
Maternal Environmental Light Conditions Affect the Morphological Allometry and Dispersal Potential of Acer palmatum Samaras

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
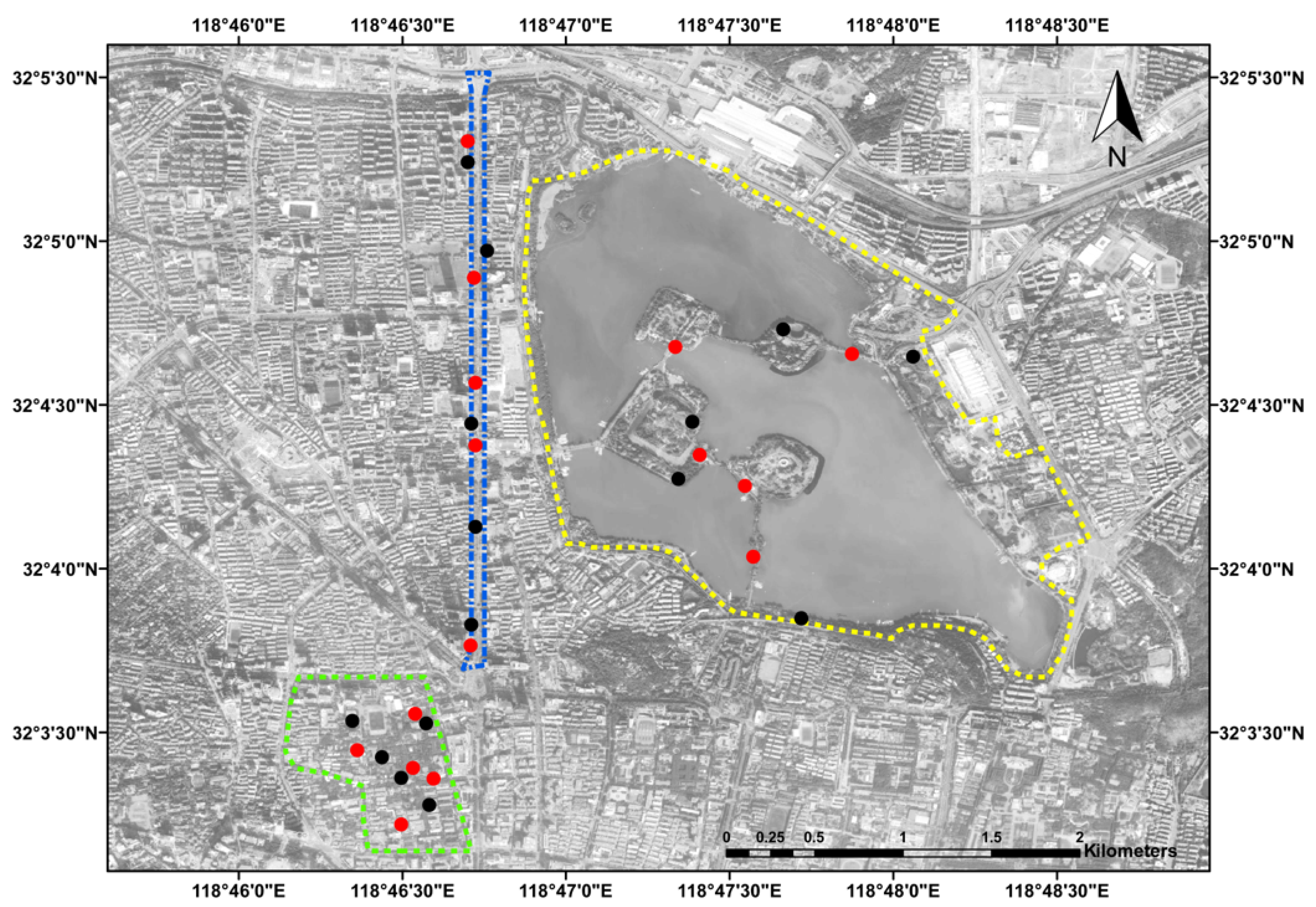
2.2. Field Collection
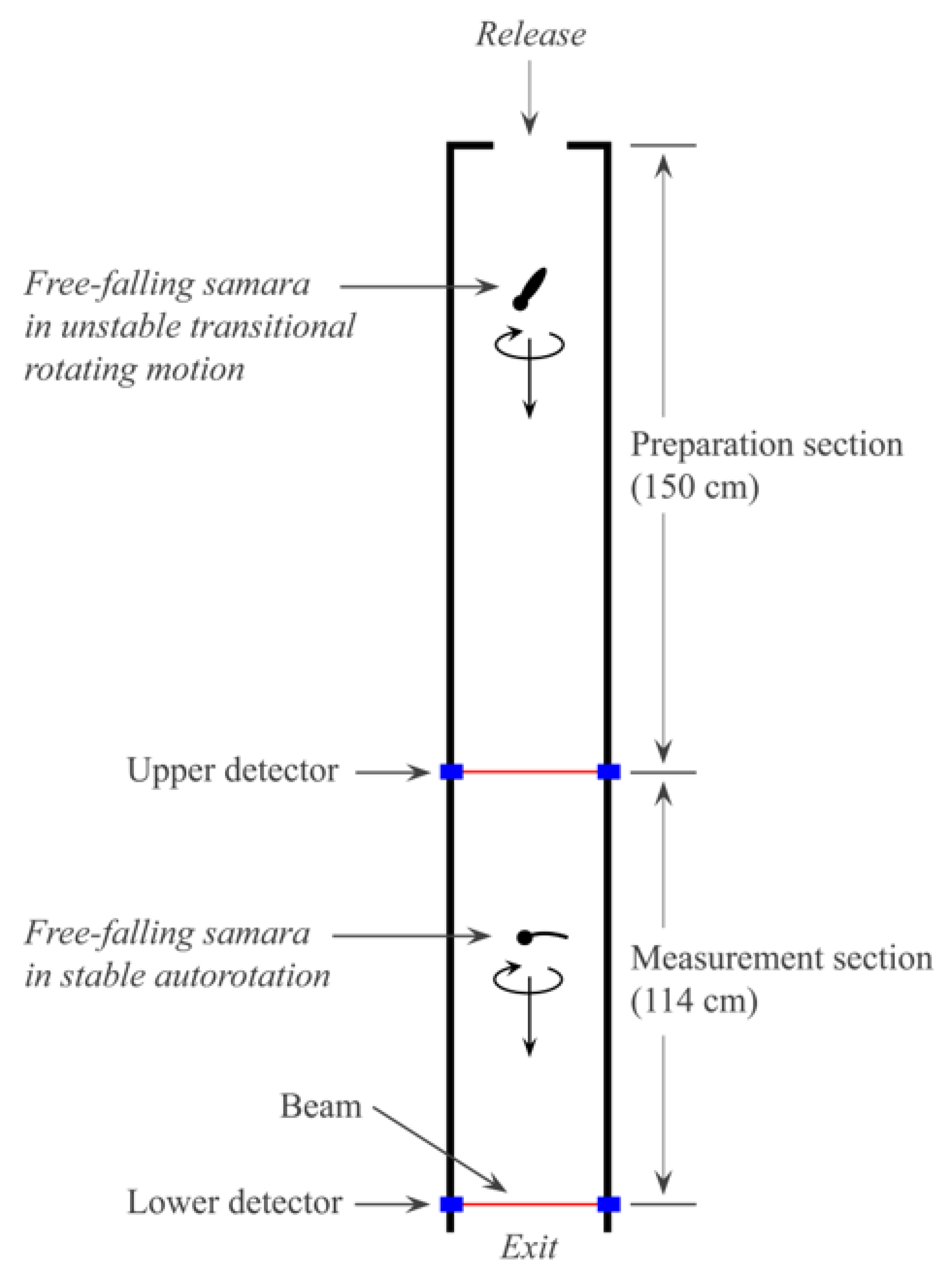
2.3. Measurements
2.4. Statistical Analyses
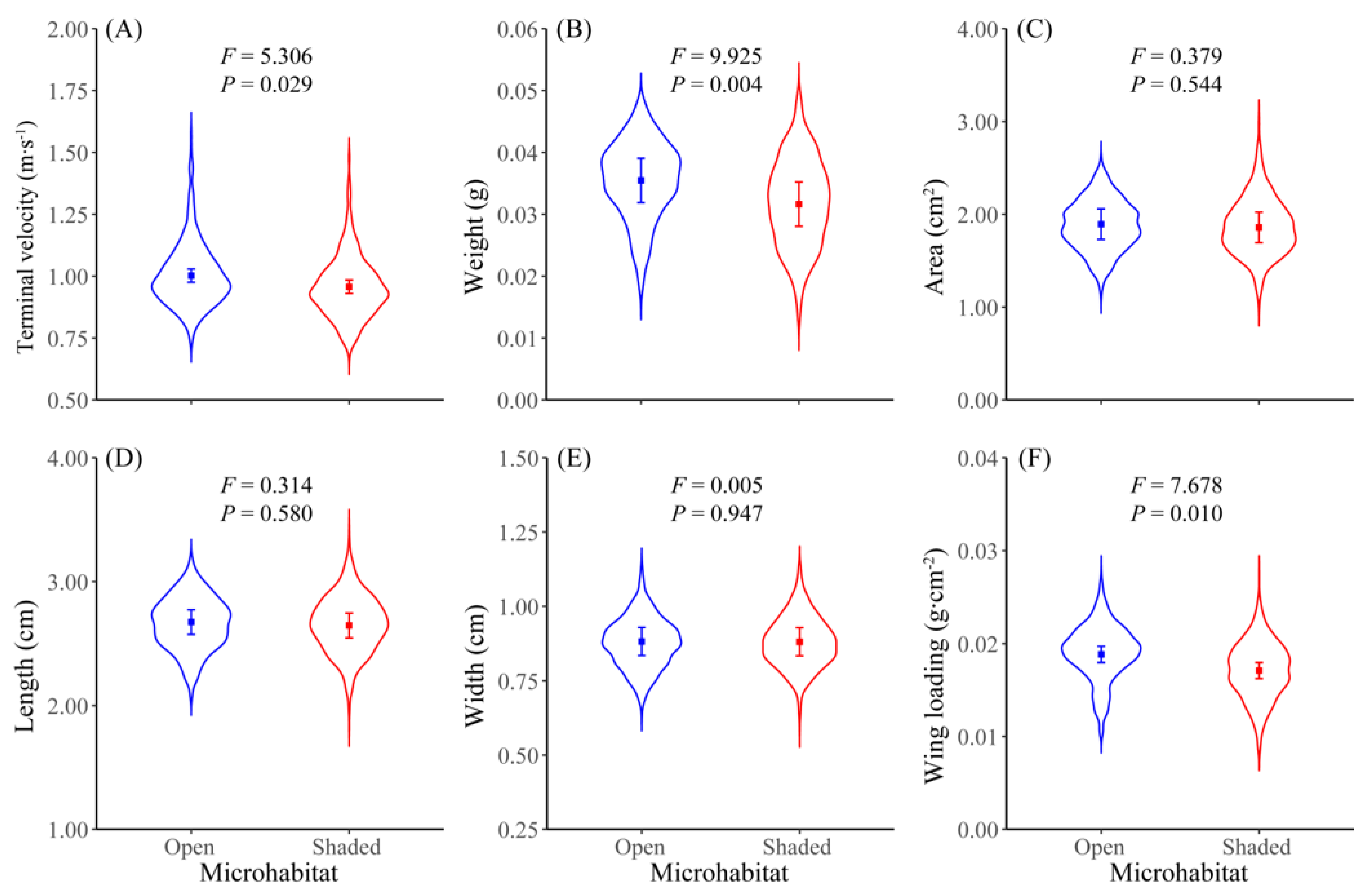
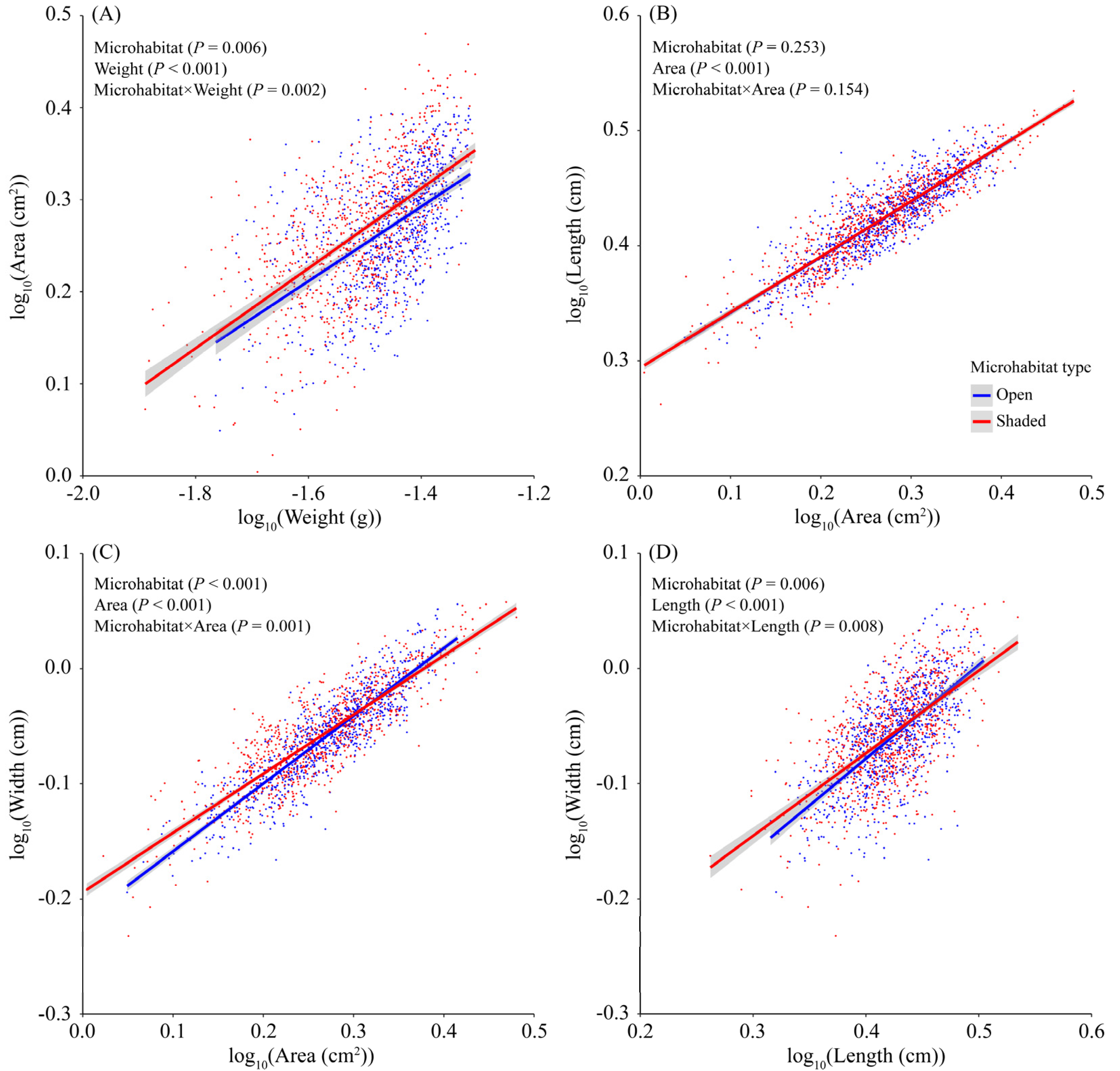
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nathan, R. Long-distance dispersal of plants. Science 2006, 313, 786–788. [Google Scholar] [CrossRef] [Green Version]
- Tackenberg, O.; Poschlod, P.; Bonn, S. Assessment of wind dispersal potential in plant species. Ecol. Monogr. 2003, 73, 191–205. [Google Scholar] [CrossRef]
- Cousens, R.; Dytham, C.; Law, R. Dispersal in Plants: A Population Perspective; Oxford University Press: New York, NY, USA, 2008. [Google Scholar]
- Harper, J.L.; Lovell, P.H.; Moore, K.G. The shapes and sizes of seeds. Annu. Rev. Ecol. Syst. 1977, 1, 327–356. [Google Scholar] [CrossRef]
- Snell, R.S.; Beckman, N.G.; Fricke, E.; Loiselle, B.A.; Carvalho, C.S.; Jones, L.R.; Lichti, N.I.; Lustenhouwer, N.; Schreiber, S.J.; Strickland, C.; et al. Consequences of intraspecific variation in seed dispersal for plant demography, communities, evolution and global change. AoB PLANTS 2019, 11, plz016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howe, H.F.; Smallwood, J. Ecology of seed dispersal. Annu. Rev. Ecol. Syst. 1982, 13, 201–228. [Google Scholar] [CrossRef]
- Nathan, R.; Katul, G.G.; Horn, H.S.; Thomas, S.M.; Oren, R.; Avissar, R.; Pacala, S.W.; Levin, S.A. Mechanisms of long-distance dispersal of seeds by wind. Nature 2002, 418, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Levine, J.M.; Murrell, D.J. The community-level consequences of seed dispersal patterns. Annu. Rev. Ecol. Syst. 2003, 34, 549–574. [Google Scholar] [CrossRef] [Green Version]
- Nathan, R.; Muller-Landau, H.C. Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends Ecol. Evol. 2000, 15, 278–285. [Google Scholar] [CrossRef]
- Saatkamp, A.; Cochrane, A.; Commander, L.; Guja, L.K.; Jimenez-Alfaro, B.; Larson, J.; Nicotra, A.; Poschlod, P.; Silveira, F.A.O.; Cross, A.T.; et al. A research agenda for seed-trait functional ecology. New Phytol. 2019, 221, 1764–1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teller, B.J.; Zhang, R.; Shea, K. Seed release in a changing climate: Initiation of movement increases spread of an invasive species under simulated climate warming. Divers. Distrib. 2016, 22, 708–716. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.S.; Cantrell, R.S.; Cosner, C.; Hartig, F.; Hastings, A.; Rogers, H.S.; Schupp, E.W.; Shea, K.; Teller, B.J.; Yu, X.; et al. Rapid changes in seed dispersal traits may modify plant responses to global change. AoB PLANTS 2019, 11, plz020. [Google Scholar] [CrossRef] [Green Version]
- Bacles, C.F.E.; Lowe, A.J.; Ennos, R.A. Effective seed dispersal across a fragmented landscape. Science 2006, 311, 628. [Google Scholar] [CrossRef] [Green Version]
- Wyse, S.V.; Hulme, P.E.; Holland, E.P. Partitioning intraspecific variation in seed dispersal potential using a low-cost method for rapid estimation of samara terminal velocity. Methods Ecol. Evol. 2019, 10, 1298–1307. [Google Scholar] [CrossRef]
- Schupp, E.W.; Zwolak, R.; Jones, L.R.; Snell, R.S.; Beckman, N.G.; Aslan, C.; Cavazos, B.R.; Effiom, E.; Fricke, E.C.; Montaño-Centellas, F.; et al. Intrinsic and extrinsic drivers of intraspecific variation in seed dispersal are diverse and pervasive. AoB PLANTS 2019, 11, plz067. [Google Scholar] [CrossRef] [PubMed]
- Levin, S.A.; Muller-Landau, H.C. The evolution of dispersal and seed size in plant communities. Evol. Ecol. Res. 2000, 2, 409–435. [Google Scholar]
- de Casas, R.R.; Willis, C.G.; Donohue, K. Plant dispersal phenotypes: A seed perspective of maternal habitat selection. In Dispersal Ecology and Evolution; Clobert, J., Baguette, M., Benton, T.G., Bullock, J.M., Eds.; Oxford University Press: New York, NY, USA, 2012; pp. 171–184. [Google Scholar]
- Donohue, K. Completing the cycle: Maternal effects as the missing link in plant life histories. Philos. Trans. R. Soc. B 2009, 364, 1059–1074. [Google Scholar] [CrossRef] [Green Version]
- Roach, D.A.; Wulff, R.D. Maternal effects in plants. Annu. Rev. Ecol. Syst. 1987, 18, 209–235. [Google Scholar] [CrossRef]
- Martorell, C.; Martínez-López, M. Informed dispersal in plants: Heterosperma pinnatum (Asteraceae) adjusts its dispersal mode to escape from competition and water stress. Oikos 2014, 123, 225–231. [Google Scholar] [CrossRef]
- Teller, B.J.; Campbell, C.; Shea, K. Dispersal under duress: Can stress enhance the performance of a passively dispersed species? Ecology 2014, 95, 2694–2698. [Google Scholar] [CrossRef] [Green Version]
- Willson, M.F.; Traveset, A. The ecology of seed dispersal. In Seeds: The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CABI Publishing: New York, NY, USA, 2000; pp. 85–110. [Google Scholar]
- Augspurger, C.K. Morphology and dispersal potential of wind-dispersed diaspores of neotropical trees. Am. J. Bot. 1986, 73, 353–363. [Google Scholar] [CrossRef]
- Willson, M.F.; Rice, B.L.; Westoby, M. Seed dispersal spectra: A comparison of temperate plant communities. J. Veg. Sci. 1990, 1, 547–562. [Google Scholar] [CrossRef]
- Song, Y.-B.; Shen-Tu, X.-L.; Dong, M. Intraspecific variation of samara dispersal traits in the endangered tropical tree Hopea hainanensis (Dipterocarpaceae). Front. Plant Sci. 2020, 11, 599764. [Google Scholar] [CrossRef] [PubMed]
- Matlack, G.R. Diaspore size, shape, and fall behavior in wind-dispersed plant species. Am. J. Bot. 1987, 74, 1150–1160. [Google Scholar] [CrossRef]
- Lorenz, R.D. Spinning Flight: Dynamics of Frisbees, Boomerangs, Samaras, and Skipping Stones; Springer: New York, NY, USA, 2006. [Google Scholar]
- Lee, S.J.; Lee, E.J.; Sohn, M.H. Mechanism of autorotation flight of maple samaras (Acer palmatum). Exp. Fluids 2014, 55, 1718. [Google Scholar] [CrossRef]
- Lentink, D.; Dickson, W.B.; van Leeuwen, J.L.; Dickinson, M.H. Leading-edge vortices elevate lift of autorotating plant seeds. Science 2009, 324, 1438–1440. [Google Scholar] [CrossRef] [Green Version]
- Arranz, G.; Gonzalo, A.; Uhlmann, M.; Flores, O.; García-Villalba, M. A numerical study of the flow around a model winged seed in auto-Rotation. Flow Turbul. Combust. 2018, 101, 477–497. [Google Scholar] [CrossRef] [Green Version]
- Norberg, R.Å. Autorotation, self-stability, and structure of single-winged fruits and seeds (samaras) with comparative remarks on animal flight. Biol. Rev. 1973, 48, 561–596. [Google Scholar] [CrossRef]
- Ortega-Jimenez, V.M.; Kim, N.S.-W.; Dudley, R. Superb autorotator: Rapid decelerations in impulsively launched samaras. J. R. Soc. Interface 2019, 16, 20180456. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, K.; Azuma, A. The autorotation boundary in the flight of samaras. J. Theor. Biol. 1997, 185, 313–320. [Google Scholar] [CrossRef]
- McCutchen, C.W. The spinning rotation of ash and tulip tree samaras. Science 1977, 197, 691–692. [Google Scholar] [CrossRef]
- Green, D.S. The terminal velocity and dispersal of spinning samaras. Am. J. Bot. 1980, 67, 1218–1224. [Google Scholar] [CrossRef]
- Sipe, T.W.; Linnerooth, A.R. Intraspecific variation in samara morphology and flight behavior in Acer saccharinum (Aceraceae). Am. J. Bot. 1995, 82, 1412–1419. [Google Scholar] [CrossRef]
- Peroni, P.A. Seed size and dispersal potential of Acer rubrum (Aceraceae) samaras produced by populations in early and late successional environments. Am. J. Bot. 1994, 81, 1428–1434. [Google Scholar] [CrossRef]
- Donohue, K. Seed dispersal as a maternally influenced character: Mechanistic basis of maternal effects and selection on maternal characters in an annual plant. Am. Nat. 1999, 154, 674–689. [Google Scholar] [CrossRef] [Green Version]
- Barstow, M.; Crowley, D. Acer palmatum. The IUCN Red List of Threatened Species 2017; 2017; e.T193845A2285627. [Google Scholar] [CrossRef]
- Xu, Z.; Chen, Y.; de Jong, P.C.; Oterdoom, H.J.; Chang, C.-S. Aceraceae. In Flora of China, Volume 11: Oxalidaceae through Aceraceae; Wu, C.Y., Raven, P.H., Eds.; Missouri Botanical Garden Press: St. Louis, MO, USA, 2008; pp. 515–553. [Google Scholar]
- Hurlbert, S.H. Pseudoreplication and the design of ecological field experiments. Ecol. Monogr. 1984, 54, 187–211. [Google Scholar] [CrossRef] [Green Version]
- Tanabata, T.; Shibaya, T.; Hori, K.; Ebana, K.; Yano, M. SmartGrain: High-throughput phenotyping software for measuring seed shape through image analysis. Plant Physiol. 2012, 160, 1871–1880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Askew, A.P.; CORKER, D.; Hodkinson, D.J.; Thompson, K. A new apparatus to measure the rate of fall of seeds. Funct. Ecol. 1997, 11, 121–125. [Google Scholar] [CrossRef]
- Lee, E.-J.; Lee, S.-J. Effect of initial attitude on autorotation flight of maple samaras (Acer palmatum). J. Mech. Sci. Technol. 2016, 30, 741–747. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Tremblay, A.; Ransijn, J. LMERConvenienceFunctions: Model Selection and Post-Hoc Analysis for (G)LMER Models. R package version 3.0. Available online: https://CRAN.R-project.org/package=LMERConvenienceFunctions (accessed on 6 October 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Fowells, H.A. Silvics of Forest Trees of the United States: Agriculture Handbook No. 271; U.S. Department of Agriculture: Washington, DC, USA, 1965.
- Weiner, J.; Martinez, S.; Muller-Scharer, H.; Stoll, P.; Schmid, B. How important are environmental maternal effects in plants? A study with Centaurea maculosa. J. Ecol. 1997, 85, 133–142. [Google Scholar] [CrossRef]
- Chen, B.J.W.; Hajiboland, R.; Bahrami-Rad, S.; Moradtalab, N.; Anten, N.P.R. Presence of belowground neighbors activates defense pathways at the expense of growth in tobacco plants. Front. Plant Sci. 2019, 10, 751. [Google Scholar] [CrossRef]
- Padilla, F.M.; Mommer, L.; de Caluwe, H.; Smit-Tiekstra, A.E.; Wagemaker, C.A.M.; Ouborg, N.J.; de Kroon, H. Early root overproduction not triggered by nutrients decisive for competitive success belowground. PLoS ONE 2013, 8, e55805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fracheboud, Y.; Luquez, V.; Björkén, L.; Sjödin, A.; Tuominen, H.; Jansson, S. The control of autumn senescence in european aspen. Plant Physiol. 2009, 149, 1982–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.J.W.; During, H.J.; Vermeulen, P.J.; Anten, N.P.R. The presence of a below-ground neighbour alters within-plant seed size distribution in Phaseolus vulgaris. Ann. Bot. 2014, 114, 937–943. [Google Scholar] [CrossRef] [Green Version]
- Larios, E.; Venable, D.L. Maternal adjustment of offspring provisioning and the consequences for dispersal. Ecology 2015, 96, 2771–2780. [Google Scholar] [CrossRef] [PubMed]
- Goulart, M.F.; Pires Lemos Filho, J.; Lovato, M.B. Variability in fruit and seed morphology among and within populations of Plathymenia (Leguminosae-Mimosoideae) in areas of the Cerrado, the atlantic forest, and transitional sites. Plant Biol. 2006, 8, 112–119. [Google Scholar] [CrossRef]
- Planchuelo, G.; Catalán, P.; Delgado, J.A.; Murciano, A. Estimating wind dispersal potential in Ailanthus altissima: The need to consider the three-dimensional structure of samaras. Plant Biosyst. 2017, 151, 316–322. [Google Scholar] [CrossRef]
- Augspurger, C.K.; Franson, S.E. Wind dispersal of artifical fruits varying in mass, area, and morphology. Ecology 1987, 68, 27–42. [Google Scholar] [CrossRef]
- Augspurger, C.K.; Franson, S.E.; Cushman, K.C.; Muller-Landau, H.C. Intraspecific variation in seed dispersal of a Neotropical tree and its relationship to fruit and tree traits. Ecol. Evol. 2016, 6, 1128–1142. [Google Scholar] [CrossRef]
- Liang, W.; Liu, Z.; Liu, M.; Qin, X.; Baskin, C.C.; Baskin, J.M.; Xin, Z.; Wang, Z.; Su, Z.; Zhou, Q. Wing loading, not terminal velocity, is the best parameter to predict capacity of diaspores for secondary wind dispersal. J. Exp. Bot. 2020, 71, 4298–4307. [Google Scholar] [CrossRef]
- Soons, M.B.; Heil, G.W.; Nathan, R.; Katul, G.G. Determinants of long-distance seed dispersal by wind in grasslands. Ecology 2004, 85, 3056–3068. [Google Scholar] [CrossRef] [Green Version]
- Thomson, F.J.; Moles, A.T.; Auld, T.D.; Kingsford, R.T. Seed dispersal distance is more strongly correlated with plant height than with seed mass. J. Ecol. 2011, 99, 1299–1307. [Google Scholar] [CrossRef]
- Augspurger, C.K.; Franson, S.E.; Cushman, K.C. Wind dispersal is predicted by tree, not diaspore, traits in comparisons of Neotropical species. Funct. Ecol. 2017, 31, 808–820. [Google Scholar] [CrossRef] [Green Version]
- Nathan, R.; Katul, G.G.; Bohrer, G.; Kuparinen, A.; Soons, M.B.; Thompson, S.E.; Trakhtenbrot, A.; Horn, H.S. Mechanistic models of seed dispersal by wind. Theor. Ecol. 2011, 4, 113–132. [Google Scholar] [CrossRef]
- Pounden, E.; Greene, D.F.; Quesada, M.; Contreras Sánchez, J.M. The effect of collisions with vegetation elements on the dispersal of winged and plumed seeds. J. Ecol. 2008, 96, 591–598. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site | Microhabitat | BD (cm) | Height (m) | CD (m) |
|---|---|---|---|---|
| NJU 1 | Open | 8.5 ± 0.4 | 3.0 ± 0.4 | 3.6 ± 0.3 |
| Shaded | 8.4 ± 0.4 | 2.9 ± 0.2 | 3.2 ± 0.3 | |
| XWL 2 | Open | 12.6 ± 1.7 | 3.4 ± 0.2 | 3.9 ± 1.0 |
| Shaded | 12.4 ± 1.3 | 3.2 ± 0.3 | 3.4 ± 0.7 | |
| ZYR 3 | Open | 6.6 ± 0.7 | 2.7 ± 0.6 | 2.9 ± 0.4 |
| Shaded | 6.5 ± 0.9 | 2.6 ± 0.5 | 2.4 ± 0.2 |
| Model | Variable | Estimate | S.E. | D.F. | t | p |
|---|---|---|---|---|---|---|
| 1 | (Intercept) † | 0.844 | 0.031 | 1757 | 27.184 | <0.001 |
| Shaded ‡ | −0.111 | 0.037 | 26 | −2.990 | 0.006 | |
| Weight | 4.473 | 0.721 | 1757 | 6.202 | <0.001 | |
| Shaded × Weight | 2.640 | 0.989 | 1757 | 2.670 | 0.008 | |
| 2 | (Intercept) † | 1.070 | 0.035 | 1757 | 30.458 | <0.001 |
| Shaded ‡ | −0.084 | 0.049 | 26 | −1.711 | 0.099 | |
| Area | −0.036 | 0.017 | 1757 | −2.100 | 0.036 | |
| Shaded × Area | 0.021 | 0.024 | 1757 | 0.851 | 0.395 | |
| 3 | (Intercept) † | 1.106 | 0.062 | 1757 | 17.772 | <0.001 |
| Shaded ‡ | −0.121 | 0.086 | 26 | −1.403 | 0.173 | |
| Length | −0.039 | 0.023 | 1757 | −1.709 | 0.088 | |
| Shaded × Length | 0.028 | 0.032 | 1757 | 0.899 | 0.369 | |
| 4 | (Intercept) † | 1.114 | 0.05 | 1757 | 22.360 | <0.001 |
| Shaded ‡ | −0.060 | 0.072 | 26 | −0.837 | 0.410 | |
| Width | −0.126 | 0.054 | 1757 | −2.329 | 0.020 | |
| Shaded × Width | 0.017 | 0.079 | 1757 | 0.222 | 0.825 | |
| 5 | (Intercept) † | 0.424 | 0.056 | 1757 | 7.630 | <0.001 |
| Shaded ‡ | −0.156 | 0.072 | 26 | −2.152 | 0.0410 | |
| Wing-loading0.5 | 4.227 | 0.394 | 1757 | 10.732 | <0.001 | |
| Shaded × Wing-loading0.5 | 1.067 | 0.534 | 1757 | 1.998 | 0.046 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, B.J.W.; Wang, X.; Dong, Y.; During, H.J.; Xu, X.; Anten, N.P.R. Maternal Environmental Light Conditions Affect the Morphological Allometry and Dispersal Potential of Acer palmatum Samaras. Forests 2021, 12, 1313. https://doi.org/10.3390/f12101313
Chen BJW, Wang X, Dong Y, During HJ, Xu X, Anten NPR. Maternal Environmental Light Conditions Affect the Morphological Allometry and Dispersal Potential of Acer palmatum Samaras. Forests. 2021; 12(10):1313. https://doi.org/10.3390/f12101313
Chicago/Turabian StyleChen, Bin J. W., Xinyu Wang, Yuting Dong, Heinjo J. During, Xia Xu, and Niels P. R. Anten. 2021. "Maternal Environmental Light Conditions Affect the Morphological Allometry and Dispersal Potential of Acer palmatum Samaras" Forests 12, no. 10: 1313. https://doi.org/10.3390/f12101313