
Lycoperoside H, a Tomato Seed Saponin, Improves Epidermal Dehydration by Increasing Ceramide in the Stratum Corneum and Steroidal Anti-Inflammatory Effect

 and
and
Abstract
:1. Introduction
2. Results
2.1. Effect of TSE and Isolated Compounds (1–11) on the mRNA Expression of Proteins Related to Epidermal Hydration in HaCaT Cells
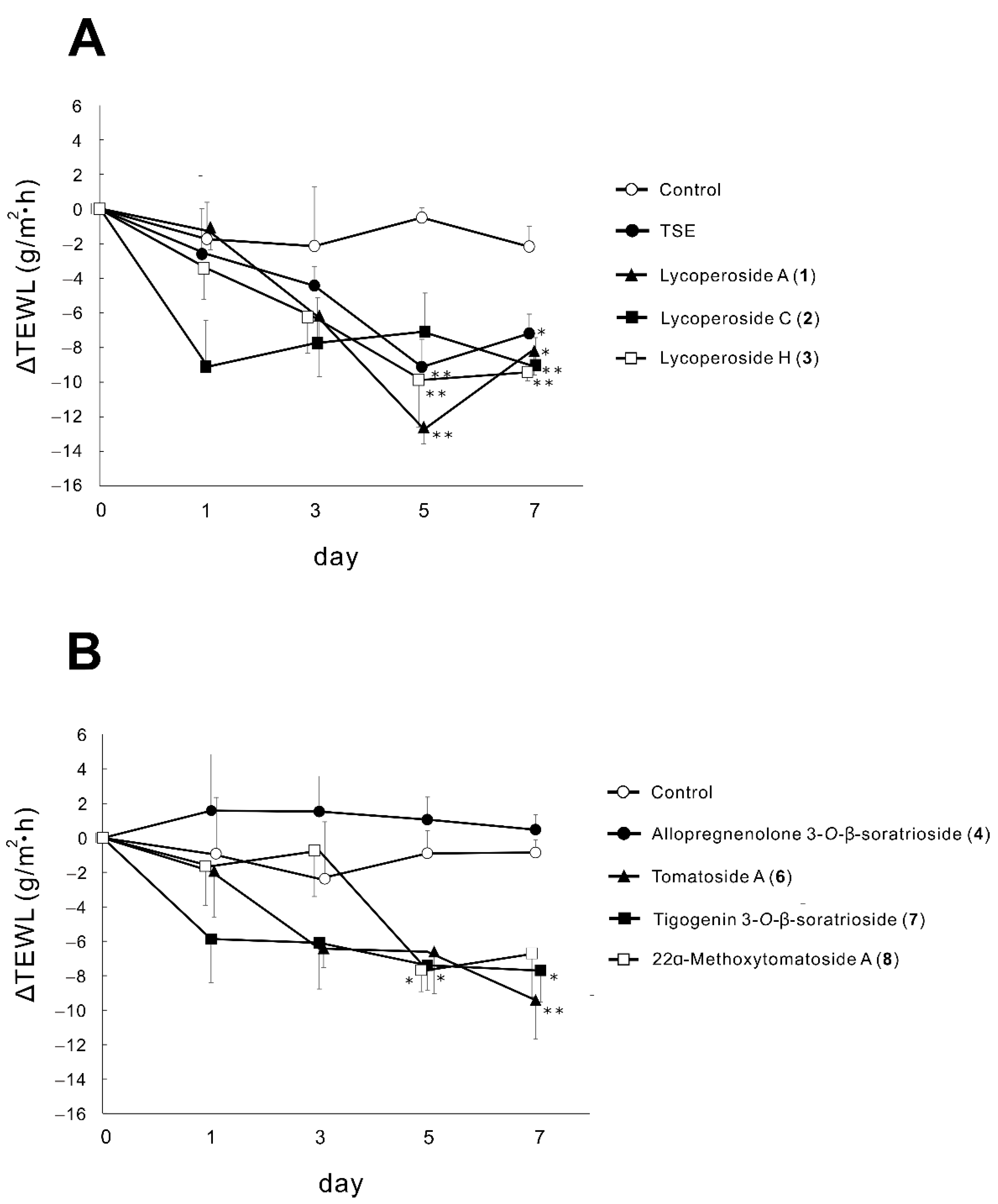
2.2. Effects of TSE and Saponins (1–4,6–8) on TEWL in RHEK Models
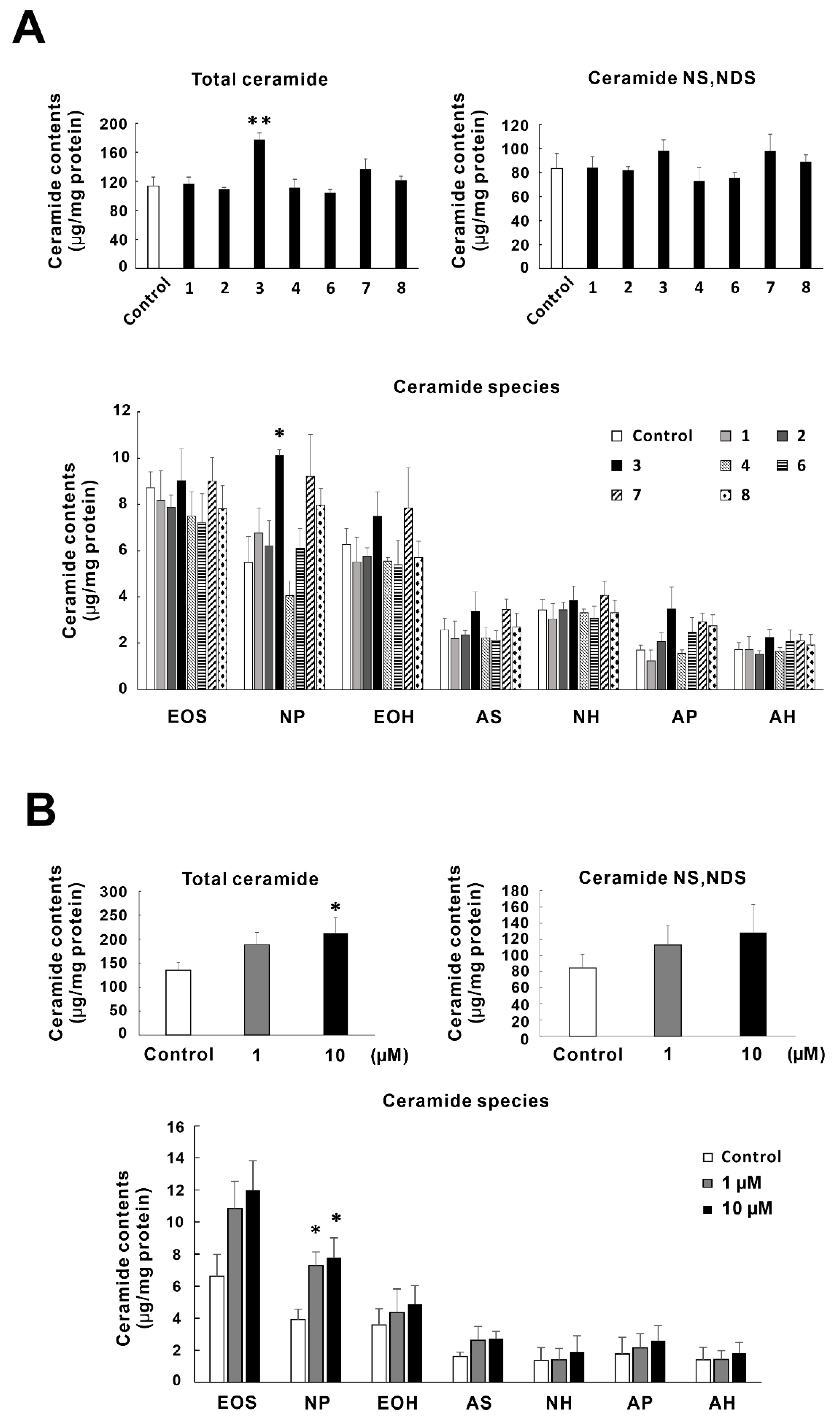
2.3. Effects of Tomato Seed Saponins (1–4,6–8) on the Ceramide Contents in SC of RHEK Models
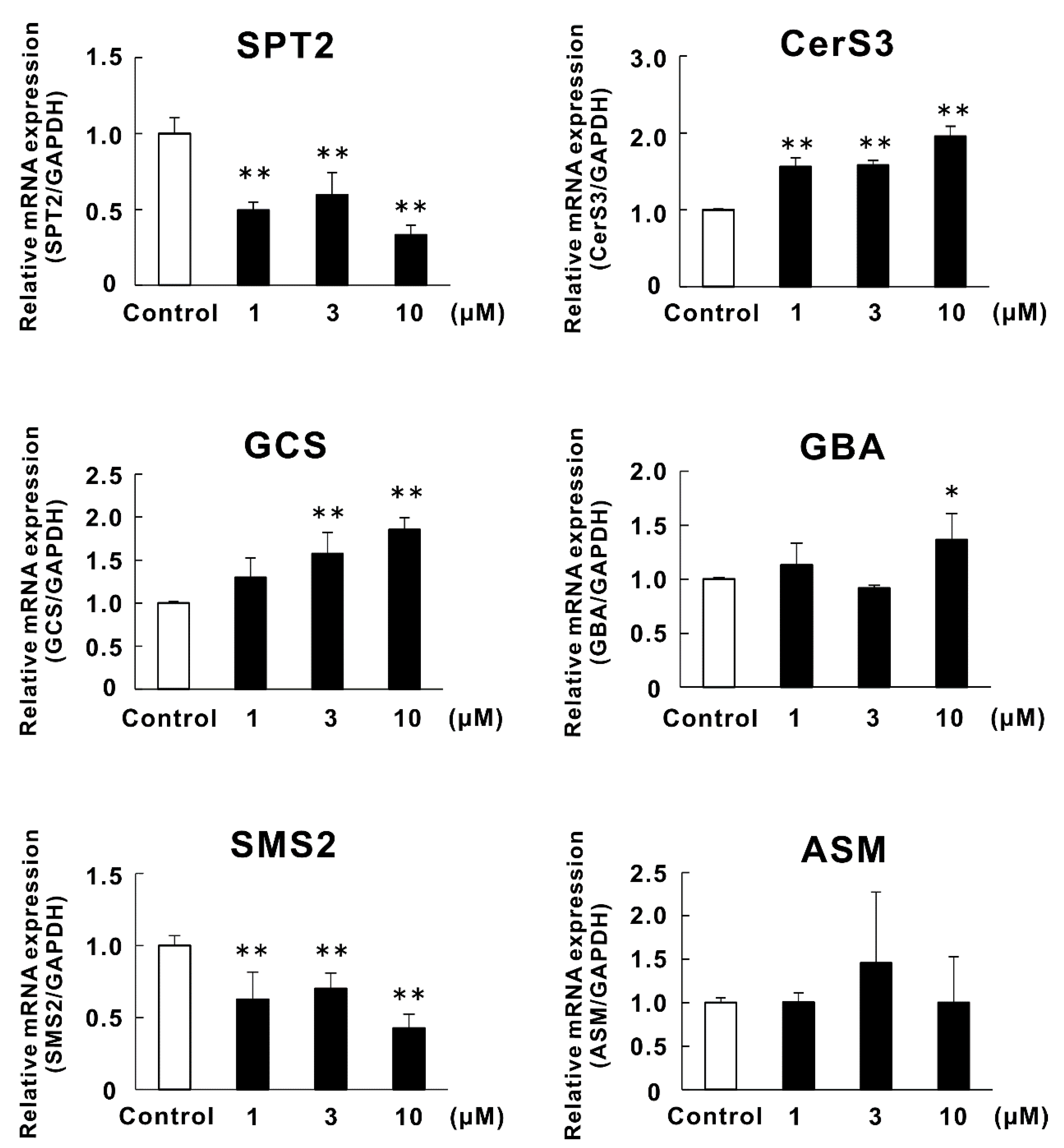
2.4. Effects of Lycoperoside H (3) on the mRNA Expression of Enzymes Related to Ceramide Synthesis in RHEK Models
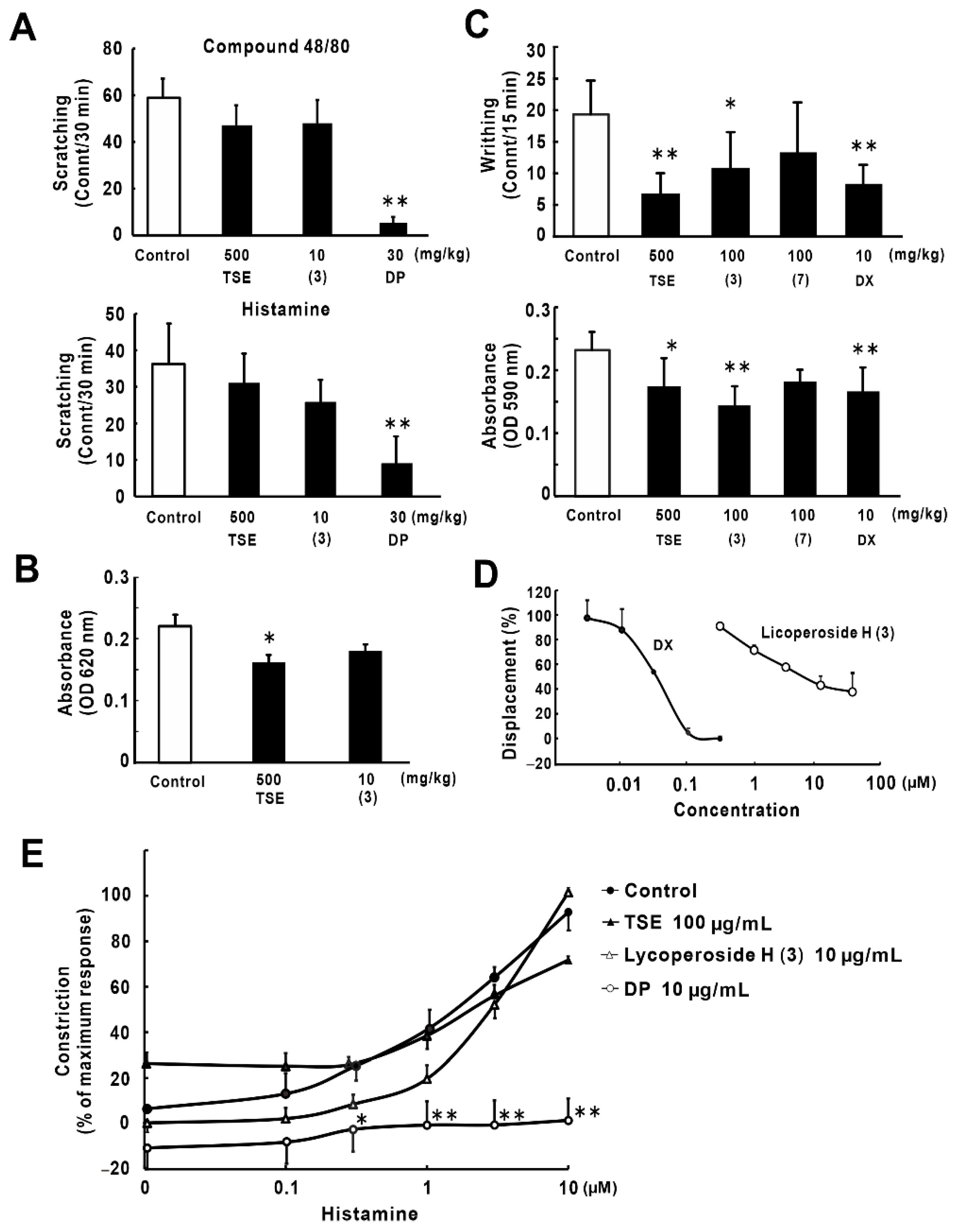
2.5. Anti-Inflammatory and Anti-Allergic Effects of Lycoperoside H (3)
3. Discussion
4. Materials and Methods
4.1. Preparation of Tomato Seed Extract and Its Compounds
4.2. Reagents
4.3. Cells and Animals
4.4. Culture of HaCaT Cells for the Screening of Candidate Compounds with Hydration Activity
4.5. Culture of the RHEK Models for the Evaluation of Hydration Factors
4.6. Measurement of TEWL in RHEK Models
4.7. Real-Time RT-PCR
4.8. Lipid Extraction and Ceramide Determination
4.9. Compound 48/80- and Histamine-Induced Mice Pruritus (In Vivo Anti-Inflammation Models)
4.10. PCA Reaction in Mice (In Vivo Anti-Allergy Model)
4.11. Acetic Acid-Induced Writhing and Vascular Permeability in Mice (In Vivo Anti-Inflammation Model)
4.12. Glucocorticoid Receptor Competitive Assay (In Vitro Assay for Glucocorticoid Receptor Ligand)
4.13. Histamine-Induced Guinea Pig Tracheal Muscle Contraction (In Vitro Anti-Histaminic Model)
4.14. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
| TSE | tomato seed extract |
| TEWL | transepidermal water loss |
| AD | atopic dermatitis |
| DP | diphenhydramine hydrochloride |
| C | Centella |
| DMEM | Dulbecco’s modified Eagle medium |
| PBS | phosphate-buffered saline |
| DX | dexamethasone |
| HPTLC | high-performance thin-layer chromatography |
| DNP-BSA | 2,4-dinitrophenylated bovine serum albumin |
| FBS | fetal bovine serum |
| RHEK | reconstructed human epidermal keratinization |
| SC | stratum corneum |
| DMSO | dimethyl sulfoxide |
| SDS | sodium lauryl sulfate |
| BCA | bicinchoninic acid |
| PCA | passive cutaneous anaphylaxis |
| SPT | serine palmitoyltransferase |
| CerS | ceramide synthase |
| GCS | glucosylceramide synthase |
| GBA | β-glucocerebrosidase |
| SMS | sphingomyelin synthase |
| ASM | acid sphingomyelinase |
References
- Chaudhary, P.; Sharma, A.; Singh, B.; Nagpal, A.K. Bioactivities of phytochemicals present in tomato. J. Food Sci. Technol. 2018, 55, 2833–2849. [Google Scholar] [CrossRef]
- Yahara, S.; Uda, N.; Nohara, T. Lycoperosides A–C, three stereoisomeric 23-acetoxyspirosolan-3β-ol β-lycotetraosides from Lycopersicon esculentum. Phytochemistry 1996, 42, 169–172. [Google Scholar] [CrossRef]
- Yoshizaki, M.; Matsushita, S.; Fujiwara, Y.; Ikeda, T.; Ono, M.; Nohara, T. Tomato new sapogenols, isoesculeogenin A and esculeogenin B. Chem. Pharm. Bull. 2005, 53, 839–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujiwara, Y.; Takaki, A.; Uehara, Y.; Ikeda, T.; Okawa, M.; Yamauchi, K.; Ono, M.; Yoshimitsu, H.; Nohara, T. Tomato steroidal alkaloid glycosides, esculeosides A and B, from ripe fruits. Tetrahedron 2004, 60, 4915–4920. [Google Scholar] [CrossRef]
- Fujiwara, Y.; Yahara, S.; Ikeda, T.; Ono, M.; Nohara, T. Cytotoxic major saponin from tomato fruits. Chem. Pharm. Bull. 2003, 51, 234–235. [Google Scholar] [CrossRef] [Green Version]
- Nohara, T.; Fujiwara, Y.; Zhou, J.R.; Urata, J.; Ikeda, T.; Murakami, K.; El-Aasr, M.; Ono, M. Saponins, esculeosides B-1 and B-2, in tomato juice and sapogenol, esculeogenin B1. Chem. Pharm. Bull. 2015, 63, 848–850. [Google Scholar] [CrossRef] [Green Version]
- Ono, M.; Takara, Y.; Egami, M.; Uranaka, K.; Yoshimitsu, H.; Matsushita, S.; Fujiwara, Y.; Ikeda, T.; Nohara, T. Steroidal alkaloid glycosides, esculeosides C and D, from the ripe fruit of cherry tomato. Chem. Pharm. Bull. 2006, 54, 237–239. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, Y.; Kiyota, N.; Hori, M.; Matsushita, S.; Iijima, Y.; Aoki, K.; Shibata, D.; Takeya, M.; Ikeda, T.; Nohara, T.; et al. Esculeogenin A, a new tomato sapogenol, ameliorates hyperlipidemia and atherosclerosis in ApoE-deficient mice by inhibiting ACAT. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2400–2406. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.R.; Urata, J.; Shiraishi, T.; Tanaka, C.; Nohara, T.; Yokomizo, K. Tomato juice saponin, esculeoside B ameliorates mice experimental dermatitis. FFHD 2018, 8, 228–241. [Google Scholar] [CrossRef]
- Takeda, S.; Miyasaka, K.; Shimoda, H. Lycoperoside H, a steroidal alkaloid saponin in tomato seeds, ameliorates atopic dermatitis-like symptoms in IL-33 transgenic mice. J. Food Biochem. 2021, 45, e13877. [Google Scholar] [CrossRef]
- Im, D.S. Pro-resolving effect of ginsenosides as an anti-inflammatory mechanism of Panax ginseng. Biomolecules 2020, 10, 444. [Google Scholar] [CrossRef] [Green Version]
- Agra, L.C.; Ferro, J.N.; Barbosa, F.T.; Barreto, E. Triterpenes with healing activity: A systematic review. J. Dermatolog. Treat. 2015, 26, 465–470. [Google Scholar] [CrossRef]
- Lee, H.J.; Cho, S.H. Therapeutic effects of Korean red ginseng extract in a murine model of atopic dermatitis: Anti-pruritic and anti-inflammatory mechanism. J. Korean Med. Sci. 2017, 32, 679–687. [Google Scholar] [CrossRef]
- Jimbo, N.; Kawada, C.; Nomura, Y. Herb extracts and collagen hydrolysate improve skin damage resulting from ultraviolet-induced aging in hairless mice. Biosci. Biotechnol. Biochem. 2015, 79, 1624–1628. [Google Scholar] [CrossRef]
- Oh, Y.; Lim, H.W.; Park, K.H.; Huang, Y.H.; Yoon, J.Y.; Kim, K.; Lim, C.J. Ginsenoside Rc protects against UVB-induced photooxidative damage in epidermal keratinocytes. Mol. Med. Rep. 2017, 16, 2907–2914. [Google Scholar] [CrossRef] [Green Version]
- Jegal, J.; Park, N.J.; Jo, B.G.; Bong, S.K.; Jegal, H.; Yang, M.H.; Kim, S.N. Anti-atopic properties of gracillin isolated from Dioscorea quinqueloba on 2,4-dinitrochlorobenzene-induced skin lesions in mice. Nutrients 2018, 10, 1205. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Guo, M.; Yu, H.; Liu, D.; Lu, Z.; Lu, Y. Propionibacterium acnes related anti-inflammation and skin hydration activities of madecassoside, a pentacyclic triterpene saponin from Centella asiatica. Biosci. Biotechnol. Biochem. 2019, 83, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Ratz-Lyko, A.; Arct, J.; Pytkowska, K. Moisturizing and antiinflammatory properties of cosmetic formulations containing Centella asiatica extract. Indian J. Pharm. Sci. 2016, 78, 27–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yahara, S.; Uda, N.; Yoshio, E.; Yae, E. Steroidal alkaloid glycosides from tomato (Lycopersicon esculentum). J. Nat. Prod. 2004, 67, 500–502. [Google Scholar] [CrossRef]
- Yamanaka, T.; Vincken, J.P.; de Waard, P.; Sanders, M.; Takada, N.; Gruppen, H. Isolation, characterization, and surfactant properties of the major triterpenoid glycosides from unripe tomato fruits. J. Agric. Food Chem. 2008, 56, 11432–11440. [Google Scholar] [CrossRef] [PubMed]
- Dall’Asta, C.; Falavigna, C.; Galaverna, G.; Sforza, S.; Dossena, A.; Marchelli, R. A multiresidual method for the simultaneous determination of the main glycoalkaloids and flavonoids in fresh and processed tomato (Solanum lycopersicum L.) by LC-DAD-MS/MS. J. Sep. Sci. 2009, 32, 3664–3671. [Google Scholar] [CrossRef] [PubMed]
- Slimestad, R.; Verheul, M. Review of flavonoids and other phenolics from fruits of different tomato (Lycopersicon esculentum Mill.) cultivars. J. Sci. Food Agric. 2009, 89, 1255–1270. [Google Scholar] [CrossRef]
- Kaunda, J.S.; Zhang, Y.J. The genus Solanum: An ethnopharmacological, phytochemical and biological properties review. Nat. Prod. Bioprospect. 2019, 9, 77–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLean, W.H. Filaggrin failure-from ichthyosis vulgaris to atopic eczema and beyond. Br. J. Dermatol. 2016, 175, 4–7. [Google Scholar] [CrossRef] [Green Version]
- Kezic, S.; Jakasa, I. Filaggrin and skin barrier function. Curr. Probl. Dermatol. 2016, 49, 1–7. [Google Scholar]
- Eckert, R.L.; Yaffe, M.B.; Crish, J.F.; Murthy, S.; Rorke, E.A.; Welter, J.F. Involucrin-structure and role in envelope assembly. J. Investig. Dermatol. 1993, 100, 613–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishifuji, K.; Yoon, J.S. The stratum corneum: The rampart of the mammalian body. Vet. Dermatol. 2013, 24, 60–72. [Google Scholar] [CrossRef]
- Huang, H.C.; Chang, T.M. Ceramide 1 and ceramide 3 act synergistically on skin hydration and the transepidermal water loss of sodium lauryl sulfate-irritated skin. Int. J. Dermatol. 2008, 47, 812–819. [Google Scholar] [CrossRef]
- Hanada, K. Serine palmitoyltransferase, a key enzyme of sphingolipid metabolism. Biochim. Biophysi. Acta 2003, 1632, 16–30. [Google Scholar] [CrossRef]
- Levy, M.; Futerman, A.H. Mammalian ceramide synthases. IUBMB Life 2010, 62, 347–356. [Google Scholar] [CrossRef]
- Hamanaka, S.; Hara, M.; Nishio, H.; Otsuka, F.; Suzuki, A.; Uchida, Y. Human epidermal glucosylceramides are major precursors of stratum corneum ceramides. J. Invest. Dermatol. 2002, 119, 416–423. [Google Scholar] [CrossRef] [Green Version]
- Takeda, S.; Shimoda, H.; Takarada, T.; Imokawa, G. Strawberry seed extract and its major component, tiliroside, promote ceramide synthesis in the stratum corneum of human epidermal equivalents. PLoS ONE 2018, 13, e0205061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Tian, Y.; Wang, T.; Lin, Q.; Feng, X.; Jiang, Q.; Liu, Y.; Chen, D. Role of the p-coumaroyl moiety in the antioxidant and cytoprotective effects of flavonoid glycosides: Comparison of astragalin and tiliroside. Molecules 2017, 22, 1165. [Google Scholar] [CrossRef] [Green Version]
- Fluhr, J.W.; Feingold, K.R.; Elias, P.M. Transepidermal water loss reflects permeability barrier status: Validation in human and rodent in vivo and ex vivo models. Exp. Dermatol. 2006, 15, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Seidenari, S.; Giusti, G. Objective assessment of the skin of children affected by atopic dermatitis: A study of pH, capacitance and TEWL in eczematous and clinically uninvolved skin. Acta Derm. Venereol. 1995, 75, 429–433. [Google Scholar]
- Tezuka, T.; Fang, K.T.; Yamaura, T.; Masaki, H.; Sakon, K.; Suzuki, K. Changes of TEWL value at various skin diseases. Ski. Res. 1989, 31, 153–156. [Google Scholar]
- Alexander, H.; Brown, S.; Danby, S.; Flohr, C. Research techniques made simple: Transepidermal water loss measurement as a research tool. J. Investig. Dermatol. 2018, 138, 2295–2300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joly-Tonetti, N.; Ondet, T.; Monshouwer, M.; Stamatas, G.N. EGFR inhibitors switch keratinocytes from a proliferative to a differentiative phenotype affecting epidermal development and barrier function. BMC Cancer 2021, 21, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, S.; Jeena, G.S.; Shukla, R.K. Recent advances in steroidal saponins biosynthesis and in vitro production. Planta 2018, 248, 519–544. [Google Scholar] [CrossRef]
- Masukawa, Y.; Narita, H.; Shimizu, E.; Kondo, N.; Sugai, Y.; Oba, T.; Homma, R.; Ishikawa, J.; Takagi, Y.; Kitahara, T.; et al. Characterization of overall ceramide species in human stratum corneum. J. Lipid Res. 2008, 49, 1466–1476. [Google Scholar] [CrossRef] [Green Version]
- Imokawa, G.; Abe, A.; Jin, K.; Higaki, Y.; Kawashima, M.; Hidano, A. Decreased level of ceramides in stratum corneum of atopic dermatitis: An etiologic factor in atopic dry skin? J. Investig. Dermatol. 1991, 96, 523–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akimoto, K.; Yoshikawa, N.; Higaki, Y.; Kawashima, M.; Imokawa, G. Quantitative analysis of stratum corneum lipids in xerosis and asteatotic eczema. J. Dermatol. 1993, 20, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Shimotoyodome, Y.; Tsujimura, H.; Ishikawa, J.; Fujimura, T.; Kitahara, T. Variations of ceramide profile in different regions of the body of Japanese females. J. Jpn. Cos. Sci. Soc. 2014, 38, 3–8. [Google Scholar]
- Takeda, S.; Terazawa, S.; Shimoda, H.; Imokawa, G. β-Sitosterol 3-O-D-glucoside increases ceramide levels in the stratum corneum via the up-regulated expression of ceramide synthase-3 and glucosylceramide synthase in a reconstructed human epidermal keratinization model. PLoS ONE 2021, 16, e0248150. [Google Scholar] [CrossRef] [PubMed]
- Takagi, Y.; Kriehuber, E.; Imokawa, G.; Elias, P.M.; Holleran, W.M. β-Glucocerebrosidase activity in mammalian stratum corneum. J. Lipid Res. 1999, 40, 861–869. [Google Scholar] [CrossRef]
- Tafesse, F.G.; Ternes, P.; Holthuis, J.C. The multigenic sphingomyelin synthase family. J. Biol. Chem. 2006, 281, 29421–29425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, R.W.; Canals, D.; Hannun, Y.A. Roles and regulation of secretory and lysosomal acid sphingomyelinase. Cell Signal. 2009, 21, 836–846. [Google Scholar] [CrossRef] [Green Version]
- James, Q.; Kimberly, C. Topical corticosteroid application and the structural and functional integrity of the epidermal barrier. Clin. Aesthet. Dermatol. 2013, 6, 20–27. [Google Scholar]
- Yoshikawa, H.; Tasaka, K. Anti-allergic action of glucocorticoids: Comparison with immunosuppressive and anti-inflammatory effects. Cur. Med. Chem. 2003, 2, 37–50. [Google Scholar] [CrossRef]
- Matsuda, H.; Li, Y.; Murakami, T.; Ninomiya, K.; Yamahara, J.; Yoshikawa, M. Effects of escins Ia, Ib, IIa, and IIb from horse chestnut, the seeds of Aesculus hippocastanum L., on acute inflammation in animals. Biol. Pharm. Bull. 1997, 20, 1092–1095. [Google Scholar] [CrossRef] [Green Version]
- Shehu, A.; Olurish, T.O.; Zezi, A.U.; Ahmed, A. Saponin and flavonoid-rich fractions of Laggera aurita Linn. F. produce central analgesia in murine models of pain. Niger. J. Pharm. Sci. 2016, 15, 60–69. [Google Scholar]
- Leung, K.W.; Cheng, Y.K.; Mak, N.K.; Chan, K.K.; Fan, T.P.; Wong, R.N. Signaling pathway of ginsenoside-Rg1 leading to nitric oxide production in endothelial cells. FEBS Lett. 2006, 580, 3211–3216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.J.; Chung, E.; Lee, K.Y.; Lee, Y.H.; Huh, B.; Lee, S.K. Ginsenoside-Rg1, one of the major active molecules from Panax ginseng, is a functional ligand of glucocorticoid receptor. Mol. Cell. Endocrinol. 1997, 133, 135–140. [Google Scholar] [CrossRef]
- Yokosuka, A.; Mimaki, Y. Steroidal saponins from the whole plants of Agave utahensis and their cytotoxic activity. Phytochemistry 2009, 70, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.Y.; Hashimoto, F.; Yahara, S.; Nohara, T.; Yoshida, N. Steroidal glycosides from Solanum dulcamara. Chem. Pharm. Bull. 1994, 42, 707–709. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, Y.; Shinoda, D.; Nakamaru, A.; Kamohara, K.; Sakagami, H.; Mimaki, Y. Steroidal glycosides from Convallaria majalis whole plants and their cytotoxic activity. Int. J. Mol. Sci. 2017, 18, 2358. [Google Scholar] [CrossRef] [Green Version]
- Tsvetkov, D.; Dmitrenok, A.; Tsvetkov, Y.; Chizhov, A.; Nifantiev, N. Polyphenol components of the knotwood extracts of Salix caprea L. Russ. Chem. Bull. 2020, 69, 2390–2395. [Google Scholar] [CrossRef]
- Wei, Y.; Xie, Q.; Fisher, D.; Sutherland, I.A. Separation of patuletin-3-O-glucoside, astragalin, quercetin, kaempferol and isorhamnetin from Flaveria bidentis (L.) Kuntze by elution-pump-out high-performance counter-current chromatography. J. Chromatogr. A 2011, 1218, 6206–6211. [Google Scholar] [CrossRef]
- Nohara, T.; Ito, Y.; Seike, H.; Komori, T.; Moriyama, M.; Gomita, Y.; Kawasaki, T. Study on the constituents of Paris quadriforia L. Chem. Pharm. Bull. 1982, 30, 1851–1856. [Google Scholar] [CrossRef]
- Biruk, S.; Kaleab, A.; Raghavendra, Y. Radical scavenging activities of the leaf extracts and a flavonoid glycoside isolated from Cineraria abyssinica Sch. Bip. Exa. Rich. J. Appl. Pharm. Sci. 2012, 2, 44–49. [Google Scholar]
- Kobayashi, Y.; Nakano, Y.; Inayama, K.; Sakai, A.; Kamiya, T. Dietary intake of the flower extracts of German chamomile (Matricaria recutita L.) inhibited compound 48/80-induced itch-scratch responses in mice. Phytomedicine 2003, 10, 657–664. [Google Scholar] [CrossRef]
- Lee, H.K.; Park, S.B.; Chang, S.Y.; Jung, S.J. Antipruritic effect of curcumin on histamine-induced itching in mice. Korean J. Physiol. Pharmacol. 2018, 22, 547–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimoda, H.; Tanaka, J.; Yamada, E.; Morikawa, T.; Kasajima, N.; Yoshikawa, M. Anti type I allergic property of Japanese butterbur extract and its mast cell degranulation inhibitory ingredients. J. Agric. Food. Chem. 2006, 54, 2915–2920. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Relative mRNA Expression | |||||
|---|---|---|---|---|---|
| Filaggrin | Involucrin | SPT2 | CerS3 | GCS | |
| Control | 1.00 ± 0.03 | 1.00 ± 0.01 | 1.00 ± 0.02 | 1.00 ± 0.02 | 1.00 ± 0.01 |
| TSE | 1.42 ± 0.06 | 1.56 ± 0.12 ** | 1.49 ± 0.07 * | 1.78 ± 0.04 ** | 1.62 ± 0.13 ** |
| Lycoperoside A (1) | 1.47 ± 0.16 | 0.91 ± 0.04 | 0.98 ± 0.05 | 1.26 ± 0.04 | 1.00 ± 0.04 |
| Lycoperoside C (2) | 1.91 ± 0.26 ** | 1.59 ± 0.14 ** | 1.70 ± 0.21 ** | 1.72 ± 0.07 ** | 1.32 ± 0.01 * |
| Lycoperoside H (3) | 1.56 ± 0.15 | 1.67 ± 0.07 ** | 1.45 ± 0.05 | 1.50 ± 0.17 ** | 1.47 ± 0.11 ** |
| Allopregnenolone 3-O-β-solatrioside (4) | 1.57 ± 0.14 | 1.40 ± 0.06 * | 1.01 ± 0.17 | 1.20 ± 0.06 | 1.01 ± 0.03 |
| Tomatoside A (6) | 1.31 ± 0.11 | 0.96 ± 0.04 | 1.35 ± 0.09 | 1.33 ± 0.07 * | 1.66 ± 0.15 ** |
| Tigogenin 3-O-β-solatrioside (7) | 1.30 ± 0.13 | 1.03 ± 0.12 | 1.59 ± 0.28 * | 1.18 ± 0.05 | 1.58 ± 0.04 ** |
| 22α-Methoxytomatoside A (8) | 1.50 ± 0.09 ** | 1.21 ± 0.08 | 1.37 ± 0.09 | 1.32 ± 0.08 * | 1.28 ± 0.08 |
| Naringenin (5) | 1.30 ± 0.11 | 1.14 ± 0.12 | 1.84 ± 0.09 * | 1.09 ± 0.08 | 0.83 ± 0.07 |
| Astragalin (9) | 1.07 ± 0.09 | 1.16 ± 0.02 | 1.82 ± 0.09 ** | 1.18 ± 0.09 | 0.81 ± 0.03 |
| Rutin (10) | 0.80 ± 0.11 | 1.01 ± 0.13 | 1.03 ± 0.03 | 0.66 ± 0.09 ** | 0.81 ± 0.05 * |
| Quercetin 3-O-β-cellobioside (11) | 0.72 ± 0.01 * | 0.82 ± 0.12 | 0.79 ± 0.02 * | 0.56 ± 0.03 ** | 0.62 ± 0.03 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takeda, S.; Miyasaka, K.; Shrestha, S.; Manse, Y.; Morikawa, T.; Shimoda, H. Lycoperoside H, a Tomato Seed Saponin, Improves Epidermal Dehydration by Increasing Ceramide in the Stratum Corneum and Steroidal Anti-Inflammatory Effect. Molecules 2021, 26, 5860. https://doi.org/10.3390/molecules26195860
Takeda S, Miyasaka K, Shrestha S, Manse Y, Morikawa T, Shimoda H. Lycoperoside H, a Tomato Seed Saponin, Improves Epidermal Dehydration by Increasing Ceramide in the Stratum Corneum and Steroidal Anti-Inflammatory Effect. Molecules. 2021; 26(19):5860. https://doi.org/10.3390/molecules26195860
Chicago/Turabian StyleTakeda, Shogo, Kenchi Miyasaka, Sarita Shrestha, Yoshiaki Manse, Toshio Morikawa, and Hiroshi Shimoda. 2021. "Lycoperoside H, a Tomato Seed Saponin, Improves Epidermal Dehydration by Increasing Ceramide in the Stratum Corneum and Steroidal Anti-Inflammatory Effect" Molecules 26, no. 19: 5860. https://doi.org/10.3390/molecules26195860