
High Throughput Screening of the NatureBank ‘Marine Collection’ in a Haemonchus Bioassay Identifies Anthelmintic Activity in Extracts from a Range of Sponges from Australian Waters

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results and Discussion
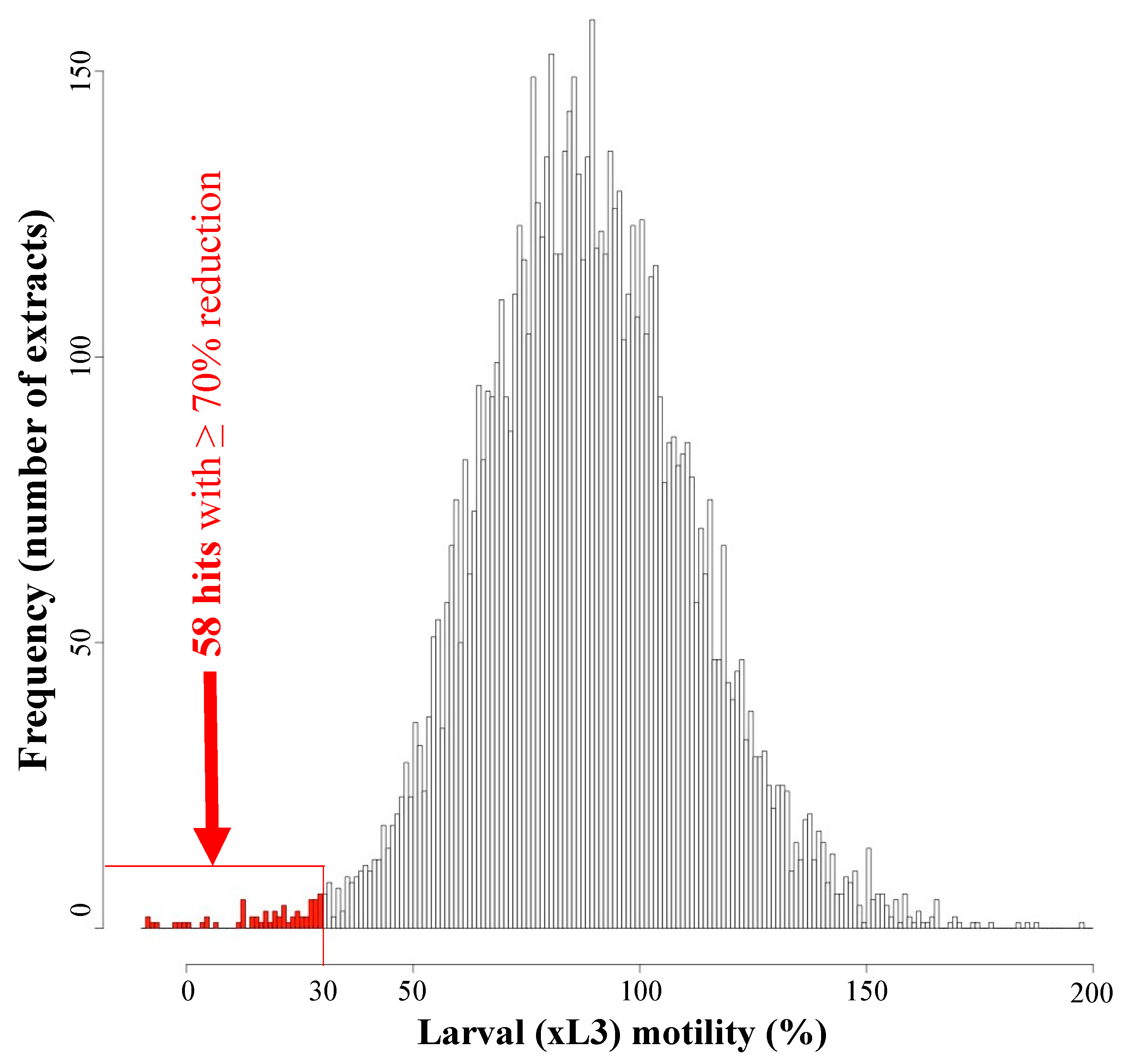
2.1. High Throughput Screening Reveals 58 of 7616 Extracts with Activity on H. contortus
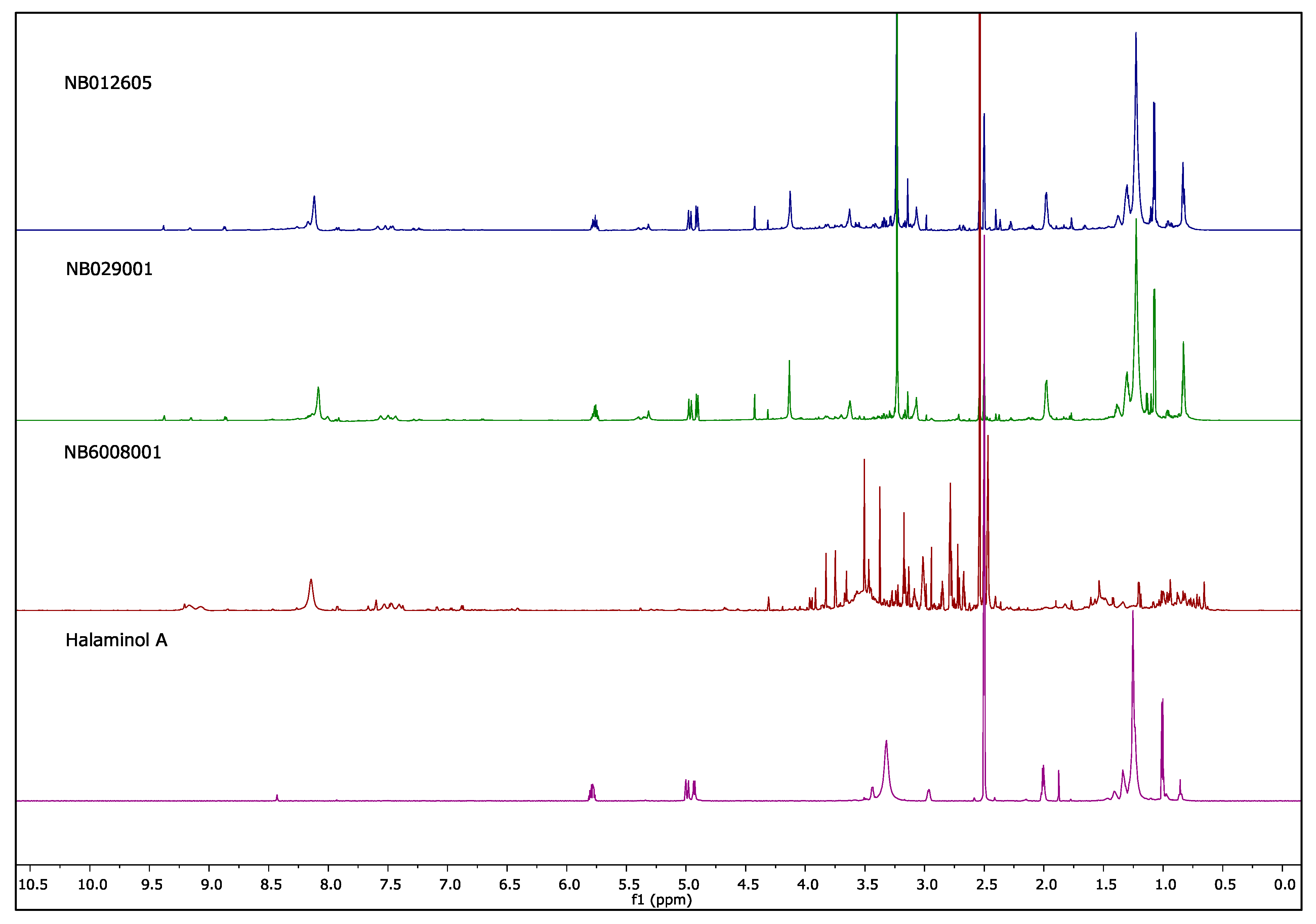
2.2. NMR Fingerprints of Extracts
3. Materials and Methods
3.1. Marine Extract Collection
3.2. Preparation of Haemonchus contortus Larvae
3.3. Screening of Extracts for Larval Motility Reduction (90 h) and Developmental/Morphological Alterations (168 h)
3.4. 1H NMR Fingerprinting of Hit Extracts
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roeber, F.; Jex, A.R.; Gasser, R.B. Impact of gastrointestinal parasitic nematodes of sheep, and the role of advanced molecular tools for exploring epidemiology and drug resistance—an Australian perspective. Parasit. Vectors 2013, 6, 153. [Google Scholar] [CrossRef] [Green Version]
- Beveridge, I.; Emery, D. Australian Animal Parasites—Inside and Out; The Australian Society for Parasitology Inc., 2014; ISBN 9780646935607. Available online: https://www.parasite.org.au/publications/australasian-animal-parasites-inside-and-out/.
- Kotze, A.C.; Prichard, R.K. Anthelmintic resistance in Haemonchus contortus: History, mechanisms and diagnosis. Adv. Parasitol. 2016, 93, 397–428. [Google Scholar]
- Besier, R.B.; Kahn, L.P.; Sargison, N.D.; Van Wyk, J.A. Diagnosis, treatment and management of Haemonchus contortus in small ruminants. Adv. Parasitol. 2016, 93, 181–238. [Google Scholar] [PubMed]
- García-Bustos, J.F.; Sleebs, B.E.; Gasser, R.B. An appraisal of natural products active against parasitic nematodes of animals. Parasit. Vectors 2019, 12, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The re-emergence of natural products for drug discovery in the genomics era. Nat. Rev. Drug. Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koehn, F.; Carter, G. The evolving role of natural products in drug discovery. Nat. Rev. Drug. Discov. 2005, 4, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Oprea, T.I. Property distribution of drug-related chemical databases. J. Comput. Aided Mol. Des. 2000, 14, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Quinn, R.J.; Carroll, A.R.; Pham, N.B.; Baron, P.; Palframan, M.E.; Suraweera, L.; Pierens, G.K.; Muresan, S. Developing a drug-like natural product library. J. Nat. Prod. 2008, 71, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Abdelaleem, E.R.; Samy, M.N.; Desoukey, S.Y.; Liu, M.; Quinn, R.J.; Abdelmohsen, U.R. Marine natural products from sponges (Porifera) of the order Dictyoceratida (2013 to 2019); a promising source for drug discovery. RSC Adv. 2020, 10, 34959–34976. [Google Scholar] [CrossRef]
- Mehbub, M.F.; Perkins, M.V.; Zhang, W.; Franco, C.M.M. New marine natural products from sponges (Porifera) of the order Dictyoceratida (2001 to 2012); a promising source for drug discovery, exploration and future prospects. Biotechnol. Adv. 2016, 34, 473–491. [Google Scholar] [CrossRef]
- Herath, H.M.P.D.; Taki, A.C.; Sleebs, B.E.; Hofmann, A.; Nguyen, N.; Preston, S.; Davis, R.A.; Jabbar, A.; Gasser, R.B. Advances in the discovery and development of anthelmintics by harnessing natural product scaffolds. Adv. Parasitol. 2021, 111, 203–251. [Google Scholar] [PubMed]
- Jiao, Y.; Preston, S.; Hofmann, A.; Taki, A.C.; Baell, J.B.; Chang, B.C.H.; Jabbar, A.; Gasser, R.B. A perspective on the discovery of selected compounds with anthelmintic activity against the barber’s pole worm–where to from here? Adv. Parasitol. 2020, 108, 1–45. [Google Scholar] [PubMed]
- Herath, H.; Preston, S.; Jabbar, A.; García-Bustos, J.; Taki, A.; Addison, R.; Hayes, S.; Beattie, K.; McGee, S.; Martin, S.; et al. Identification of fromiamycalin and halaminol A from Australian marine sponge extracts with anthelmintic activity against Haemonchus contortus. Mar. Drugs 2019, 17, 598. [Google Scholar] [CrossRef] [Green Version]
- Blunt, J.W.; Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2018, 35, 8–53. [Google Scholar] [CrossRef] [Green Version]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2019, 36, 122–173. [Google Scholar] [CrossRef] [Green Version]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2020, 37, 175–223. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2021, 38, 362–413. [Google Scholar] [CrossRef]
- Casertano, M.; Menna, M.; Imperatore, C. The Ascidian-derived metabolites with antimicrobial properties. Antibiotics 2020, 9, 510. [Google Scholar] [CrossRef] [PubMed]
- Taki, A.C.; Byrne, J.J.; Wang, T.; Sleebs, B.E.; Nguyen, N.; Hall, R.S.; Korhonen, P.K.; Chang, B.C.H.; Jackson, P.; Jabbar, A.; et al. High-throughput phenotypic assay to screen for anthelmintic activity on Haemonchus contortus. Pharmaceuticals 2021, 14, 616. [Google Scholar] [CrossRef]
- Camp, D.; Campitelli, M.; Carroll, A.R.; Davis, R.A.; Quinn, R.J. Front-loading natural-product-screening libraries for log P: Background, development, and implementation. Chem. Biodivers. 2013, 10, 524–537. [Google Scholar] [CrossRef] [Green Version]
- Camp, D.; Davis, R.A.; Campitelli, M.; Ebdon, J.; Quinn, R.J. Drug-like properties: Guiding principles for the design of natural product libraries. J. Nat. Prod. 2012, 75, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Le, T.G.; Kundu, A.; Ghoshal, A.; Nguyen, N.H.; Preston, S.; Jiao, Y.; Ruan, B.; Xue, L.; Huang, F.; Keiser, J.; et al. Optimization of novel 1-methyl-1 h -pyrazole-5-carboxamides leads to high potency larval development inhibitors of the barber’s pole worm. J. Med. Chem. 2018, 61, 10875–10894. [Google Scholar] [CrossRef]
- Simonetta, S.H.; Golombek, D.A. An automated tracking system for Caenorhabditis elegans locomotor behavior and circadian studies application. J. Neurosci. Methods 2007, 161, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.H.; Chung, T.D.; Oldenburg, K.R. A simple statistical parameter for use in evaluation and validation of high throughput screening assays. J. Biomol. Screen. 1999, 4, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Sui, Y.; Wu, Z. Alternative statistical parameter for high-throughput screening assay quality assessment. J. Biomol. Screen. 2007, 12, 229–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommerville, R.I. The development of Haemonchus contortus to the fourth stage in vitro. J. Parasitol. 1966, 52, 127–136. [Google Scholar] [CrossRef]
- Preston, S.; Jabbar, A.; Nowell, C.; Joachim, A.; Ruttkowski, B.; Baell, J.; Cardno, T.; Korhonen, P.K.; Piedrafita, D.; Ansell, B.R.; et al. Low cost whole-organism screening of compounds for anthelmintic activity. Int. J. Parasitol. 2015, 45, 333–343. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| NatureBank extract code | Species | Family (group*) | xL3 motility inhibition at 90 h | L4 development inhibition at 168 h | Abnormal phenotype detected (%) at 168 h |
|---|---|---|---|---|---|
| NB5866465 | Acanthophora muscoides | Rhodomelaceae (s) | 71 | No | nd |
| NB028803 | Agelas axifera | Agelasidae (s) | 96 | Yes | Cur (80) |
| NB029537 | Callyspongia (Callyspongia) sp. | Callyspongiidae (s) | 76 | No | nd |
| NB015513 | Callyspongia (Callyspongia) sp. | Callyspongiidae (s) | 75 | No | nd |
| NB007753 | Callyspongia (Euplacella) sp. | Callyspongiidae (s) | 79 | No | nd |
| NB6016992 | Callyspongia (Toxachalina) sp. | Callyspongiidae (s) | 82 | Yes | Cur (95) |
| NB6018007 | Ceratopsion clavatum | Raspailiidae (s) | 107 | No | nd |
| NB6004722 | Chalinula sp. | Chalinidae (s) | 75 | No | nd |
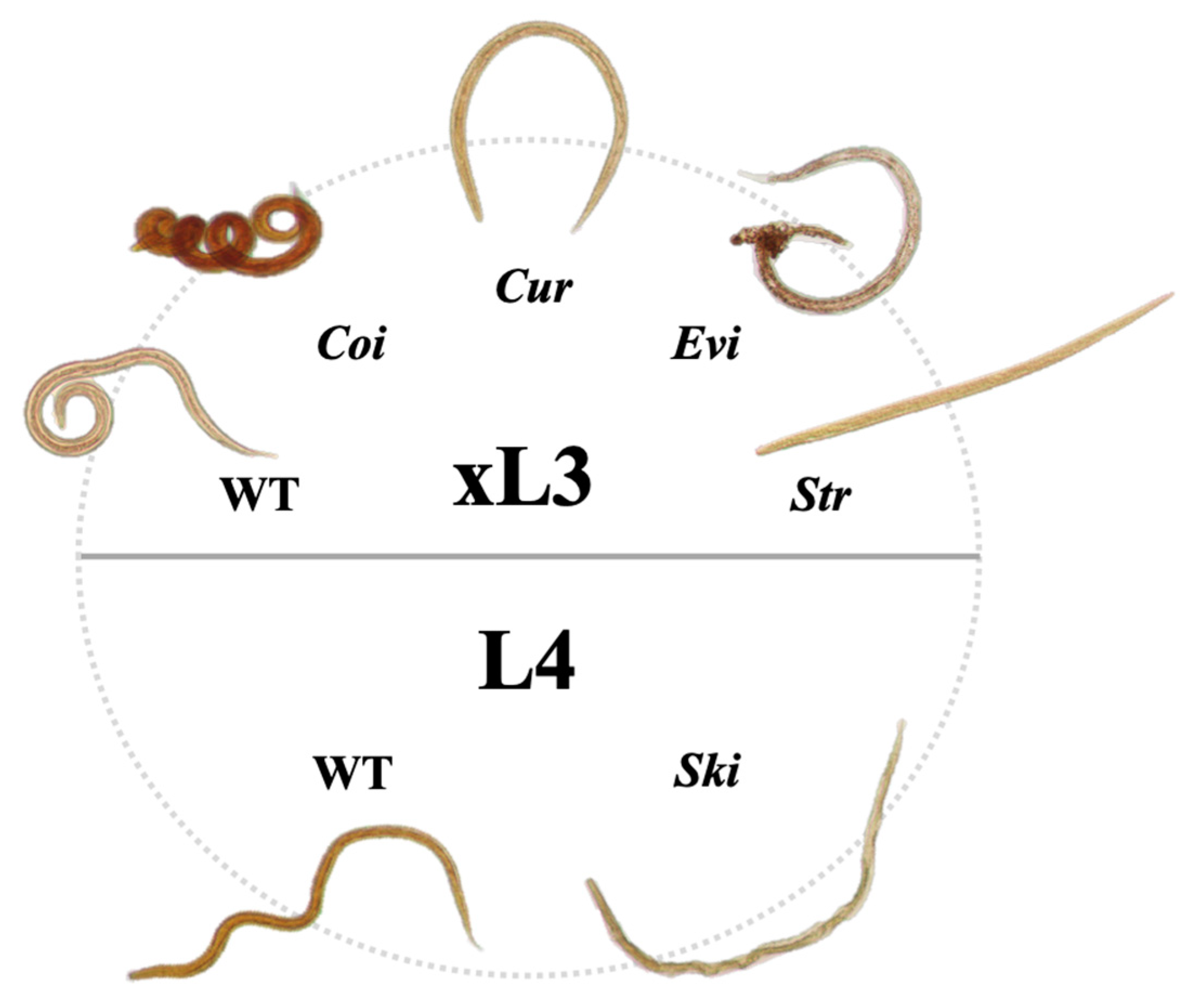
| NB031644 | Cinachyrella (Raphidotethya) enigmatica | Tetillidae (s) | 72 | Yes | Cur (40), Coi (20) |
| NB6018049 | Citronia sp. | Dysideidae (s) | 70 | No | Ski (60), Cur (40) |
| NB6014898 | Clathria (Thalysias) reinwardti | Microcionidae (s) | 72 | No | nd |
| NB6020295 | Clathria sp. | Microcionidae (s) | 73 | No | nd |
| NB6013853 | Coscinoderma mathewsi | Spongiidae (s) | 102 | No | Cur (30), Evi (25) |
| NB5866375 | Coscinoderma mathewsi | Spongiidae (s) | 97 | No | nd |
| NB6007999 | Coscinoderma mathewsi | Spongiidae (s) | 88 | Yes | nd |
| NB6013552 | Coscinoderma mathewsi | Spongiidae (s) | 87 | No | Cur (40), Evi (10) |
| NB6008047 | Coscinoderma mathewsi | Spongiidae (s) | 84 | No | nd |
| NB6009651 | Coscinoderma mathewsi | Spongiidae (s) | 79 | No | nd |
| NB5866277 | Coscinoderma mathewsi | Spongiidae (s) | 72 | No | nd |
| NB6008378 | Coscinoderma mathewsi | Spongiidae (s) | 71 | No | nd |
| NB6009654 | Coscinoderma mathewsi | Spongiidae (s) | 70 | Yes | nd |
| NB5379207 | Cymbastela coralliophila | Axinellidae (s) | 80 | No | Ski (90) |
| NB6009659 | Desmacella sp. | Desmacellidae (s) | 83 | No | Cur (70), Evi (10) |
| NB6005201 | Echinochalina (Protophlitaspongia) sp. | Microcionidae (s) | 100 | Yes | Cur (95) |
| NB6020433 | Endectyon sp. | Raspailiidae (s) | 75 | No | nd |
| NB6003967 | Erylus amissus | Geodiidae (s) | 85 | No | Cur (50) |
| NB5866232 | Fascaplysinopsis reticulata | Thorectidae (s) | 73 | No | Ski (95) |
| NB6018006 | Gelliodes sp. | Niphatidae (s) | 71 | No | nd |
| NB012605 | Haliclona (Haliclona) sp. | Chalinidae (s) | 85 | Yes | Str (90), Cur (10) |
| NB029001 | Haliclona (Reniera) sp. | Chalinidae (s) | 83 | Yes | Str (99) |
| NB6008001 | Haliclona (Reniera) sp. | Chalinidae (s) | 72 | No | nd |
| NB031634 | Ianthella flabelliformis | Ianthellidae (s) | 88 | No | nd |
| NB5820466 | Leucetta microraphis | Leucettidae (s) | 78 | Yes | Coi (80) |
| NB6009581 | Oceanapia sp. | Phloeodictyidae (s) | 80 | Yes | Cur (30), Coi (30) |
| NB6009479 | Oceanapia sp. | Phloeodictyidae (s) | 74 | No | Cur (90), Coi (20), Evi (10) |
| NB027467 | Petromica (Chaladesma) pacifica | Desmanthidae (s) | 77 | No | Ski (60), Cur (20) |
| NB6020712 | Phyllospongia foliascens | Thorectidae (s) | 81 | Yes | Cur (80) |
| NB6021239 | Phyllospongia foliascens | Thorectidae (s) | 72 | No | Cur (80) |
| NB5376298 | Phyllospongia bergquistae | Thorectidae (s) | 108 | No | Cur (30), Evi (20), Coi (20) |
| NB6005361 | Phyllospongia bergquistae | Thorectidae (s) | 107 | No | Cur (30), Evi (10) |
| NB5818101 | Phyllospongia bergquistae | Thorectidae (s) | 101 | Yes | nd |
| NB028821 | Phyllospongia bergquistae | Thorectidae (s) | 88 | Yes | Cur (100) |
| NB2434682 | Phyllospongia bergquistae | Thorectidae (s) | 79 | Yes | nd |
| NB6017543 | Phyllospongia bergquistae | Thorectidae (s) | 108 | No | Cur (30), Evi (10) |
| NB010981 | Phyllospongia papyracea | Thorectidae (s) | 93 | No | Cur (60) |
| NB5818080 | Phyllospongia papyracea | Thorectidae (s) | 88 | No | nd |
| NB6017542 | Phyllospongia papyracea | Thorectidae (s) | 85 | No | Cur (30), Coi (20) |
| NB6013898 | Polyfibrospongiaflabellifera | Thorectidae (s) | 99 | Yes | Cur (60) |
| NB5867103 | Psammocinia halmiformis | Irciniidae (s) | 75 | No | nd |
| NB6005306 | Psammocinia sp. | Irciniidae (s) | 84 | Yes | Cur (95) |
| NB008063 | Pseudoceratina sp. | Pseudoceratinidae (s) | 80 | No | nd |
| NB023362 | Rhabdastrella globostellata | Ancorinidae (s) | 95 | No | Ski (90) |
| NB6008040 | Rhabdastrella globostellata | Ancorinidae (s) | 81 | No | nd |
| NB5818959 | Rhabdastrella globostellata | Ancorinidae (s) | 78 | No | Ski (90) |
| NB6018071 | Didemnum molle | Didemnidae (t) | 71 | No | nd |
| NB5867348 | Didemnum perplexum | Didemnidae (t) | 73 | No | nd |
| NB6021174 | Polycarpa aurata | Styelidae (t) | 73 | No | nd |
| NB029740 | Sarcophyton cherbonnieri | Alcyoniidae (c) | 80 | No | nd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taki, A.C.; Byrne, J.J.; Jabbar, A.; Lum, K.Y.; Hayes, S.; Addison, R.S.; Ramage, K.S.; Hofmann, A.; Ekins, M.G.; Wang, T.; et al. High Throughput Screening of the NatureBank ‘Marine Collection’ in a Haemonchus Bioassay Identifies Anthelmintic Activity in Extracts from a Range of Sponges from Australian Waters. Molecules 2021, 26, 5846. https://doi.org/10.3390/molecules26195846
Taki AC, Byrne JJ, Jabbar A, Lum KY, Hayes S, Addison RS, Ramage KS, Hofmann A, Ekins MG, Wang T, et al. High Throughput Screening of the NatureBank ‘Marine Collection’ in a Haemonchus Bioassay Identifies Anthelmintic Activity in Extracts from a Range of Sponges from Australian Waters. Molecules. 2021; 26(19):5846. https://doi.org/10.3390/molecules26195846
Chicago/Turabian StyleTaki, Aya C., Joseph J. Byrne, Abdul Jabbar, Kah Yean Lum, Sasha Hayes, Russell S. Addison, Kelsey S. Ramage, Andreas Hofmann, Merrick G. Ekins, Tao Wang, and et al. 2021. "High Throughput Screening of the NatureBank ‘Marine Collection’ in a Haemonchus Bioassay Identifies Anthelmintic Activity in Extracts from a Range of Sponges from Australian Waters" Molecules 26, no. 19: 5846. https://doi.org/10.3390/molecules26195846