BM, Michailides TJ (2002) Morphological, pathogenic, and molecular characterization of Alternaria isolates associated with Alternaria late blight of pistachio. Phytopathology 92(4):406–416
Pugh GJF, Jones EBG (1986) Antarctic marine fungi: a preliminary account. In: Moss ST (ed) The biology of marine fungi. Cambridge Uiniversity, Cambridge, pp 323–225
Pugh GJF (1969) Some problems in the classification of soil fungi. In: Sheals JG (1969) The Soil Ecosystem. A Symposium, Systematics Association Publication No. 8, Academic Press, London 119–130.
Purahong W, Hyde KD (2011) Effects of fungal endophytes on grass and non-grass litter decomposition rates. Fungal Divers 47:1–7
Purahong W, Wubet T, Lentendu G, Schloter M, Pecyna MJ, Kapturska D, Hofrichter M, Krüger D, Buscot F (2016) Life in leaf litter: novel insights into community dynamics of bacteria and fungi during litter decomposition. Mol Ecol 25(16):4059–4074
Purahong W, Pietsch KA, Lentendu G, Schöps R, Bruelheide H, Wirth C, Buscot F, Wubet T (2017) Characterization of unexplored deadwood mycobiome in highly diverse subtropical forests using culture-independent molecular technique. Front Microbiol 8:574. https://doi.org/10.3389/fmicb.2017.00574
Purvis A, Hector A (2000) Getting the measure of biodiversity. Nature 405:212–219
Queiroz K, Donoghue MJ (1988) Phylogenetic systematics and the species problem. Cladistics 4(4):317–338
Raghukumar C, Ravindram J (2012) Fungi and their role in corals and coral reef ecosystems. In: Raghukumar C (ed) Biology of marine fungi. Springer, Berlin, pp 89–113
Raja HA, Miller AN, Pearce CJ, Oberlies NH (2017) Fungal identification using molecular tools: a primer for the natural products research community. J Nat Prod 80(3):756–770
Rajamanikyam M, Vadlapudi V, Amanchy R, Upadhyayula SM (2018) Endophytic fungi as novel resources of natural therapeutics. Braz Arch Biol Technol. https://doi.org/10.1590/1678-4324-2017160542
Rämä T, Quandt CA (2021) Improving fungal cultivability for natural products discovery. Front Microbiol 12:2673. https://doi.org/10.3389/fmicb.2021.706044
Rappaz F (1987) Taxonomy and nomenclature of Diatrypaceae with 8-spored asci. Mycol Helv 2:285–648
Réblová M, Miller AN, Rossman AY, Seifert KA, Crous PW, Hawksworth DL, Beer ZWD (2016) Recommendations for competing sexual-asexually typified generic names in Sordariomycetes (except Diaporthales, Hypocreales, and Magnaporthales). IMA Fungus 7(1):131–153
Réblová M, Kolařík M, Nekvindová J, Miller AN, Hernández-Restrepo M (2021) Phylogeny, global biogeography and Pleomorphism of Zanclospora. Microorganisms 9(4):706. https://doi.org/10.3390/microorganisms9040706
Réblová M, Štěpánek V (2018) Introducing the Rhamphoriaceae, fam. nov. (Sordariomycetes), two new genera, and new life histories for taxa with Phaeoisaria- and Idriella-like anamorphs. Mycologia. 110(4): 750–770.
Reece JB, Urry LA, Cain ML, Wasserman SA, Minorsky PV, Jackson RB (2011) Campbell Biology, 9th ed., Pearson, Boston, Massachusetts, 1309pp.
Restrepo S, Tabima JF, Mideros MF, Grunwald NJ, Matute DR (2014) Speciation in fungal and Oomycete plant pathogens. Annu Rev Phytopathol 52:289–316
Reynolds DR, Taylor JW (1993) The fungal holomorph: mitotic, meiotic and pleomorphic speciation in fungal systematic. CAB International, Wallingford, p 375
Richardson AE (2001a) Prospects for using soil microorganisms to improve the acquisition of phosphorus by plants. Aust J Plant Physiol 28:897–906
Richardson MJ (2001b) Diversity and occurrence of coprophilous fungi. Mycol Res 105:387–402
Richardson DHS (2002) Reflections on lichenology: achievements over the last 40 years and challenges for the future. Can J Bot 80:101–113
Richardson MJ (2019) Coprophilous Ascomycetes. Ascomycete.org 11(6): 205–209
Risse-Buhl U, Herrmann M, Lange P, Akob DM, Pizani N, Schönborn W, Totsche KU, Küsel K (2013) Phagotrophic protest diversity in the groundwater of a karstified aquifer—morphological and molecular analysis. J Eukaryot Microbiol 60:467–479
Robbertse B, Strope PK, Chaverri P, Gazis R, Ciufo S, Domrachev M, Schoch CL (2017) Improving taxonomic accuracy for fungi in public sequence databases: Applying “one name one species” in well-defined genera with Trichoderma/Hypocrea as a test case. Database, Article bax072.
Rodolfi M, Longa CMO, Pertot I, Tosi S, Savino E, Guglielminetti M, Altobelli E, Del Frate G, Picco AM (2016) Fungal biodiversity in the periglacial soil of Dosdè Glacier (Valtellina, Northern Italy). J Basic Microb 56:263–274
Roe AD, Sperling FAH (2007) Population structure and species boundary delimitation of cryptic Dioryctria moths: an integrative approach. Mol Ecol 16(17):3617–3633
Romaní AM, Fischer H, Mille-Lindblom C, Tranvik LJ (2006) Interactions of bacteria and fungi on decomposing litter: differential extracellular enzyme activities. Ecology 87(10):2559–2569
Roskov Y, Ower G, Orrell T, Nicolson D, Bailly N, Kirk PM, Bourgoin T, DeWalt RE, Decock W, Nieukerken E. van, Zarucchi J, Penev L (2019) Species 2000 & ITIS Catalogue of Life, 2019 Annual Checklist. Species 2000: Naturalis, Leiden, the Netherlands. ISSN 2405-884X
Rosling A (2011) Archaeorhizomycetes–a new class of basal fungi. Svensk Mykologisk 32:11–14
Rossman AY, Adams GC, Cannon PF, Castlebury LA, Crous PW, Gryzenhout M, Jaklitsch WM, Mejia LC, Stoykov D, Udayanga D, Voglmayr H, Walker DM (2015a) Recommendations of generic names in Diaporthales competing for protection or use. IMA Fungus 6(1):145–154
Rossman AY, Crous PW, Hyde KD, Hawksworth DL, Aptroot A, Bezerra JL, Bhat JD, Boehm E, Braun U, Boonmee S, Camporesi E, Chomnunti P, Dai DQ, D’souza MJ, Dissanayake A, Gareth Jones EB, Groenewald JZ, Hernández-Restrepo M, Hongsanan S, Jaklitsch WM, Jayawardena R, Jing LW, Kirk PM, Lawrey JD, Mapook A, McKenzie EH, Monkai J, Phillips AJ, Phookamsak R, Raja HA, Seifert KA, Senanayake I, Slippers B, Suetrong S, Taylor JE, Thambugala KM, Tian Q, Tibpromma S, Wanasinghe DN, Wijayawardene NN, Wikee S, Woudenberg JH, Wu HX, Yan J, Yang T, Zhang Y (2015b) Recommended names for pleomorphic genera in Dothideomycetes. IMA Fungus 6(2):507–523
Rossman AY (1994) A strategy for an all-taxa inventory of fungal diversity. In: Peng CI, Chen CH (eds.), Biodiversity and Terrestrial Ecosystems, Monograph Series No. 14, Institute of Botany, Academia Sinica, Taipei 169–194.
Ryberg M, Nilsson RH (2018) New light on names and naming of dark taxa. MycoKeys 23:31–39
Ryberg M, Nilsson RH, Kristiansson E, Töpel M, Jacobsson S, Larsson E (2008) Mining metadata from unidentified ITS sequences in GenBank: a case study in Inocybe (Basidiomycota). BMC Evol Biol 8:50. https://doi.org/10.1186/1471-2148-8-50
Saikkonen K, Saari S, Helander M (2010) Defensive mutualism between plants and endophytic fungi? Fungal Divers 41:101–113
Sainz MJ, González-Jartín JM, Aguín O, Mansilla JP, Botana LM (2018) Isolation, characterization, and identification of mycotoxin-producing fungi. In: Botana LM (ed) Environmental toxicology. De Gruyter, Berlin, Boston, pp 202–245
Samson RA, Visagie CM, Houbraken J, Hong SB, Hubka V, Klaassen CHW, Perrone G, Seifert KA, Susca A, Tanney JB, Varga J, Kocsubé S, Szigeti G, Yaguchi T, Frisvad JC (2014) Phylogeny, identification and nomenclature of the genus Aspergillus. Stud Mycol 78:141–173
Saumell CA, Padilha T (2000) Influence of weather and time of deposition on sheep faeces colonization by nematophagous fungi in the Mata region of Minas Gerais State. Brazilian Appl Soil Ecol 14:63–70
Saumell CA, Padilha T, Santos C, Roque MV (1999) Nematophagous fungi in fresh faeces of cattle in the Mata region of Minas Gerais state. Brazil Vet Parasitol 82(3):217–220
Saumell CA, Padilha T, Santos C (2000) Nematophagous fungi in sheep faeces in Minas Gerais. Brazil Mycol Res 104(8):1005–1008
Schardl CL, Craven KD (2003) Interspecific hybridization in plant-associated fungi and Oomycetes: a review. Mol Ecol 12:2861–2873
Schmit JP, Lodge DJ (2005) Classical methods and modern analysis for studying fungal diversity. In: Schmit JP (ed) Fungal community: its organization and role in the ecosystem. Taylor & Francis, Boca Raton, pp 193–214
Schmit JP, Mueller GM (2007) An estimate of the lower limit of global fungal diversity. Biodivers Conserv 16:99–111
Schmitt I (2011) 8 fruiting body evolution in the ascomycota: a molecular perspective integrating lichenized and non-lichenized groups. In: Pöggeler S, Wöstemeyer J (eds) Evolution of fungi and fungal-like organisms. The Mycota. Springer, Berlin, pp 187–204
Schoch CL, Sung GH, López-Giráldez F, Townsend JP, Miadlikowska J, Hofstetter V, Robbertse B, Matheny PB, Kauff F, Wang Z, Gueidan C (2009) The Ascomycota tree of life: a phylum-wide phylogeny clarifies the origin and evolution of fundamental reproductive and ecological traits. Syst Biol 58(2):224–239
Schoch CL, Seifert KA, Huhndorf S, Robert V, Spouge JL, Levesque CA, Chen W (2012) Fungal Barcoding Consortium: Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc Natl Acad Sci USA 109(16):6241–6246
Schrank FVP (1789) Baierische. Flora 1: 1–753
Schubert K, Groenewald JZ, Braun U, Dijksterhuis J, Starink M, Hill CF, Zalar P, de Hoog G, Crous PW (2007) Biodiversity in the Cladosporium herbarum complex (Davidiellaceae, Capnodiales), with standardisation of methods for Cladosporium taxonomy and diagnostics. Stud Mycol 58:105–156
Seena S, Bärlocher F, Sobral O, Gessner MO, Dudgeon D, McKieh BG, Chauvet E, Boyero L, Ferreiraam V, Frainerno A, Bruderp A, Matthaeip CD, Fenoglio S, Sridharr KR, Albariños RJ, Douglastu MM, Encaladaav AC, Garciau E, Graçaam MAS (2019) Biodiversity of leaf litter fungi in streams along a latitudinal gradient. Sci Total Environ 661:306–315
Seifert KA (2009) Progress towards DNA barcoding of fungi. Mol Ecol Resour 9:83–89
Seifert KA, Rossman AY (2010) How to describe a new fungal species. IMA Fungus 1(2):109–116
Seifert KA, Samuels GJ (2000) How should we look at anamorphs? Stud Mycol 45:5–18
Senanayake IC, Maharachchikumbura SSN, Hyde KD, Bhat JD, Jones EBG, McKenzie EH, Dai DQ, Daranagama DA, Dayarathne MC, Goonasekara ID, Konta S (2015) Towards unravelling relationships in Xylariomycetidae (Sordariomycetes). Fungal Divers 73:73–144
Senanayake IC, Crous PW, Groenewald JZ, Maharachchikumbura SSN, Jeewon R, Phillips AJL, Bhat JD, Perera RH, Li QR, Li WJ, Tangthirasunun N, Norphanphoun C, Karunarathna SC, Camporesi E, Manawasighe I, Al-Sadi AM, Hyde KD (2017) Families of Diaporthales based on morphological and phylogenetic evidence. Stud Mycol 86:217–296
Senanayake IC, Jeewon R, Chomnunti P, Wanasingha DN, Norphanphoun C, Karunarathna A, Pem D, Perera RH, Camporesi E, McKenzie EHC, Hyde KD, Karunarathna SC (2018) Taxonomic circumscription of Diaporthales based on multigene phylogeny and morphology. Fungal Divers 93(1):241–443
Senanayake IC, Rathnayaka AR, Marasinghe DS, Calabon MS, Gentekaki E, Lee HB, Hurdeal VG, Pem D, Dissanayake LS, Wijesinghe SN, Bundhun D, Nguyen TT, Goonasekara ID, Abeywickrama PD, Bhunjun CS, Jayawardena RS, Wanasinghe DN, Jeewon R, Bhat DJ, Xiang MM (2020b) Morphological approaches in studying fungi: collection, examination, isolation, sporulation and preservation. Mycosphere 11(1):2678–2754
Senanayake IC, Al-Sadi AM, Bhat JD, Camporesi E, Dissanayake AJ, Lumyong S, Maharachchikumbura SSN, Hyde KD (2016) Phomatosporales ord. nov. and Phomatosporaceae fam. nov., to accommodate Lanspora, Phomatospora and Tenuimurus, gen. nov. Mycosphere 7(5):628–641
Senanayake IC, Jeewon R, Hyde KD, Bhat JD, Cheewangkoon R (2020) Taxonomy and phylogeny of Leptosillia cordylinea sp. nov. from China. Phytotaxa 435(3):213–226
Shaw KL, Mullen SP (2011) Genes versus phenotypes in the study of speciation. Genetica 139:649–661
Shearer CA, Descals E, Kohlmeyer B, Kohlmeyer J, Marvanova L, Padgett D, Porter D, Raja HA, Schmit JP, Thorton HA, Voglymayr H (2007) Fungal biodiversity in aquatic habitats. Biodivers Conserv 16:49–67
Shearer CA, Raja HA, Miller AN, Nelson P, Tanaka K, Hirayama K, Marvanová L, Hyde KD, Zhang Y (2009) The molecular phylogeny of freshwater Dothideomycetes. Stud Mycol 64:145–153
Shearer CA, Pang KL, Suetrong S, Raja HA (2014) Phylogeny of Dothideomycetes and other classes of freshwater fissitunicate Ascomycota. In: Jones EBG, Hyde KD, Pang KL (eds) Freshwater fungi and fungus-like organisms. De Gruyter, Berlin, Boston, pp 25–46
Shearer CA, Zelski SE, Raja HA, Schmit JP, Miller AN, Janovec JP (2015) Distributional patterns of freshwater ascomycetes communities along an Andes to Amazon elevational gradient in Peru. Biodivers Conserv 24:1877–1897
Shearer CA, Raja HA (2021) Freshwater ascomycetes and their anamorphs. http://fungi.life.illinois.edu. Accessed 22 Jul 2021
Shibata H, Branquinho C, McDowell WH, Mitchell MJ, Monteith DT, Tang J, Arvola L, Cruz C, Cusack DF, Halada L, Kopáček J, Máguas C, Sajidu S, Schubert H, Tokuchi N, Záhora J (2015) Consequence of altered nitrogen cycles in the coupled human and ecological system under changing climate: the need for long-term and site-based research. Ambio 44:178–193
Silberfeld T, Leigh JW, Verbruggen H, Cruaud C, Reviers B, Rousseau F (2010) A multi-locus time-calibrated phylogeny of the brown algae (Heterokonta, Ochrophyta, Phaeophyceae): Investigation the evolutionary nature of the “brown algal crown radiation.” Mol Phylogenet Evol 56:659–674
Singh R, Dubey AK (2015) Endophytic actinomycetes as emerging source for therapeutic compounds. Indo Global J Pharm Sci 5:106–116
Singtripop C, Hongsanan S, Li J, Silva NID, Phillips AJL, Jones GEB, Bahkali A, Hyde KD (2016) Chaetothyrina mangiferae sp. nov., a new species of Chaetothyrina. Phytotaxa 255:21–33
Sinsabaugh RL, Carreiro MM, Repert DA (2002) Allocation of extracellular enzymatic activity in relation to litter composition, deposition, and mass loss. Biogeochemistry 60:1–24
Smith M, Jaffee B (2009) PCR primers with enhanced specificity for nematode-trapping fungi (Orbiliales). Microb Ecol 58:117–128
Song X, Li Y, Hu Y, Guo W, Wu Z, Zhang Y, Cao Z (2021) Endophytes from blueberry roots and their antifungal activity and plant growth enhancement effects. Rhizosphere. https://doi.org/10.1016/j.rhisph.2021.100454
Splivallo R, Ottonello S, Mello A, Karlovsky P (2010) Truffle Volatiles: from chemical ecology to aroma biosynthesis. New Phytol 198(3):688–699
Stadler M, Læssøe T, Fournier J, Decock C, Schmieschek B, Tichy HVPD (2014) A polyphasic taxonomy of Daldinia (Xylariaceae). Stud Mycol 77:1–143
Stearns SC, Hoekstra RJ (2005) Evolution. 2ndEd. Oxford University Press, New York.
Stewart EJ (2012) Growing unculturable bacteria. J Bacteriol 194:4151–4160
Stielow JB, Lévesque CA, Seifert KA, Meyer W, Iriny L, Smits D, Robert V (2015) One fungus, which genes? Development and assessment of universal primers for potential secondary fungal DNA barcodes. Persoonia 35:242–263
Stork NE (1988) Insect diversity—facts, fiction and speculation. Biol J Linn Soc 35:321–337
Stork NE (2018) How many species of insects and other terrestrial arthropods are there on earth? Annu Rev Entomol 63:31–45
Stork NE, McBroom J, Gely C, Hamilton AJ (2015) New approaches narrow global species estimates for beetles, insects, and terrestrial arthropods. PNAS 112:7519–7523
Straatsma G, Ayer F, Egli S (2001) Species richness, abundance, and phenology of fungal fruit bodies over 21 years in a Swiss forest plot. Mycol Res 105(5):515–523
Strullu-Derrien C, Selosse MA, Kenrick P, Martin FM (2018) The origin and evolution of mycorrhizal symbioses: from palaeomycology to phylogenomics. New Phytol 220:1012–1030
Stukenbrock EH (2013) Evolution, selection and isolation: A genomic view of speciation in fungal plant pathogens. New Phytol 199:895–907
Stukenbrock EH (2016) The role of hybridization in the evolution and emergence of new fungal pathogens. Phytopathology 106:104–112
Su YY, Guo LD, Hyde KD (2010) Response of endophytic fungi of Stipa grandis to experimental plant function group removal in Inner Mongolia steppe. China Fungal Divers 43:93–101
Subramanian CV (1986) The progress and status of mycology in India. Proc Plant Sci 96:379–392
Suh SO, Noda H, Blackwell M (2001) Insect symbiosis: derivation of yeast-like endosymbionts within an entomophathogenic filamentous lineage. Mol Biol Evol 18:995–1000
Sullivan DJ, Berman J, Myers LC, Moran GP (2015) Telomeric ORFS in Candida albicans: does mediator tail wag the yeast? PLoS Pathog 11(2):e1004614. https://doi.org/10.1371/journal.ppat.1004614
Sun X, Guo LD (2012) Endophytic fungal diversity: review of traditional and molecular techniques. Mycology 3(1):65–76
Sun S, Heitman J (2011) Is sex necessary? BMC Biol 9:56. https://doi.org/10.1186/1741-7007-9-56
Sun X, Guo LD, Hyde KD (2011) Community composition of endophytic fungi in Acer truncatum and their role in decomposition. Fungal Divers 47:85–95
Sun JZ, Liu XZ, McKenzie EHC, Jeewon R, Liu JK, Zhang XL, Zhao Q, Hyde KD (2019) Correction to: Fungicolous fungi: terminology, diversity, distribution, evolution, and species checklist. Fungal Divers 95:431–432
Sung GH, Sung JM, Hywel-Jones NL, Spatafora JW (2007) A multi-gene phylogeny of Clavicipitaceae (Ascomycota, Fungi): Identification of localized incongruence using a combinational bootstrap approach. Mol Phylogenet Evol 44(3):1204–1223
Szink I, Davis EL, Ricks KD, Koide RT (2016) New evidence for broad trophic status of leaf endophytic fungi of Quercus gambelii. Fungal Ecol 22:2–9
Taberlet P, Prud’Homme SM, Campione E, Roy J, Miquel C, Shehzad W, Gielly L, Rioux D, Choler P, Clément JC, Melodelima C, Pompanon F, Coissac E (2012) Soil sampling and isolation of extracellular DNA from large amount of starting material suitable for metabarcoding studies. Mol Ecol 21:1816–1820
Talgø V, Chastagner G, Thomsen IM, Cech T, Riley K, Lange K, Klemsdal SS, Stensvand A (2010) Sydowia polyspora associated with current season needle necrosis (CSNN) on true fir (Abies spp). Fungal Biol 114(7):545–554
Tanney JB, Seifert KA (2018) Phacidiaceae endophytes of Picea rubens in Eastern Canada. Botany 96(9):555–588
Tarman K, Palm GJ, Porzel A, Merzweiler K, Arnold N, Wessjohann LA, Unterseher M, Lindequist U (2012) Helicascolide C, a new lactone from an Indonesian marine algicolous strain of Daldinia eschscholzii (Xylariaceae, Ascomycota). Phytochem Lett 5:83–86
Taylor DL, Sinsabaugh R (2015) The soil fungi: occurrence, phylogeny and ecology. In: Paul E (ed) Soil microbiology, ecology and biochemistry. Elsevier Inc, New York, p 604
Taylor DL, Herriott IC, Stone KE, McFarland JW, Booth MG, Leigh MB (2010) Structure and resilience of fungal communities in Alaskan boreal forest soils. Can J Res 40:1288–1301
Taylor DL, Hollingsworth TN, McFarland JW, Lennon NJ, Nusbaum C, Ruess RW (2014) A first comprehensive census of fungi in soil reveals both hyperdiversity and fine-scale niche partitioning. Ecol Monogr 84:3–20
Taylor JW, Göker M, Pitt JI (2016) Choosing one name for pleomorphic fungi: the example of Aspergillus versus Eurotium, Neosartorya and Emericella. Taxon 65(3):593–601
Taylor JW, Spatafora J, Berbee M (2006) Ascomycota. Available at http://tolweb.org/Ascomycota. Accessed 9 Dec 2020
Tedersoo L, Smith ME (2017) Ectomycorrhizal fungal lineages: detection of four new groups and notes on consistent recognition of ectomycorrhizal taxa in high-throughput sequencing studies. In: Tedersoo L (ed) Biogeography of mycorrhizal symbiosis. Springer, Cham, pp 125–142
Tedersoo L, May TW, Smith ME (2010) Ectomycorrhizal lifestyle in fungi: global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhiza 20:217–263
Tedersoo L, Bahram M, Puusepp R, Nilsson RH, James TY (2017) Novel soil-inhabiting clades fill gaps in the fungal tree of life. Microbiome 5:42
Tedersoo L, Bahram M, Põlme S, Kõljalg U, Yorou N, Wijesundera R, Abarenkov K (2014) Fungal biogeography, global diversity and geography of soil fungi. Science (New York, NY). https://doi.org/10.1126/science.1256688
Tehler A, Irestedt M (2007) Parallel evolution of lichen growth forms in the family Roccellaceae (Arthoniales, Ascomycota). Cladistics 23(5):432–454
Tejesvi MV, Kajula M, Mattila S, Pirttilä AM (2011) Bioactivity and genetic diversity of endophytic fungi in Rhododendron tomentosum Harmaja. Fungal Divers 47:97–107
Terhonen E, Blumenstein K, Kovalchuk A, Asiegbu FO (2019) Forest tree microbiomes and associated fungal endophytes: functional roles and impact on forest health. Forests 10:42. https://doi.org/10.3390/f10010042
Thines M, Crous PW, Aime MC, Aoki T, Cai L, Hyde KD, Miller AN, Zhang N, Stadler M (2018) Ten reasons why a sequencebased nomenclature is not useful for fungi anytime soon. IMA Fungus 9:177–183
Thomas DC, Vandegrift R, Ludden A, Carroll GC, Roy BA (2016) Spatial ecology of the fungal genus Xylaria in a tropical cloud forest. Biotropica 48:381–393
Tibell S, Tibell L, Pang K-L, Jones EBG (2020) A conspectus of the filamentous marine fungi of Sweden. Bot Mar 63(2):141–153
Todd RB, Zhou M, Ohm RA, Leeggangers HACF, Visser L, de Vries RP (2014) Prevalence of transcription factors in ascomycete and basidiomycete fungi. BMC Genomics 15:214. https://doi.org/10.1186/1471-2164-15-214
Torres-Montúfar A, Borsch T, Ochoterena H (2018) When homoplasy is not homoplasy: Dissecting trait evolution by contrasting composite and reductive coding. Syst Biol 67(3):543–551
Torta L, Lo Piccolo S, Piazza G, Burruano S, Colombo P, Ottonello D, Perrone R, Di Maida G, Pirrotta M, Tomasello A, Calvo S (2015) Lulwoana sp, a dark septate endophyte in roots of Posidonia oceanica (L.). Delile Seagrass 17(2):505–511
Tretter ED, Johnson EM, Benny GL, Lichtwardt RW, Wang Y, Kandel P, Novak SJ, Smith JF, White MM (2014) An eight-gene molecular phylogeny of the Kickxellomycotina, including the first phylogenetic placement of Asel-lariales. Mycologia 106:912–935
Tripathi M, Joshi Y (2019) Endolichenic fungi: present and future trends. 1st edn., Springer Singapore, 180pp.
Trouillas FP, Úrbez-Torres JR, Gubler WD (2010) Diversity of diatrypaceous fungi associated with grapevine canker diseases in California. Mycologia 102:319–336
Tsui CKM, Baschien C, Goh TK (2016) Biology and ecology of freshwater fungi. In: Li DW (ed) Biology of microfungi. Springer, New York, pp 284–314
Turland NJ, Wiersema JH, Barrie FR, Greuter W, Hawksworth DL, Herendeen PS, Knapp S, Kusber WH, Li DZ, Marhold K, May TW, McNeill J, Monro AM, Prado J, Price MJ, Smith GF (2018) International Code of Nomenclature for algae, fungi, and plants (Shenzhen Code) adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017. Koeltz Botanical Books
Valladares F, Gianoli E, Gómez JM (2007) Ecological limits to plant phenotypic plasticity. New Phytol 176(4):749–763
Van der Heijden MGA, Bardgett RD, van Straalen NM (2008) The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol Lett 11:296–310
Vartoukian S, Palmer RM, Wade WG (2010) Strategies for culture of ‘unculturable’ bacteria. FEMS Microbiol Ecol 309:1–7
Vega FE, Dowd PF (2005) The role of yeasts as insect endosymbionts. In: Vega FE, Blackwell M (eds) Insect Fungal associations: ecology and evolution. Oxford University Press, New York, pp 211–243
Vega FE, Meyling N, Luangsa-ard J, BlackwelL M (2012) Fungal Entomopathogens. Insect Pathol 2(4):171–220
Vega FE, Blackwell M (2005) Insect-Fungal Associations: Ecology and Evolution, Oxford University Press, 1st edition 352pp.
Vellinga EC, Kuyper TW, Ammirati J, Desjardin DE, Halling RE, Justo A, Læssøe T, Lebel T, Lodge DJ, Matheny PB, Methven AS, Moreau PA, Mueller GM, Noordeloos ME, Nuytinck J, Ovrebo CL, Verbeken A (2015) Six simple guidelines for introducing new genera of fungi. IMA Fungus 6(2):65–68
Venter JC, Remington K, Heidelberg JF, Halpern AL, Rusch D, Eisen JA, Wu DY, Paulsen I, Nelson KE, Nelson W, Fouts DE, Levy S, Knap AH, Lomas MW, Nealson K, White O, Peterson J, Hoffman J, Parsons R, Baden-Tillson H, Pfannkoch C, Rogers YH, Smith HO (2004) Environmental genome shotgun sequencing of the Sargasso Sea. Science 304:66–74
Větrovský T, Kohout P, Kopecký M, Machac A, Man M, Bahnmann BD, Brabcová V, Choi J, Meszárošová L, Human ZR, Lepinay C, Lladó S, López-Mondéjar R, Martinović T, Mašínová T, Morais D, Navrátilová D, Odriozola I, Štursová M, Švec K, Tláskal V, Urbanová M, Wan J, Žifčáková L, Howe A, Ladau J, Peay KG, Storch D, Wild J, Baldrian P (2019) A meta-analysis of global fungal distribution reveals climate-driven patterns. Nat Commun 10:5142
Vijaykrishna D, Jeewon R, Hyde KD (2006) Molecular taxonomy, origins and evolution of freshwater ascomycetes. Fungal Divers 23:351–390
Vleugels T, de Riek J, Heungens K, van Bockstaele E, Baert J (2012) Genetic diversity and population structure of Sclerotinia species from European red clover crops. J Plant Pathol 94(3):493–503
Voglmayr H, Friebes G, Gardiennet A, Jaklitsch WM (2018) Barrmaelia and Entosordaria in Barrmaeliaceae (fam. nov., Xylariales) and critical notes on Anthostomella-like genera based on multigene phylogenies. Mycol Progress 17:155–177
Voglmayr H, Fournier J, Jaklitsch WM (2019) Two new classes of Ascomycota: Xylobotryomycetes and Candelariomycetes. Pers.: Mol. Phylogeny Evol Fungi 42:36–49
Volk TJ (2013) Fungi. Encyclopedia Biodivers 3(624):640
Voříšková J, Baldrian P (2013) Fungal community on decomposing leaf litter undergoes rapid successional changes. ISME J 7(3):477–486
Wagner CE, Keller I, Wittwer S, Selz OM, Mwaiko S, Greuter L, Sivasundar A, Seehausen O (2013) Genome-wide RAD sequence data provide unprecedented resolution of species boundaries and relationships in the Lake Victoria cichlid adaptive radiation. Mol Ecol 22:787–798
Walker AK, Campbell J (2010) Marine fungal diversity: a comparison of natural and created salt marshes of the north-central Gulf of Mexico. Mycologia 102:513–521
Walker AK, Robicheau BM (2021) Fungal diversity and community structure from Coastal and barrier island beaches in the United States Gulf of Mexico. Nature Portfolio 11:3889
Walker AK, Velez P, González MC (2017) Marine fungi. eLS 1–6.
Wallen RM, Michael HP (2018) An Overview of the function and maintenance of sexual reproduction in dikaryotic fungi. Front Microbiol 9:503. https://doi.org/10.3389/fmicb.2018.00503
Wanasinghe DN, Hyde KD, Jeewon R, Crous PW, Wijayawardene NN, Jones EBG, Bhat DJ, Phillips AJL, Groenewald JZ, Dayarathne MC, Phukhamsakda C, Thambugala KM, Bulgakov TS, Camporesi E, Gafforov YS, Mortimer PE, Karunarathna SC (2017) Phylogenetic revision of Camarosporium (Pleosporineae, Dothideomycetes) and allied genera. Stud Mycol 87:207–256
Wanasinghe DN, Phukhamsakda C, Hyde KD, Jeewon R, Lee HB, Jones EBG, Tibpromma S, Tennakoon DS, Dissanayake AJ, Jayasiri SC, Gafforov Y, Camporesi E, Bulgakov TS, Ekanayake AH, Perera RH, Samarakoon MC, Goonasekara ID, Mapook A, Li WJ, Indunil C, Senanayake IC, Li JF, Norphanphoun C, Doilom M, Bahkali AH, Xu JC, Mortimer PE, Tibell L, Tibell S, Karunarathna SC (2018) Fungal diversity notes 709–839: taxonomic and phylogenetic contributions to fungal taxa with an emphasis on fungi on Rosaceae. Fungal Divers 89:1–238
Wang DYC, Kumar S, Hedges SB (1999) Divergence time estimates for the early history of animal phyla and the origin of plants, animals and fungi. Proc R Soc Lond b 266(1415):163–171
Wang Z, Nilsson RH, James TY, Dai Y, Townsend JP (2016) Future perspectives and challenges of fungal systematics in the age of big data. In: Li DW (ed) Biology of microfungi. Springer, Cham, pp 25–46
Wang MM, Chen Q, Diao YZ, Duan WJ, Cai L (2019) Fusarium incarnatum-equiseti complex from China. Persoonia 43:70–89
Wardle DA, Lindahl BD (2014) Disentangling global soil fungal diversity. Science 346:1052–1053
Watanabe T (1994) Pictorial atlas of soil and seed fungi. Lewis Publishers, Boca Raton, p 504
Watkinson S, Boddy L, Money N (2015) The Fungi, 3rd Edn, Academic Press, 466pp.
Weber CF, Zak DR, Hungate BA, Jackson RB, Vilgalys R, Evans RD, Schadt CW, Megonigal JP, Kuske CR (2011) Responses of soil cellulolytic fungal communities to elevated atmospheric CO2 are complex and variable across five ecosystems. Environ Microbiol 13:2778–2793
Weber B, Büdel B (2011) Fungi and lichens. In: Encyclopedia of Earth Sciences Series; Springer Amsterdam, The Netherlands, 401–409.
Webster J (1970) Coprophilous fungi. Trans Br Mycol Soc 54:161–180
Wehmeyer LE (1941) A Revision of Melanconis, Pseudovalsa, Prosthecium, and Titania. Univ Mich Stud Sci Ser 14:1–161
Wei Y, Liu W, Hu W, Yan Y, Shi H (2020) The chaperone MeHSP90 recruits MeWRKY20 and MeCatalase1 to regulate drought stress resistance in cassava. New Phytol 226(2):476–491
Wendt L, Benjamin ES, Kuhnert E, Heitkämper S, Lambert C, Hladki AI, Romero AI, Luangsa-ard JJ, Srikitikulchai P, Peršoh D, Stadler M (2018) Resurrection and emendation of the Hypoxylaceae, recognised from a multigene phylogeny of the Xylariales. Mycol Prog 17:115–154
White JF, Cole GT (1986) Endophyte-host associations in forage grasses. IV. The Endophyte of Festuca Versuta. Mycologia 78:102–107
White T, Bruns T, Lee S, Taylor J, Innis M, Gelfand D, Sninsky J (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ (eds) PCR protocols: a guide to methods and applications. Academic Press, New York, pp 315–322
Wicklow DT (1992) The coprophilous fungal community: an experimental system. In: Carrol GC, Wicklow DT (eds) The fungal community, its organization and role in the ecosystem. Marcel Dekker, New York, pp 715–728
Wijayawardene NN, Mckenzie EHC, Hyde KD (2012) Towards incorporating anamorphic fungi in a natural classificationchecklist and notes for 2011. Mycosphere 3:157–228
Wijayawardene NN, Hyde KD, Rajeshkumar KC, Hawksworth DL, Madrid H, Kirk PM, Braun U, Singh RV, Crous PW, Kukwa M, Lücking R, Kurtzman CP, Yurkov A, Haelewaters D, Aptroot A, Lumbsch HT, Timdal E, Ertz D, Etayo J, Phillips AJL, Groenewald JZ, Papizadeh M, Selbmann L, Dayarathne MC, Weerakoon G, Jones EBG, Suetrong S, Tian Q, Castañeda-Ruiz RF, Bahkali AH, Pang KL, Tanaka K, Dai DQ, Sakayaroj J, Hujslová M, Lombard L, Shenoy BD, Suija A, Maharachchikumbura SSN, Thambugala KM, Wanasinghe DN, Sharma BO, Gaikwad S, Pandit G, Zucconi L, Onofri S, Egidi E, Raja HA, Kodsueb R, Cáceres MES, Pérez-Ortega S, Fiuza PO, Monteiro JS, Vasilyeva LN, Shivas RG, Prieto M, Wedin M, Olariaga I, Lateef AA, Agrawal Y, Fazeli SAS, Amoozegar MA, Zhao GZ, Pfliegler WP, Sharma G, Oset M, Abdel-Wahab MA, Takamatsu S, Bensch K, de Silva NI, De Kesel A, Karunarathna A, Boonmee S, Pfister DH, Lu YZ, Luo ZL, Boonyuen N, Daranagama DA, Senanayake IC, Jayasiri SC, Samarakoon MC, Zeng XY, Doilom M, Quijada L, Rampadarath S, Heredia G, Dissanayake AJ, Jayawardana RS, Perera RH, Tang LZ, Phukhamsakda C, Hernández-Restrepo M, Ma X, Tibpromma S, Gusmao LFP, Weerahewa D, Karunarathna SC (2017) Notes for Genera-Ascomycota. Fungal Divers 86:1–594
Wijayawardene NN, Hyde KD, Lumbsch HT, Liu JK, Maharachchikumbura SSN, Ekanayaka AH, Tian Q, Phookamsak R (2018) Outline of Ascomycota: 2017. Fungal Divers 88:167–263
Williams SE, Shoo LP, Isaac JL, Hoffmann AA, Langham G, Moritz C (2008) Towards an integrated framework for assessing the vulnerability of species to climate change. PLoS Biol 6(12):2621–2626
Williamson PM, Sivasithamparam K, Cowling WA (1991) Formation of subcuticular coralloid hyphae by Phomopsis leptostromiformis. Plant Dis 75:1023–1026
Williamson PM, Highet AS, Gams W, Sivasithamparam K, Cowling WA (1994) Diaporthe toxica sp nov, the cause of lupinosis in sheep. Mycol Res 98(12):1364–1368
Willis KJ (2018) State of the World’s Fungi 2018, Royal Botanic Gardens, Kew.
Wilson AM, Wilken PM, van der Nest MA, Wingfield MJ, Wingfield BD (2019) It’s All in the Genes: the regulatory pathways of sexual reproduction in filamentous ascomycetes. Genes 10(5):330
Wingfield MJ, De Beer ZW, Slippers B (2012) One fungus, one name promotes progressive plant pathology. Mol Plant Pathol 13:604–613
Wu HX, Schoch CL, Boonmee S, Bahkali AH, Chomnunti P, Hyde KD (2011) A Reappraisal of Microthyriaceae. Fungal Divers 51:189–248
Wu B, Tian J, Bai C, Xiang M, Sun J, Liu X (2013) The biogeography of fungal communities in wetland sediments along the Chang jiang river and other sites in China. ISME J 7:1299–1309
Wu B, Hussain M, Zhang W, Stadler M, Liu X, Xiang M (2019a) Current insights into fungal species diversity and perspective on naming the environmental DNA sequences of fungi. Mycology 10(3):127–140
Wu MS, Aquino LBB, Barbaza MYU, Hsieh CL, Castro-Cruz KA, Yang LL, Tsai PW (2019) Anti-inflammatory and anticancer properties of bioactive compounds from Sesamum indicum L.—A review. Molecules 24(24):4426. https://doi.org/10.3390/molecules24244426
Xu J, Ebada SS, Proksch P (2010) Pestalotiopsis a highly creative genus: chemistry and bioactivity of secondary metabolites. Fungal Divers 44:15–31
Xu W, Gong LF, Pang KL, Luo ZH (2018) Fungal diversity in deep-sea sediments of a hydrothermal vent system in the Southwest Indian Ridge. Deep-Sea Res 131:16–26
Yang ZL (2011) Molecular techniques revolutionize knowledge of basidiomycete evolution. Fungal Divers 50:47–58
Yang X, Feng P, Yin Y, Bushley K, Spatafora JW, Wang C (2018) Cyclosporine biosynthesis in Tolypocladium inflatum benefits fungal adaptation to the environment. mBio. https://doi.org/10.1128/mBio.01211-18
Zachow C, Berg C, Mueller H, Meincke R, Komon-Zelazowska M, Druzhinina IS, Kubicek CP, Berg G (2009) Fungal diversity in the rhizosphere of endemic plant species of Tenerife (Canary Islands): relationship to vegetation zones and environmental factors. ISME J 3:79–92
Zamora JC, Svensson M, Kirschner R, Olariaga I et al (2018) Considerations and consequences of allowing DNA sequence data as types of fungal taxa. IMA Fungus 9:167–175
Zhang YL, Zhang J, Jiang N, Lu YH, Wang L, Xu SH, Wang W, Zhang GF, Xu Q, Ge HM (2011) Immunosuppressive polyketides from mantis-associated Daldinia eschscholzii. J Am Chem Soc 133:5931–5940
Zhang T, Wei XL, Zhang YQ, Liu HY, Yu LY (2015) Diversity and distribution of lichen-associated fungi in the Ny-Alesund Region (Svalbard, High Arctic) as revealed by 454 pyrosequencing. Sci Rep 5:148–150
Zhang Y, Wu Y, Zhang Y, Ozcan A (2016) Color calibration and fusion of lens-free and mobile-phone microscopy images for high-resolution and accurate color reproduction. Sci Rep 6:27811
Zhang X, Zhou Y, Li Y, Fu X, Wang Q (2017) Screening and characterization of endophytic Bacillus for biocontrol of grapevine downy mildew. Crop Prot 96:173–179
Zhang N, Li Y, Wubet T, Bruelheide H, Liang Y, Purahong W, Buscot F, Ma K (2018) Tree species richness and fungi in freshly fallen leaf litter: unique patterns of fungal species composition and their implications for enzymatic decomposition. Soil Biol Biochem 127:120–126
Zhang YJ, Zhang S, Wang M, Bai FY, Liu XZ (2010) High diversity of the fungal community structure in naturally occurring Ophiocordyceps sinensis. PLoS ONE 5:e15570
Zin NA, Badaluddin NA (2020) Biological functions of Trichoderma spp for agriculture applications. Ann Agric Sci 65(2):168–178
Acknowledgements
Ning Xie thanks National Key R&D Program of China (2021YFA0910800), National Natural Science Foundation of China (No. 31601014), Basic and applied basic research fund of Guangdong Province (2121A1515012166), Stability Support Project for Universities in Shenzhen (20200812173625001) and Project of DEGP (2019KTSCX150) for funding. Senanayake thanks to Paul Kirk, Samantha C. Karunarathna for data contribution. S.N. Wijesinghe would like to acknowledge Thailand Science Research and Innovation (TSRI) grant for Macrofungi diversity research from the Lancang-Mekong Watershed and Surrounding areas (Grant No. DBG6280009). Dhanushka Wanasinghe thanks the CAS President’s International Fellowship Initiative (PIFI) for funding his postdoctoral research (number 2021FYB0005), the Postdoctoral Fund from Human Resources and Social Security Bureau of Yunnan Province and the National Science Foundation of China. Saowaluck Tibpromma would like to thank the International Postdoctoral Exchange Fellowship Program (Number Y9180822S1), CAS President’s International Fellowship Initiative (PIFI) (Number 2020PC0009), China Postdoctoral Science Foundation and the Yunnan Human Resources, and Social Security Department Foundation for funding her postdoctoral research. Rungtiwa Phookamsak thanks to CAS President’s International Fellowship Initiative (PIFI) for young staff (Grant No. 2019FYC0003) and “High-level Talent Support Plan” Young Top Talent Special Project of Yunnan Province.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that there is no confict of interest.
Additional information
Handling Editor: Antonio Roberto Gomes de Farias
The original version of this article has been revised: In the section Biodiversity hotspots, the wording of the origin of most ascomycetous type collections has been corrected.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Senanayake, I.C., Pem, D., Rathnayaka, A.R. et al. Predicting global numbers of teleomorphic ascomycetes. Fungal Diversity 114, 237–278 (2022). https://doi.org/10.1007/s13225-022-00498-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13225-022-00498-w