
Genetic Comparison of ESBL-Producing Escherichia coli from Workers and Pigs at Vietnamese Pig Farms

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
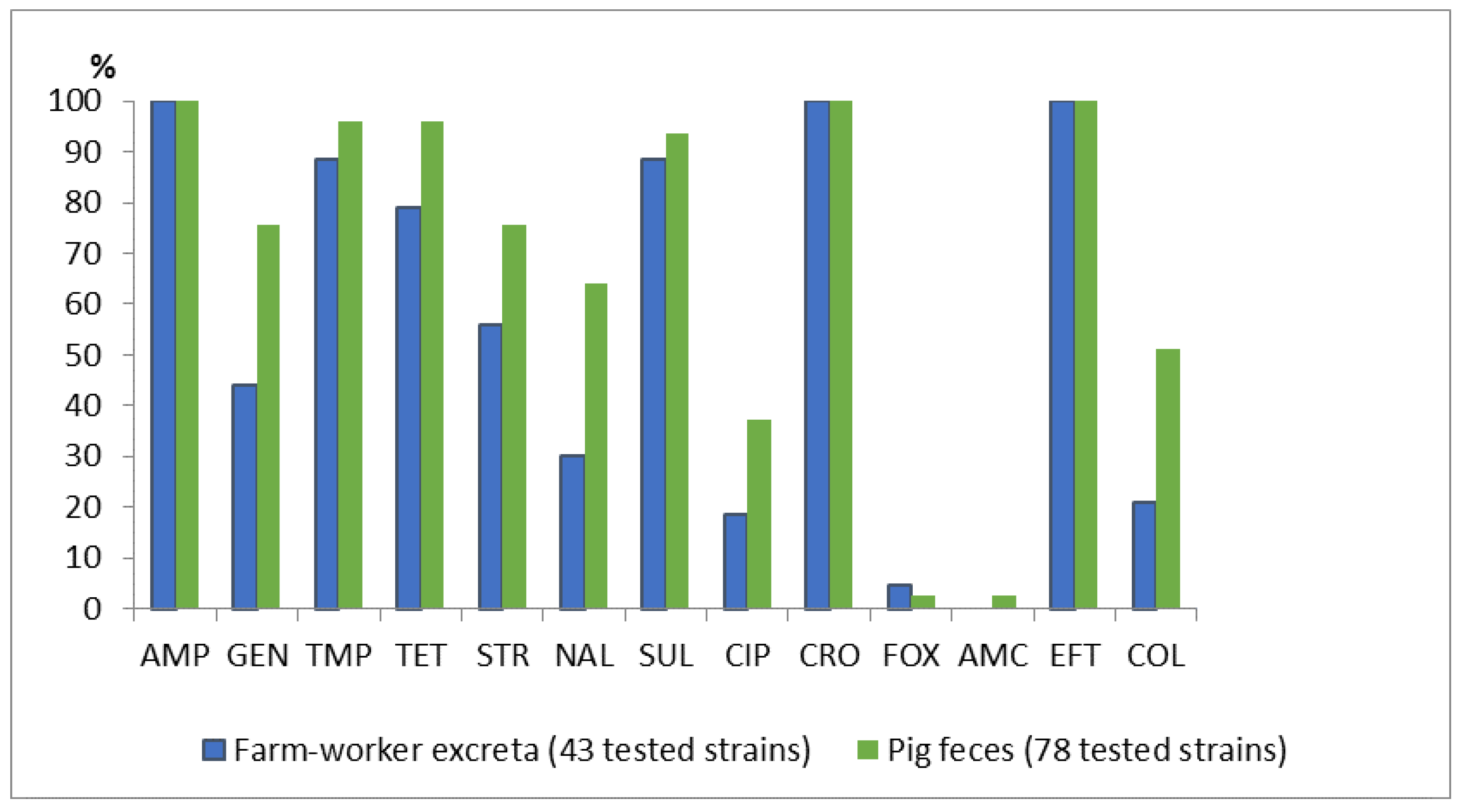
2.1. Antimicrobial Resistance in E. coli Isolated from Pigs and Farm Workers
2.2. Occurrence of ESBL-Producing E. coli in Pigs and Pig Farm Workers
2.3. Antimicrobial Resistance Genes in ESBL-Producing E. coli
2.3.1. ESBL Genes
2.3.2. Colistin Resistance Genotypes
2.3.3. Quinolone Resistance
2.3.4. Resistance Genotypes to Other Antimicrobials
2.4. Plasmid Replicons in ESBL-Producing E. coli
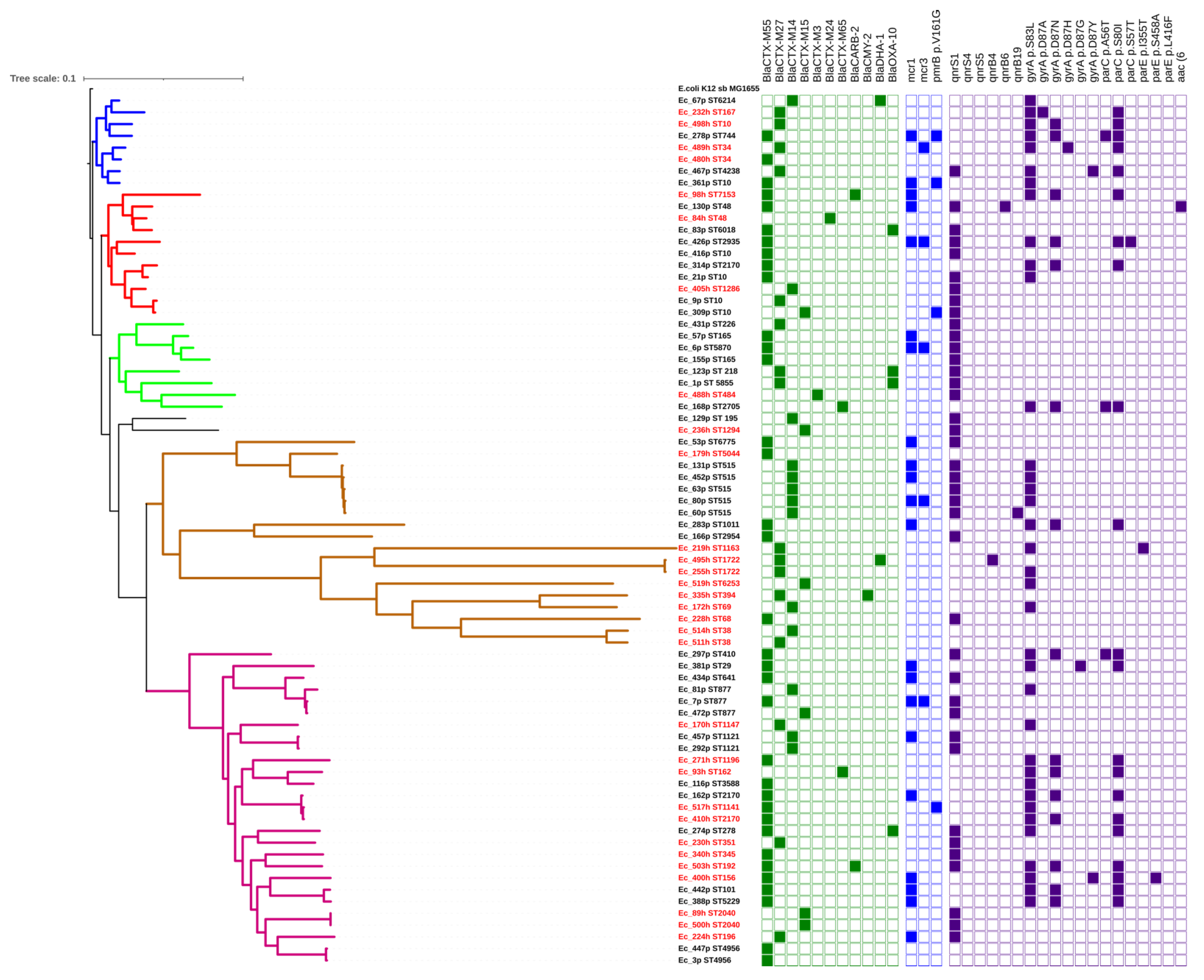
2.5. Genetic Diversity of ESBL-Producing E. coli
2.6. Virulence Genes and Serotypes
3. Discussion
4. Materials and Methods
4.1. Collection of Fecal Samples
4.2. Sampling and Isolation of E. coli
4.3. Antimicrobial Susceptibility Testing
4.4. Whole-Genome Sequencing and Sequence Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Global Action Plan on Antimicrobial Resistance; WHO: Geneva, Switzerland, 2017; pp. 1–28. [Google Scholar]
- Chereau, F.; Opatowski, L.; Tourdjman, M.; Vong, S. Risk Assessment for Antibiotic Resistance in South East Asia. BMJ 2017, 358. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5598255/ (accessed on 5 September 2017).
- Vounba, P.; Arsenault, J.; Bada-Alambédji, R.; Fairbrother, J.M. Pathogenic potential and the role of clones and plasmids in beta-lactamase-producing E. coli from chicken faeces in Vietnam. BMC Vet. Res. 2019, 15, 106. [Google Scholar] [CrossRef]
- Mburu, J.; Egelyng, H.; Ackello-ogutu, C. Household Pork Consumption Behaviour in Vietnam: Implications for pro-Smallholder Pig Value Chain Upgrading. In Proceedings of the Tropentag 2015, Berlin, Germany, 16–18 September 2015. [Google Scholar]
- Lucila, M.; Lapar, A. The REVALTER Project "Multi-Scale Assessment of Livestock Development Pathways in Vietnam"; Review of the Pig Sector in Vietnam; ILRI: Nairobi, Kenya, 2014. [Google Scholar]
- Carrique-Mas, J.J.; Choisy, M.; Van Cuong, N.; Thwaites, G.; Baker, S. An estimation of total antimicrobial usage in humans and animals in Vietnam. Antimicrob. Resist. Infect. Control 2020, 9, 16. [Google Scholar] [CrossRef] [Green Version]
- Van Cuong, N.; Nhung, N.T.; Nghia, N.H.; Mai Hoa, N.T.; Trung, N.V.; Thwaites, G.; Carrique-Mas, J. Antimicrobial Consumption in Medicated Feeds in Vietnamese Pig and Poultry Production. Ecohealth 2016, 13, 490–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. List of Critically Important Antimicrobials for Human Medicine; Who: Geneva, Switzerland, 2017; ISBN 978 92 4 150448 5. [Google Scholar]
- Nguyen, N.T.; Nguyen, H.M.; Nguyen, C.V.; Nguyen, T.V.; Nguyen, M.T.; Thai, H.Q.; Ho, M.H.; Thwaites, G.; Ngo, H.T.; Baker, S.; et al. Use of Colistin and Other Critical Antimicrobials on Pig and Chicken Farms in Southern Vietnam and Its Association with Resistance in Commensal Escherichia coli Bacteria. Appl. Environ. Microbiol. 2016, 82, 3727–3735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, S.T.T.; Bortolaia, V.; Tran, N.T.; Le, H.Q.; Dalsgaard, A. Cephalosporin-resistant Escherichia coli isolated from farm workers and pigs in northern Vietnam. Trop. Med. Int. Health 2018, 23, 415–424. [Google Scholar] [CrossRef]
- Nguyen, V.T.; Carrique-Mas, J.J.; Ngo, T.H.; Ho, H.M.; Ha, T.T.; Campbell, J.I.; Nguyen, T.N.; Hoang, N.N.; Pham, V.M.; Wagenaar, J.A.; et al. Prevalence and risk factors for carriage of antimicrobial-resistant Escherichia coli on household and small-scale chicken farms in the Mekong Delta of Vietnam. J. Antimicrob. Chemother. 2015, 70, 2144–2152. [Google Scholar] [PubMed] [Green Version]
- Mitchell, M.E.; Alders, R.; Unger, F.; Nguyen-Viet, H.; Le TT, H.; Toribio, J.A. The challenges of investigating antimicrobial resistance in Vietnam—what benefits does a One Health approach offer the animal and human health sectors? BMC Public Health 2020, 20, 1–12. Available online: https://bmcpublichealth.biomedcentral.com/articles/10.1186/s12889-020-8319-3 (accessed on 24 September 2021). [CrossRef] [PubMed] [Green Version]
- Ueda, S.; Ngan, B.T.K.; Huong, B.T.M.; Hirai, I.; Tuyen, L.D.; Yamamoto, Y. Limited Transmission of blaCTX-M-9-Type-Positive Escherichia coli between Humans and Poultry in Vietnam. Antimicrob. Agents Chemother. 2015, 59, 3574–3577. [Google Scholar] [CrossRef] [Green Version]
- Börjesson, S.; Ny, S.; Egervärn, M.; Bergström, J.; Rosengren, Å.; Englund, S.; Löfmark, S.; Byfors, S. Limited Dissemination of Extended-Spectrum β-Lactamase– and Plasmid-Encoded AmpC–Producing Escherichia coli from Food and Farm Animals, Sweden. Emerg. Infect. Dis. 2016, 22, 634–640. [Google Scholar] [CrossRef] [Green Version]
- Dorado-García, A.; Smid, J.H.; van Pelt, W.; Bonten, M.J.M.; Fluit, A.C.; van den Bunt, G.; Wagenaar, J.A.; Hordijk, J.; Dierikx, C.M.; Veldman, K.T.; et al. Molecular relatedness of ESBL/AmpC-producing Escherichia coli from humans, animals, food and the environment: A pooled analysis. J. Antimicrob. Chemother. 2018, 73, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.T.; Jamrozy, D.; Matamoros, S.; Carrique-Mas, J.J.; Ho, H.M.; Thai, Q.H.; Nguyen, T.N.M.; Wagenaar, J.A.; Thwaites, G.; Parkhill, J.; et al. Limited Contribution of Non-Intensive Chicken Farming to Esbl-Producing Escherichia Coli Colonization in Humans in Vietnam: An Epidemiological and Genomic Analysis. J. Antimicrob. Chemother. 2019, 74, 561–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludden, C.; Raven, K.E.; Jamrozy, D.; Gouliouris, T.; Blane, B.; Coll, F.; de Goffau, M.; Naydenova, P.; Horner, C.; Hernandez-Garcia, J.; et al. One Health Genomic Surveillance of Escherichia coli Demonstrates Distinct Lineages and Mobile Genetic Elements in Isolates from Humans versus Livestock. MBio 2019, 10, e02693-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; Murray, A.C.; Gajewski, A.; Sullivan, M.; Snippes, P.; Kuskowski, M.A.; Smith, K.E. Isolation and Molecular Characterization of Nalidixic Acid-Resistant Extraintestinal Pathogenic Escherichia coli from Retail Chicken Products. Antimicrob. Agents Chemother. 2003, 47, 2161–2168. [Google Scholar] [CrossRef] [Green Version]
- Spurbeck, R.R.; Dinh, P.C.; Walk, S.T.; Stapleton, A.E.; Hooton, T.M.; Nolan, L.K.; Kim, K.S.; Johnson, J.R.; Mobley, H.L.T. Escherichia coli Isolates That Carry vat, fyuA, chuA, and yfcV Efficiently Colonize the Urinary Tract. Infect. Immun. 2012, 80, 4115–4122. [Google Scholar] [CrossRef] [Green Version]
- Day, M.J.; Rodríguez, I.; van Essen-Zandbergen, A.; Dierikx, C.; Kadlec, K.; Schink, A.-K.; Wu, G.; Chattaway, M.A.; DoNascimento, V.; Wain, J.; et al. Diversity of STs, plasmids and ESBL genes among Escherichia coli from humans, animals and food in Germany, the Netherlands and the UK. J. Antimicrob. Chemother. 2016, 71, 1178–1182. [Google Scholar] [CrossRef] [Green Version]
- Flament-Simon, S.-C.; Nicolas-Chanoine, M.-H.; García, V.; Duprilot, M.; Mayer, N.; Alonso, M.P.; García-Meniño, I.; Blanco, J.E.; Blanco, M.; Blanco, J. Clonal Structure, Virulence Factor-encoding Genes and Antibiotic Resistance of Escherichia coli, Causing Urinary Tract Infections and Other Extraintestinal Infections in Humans in Spain and France during 2016. Antibiotics 2020, 9, 161. [Google Scholar] [CrossRef] [Green Version]
- Chattaway, M.A.; Jenkins, C.; Ciesielczuk, H.; Day, M.; DoNascimento, V.; Day, M.; Rodríguez, I.; van Essen-Zandbergen, A.; Schink, A.-K.; Wu, G.; et al. Evidence of Evolving Extraintestinal Enteroaggregative Escherichia coli ST38 Clone. Emerg. Infect. Dis. 2014, 20, 1935–1937. [Google Scholar] [CrossRef]
- Ahmed, S.; Olsen, J.E.; Herrero-Fresno, A. The genetic diversity of commensal Escherichia coli strains isolated from non-antimicrobial treated pigs varies according to age group. PLoS ONE 2017, 12, e0178623. [Google Scholar] [CrossRef]
- Bui, T.K.N.; Bui, T.M.H.; Ueda, S.; Le, D.T.; Yamamoto, Y.; Hirai, I. Potential transmission opportunity of CTX-M-producing Escherichia coli on a large-scale chicken farm in Vietnam. J. Glob. Antimicrob. Resist. 2018, 13, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Hoang, T.a.V.; Nguyen, T.N.H.; Ueda, S.; Le, Q.P.; Tran, T.T.N.; Nguyen, T.N.D.; Dao, T.V.K.; Tran, M.T.; Le, T.T.T.; Le, T.L.; et al. Common findings of bla CTX-M-55-encoding 104–139 kbp plasmids harbored by extended-spectrum β-lactamase-producing Escherichia coli in pork meat, wholesale market workers, and patients with urinary tract infection in Vietnam. Curr. Microbiol. 2017, 74, 203–211. [Google Scholar] [CrossRef]
- Kawahara, R.; Khong, D.T.; Le, H.V.; Phan, Q.N.; Nguyen, T.N.; Yamaguchi, T.; Kumeda, Y.; Yamamoto, Y. Prevalence Of mcr-1 Among Cefotaxime-Resistant Commensal Escherichia coli In Residents Of Vietnam. Infect. Drug Resist. 2019, 12, 3317–3325. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, T.; Ueda, S.; Huong, B.T.M.; Tuyen, L.D.; Komalamisra, C.; Kusolsuk, T.; Hirai, I.; Yamamoto, Y. Wide dissemination of extended-spectrum β-lactamase-producing Escherichia coli in community residents in the Indochinese peninsula. Infect. Drug Resist. 2015, 8, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.P.; Nguyen, T.A.D.; Le, T.H.; Tran, N.M.D.; Ngo, T.P.; Dang, V.C.; Kawai, T.; Kanki, M.; Kawahara, R.; Jinnai, M.; et al. Dissemination of Extended-Spectrum β-Lactamase-and AmpC β-Lactamase-Producing Escherichia coli within the Food Distribution System of Ho Chi Minh City, Vietnam. BioMed Res. Int. 2016. [Google Scholar] [CrossRef] [Green Version]
- Hinenoya, A.; Tran, S.T.T.; Nguyen, N.T.; Nguyen, H.C.; Nguyen, D.D.L.; Hoang, P.H.; Awasthi, S.P.; Hassan, J.; Sumimura, Y.; Yamamoto, Y.; et al. Isolation and molecular characterization of extended-spectrum β-lactamase producing Escherichia coli from industrial food animals in Mekong Delta, Vietnam. Jpn. J. Vet. Res. 2018, 66, 1–12. [Google Scholar]
- LaPlante, K.; Cusumano, J.; Tillotson, G. Colistin for the treatment of multidrug-resistant infections. Lancet Infect. Dis. 2018, 18, 1174–1175. [Google Scholar] [CrossRef]
- Teramae, M.; Osawa, K.; Shigemura, K.; Kitagawa, K.; Shirakawa, T.; Fujisawa, M.; Miyara, T. Prevalence of Quinolone Resistance of Extended-Spectrum β-Lactamase-Producing Escherichia coli with ST131-fimH30 in a City Hospital in Hyogo, Japan. Int. J. Mol. Sci. 2019, 20, 5612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peirano, G.; Pitout, J.D.D. Fluoroquinolone-Resistant Escherichia coli Sequence Type 131 Isolates Causing Bloodstream Infections in a Canadian Region with a Centralized Laboratory System: Rapid Emergence of the H30-Rx Sublineage. Antimicrob. Agents Chemother. 2014, 58, 2699–2703. [Google Scholar] [CrossRef] [Green Version]
- Matsumura, Y.; Johnson, J.R.; Yamamoto, M.; Nagao, M.; Tanaka, M.; Takakura, S.; Ichiyama, S.; Kyoto–Shiga Clinical Microbiology Study Group. Kyoto-Shiga Clinical Microbiology Study Group CTX-M-27- and CTX-M-14-producing, ciprofloxacin-resistant Escherichia coli of the H30 subclonal group within ST131 drive a Japanese regional ESBL epidemic. J. Antimicrob. Chemother. 2015, 70, 1639–1649. [Google Scholar]
- Zurfluh, K.; Abgottspon, H.; Hächler, H.; Nüesch-Inderbinen, M.; Stephan, R. Quinolone Resistance Mechanisms among Extended-Spectrum Beta-Lactamase (ESBL) Producing Escherichia coli Isolated from Rivers and Lakes in Switzerland. PLoS ONE 2014, 9, e95864. [Google Scholar]
- Nhung, N.T.; Cuong, N.V.; Thwaites, G.; Carrique-Mas, J. Antimicrobial Usage and Antimicrobial Resistance in Animal Production in Southeast Asia: A Review. Antibiotics 2016, 5, 37. [Google Scholar] [CrossRef] [Green Version]
- Mathers, A.J.; Peirano, G.; Pitout, J.D.D. The Role of Epidemic Resistance Plasmids and International High-Risk Clones in the Spread of Multidrug-Resistant Enterobacteriaceae. Clin. Microbiol. Rev. 2015, 28, 565–591. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Kawahara, R.; Fujiya, Y.; Sasaki, T.; Hirai, I.; Khong, D.T.; Nguyen, T.N.; Nguyen, B.X. Wide dissemination of colistin-resistant Escherichia coli with the mobile resistance gene mcr in healthy residents in Vietnam. J. Antimicrob. Chemother. 2019, 74, 523–524. [Google Scholar] [CrossRef]
- Bui, T.M.H.; Hirai, I.; Ueda, S.; Bui, T.K.N.; Hamamoto, K.; Toyosato, T.; Le, D.T.; Yamamoto, Y. Carriage of Escherichia coli Producing CTX-M-Type Extended-Spectrum β-Lactamase in Healthy Vietnamese Individuals. Antimicrob. Agents Chemother. 2015, 59, 6611–6614. [Google Scholar] [CrossRef] [Green Version]
- Zając, M.; Sztromwasser, P.; Bortolaia, V.; Leekitcharoenphon, P.; Cavaco, L.M.; Ziȩtek-Barszcz, A.; Hendriksen, R.S.; Wasyl, D. Occurrence and Characterization of mcr-1-Positive Escherichia coli Isolated From Food-Producing Animals in Poland, 2011–2016. Front. Microbiol. 2019, 10, 1753. [Google Scholar] [CrossRef] [Green Version]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance—PubMed. Clin. Microbiol. Infect 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, S.; Le, T.D.; Vien, M.Q.; Van Dang, C.; Yamamoto, Y. Prevalence of extended-spectrum β-lactamase-producing Escherichia coli and residual antimicrobials in the environment in Vietnam. Anim. Health Res. Rev. 2017, 18, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Vounba, P.; Rhouma, M.; Arsenault, J.; Bada Alambédji, R.; Fravalo, P.; Fairbrother, J.M. Prevalence of colistin resistance and mcr-1/mcr-2 genes in extended-spectrum β-lactamase/AmpC-producing Escherichia coli isolated from chickens in Canada, Senegal and Vietnam. J. Glob. Antimicrob. Resist. 2019, 19, 222–227. [Google Scholar] [CrossRef]
- Coyne, L.; Arief, R.; Benigno, C.; Giang, V.N.; Huong, L.Q.; Jeamsripong, S.; Kalpravidh, W.; McGrane, J.; Padungtod, P.; Patrick, I.; et al. Characterizing Antimicrobial Use in the Livestock Sector in Three South East Asian Countries (Indonesia, Thailand, and Vietnam). Antibiotics 2019, 8, 33. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, K.V.; Thi Do, N.T.; Chandna, A.; Nguyen, T.V.; Pham, C.V.; Doan, P.M.; Nguyen, A.Q.; Thi Nguyen, C.K.; Larsson, M.; Escalante, S.; et al. Antibiotic use and resistance in emerging economies: A situation analysis for Viet Nam. BMC Public Health 2013, 13, 1158. [Google Scholar] [CrossRef] [Green Version]
- The List of Veterinary Drugs has been Licensed for Circulation in Vietnam until March 2019 (Vietnamese). Available online: http://cucthuy.gov.vn/Pages/danh-muc-thuoc-thu-y-da-duoc-cap-giay-chung-nhan-luu-hanh-tai-viet-nam-.aspx (accessed on 3 May 2020).
- Yang, S.; Deng, W.; Liu, S.; Yu, X.; RazaMustafa, G.; Chen, S.; He, L.; Ao, X.; Yang, Y. Presence of Heavy Metal Resistance Genes in Escherichia coli and Salmonella, and Analysis of Resistance Gene Structure in E. coli E308. J. Glob. Antimicrob. Resist. 2020, 21, 420–426. [Google Scholar] [CrossRef]
- Comparative analysis of the susceptibility to biocides and heavy metals of extended-spectrum β-lactamase-producing Escherichia coli isolates of human and avian origin, Germany. Diagn. Microbiol. Infect. Dis. 2017, 88, 88–92. [CrossRef]
- Yu, Z.; Gunn, L.; Wall, P.; Fanning, S. Antimicrobial resistance and its association with tolerance to heavy metals in agriculture production. Food Microbiol. 2017, 64, 23–32. [Google Scholar] [CrossRef]
- Yang, Q.E.; Agouri, S.R.; Tyrrell, J.M.; Walsh, T.R. Heavy Metal Resistance Genes Are Associated with blaNDM-1- and blaCTX-M-15-Carrying Enterobacteriaceae. Antimicrob. Agents Chemother. 2018, 62, e02642-17. [Google Scholar] [CrossRef] [Green Version]
- EUCAST: Clinical Breakpoints and Dosing of Antibiotics. Available online: https://eucast.org/clinical_breakpoints/ (accessed on 12 April 2020).
- Farmer, J.J.; Farmer, M.K.; Holmes, B. The Enterobacteriaceae: General Characteristics. In Topley & Wilson’s Microbiology and Microbial Infections; American Cancer Society: Chichester, UK, 2010; ISBN 978-0-470-68861-8. [Google Scholar]
- CLSI M100|Performance Standards for Antimicrobial Susceptibility Testing, 28th Edition. Available online: https://clsi.org/standards/products/microbiology/documents/m100/ (accessed on 12 April 2020).
- Naseer, F.; Iqbal, R.; Ikram, N.; Shoaib, M.; asad, M.J.; Mehmood, R.T.; Niazi, A.; Niazi, A.; Ishfaq, B. Phenotypic cofirmatory disc diffusion test (PCDDT), double disc synergy test (DDST), E-test OS diagnostic tool for detection of extended spectrum beta lactamase (ESΒL) producing Uropathogens. Appl. Biotechnol. Bioeng. Appl. Biotechnol 2017, 3, 344–349. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. J. Comput. Mol. Cell Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinform. Oxf. Engl. 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
- Malberg Tetzschner, A.M.; Johnson, J.R.; Johnston, B.D.; Lund, O.; Scheutz, F. In Silico Genotyping of Escherichia coli Isolates for Extraintestinal Virulence Genes by Use of Whole-Genome Sequencing Data. J. Clin. Microbiol. 2020, 58, e01269-20. Available online: https://journals.asm.org/doi/10.1128/JCM.01269-20 (accessed on 20 August 2021). [CrossRef]
- RASTtk: A Modular and Extensible Implementation of the RAST Algorithm for Building Custom Annotation Pipelines and Annotating Batches of Genomes. Sci. Rep. 2015, 5, 1–6. Available online: https://www.nature.com/articles/srep08365 (accessed on 3 May 2020).
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the Problem of Comparing Whole Bacterial Genomes across Different Sequencing Platforms. PLoS ONE 2014, 9, e104984. [Google Scholar] [CrossRef] [Green Version]
- Interactive Tree of Life (iTOL) v3: An Online Tool for the Display and Annotation of Phylogenetic and other Trees. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4987883/ (accessed on 12 April 2020).



{kind=link}
{kind=link}
| Antimicrobial | Farm Worker Feces (N = 94) | Pig Manure (N = 116) | p-Value |
|---|---|---|---|
| Number of Resistant Isolates n (%) | Number of Resistant Isolates n (%) | ||
| Ampicillin (AMP) | 68 (72.3) | 100 (86.2) | 0.02 |
| Gentamycin (GEN) | 11 (11.7) | 34 (29.3) | 0.003 |
| Trimethoprim (TMP) | 61 (64.9) | 84 (72.4) | 0.30 |
| Tetracycline (TET) | 68 (72.3) | 98 (84.5) | 0.04 |
| Streptomycin (STR) | 40 (42.6) | 65 (56.0) | 0.07 |
| Nalidixic acid (NAL) | 25 (26.6) | 32 (27.6) | 0.99 |
| Sulphonamide (SUL) | 63 (67.0) | 96 (82.8) | 0.01 |
| Ciprofloxacin (CIP) | 11 (11.7) | 17 (14.7) | 0.67 |
| Ceftriaxone (CRO) | 13 (13.8) | 1 (0.9) | 0.0005 |
| Cefoxitin (FOX) | 4 (4.3) | 0 (0.0) | |
| Amoxicillin-clavulanic acid (AMC) | 4 (4.3) | 3 (2.6) | |
| Ceftiofur (EFT) | 13 (13.8) | 1 (0.9) | 0.0005 |
| Colistin (COL) | 14 (14.9) | 44 (37.9) | 0.003 |
| Topoisomearase Gene | Mutation Region and Amino Acid Change | Number of Pig Isolates (n) (N = 43) | Number of Human Isolates (n) (N = 31) |
|---|---|---|---|
| gyrA | Ser83→Leu | 19 | 17 |
| Asp87→Ans | 9 | 6 | |
| Asp87→Ala | 1 | ||
| Asp87→Gly | 1 | ||
| Asp87→His | 1 | ||
| Asp87→Tyr | 2 | ||
| parC | Ser80→Ile | 11 | 10 |
| Ser56→Thr | 3 | ||
| Ser57→Thr | 1 | ||
| parE | Ser458→Ala | 1 | 4 |
| Iso335→Thr | 1 | ||
| Leu416→Phe | 1 |
| Strain ID | MLST | Genome Size | GC% | N50 | Phylogroup | Serotype | Virulence Genes | Plasmid Replicons |
|---|---|---|---|---|---|---|---|---|
| Ec_1 p * | ST5855 | 4,548,521 | 51.12 | 21,866 | A | H38 | ompT, terC | IncY |
| Ec_3 p | ST4956 | 4,647,525 | 51.1 | 23,906 | B1 | H37 | lpfA, terC | ND |
| Ec_6 p | ST5870 | 4,899,146 | 51.03 | 50,792 | A | O147:H40 | astA, terC, traT | IncFIA (HI1), IncFIB (K), IncFII, IncN, IncX1 |
| Ec_7 p | ST877 | 4,728,224 | 50.94 | 29,266 | B1 | O45:H10 | iss, lpfA, ompT, terC, traT, | IncFII, IncR |
| Ec_9 p | ST10 | 4,873,762 | 50.71 | 71,377 | A | H32 | terC | IncFIA (HI1), IncFIB (K), IncHI1A, IncHI1B (R27) |
| Ec_21 p | ST10 | 4,770,784 | 50.89 | 72,227 | A | O45:H45 | kpsE, kpsMII, terC | IncFIA (HI1), IncFIB (K), IncX1 |
| Ec_219 h* | ST1163 | 4,905,795 | 51.02 | 73,212 | G | H23 | iss, lpfA, senB, chuA, neuC, ompT, terC, traT | Col156, IncFIA, IncFIB (AP001918), IncFII |
| Ec_309 p | ST10 | 4,820,803 | 50.9 | 43,099 | A | H32 | iss, terC | ND |
| Ec_335 h | ST394 | 5,103,214 | 50.52 | 72,314 | D | O17/O77:H18 | aap, air, astA, eilA, lpfA, chuA, kpsE, kpsMII_K1, neuC, terC, traT | IncFII, IncFII (pHN7A8) |
| Ec_314 p | ST2170 | 5,061,254 | 50.7 | 43,068 | A | H51 | cma, iss, lpfA, fyuA, hlyF, irp2, iucC, iutA, ompT, sitA, terC, traT | IncFIB (AP001918), IncFIB (Plf82-PhagePlasmid), IncFIC (FII), IncI1-I (gamma) |
| Ec_340 h | ST345 | 4,880,303 | 50.61 | 82,846 | B1 | O8:H21 (53) | lpfA, terC | IncI1, IncY |
| Ec_116 p | ST3588 | 4,866,794 | 50.99 | 23,092 | ND | O8:H7 | iroN, iss, lpfA, mchF, tsh, cvaC, etsC, hlyF, iucC, iutA, ompT, sitA, terC, traT | IncFIB (AP001918), IncFIC (FII), IncFII (pHN7A8), IncN |
| Ec_224 h | ST196 | 5,127,497 | 50.46 | 85,977 | B1 | O8:H7 | iroN, iss, lpfA, hlyF, ompT, sitA, terC, traT | IncFIA (HI1), IncFIB (AP001918), IncFII (pCoo), IncHI1A, IncHI1B (R27), IncI2, p0111 |
| Ec_123 p | ST218 | 4,902,464 | 51 | 38,282 | A | H23 | TerC, TraT | IncFIA (HI1), IncFIB (K), IncFII, IncN, IncX1 |
| Ec_129 p | ST195 | 4,641,276 | 50.92 | 27,405 | ND | H4 | lpfA, terC | ND |
| Ec_228 h | ST68 | 4,995,853 | 50.73 | 68,496 | D | H6 | air, eilA, iss, lpfA, chuA, kpsE, kpsMII, sitA, terC, | IncFIB (K) |
| Ec_130 p | ST48 | 4,963,599 | 50.75 | 51,687 | B1 | O15:H11 | astA, ompT, terC | IncN, IncX1, p0111 |
| Ec_131 p | ST515 | 5,015,450 | 50.61 | 73,216 | B1 | H12 | fyuA, irp2, terC | IncFIA (HI1), IncHI1A, IncHI1B (R27) |
| Ec_274 p | ST278 | 4,936,780 | 50.61 | 61,512 | B1 | H7 | fim41A, lpfA, terC | IncFIB (pB171), IncFII, IncX1 |
| Ec_230 h | ST351 | 5,181,680 | 50.5 | 68,811 | B1 | O18:H7 | cma, iroN, iss, cvaC, etsC, hlyF, ompT, papC, sitA, terC | IncFIA (HI1), IncFIB (AP001918), IncFII, IncHI1A, IncHI1B (R27) |
| Ec_400 h | ST156 | 5,241,723 | 50.5 | 42,932 | A | O76:H45 | cma, astA, hlyF, hra, iucC, iutA, ompT, papC, sitA, terC, traT | IncFIB (AP001918), IncFIC (FII) |
| Ec_361 p | ST10 | 5,207,190 | 50.7 | 75,974 | A | O29:H10 | astA, sitA, terC | IncHI2, IncHI2A, IncQ1, IncY |
| Ec_405 h | ST1286 | 5,984,169 | 50.34 | 26,744 | B1 | O71:H32 | aap, astA, iha, mchB, mchC, mchF, fyuA, irp2, terC, traT | IncB/O/K/Z, IncFII, IncFII (pHN7A8) |
| Ec_381 p | ST29 | 5,102,316 | 50.59 | 73,837 | B1 | O123/186:H11 | astA, cif, eae, efa1, espA, espB, espJ, iha, iss, katP, lpfA, nleA, nleB, nleC, sepA, tir, cea, iucC, iutA, ompT, terC, traT, fyuA, irp2 | IncFIB (AP001918), IncFII (pHN7A8), IncHI2, IncHI2A |
| Ec_410 h | ST2170 | 5,023,174 | 50.9 | 32,449 | B1 | H51 | cma, iss, lpfA, fyuA, hlyF, irp2, iucC, iutA, ompT, sitA, terC, traT | IncFIB (AP001918), IncFIB (pLF82), IncFIC (FII), IncI1 |
| Ec_388 p | ST5229 | 4,859,334 | 50.86 | 30,295 | ND | O177:H34 | astA, lpfA, fyuA, irp2, iucC, iutA, papC, sitA, terA, traT | IncHI2, IncFIB (AP001918), IncFII (pHN7A8), IncN |
| Ec_155 p | ST165 | 4,879,514 | 50.98 | 37,635 | ND | H26 | terC | IncQ1, IncR, IncX1, IncY |
| Ec_170 h | ST1147 | 5,104,121 | 50.7 | 91,960 | B1 | H35 | aap, aatA, iha, iss, lpfA, fyuA, irp2, terC, traT | IncFIB (AP001918), IncFII (29) |
| Ec_278 p | ST744 | 4,925,781 | 50.7 | 103,452 | A | O162/89:H9 | cma, cvaC, hlyF, iucC, iutA, ompT, sitA, terC, traT | IncFIB (AP001918), IncFIC (FII), IncN, IncQ1, IncX4 |
| Ec_172 h | ST69 | 5,391,031 | 50.77 | 72,381 | D | O15:H18 | air, eilA, iha, iss, lpfA, sat, senB, chuA, fyuA, irp2, iucC, iutA, kpsE, kpsMII_K52, ompT, papA_fsiA_F16, papC, sitA, terC, traT | Col156, IncB/O/K/Z, IncFIB (AP001918) |
| Ec_162 p | ST2170 | 4,901,850 | 50.7 | 65,441 | B1 | O78:H51 | cma, iss, lpfA, fyuA, hlyF, irp2, iucC, iutA, ompT, sitA, terC, traT | IncFIB (AP001918), IncFIC (FII), IncI2 |
| Ec_166 p | ST2954 | 4,803,218 | 50.87 | 19,954 | ND | O160:H9 | air, chuA, terC | IncFIB (pLF82), p0111 |
| Ec_232 h | ST167 | 4,956,803 | 50.84 | 70,019 | A | O-89 (162):H10 | fyuA, irp2, terC, traT, sitA | Col (BS512), IncFIA, IncFIB (AP001918), IncFII |
| Ec_168 p | ST2705 | 4,726,759 | 50.8 | 35,199 | A | H10 | ompT, TerC | p0111 |
| Ec_179 h | ST5044 | 4,523,830 | 50.79 | 93,960 | B1 | H29 | terC | ND |
| Ec_283 p | ST1011 | 5,130,567 | 50.55 | 61,007 | D | O8:H16 | air, cma, eilA, iroB, iss, chuA, cvaC, hlyF, iucC, iutA, ompT, sitA, terC, traT | IncFIB (AP001918), IncFIB (pLF82), IncFIC (FII), IncX4 |
| Ec_53 p | ST6775 | 4,825,481 | 50.8 | 26,621 | B1 | O65:H49 | iss, terC | IncHI2, IncHI2A, IncX1 |
| Ec_236 h | ST1294 | 4,511,999 | 50.78 | 126,228 | A | O9:H30 | terC | ND |
| Ec_57 p | ST165 | 5,115,907 | 50.56 | 52,960 | A | H27 | terC | IncHI2, IncHI2A, IncQ1, IncR, IncX1 |
| Ec_292 p | ST1121 | 4,893,986 | 50.7 | 83,326 | A | H48 | AstA, lpfA, terC | IncN, IncY |
| Ec_416 p | ST10 | 4,680,174 | 50.7 | 97,966 | A | O25:H32 | terC | IncFIB (K) |
| Ec_480 h | ST34 | 4,650,258 | 50.92 | 60,644 | A | H30 | terC | IncFIA (HI1), IncFIB (K) |
| Ec_426 p | ST2935 | 4,875,048 | 50.86 | 54,178 | A | H32 | astA, terC | IncP1, IncQ1, IncX1, IncY |
| Ec_488 h | ST484 | 4,877,138 | 50.57 | 68,561 | A | H4 | aap, astA, iss, kpsE, kpsMII, ompT, terC, traT | IncFIC (FII), IncFII (pCoo) |
| Ec_431 p | ST226 | 4,694,058 | 51 | 45,154 | A | O15:H10 | fyuA, irp2, terC | ND |
| Ec_489 h | ST34 | 5,015,072 | 50.6 | 59,062 | A | O68 (62):H30 | astA, TerC | IncFIA (HI1), IncFIB (K), IncHI1A, IncHI1B (R27), IncR, IncX1 |
| Ec_434 p | ST641 | 4,959,009 | 50.7 | 29,769 | B1 | H21 | astA, lpfA, terC | IncHI2, IncHI2A |
| Ec_495 h | ST1722 | 5,038,475 | 50.7 | 83,575 | F | H24 | air, eclb, eilA, iss, lpfA, chuA, fyuA, irp2, kpsE, sitA, terC, traT | Col (IMGS31), Col156, IncFIA, IncFIB (AP001918), IncFII |
| Ec_297 p | ST410 | 5,069,502 | 50.6 | 48,156 | C | O33:H26 | iss, lpfA, ompT, TerC, traT | IncFIA (HI1), IncFIB (AP001918), IncFII (pHN7A8), IncHI1A, IncHI1B (R27), IncI1 |
| Ec_60 p | ST515 | 5,084,460 | 50.9 | 65,609 | B1 | O128:H12 | fyuA, irp2, terC, traT | IncFIB (AP001918) |
| Ec_84 h | ST48 | 4,947,358 | 50.76 | 72,323 | A | O20 (137):H45 | cma, iroN, iss, katP, mchF, cvaC, hlyF, iucC, iutA, ompT, sitA, terC, traT | IncFIB (AP001918), IncP1 |
| Ec_63 p | ST515 | 4,812,948 | 50.9 | 55,529 | B1 | O128:H12 | fyuA, irp2, terC | Col440II |
| Ec_89 h | ST2040 | 4,932,400 | 50.7 | 69,363 | A | O159:H20 | astA, lpsA, cib, sitA, terC | IncB/O/K/Z, IncFII (pCoo) |
| Ec_67 p | ST6214 | 5,112,157 | 50.9 | 56,813 | B1 | O162/89:H10 | iss, nfaE, afaA, afaB, afaC, afaD, afaE, fyuA, irp2, iucC, iutA, sitA, TerC, TraT | IncFIB (AP001918), IncFII (pRSB107) |
| Ec_93 h | ST162 | 5,050,320 | 50.6 | 72,932 | B1 | O8:H21 | astA, iss, lpfA, hlyF, hra, iucC, iutA, ompT, papA-NEW, papC, sitA, terC, traT | IncFIB (AP001918), IncFIC (FII) |
| Ec_255 h | ST1722 | 4,823,623 | 50.62 | 78,611 | F | O1:H25 | air, astA, eilA, iss, lpfA, chuA, hra, kpsE, terC, yfcV | IncQ1 |
| Ec_98 h | ST7153 | 4,746,746 | 50.75 | 49,123 | A | O148:H30 | astA, terC, traT | IncFIA (HI1), IncFIB (K), IncX1, IncY |
| Ec_80 p | ST515 | 4,962,369 | 50.9 | 38,744 | B1 | O128:H12 | fyuA, irp2, terC, traT | IncFIB (AP001918), p0111 |
| Ec_271 h | ST1196 | 4,931,876 | 50.84 | 129,873 | B1 | O29:H8 | cma, lpfA, cea, cvaC, hlyF, iucC, iutA, ompT, sitA, terC, traT | IncFIB (AP001918), IncFIC (FII) |
| Ec_81 p | ST877 | 4,744,811 | 50.8 | 59,541 | B1 | O28ac/O42:H32 | lpfA, ompT, TerC | IncFIB (AP001918), IncFIA (HI1), IncFIB (K), Inc1 |
| Ec_447 p | ST4956 | 4,805,635 | 50.9 | 54,620 | B1 | O156:H37 | lpfA, terC, traT | IncHI2, IncFIB (AP001918), IncFIC (FII) |
| Ec_498 h | ST10 | 4,743,370 | 50.91 | 77,783 | A | O9 (89,162):H9 | sitA, terC, traT | IncFIA, IncFIB (AP001918), IncFII |
| Ec_83 p | ST6018 | 4,752,755 | 50.79 | 69,914 | A | O8:H11 | ompT, terC | IncX1, p0111 |
| Ec_442 p | ST101 | 5,046,847 | 50.26 | 71,098 | B1 | O118:H21 | cma, iss, lpfA, cvaC, hlyF, iucC, iutA, ompT, sitA, terC, traT | IncFIB (AP001918), IncFIC (FII), IncFII (29), IncI2 |
| Ec_452 p | ST515 | 4,878,968 | 51 | 42,049 | B1 | O128ac:H12 | fyuA, irp2, terC, traT | IncFIB (AP001918), IncX4 |
| Ec_500 h | ST2040 | 4,895,059 | 50.77 | 26,675 | A | O159:H20 | astA, lpfA, cib, sitA, terC | IncB/O/K/Z, IncFII (pCoo) |
| Ec_457 p | ST1121 | 4,935,661 | 50.57 | 131,601 | A | H48 | lpfA, terC | IncX4, IncY |
| Ec_503 h | ST192 | 4,868,610 | 50.73 | 79,109 | B1 | O124 (164):H34 | iss, lpfA, ompT, terC | IncX2, p0111 |
| Ec_511 h | ST38 | 5,198,810 | 50.66 | 73,146 | D | O1:H15 | air, iha, iss, sat, senB, chuA, fyuA, hra, irp2, iucC, iutA, kpsEkpsMII_K5, papA_F43, terC, traT, kpsE | Col156, IncFIA, IncFIB (AP001918), IncFII (pRSB107) |
| Ec_467 p | ST4238 | 4,663,515 | 50.71 | 92,493 | A | H2 | capU, terC | IncFIB (K) |
| Ec_514 h | ST38 | 5,084,740 | 50.7 | 54,474 | D | O86:H18 | air, astA, eilA, iss, afaD, chuA, fyuA, irp2, kpsE, kpsMII_K5, terC | IncL/M (pOXA-48) |
| Ec_472 p | ST877 | 4,948,850 | 50.7 | 72,864 | B1 | H10 | lpfA, ompT, TerC, traT | IncFII (pCoo), IncI1, IncX1 |
| Ec_517 h | ST1141 | 5,068,761 | 50.82 | 55,942 | B1 | O13:H11 | astA, terC, traT | Col (BS512), Col (IRGK), IncFIB (K), IncR, IncY |
| Ec_519 h | ST6253 | 5,382,277 | 50.8 | 35,706 | D | H15 | aap, air, astA, capU, eatA, eilA, chuA, kpsE, kpsMII_K5, terC, traT | IncFII |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Truong, D.T.Q.; Hounmanou, Y.M.G.; Dang, S.T.T.; Olsen, J.E.; Truong, G.T.H.; Tran, N.T.; Scheutz, F.; Dalsgaard, A. Genetic Comparison of ESBL-Producing Escherichia coli from Workers and Pigs at Vietnamese Pig Farms. Antibiotics 2021, 10, 1165. https://doi.org/10.3390/antibiotics10101165
Truong DTQ, Hounmanou YMG, Dang STT, Olsen JE, Truong GTH, Tran NT, Scheutz F, Dalsgaard A. Genetic Comparison of ESBL-Producing Escherichia coli from Workers and Pigs at Vietnamese Pig Farms. Antibiotics. 2021; 10(10):1165. https://doi.org/10.3390/antibiotics10101165
Chicago/Turabian StyleTruong, Duong Thi Quy, Yaovi Mahuton Gildas Hounmanou, Son Thi Thanh Dang, John Elmerdahl Olsen, Giang Thi Huong Truong, Nhat Thi Tran, Flemming Scheutz, and Anders Dalsgaard. 2021. "Genetic Comparison of ESBL-Producing Escherichia coli from Workers and Pigs at Vietnamese Pig Farms" Antibiotics 10, no. 10: 1165. https://doi.org/10.3390/antibiotics10101165