
Juglone Inactivates Pseudomonas aeruginosa through Cell Membrane Damage, Biofilm Blockage, and Inhibition of Gene Expression


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Culture
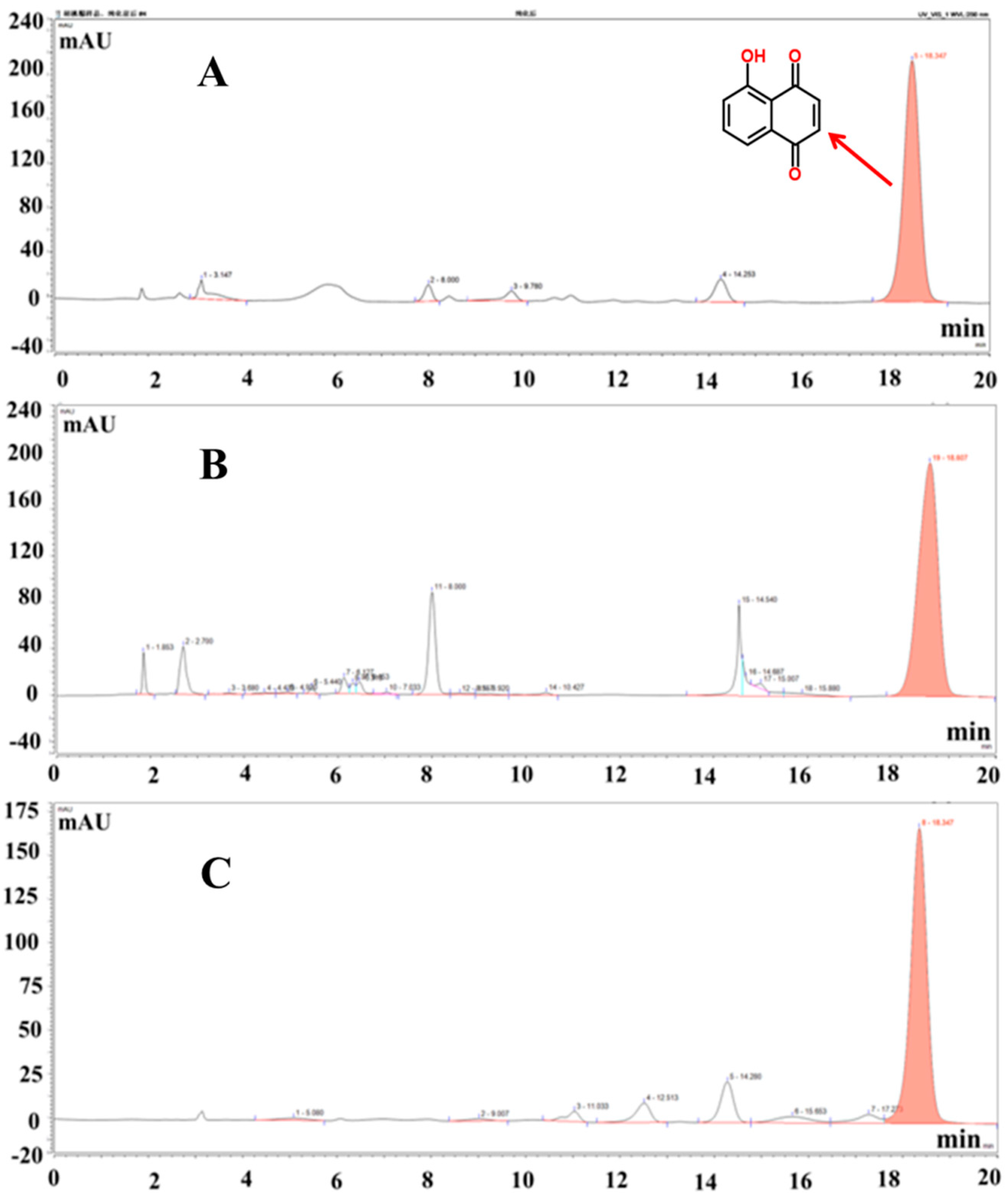
2.2. Extraction, Purification of Juglone, and Content Analysis
2.3. Antibacterial Activity against P. aeruginosa
2.3.1. Determination of Bacteriostatic Zone Diameter (DIZ)
2.3.2. Determination of Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC)
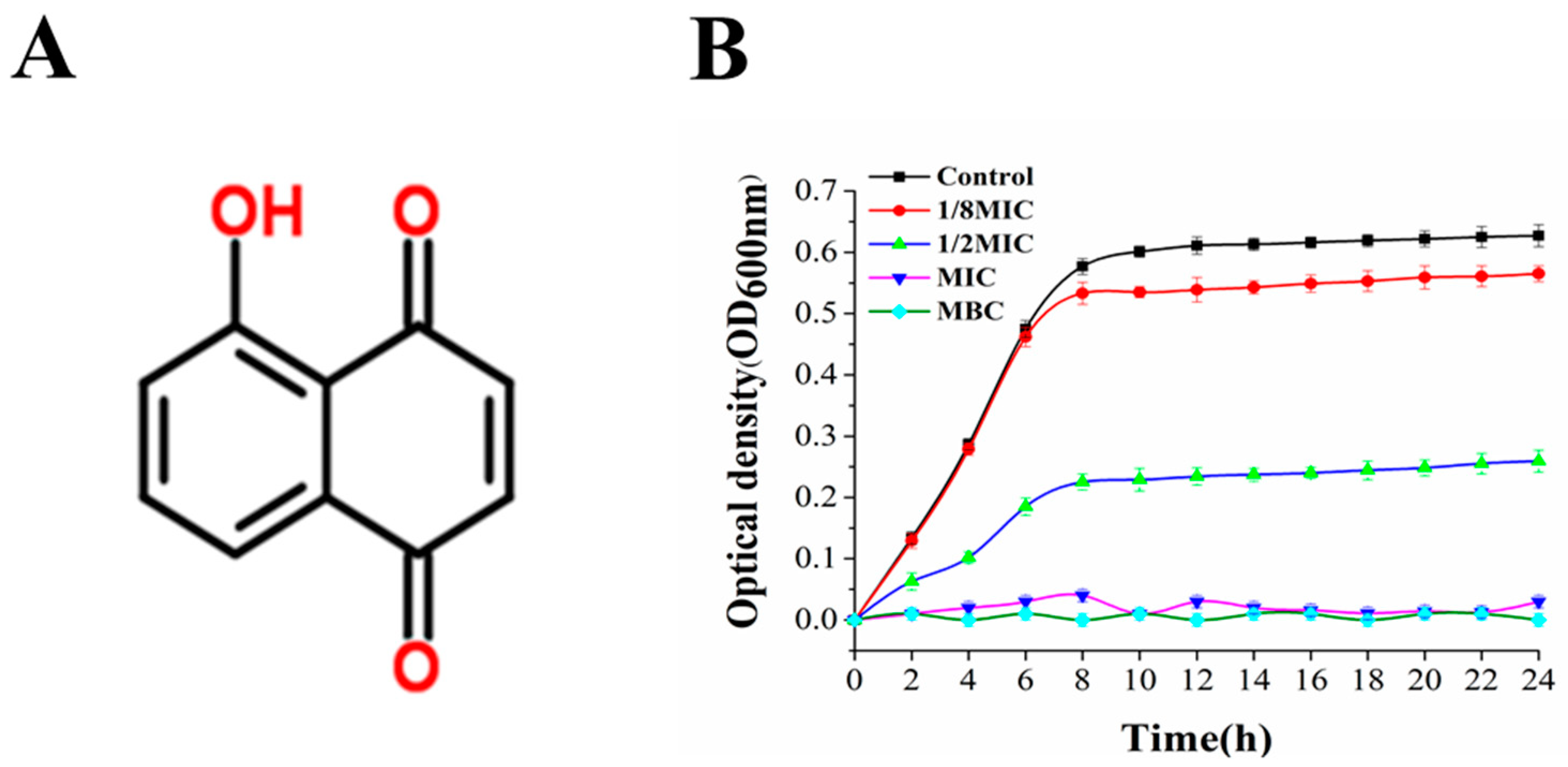
2.4. Measurement of Growth Curve
2.5. Antimicrobial Mechanism of Juglone against P. aeruginosa
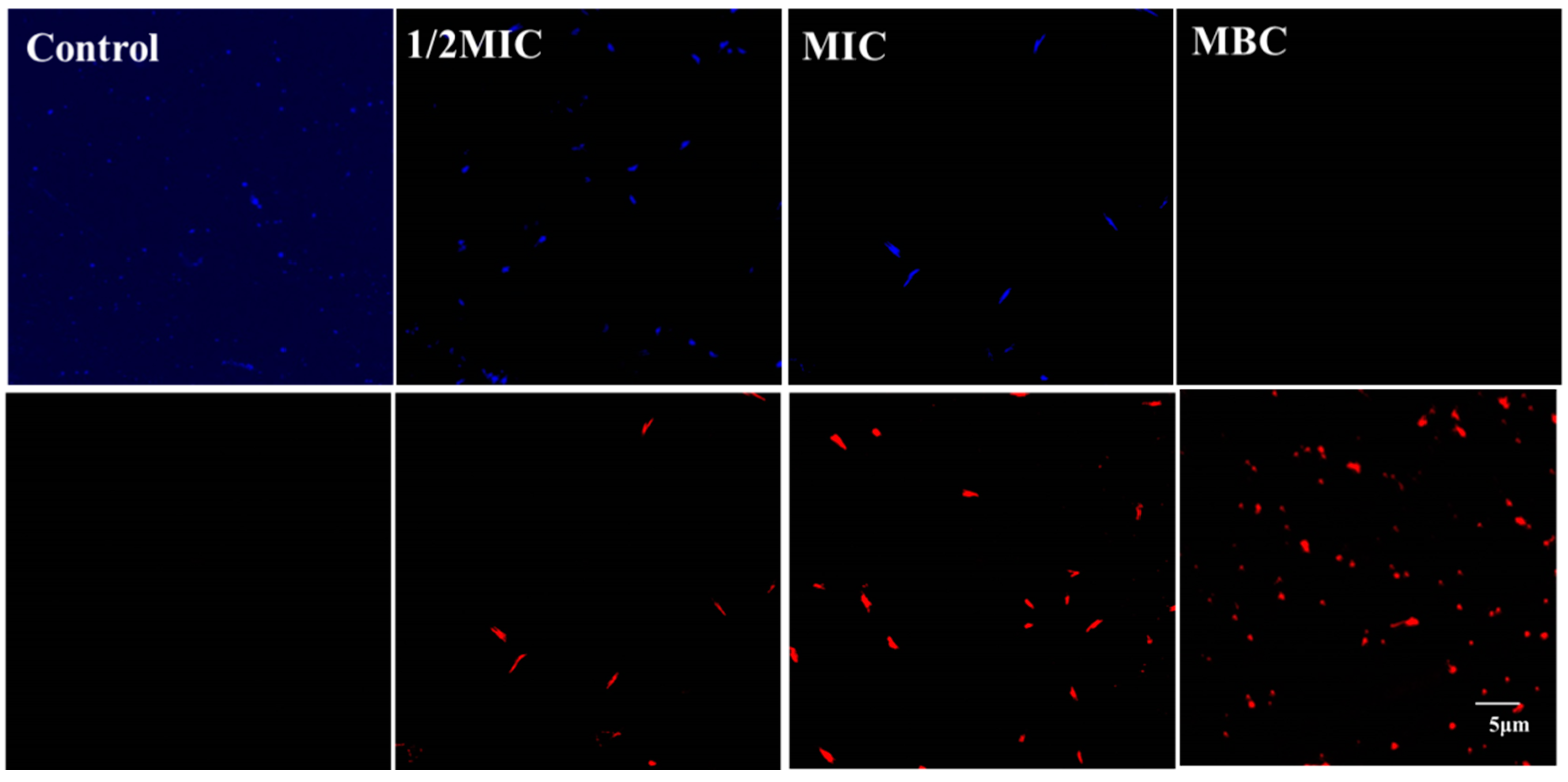
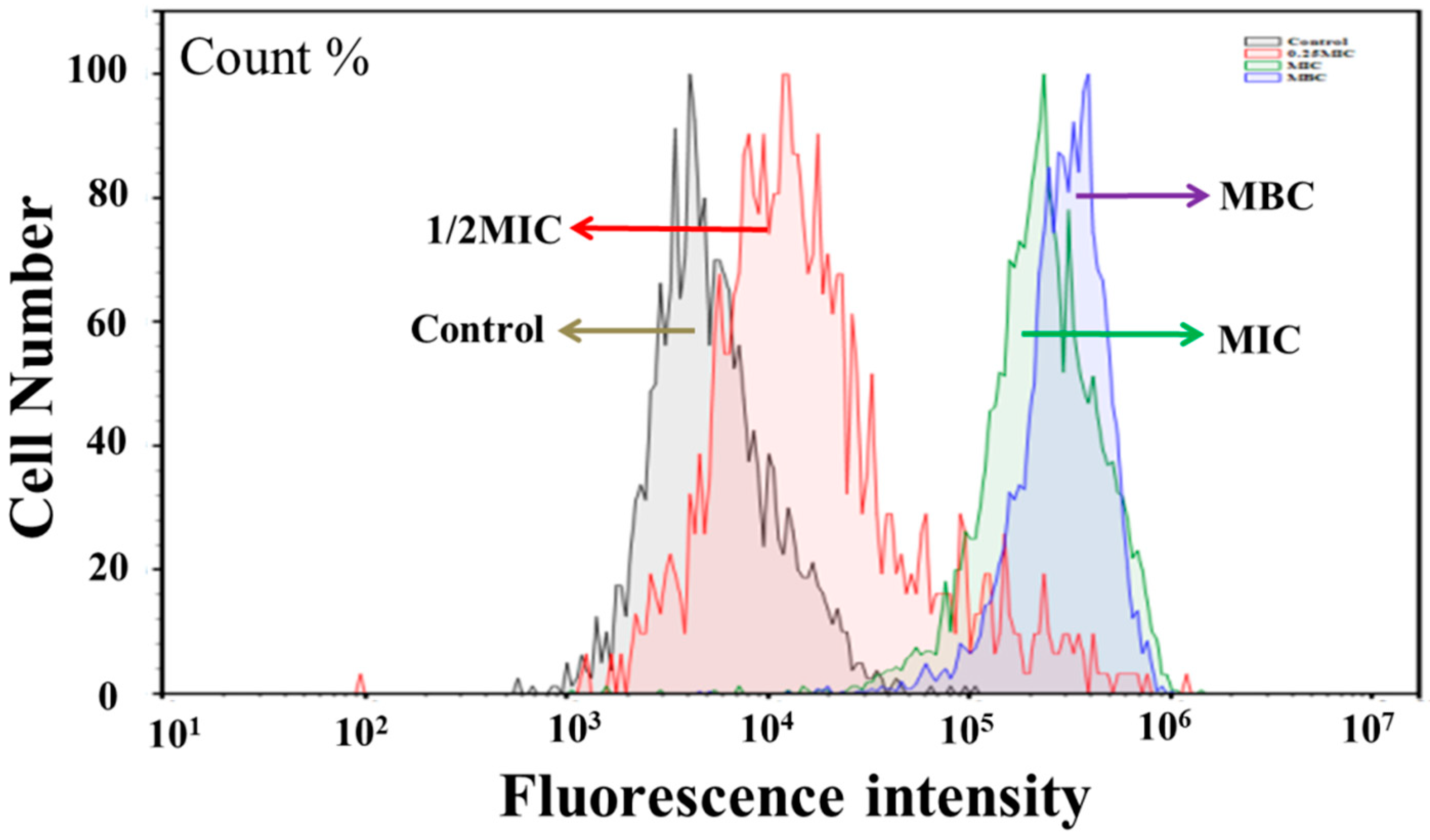
2.5.1. Confocal Laser Scanning Microscopy (CLSM)
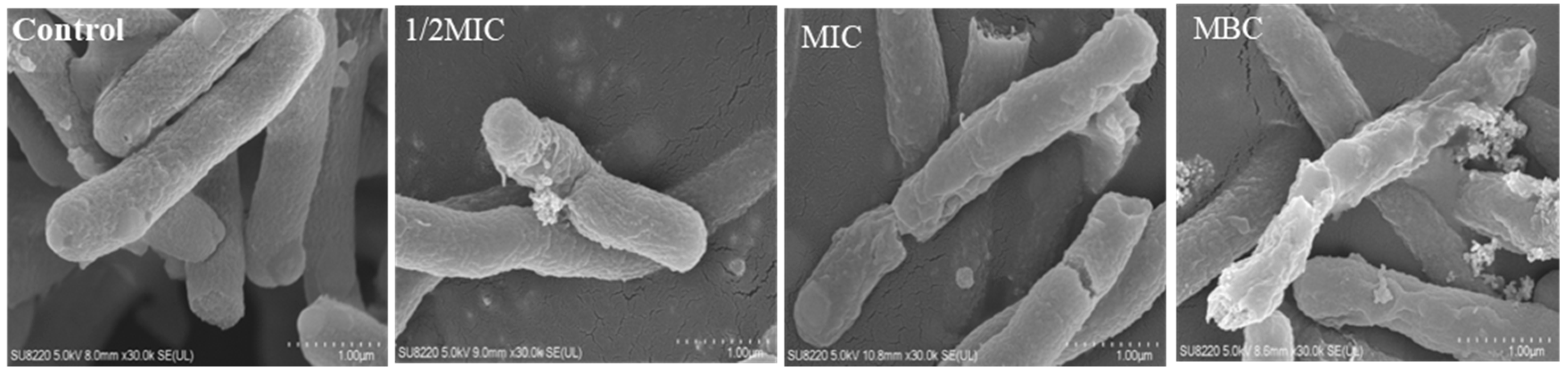
2.5.2. Field-Emission Scanning Electron Microscopy (FE-SEM)
2.5.3. Determination of Cell Membrane Fluidity
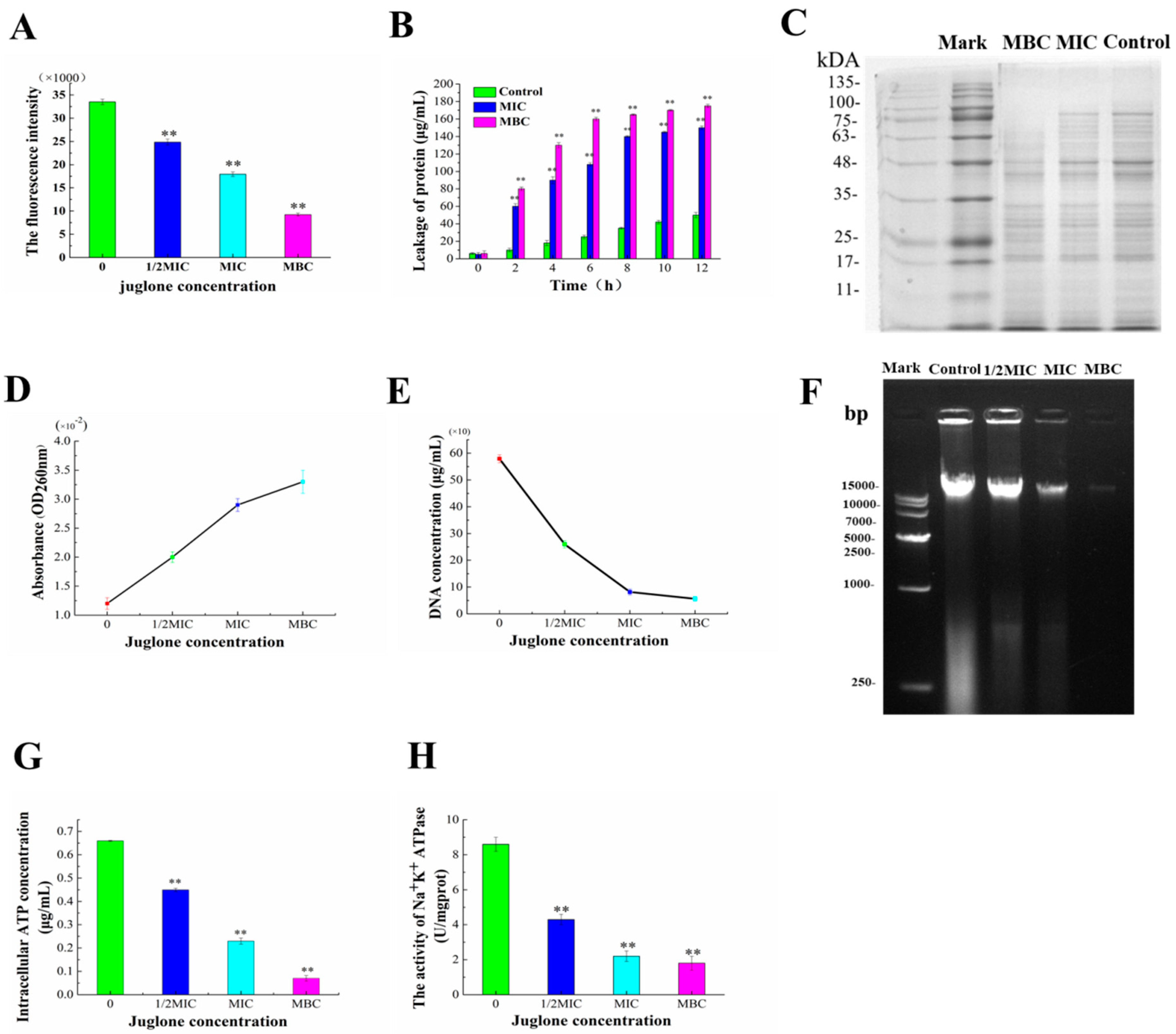
2.5.4. Whole-Cell Protein Analysis and Determination of Protein Leakage
2.5.5. Intracellular and Extracellular Nucleic Acid Leakage Analysis and DNA Gel Electrophoresis
2.5.6. Determination of Intracellular ATP Concentration and ATPase Activity
2.6. Reactive Oxygen Species (ROS) Analysis
2.7. Inhibition of Biofilm Formation by Juglone
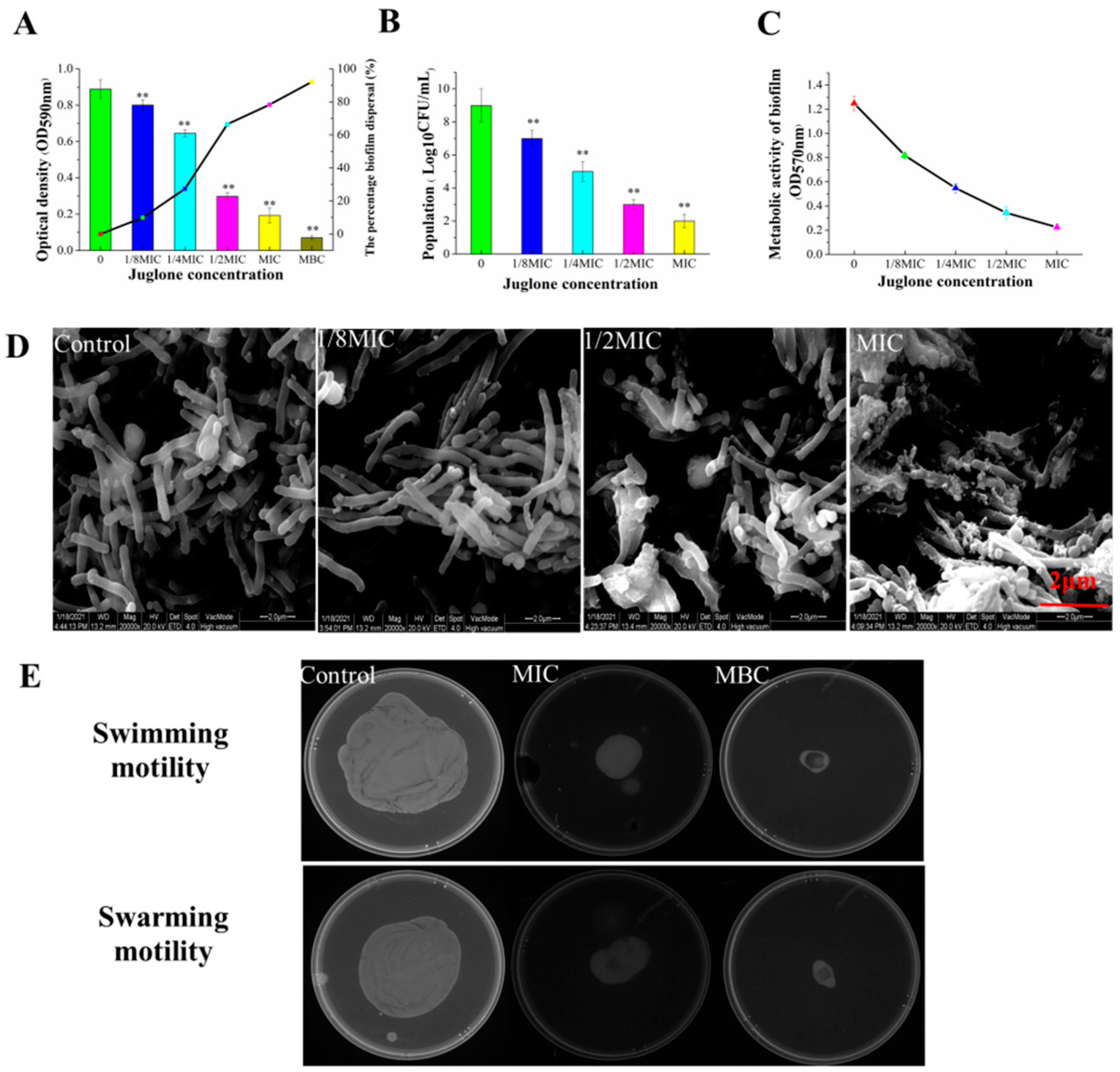
2.7.1. Biofilm Formation
2.7.2. Determination of Living Cells
2.7.3. ESEM Morphological Analysis of Biofilm
2.7.4. Determination of Bacterial Metabolic Activity in Biofilms
2.7.5. Swimming and Swarming Motilities Assays
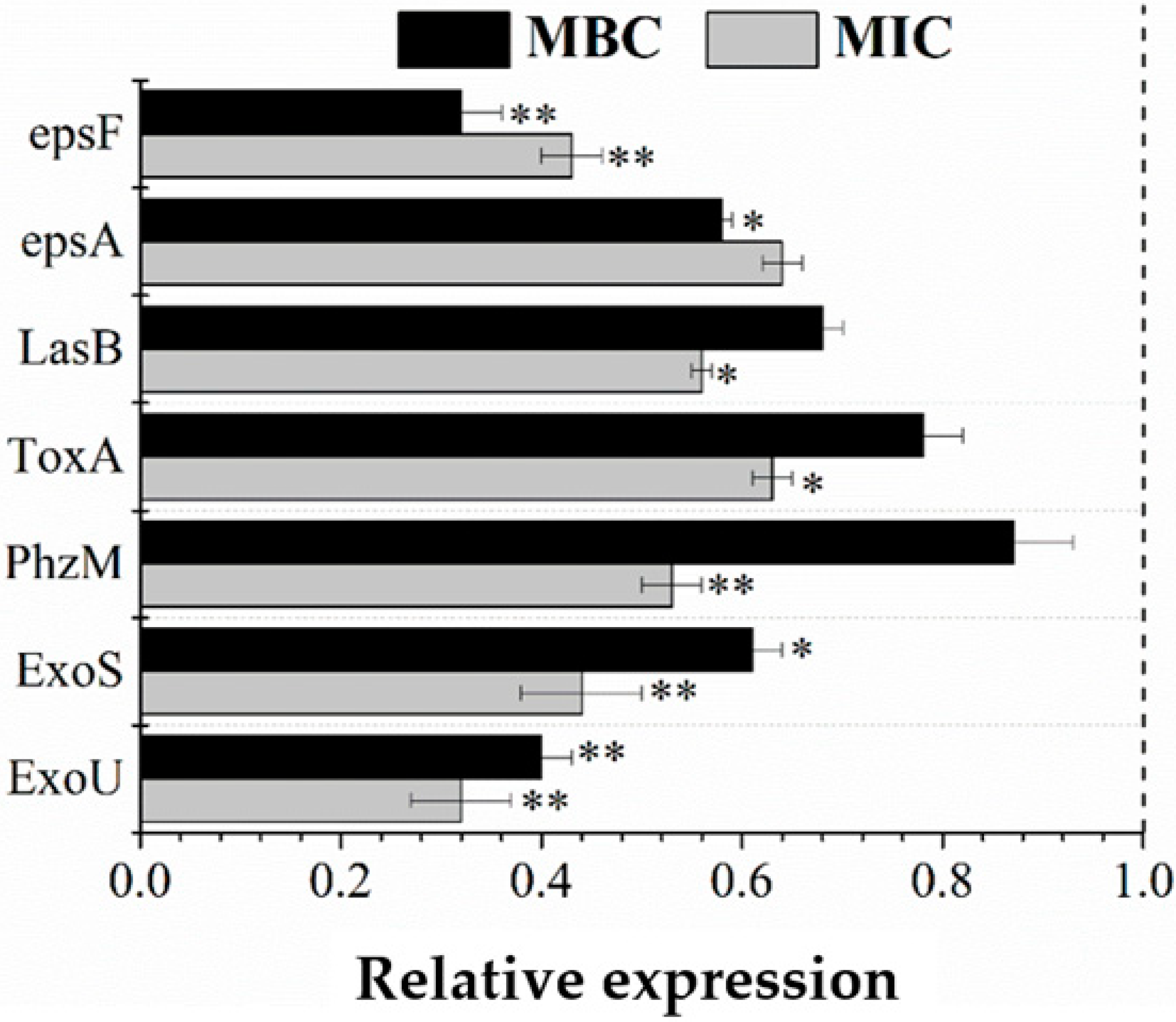
2.8. RNA Extraction and Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR) Validation
2.9. Statistical Analysis
3. Results
3.1. Juglone Extraction Rate, Purity, and HPLC Content Analysis
3.2. Antimicrobial Activity of Juglone and Its Effect on the Growth of P. aeruginosa
3.3. Effect of Juglone on Cell Viability
3.4. Effect of Juglone on the Morphology of P. aeruginosa
3.5. Destruction of Cell Membrane of P. aeruginosa by Juglone
3.6. Intracellular Oxidative Stress Induced by Juglone
3.7. Effect of Juglone on Biofilm Formation of P. aeruginosa
3.8. RT-qPCR Validation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Liu, H.; Pei, H.; Han, Z.; Feng, G.; Li, D. The antimicrobial effects and synergistic antibacterial mechanism of the combination of ε-Polylysine and nisin against Bacillus subtilis. Food Control 2015, 47, 444–450. [Google Scholar] [CrossRef]
- Liu, X.; Cai, J.; Chen, H.; Zhong, Q.; Chen, W. Antibacterial activity and mechanism of linalool against pseudomonas aeruginosa. Microb. Pathog. 2020, 141, 103980. [Google Scholar] [CrossRef]
- Ma, Q.; Davidson, P.M.; Zhong, Q. Antimicrobial properties of lauric arginate alone or in combination with essential oils in tryptic soy broth and 2% reduced fat milk. Int. J. Food Microbiol. 2013, 166, 77–84. [Google Scholar] [CrossRef]
- Kito, M.; Onji, Y.; Yoshida, T.; Nagasawa, T. Occurrence of ε-poly-l-lysine-degrading enzyme in?-poly-l-lysine-tolerant Sphingobacterium multivorum OJ10: Purification and characterization. FEMS Microbiol. Lett. 2002, 207, 147–151. [Google Scholar] [CrossRef]
- Liu, T.; Kang, J.; Liu, L. Thymol as a critical component of Thymus vulgaris L. essential oil combats Pseudomonas aeruginosa by intercalating DNA and inactivating biofilm. LWT-Food Sci. Technol. 2021, 136, 110354. [Google Scholar] [CrossRef]
- Lavenir, R.; Jocktane, D.; Laurent, F.; Nazaret, S.; Cournoyer, B. Improved reliability of Pseudomonas aeruginosa PCR detection by the use of the species-specific ecfX gene target. J. Microbiol. Methods 2007, 70, 20–29. [Google Scholar] [CrossRef]
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouységu, L. Plant Polyphenols: Chemical Properties, Biological Activities, and Synthesis. Angew. Chem. Int. Ed. 2011, 50, 586–621. [Google Scholar] [CrossRef]
- Stampar, F.; Solar, A.; Hudina, M.; Veberic, R.; Colaric, M. Traditional walnut liqueur–cocktail of phenolics. Food Chem. 2006, 95, 627–631. [Google Scholar] [CrossRef]
- Salejda, A.M.; Janiewicz, U.; Korzeniowska, M.; Kolniak-Ostek, J.; Krasnowska, G. Effect of walnut green husk addition on some quality properties of cooked sausages. LWT 2016, 65, 751–757. [Google Scholar] [CrossRef]
- Boussetta, T.; Gougerot-Pocidalo, M.A.; Hayem, G.; Ciappelloni, S.; El-Benna, J. The prolyl isomerase Pin1 acts as a novel molecular switch for TNF-α–induced priming of the NADPH oxidase in human neutrophils. Blood 2010, 116, 5795. [Google Scholar] [CrossRef] [Green Version]
- Catanzaro, E.; Greco, G.; Potenza, L.; Calcabrini, C. Natural Products to Fight Cancer: A Focus on Juglans regia. Toxins 2018, 10, 469. [Google Scholar] [CrossRef] [Green Version]
- Sousa, A. Walnut (Juglans regia L.) leaves: Phenolic compounds, antibacterial activity and antioxidant potential of different cultivars. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2007, 45, 2287–2295. [Google Scholar]
- Wang, J.; Liu, D.; Sun, X.; Bai, B.; Jiang, D.; Wu, Z. Label-free quantitative proteomic analysis of the inhibitory activities of juglone against translation and energy metabolism in Escherichia coli. Phytochem. Lett. 2016, 18, 55–58. [Google Scholar] [CrossRef]
- Caballero, E.; Soto, C.; Jara, J. Thermal stability data of juglone from extracts of walnut (Juglans regia) green husk, and technologies used to concentrate juglone. Data Brief 2019, 25, 104081. [Google Scholar] [CrossRef]
- Kang, J.; Liu, L.; Wu, X.; Sun, Y.; Liu, Z. Effect of thyme essential oil against Bacillus cereus planktonic growth and biofilm formation. Appl. Microbiol. Biotechnol. 2018, 102, 10209–10218. [Google Scholar] [CrossRef]
- Kang, J.; Liu, L.; Liu, Y.; Wang, X. Ferulic Acid Inactivates Shigella flexneri through Cell Membrane Destruction, Biofilm Retardation, and Altered Gene Expression. J. Agric. Food Chem. 2020, 68, 7121–7131. [Google Scholar] [CrossRef]
- Ning, H.Q.; Li, Y.Q.; Tian, Q.W.; Wang, Z.S.; Mo, H.Z. The apoptosis of Staphylococcus aureus induced by glycinin basic peptide through ROS oxidative stress response. LWT-Food Sci. Technol. 2018, 99, 62–68. [Google Scholar] [CrossRef]
- Trevors, J.T. Fluorescent probes for bacterial cytoplasmic membrane research. J. Biochem. Biophys. Methods 2003, 57, 87–103. [Google Scholar] [CrossRef]
- Abozahra, R.; El-Kholy, M.A.; Baraka, K. Virulence genotyping of drug resistant Pseudomonas aeruginosa clinical isolates in Egypt using multiplex PCR. Gene Rep. 2020, 22, 101000. [Google Scholar]
- Chen, Z.; Liu, P.; Li, Z.; Yu, W.; Wang, Z.; Yao, H.; Wang, Y.; Li, Q.; Deng, X.; He, N. Identification of Key Genes Involved in Polysaccharide Bioflocculant Synthesis in Bacillus Licheniformis. Biotechnol. Bioeng. 2017, 114, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Zmantar, T.; Miladi, H.; Kouidhi, B.; Chaabouni, Y.; Slama, R.B.; Bakhrouf, A.; Mahdouani, K.; Chaieb, K. Use of juglone as antibacterial and potential efflux pump inhibitors in staphylococcus aureus isolated from the oral cavity. Microb. Pathog. 2016, 101, 44–49. [Google Scholar] [CrossRef]
- Dong, D.; Zou, D.; Hui, L.; Yang, Z.; Huang, S.; Liu, N.; Huang, L. Rapid detection of Pseudomonas aeruginosa targeting the toxA gene in intensive care unit patients from Beijing, China. Front. Microbiol. 2015, 6, 1100. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Du, Y.; Wang, X.; Sun, L. Chitosan kills bacteria through cell membrane damage. Int. J. Food Microbiol. 2004, 95, 147–155. [Google Scholar] [CrossRef]
- Rurián-Henares, J.; Morales, F.J. Antimicrobial activity of melanoidins against Escherichia coli is mediated by a membrane-damage mechanism. J. Agric. Food Chem. 2008, 56, 2357–2362. [Google Scholar] [CrossRef] [Green Version]
- Stiefel, P.; Schmidt-Emrich, S.; Maniura-Weber, K.; Ren, Q. Critical aspects of using bacterial cell viability assays with the fluorophores SYTO9 and propidium iodide. BMC Microbiol. 2015, 15, 36. [Google Scholar] [CrossRef] [Green Version]
- Dong, G.L.; Kim, P.I.; Park, Y.; Woo, E.R.; Ji, S.C.; Choi, C.H.; Hahm, K.S. Design of novel peptide analogs with potent fungicidal activity, based on PMAP-23 antimicrobial peptide isolated from porcine myeloid. Biochem. Biophys. Res. Commun. 2002, 293, 231–238. [Google Scholar]
- Silva, N.; Alves, S.; Goncalves, A.; Amaral, J.S.; Poeta, P. Antimicrobial activity of essential oils from mediterranean aromatic plants against several foodborne and spoilage bacteria. Food Sci. Technol. Int. 2013, 19, 503–510. [Google Scholar] [CrossRef]
- Portet, T.; Franc, C.; Escoffre, J.M.; Favard, C.; Rols, M.P.; Dean, D.S. Visualization of Membrane Loss during the Shrinkage of Giant Vesicles under Electropulsation. Biophys. J. 2009, 96, 4109–4121. [Google Scholar] [CrossRef] [Green Version]
- Mytle, N.; Anderson, G.L.; Doyle, M.P.; Smith, M.A. Antimicrobial activity of clove (Syzgium aromaticum) oil in inhibiting Listeria monocytogenes on chicken frankfurters. Food Control 2006, 17, 102–107. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, X.; Wang, Y.; Jiang, P.; Quek, S. Antibacterial activity and mechanism of cinnamon essential oil against Escherichia coli and Staphylococcus aureus. Food Control 2016, 59, 282–289. [Google Scholar] [CrossRef]
- Gonzalez, B.; Glaasker, E.; Kunji, E.; Driessen, A.; Suarez, J.E.; Konings, W.N. Bactericidal mode of action of plantaricin C. Appl. Environ. Microbiol. 1996, 62, 2701–2709. [Google Scholar] [CrossRef] [Green Version]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Paredes, J.; Alonso-Arce, M.; Schmidt, C.; Valderas, D.; Sedano, B.; Legarda, J.; Pozo, J. Smart central venous port for early detection of bacterial biofilm related infections. Biomed. Microdevices 2014, 16, 365–374. [Google Scholar] [CrossRef]
- Dakheel, M.M.; Alkandari, F.; Mueller-Harvey, I.; Woodward, M.J.; Rymer, C. Antimicrobial in vitro activities of condensed tannin extracts on avian pathogenic Escherichia coli. Lett. Appl. Microbiol. 2020, 70, 165–172. [Google Scholar] [CrossRef]
- Shaver, C.M.; Hauser, A.R. Relative Contributions of Pseudomonas aeruginosa ExoU, ExoS, and ExoT to Virulence in the Lung. Infect. Immun. 2004, 72, 6969–6977. [Google Scholar] [CrossRef] [Green Version]
- Lau, G.W.; Ran, H.; Kong, F.; Hassett, D.J.; Mavrodi, D. Pseudomonas aeruginosa Pyocyanin Is Critical for Lung Infection in Mice. Infect. Immun. 2004, 72, 4275–4278. [Google Scholar] [CrossRef] [Green Version]
- Abdelali, D.; Sandy, F.; Matilde, F.; Zulema, U.; Tino, K.; Ramos, J.L.; Valerie, C.L. Genes for Carbon Metabolism and the ToxA Virulence Factor in Pseudomonas aeruginosa Are Regulated through Molecular Interactions of PtxR and PtxS. PLoS ONE 2012, 7, e39390. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer Sequence | Reverse Primer Sequence |
|---|---|---|
| Exou | CCAACACATTAGCAGCGAGA | TGGGAGTACATTGAGCAGCA |
| Exos | CATCCTCAGGCGTACATCCT | ATCGATGTCAGCGGGATATC |
| Phzm | CGGCGAAGACTTCTACAGCT | AGGTAGATATCGCCGTTGGA |
| Toxa | ATGGTGTAGATCGGCGACAT | AAGCCTTCGACCTCTGGAAC |
| Lasb | ACATCGCCCAACTGGTCTAC | ACCAGCGGATAGAACATGGT |
| Ecfx | ATGCCTATCAGGCGTTCCAT | GGCGATCTGGAAAAGAAATG |
| epsA | TTTATCGATGATACGGTTGCAAG | CTAATAGCCAAGCGGCTCACTC |
| epsF | CGGCTTTGAACGGTGGG | TCACTGTCCTTCTGCCGCG |
| DIZ (mm) | MIC (μg/mL) | MBC (μg/mL) | ||
|---|---|---|---|---|
| juglone | juglone | Gallic acid | juglone | Gallic acid |
| 22.89 ± 1.00 | 35 | 600 | 45 | 5000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Q.; Yan, X.; Zhang, R.; Wang, G.; Zhang, Y. Juglone Inactivates Pseudomonas aeruginosa through Cell Membrane Damage, Biofilm Blockage, and Inhibition of Gene Expression. Molecules 2021, 26, 5854. https://doi.org/10.3390/molecules26195854
Han Q, Yan X, Zhang R, Wang G, Zhang Y. Juglone Inactivates Pseudomonas aeruginosa through Cell Membrane Damage, Biofilm Blockage, and Inhibition of Gene Expression. Molecules. 2021; 26(19):5854. https://doi.org/10.3390/molecules26195854
Chicago/Turabian StyleHan, Qiqi, Xinpeng Yan, Runguang Zhang, Guoliang Wang, and Youlin Zhang. 2021. "Juglone Inactivates Pseudomonas aeruginosa through Cell Membrane Damage, Biofilm Blockage, and Inhibition of Gene Expression" Molecules 26, no. 19: 5854. https://doi.org/10.3390/molecules26195854