
Depth-Dependent C-N-P Stocks and Stoichiometry in Ultisols Resulting from Conversion of Secondary Forests to Plantations and Driving Forces



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Soil Sampling
2.3. Chemical Analysis
2.4. Calculation of Stocks
2.5. Statistical Analysis
3. Results
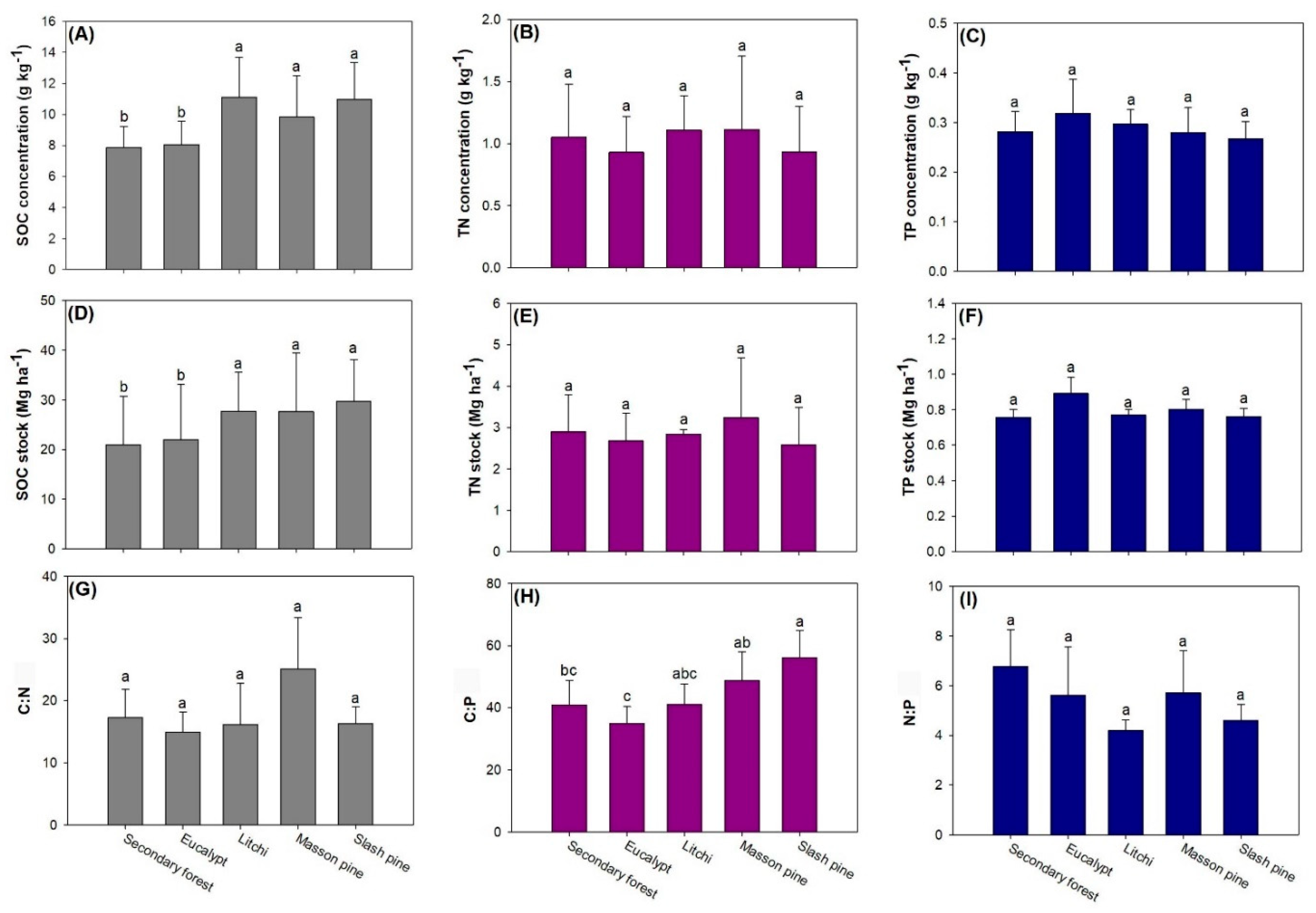
3.1. Difference among Forest Types
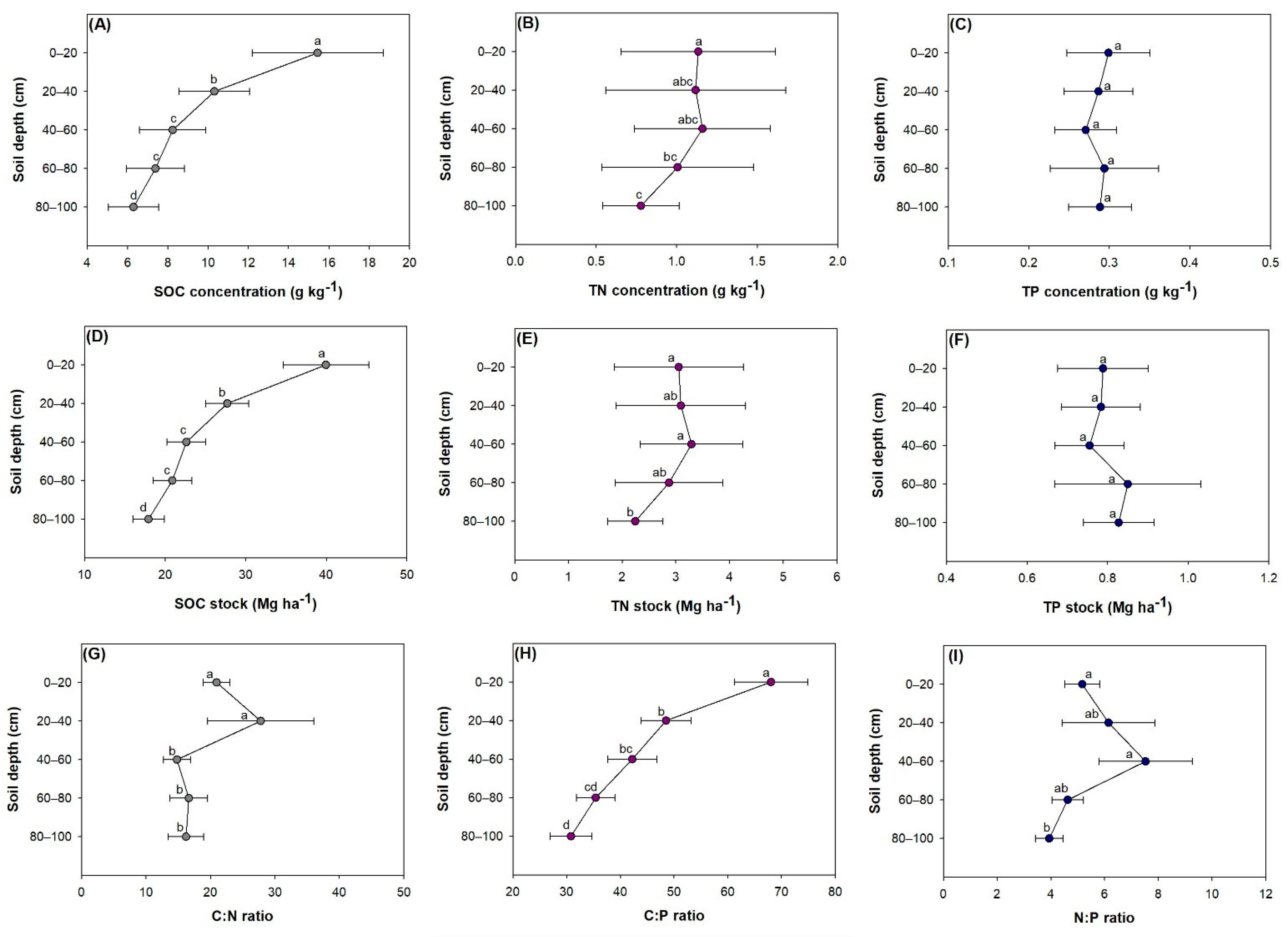
3.2. Variation among Soil Layer Depths
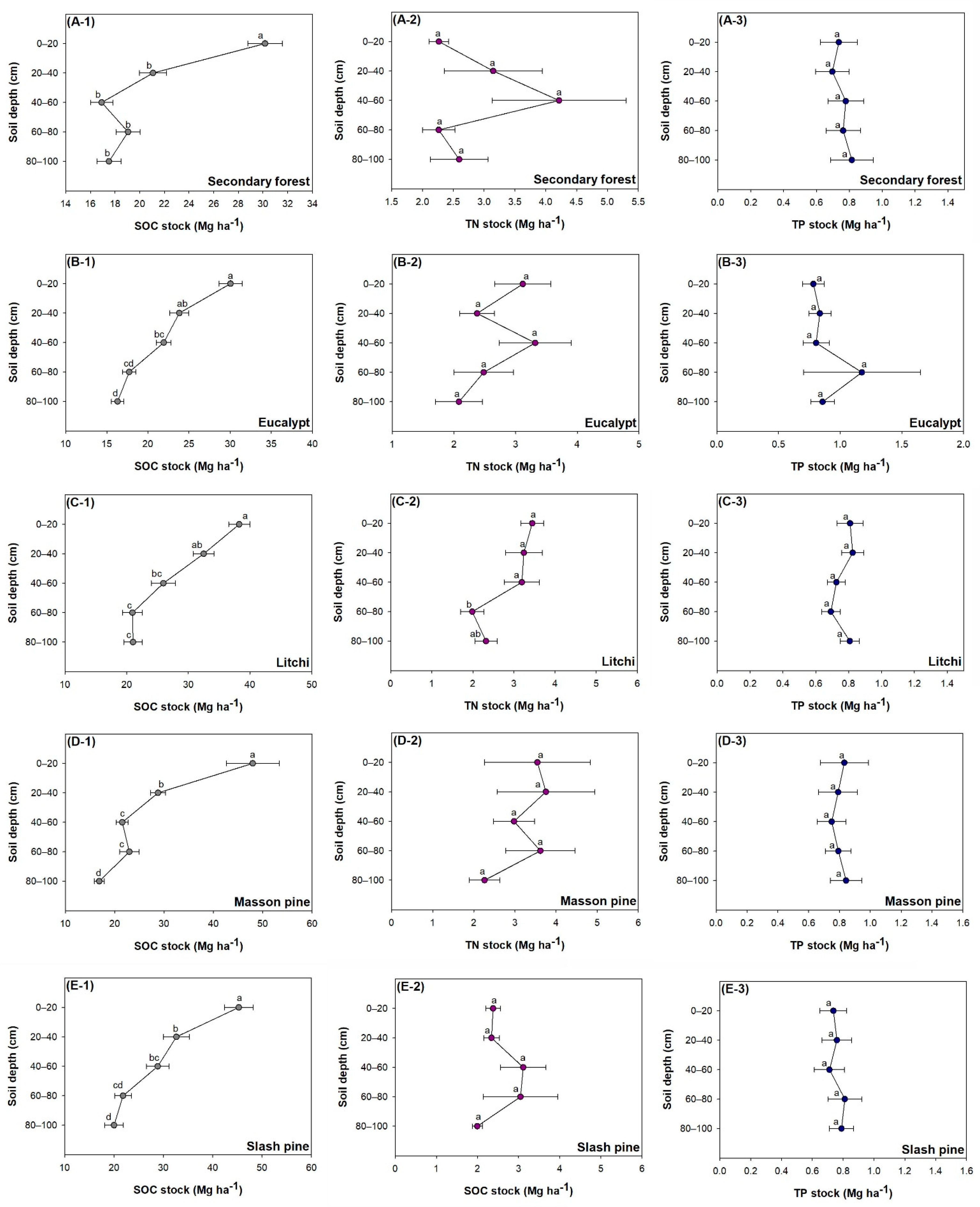
3.3. Variation of Concentration along Soil Depths for Different Species
3.4. Variation of Stock along Soil Depths for Different Species
3.5. Regression of Soil Variable Stock
4. Discussion
4.1. Failure of Interactive Effects
4.2. Variation among Forest Types
4.3. Vertical Distribution across Soil Layers
4.4. Vertical Distribution among Differnet Forest Types
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lal, R. Managing Soils and Ecosystems for Mitigating Anthropogenic Carbon Emissions and Advancing Global Food Security. BioScience 2010, 60, 708–721. [Google Scholar] [CrossRef]
- Pires, C.V.; Schaefer, C.E.R.G.; Hashigushi, A.K.; Thomazini, A.; Filho, E.I.F.; Mendonça, E.S. Soil organic carbon and nitrogen pools drive soil C-CO2 emissions from selected soils in Maritime Antarctica. Sci. Total Environ. 2017, 596–597, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Zhang, P.; Chen, Y.M. Soil C:N:P stoichiometry in plantations of N-fixing black locust and indigenous pine, and secondary oak forests in Northwest China. J. Soils Sediments 2018, 18, 1478–1489. [Google Scholar] [CrossRef]
- Jobbagy, E.G.; Jackson, R.B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Dixon, R.K.; Brown, S.; Houghton, R.A.; Solomon, A.M.; Trexler, M.C.; Wisniewski, J. Carbon pools and flux of global forest ecosystems. Science 1994, 263, 185–190. [Google Scholar] [CrossRef]
- Lewis, S.L.; Lopez-Gonzalez, G.; Sonke, B.; Affum-Baffoe, K.; Baker, T.R.; Ojo, L.O.; Phillips, O.L.; Reitsma, J.M.; White, L.; Comiskey, J.A.; et al. Increasing carbon storage in intact African tropical forests. Nature 2009, 457, 1003. [Google Scholar] [CrossRef]
- Malhi, Y. The carbon balance of tropical forest regions, 1990–2005. Curr. Opin. Environ. Sustain. 2010, 2, 237–244. [Google Scholar] [CrossRef]
- Beer, C.; Reichstein, M.; Tomelleri, E.; Ciais, P.; Jung, M.; Carvalhais, N.; Rodenbeck, C.; Arain, M.A.; Baldocchi, D.; Bonan, G.B.; et al. Terrestrial Gross Carbon Dioxide Uptake: Global Distribution and Covariation with Climate. Science 2010, 329, 834–838. [Google Scholar] [CrossRef] [Green Version]
- Raich, J.W.; Schlesinger, W.H. The global carbon-dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus Ser. B Chem. Phys. Meteorol. 1992, 44, 81–99. [Google Scholar] [CrossRef] [Green Version]
- Cleveland, C.C.; Liptzin, D. C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 2007, 85, 235–252. [Google Scholar] [CrossRef]
- Hu, L.; Ade, L.; Wu, X.; Zi, H.; Luo, X.; Wang, C. Changes in Soil C:N:P Stoichiometry and Microbial Structure along Soil Depth in Two Forest Soils. Forests 2019, 10, 113. [Google Scholar] [CrossRef] [Green Version]
- Reich, P.B.; Hobbie, S.E.; Lee, T.; Ellsworth, D.S.; West, J.B.; Tilman, D.; Knops, J.M.H.; Naeem, S.; Trost, J. Nitrogen limitation constrains sustainability of ecosystem response to CO2. Nature 2006, 440, 922–925. [Google Scholar] [CrossRef] [PubMed]
- Batjes, N.H. Total carbon and nitrogen in the soils of the world. Eur. J. Soil Sci. 1996, 47, 151–163. [Google Scholar] [CrossRef]
- Stevenson, F.J.; Cole, M.A. Cycles of Soils: Carbon, Nitrogen, Phosphorus, Sulfur, Micronutrients; John Wiley and Sons: Hoboken, NJ, USA, 1999. [Google Scholar]
- Walker, T.W.; Syers, J.K. The fate of phosphorus during pedogenesis. Geoderma 1976, 15, 1–19. [Google Scholar] [CrossRef]
- Trudgill, S.T. Soil and Vegetataion Systems; Clarendon Press: Oxford, UK, 1988. [Google Scholar]
- Groppo, J.D.; Lins, S.R.M.; Camargo, P.B.; Assad, E.D.; Pinto, H.S.; Martins, S.C.; Salgado, P.R.; Evangelista, B.; Vasconcellos, E.; Sano, E.E.; et al. Changes in soil carbon, nitrogen, and phosphorus due to land-use changes in Brazil. Biogeosciences 2015, 12, 4765–4780. [Google Scholar] [CrossRef] [Green Version]
- Fan, H.B.; Wu, J.P.; Liu, W.F.; Yuan, Y.H.; Hu, L.; Cai, Q.K. Linkages of plant and soil C:N:P stoichiometry and their relationships to forest growth in subtropical plantations. Plant Soil 2015, 392, 127–138. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Reich, P.B.; Khachane, A.N.; Campbell, C.D.; Thomas, N.; Freitag, T.E.; Abu Al-Soud, W.; Sorensen, S.; Bardgett, R.D.; Singh, B.K. It is elemental: Soil nutrient stoichiometry drives bacterial diversity. Environ. Microbiol. 2017, 19, 1176–1188. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, S.; Yu, X. Decline of soil fertility during forest conversion of secondary forest to Chinese fir plantations in subtropical China. Land Degrad. Dev. 2011, 22, 444–452. [Google Scholar] [CrossRef]
- Lewis, T.; Smith, T.E.; Hogg, B.; Swift, S.; Verstraten, L.; Bryant, P.; Wehr, B.J.; Tindale, N.; Menzies, N.W.; Dalal, R.C. Conversion of sub-tropical native vegetation to introduced conifer forest: Impacts on below-ground and above-ground carbon pools. For. Ecol. Manag. 2016, 370, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, L.; Xiang, W.H.; Ouyang, S.; Zhang, T.D.; Zhang, X.L.; Zeng, Y.L.; Hu, Y.T.; Luo, G.W.; Kuzyakov, Y. Forest conversion to plantations: A meta-analysis of consequences for soil and microbial properties and functions. Glob. Chang. Biol. 2021. Early Access. [Google Scholar] [CrossRef]
- Ngo, K.M.; Turner, B.L.; Muller-Landau, H.C.; Davies, S.J.; Larjavaara, M.; Hassan, N.F.B.; Lum, S. Carbon stocks in primary and secondary tropical forests in Singapore. For. Ecol. Manag. 2013, 296, 81–89. [Google Scholar] [CrossRef]
- Lal, R. Soil carbon sequestration to mitigate climate change. Geoderma 2004, 123, 1–22. [Google Scholar] [CrossRef]
- University of Idaho. The Twelve Soil Orders: Ultisols. Available online: https://www.uidaho.edu/cals/soil-orders/ultisols#:~:text=Ultisols%20(from%20Latin%20ultimus%2C%20%22,typically%20on%20older%2C%20stable%20landscapes (accessed on 3 January 2021).
- Wikipedia. The Red Clay Soils: Ultisol. Available online: https://en.wikipedia.org/wiki/Ultisol (accessed on 4 January 2021).
- Fujii, K.; Uemura, M.; Hayakawa, C.; Funakawa, S.; Sukartiningsih; Kosaki, T.; Ohta, S. Fluxes of dissolved organic carbon in two tropical forest ecosystems of East Kalimantan, Indonesia. Geoderma 2009, 152, 127–136. [Google Scholar] [CrossRef]
- Lu, D.; Moran, E.; Mausel, P. Linking amazonian secondary succession forest growth to soil properties. Land Degrad. Dev. 2002, 13, 331–343. [Google Scholar] [CrossRef]
- Sotomayor-Ramirez, D.; Espinoza, Y. Soil aggregation and organic carbon fractions in the Rio Grande de Arecibo watershed. J. Agric. Univ. P. R. 2010, 94, 25–39. [Google Scholar] [CrossRef]
- Qiao, Y.; Wang, J.; Liu, H.M.; Huang, K.; Yang, Q.S.; Lu, R.L.; Yan, L.M.; Wang, X.H.; Xia, J.Y. Depth-dependent soil C-N-P stoichiometry in a mature subtropical broadleaf forest. Geoderma 2020, 370, 9. [Google Scholar] [CrossRef]
- Ding, X.G.; Zhao, Z.Y.; Yang, Q.; Chen, L.N.; Tian, Q.Y.; Li, X.C.; Meng, F.R. Model prediction of depth-specific soil texture distributions with artificial neural network: A case study in Yunfu, a typical area of Udults Zone, South China. Comput. Electron. Agric. 2020, 169, 13. [Google Scholar] [CrossRef]
- Luo, X.Z.; Hou, E.Q.; Chen, J.Q.; Li, J.; Zhang, L.L.; Zang, X.W.; Wen, D.Z. Dynamics of carbon, nitrogen, and phosphorus stocks and stoichiometry resulting from conversion of primary broadleaf forest to plantation and secondary forest in subtropical China. Catena 2020, 193, 104606. [Google Scholar] [CrossRef]
- Zhao, F.Z.; Zhang, L.; Ren, C.J.; Sun, J.; Han, X.H.; Yang, G.H.; Wang, J. Effect of Microbial Carbon, Nitrogen, and Phosphorus Stoichiometry on Soil Carbon Fractions under a Black Locust Forest within the Central Loess Plateau of China. Soil Sci. Soc. Am. J. 2016, 80, 1520–1530. [Google Scholar] [CrossRef]
- Li, X.; Ding, X.; Ceng, S.; Zhang, C.; Yang, H. Forestry Soil Survey of Yunfu, Guangdong Province; China Forestry Publishing House: Beijing, China, 2018. [Google Scholar]
- Tang, Z.H.; Ouyang, T.P.; Li, M.K.; Huang, N.S.; Kuang, Y.Q.; Hu, Q.; Zhu, Z.Y. Potential effects of exploiting the Yunfu Pyrite mine (southern China) on soil: Evidence from analyzing trace elements in surface soil. Environ. Monit. Assess. 2019, 191, 1–18. [Google Scholar] [CrossRef]
- Wang, W.; Shu, J.; Chen, W.; Ding, J. Holocene environmental changes and human impact in the Yangtze River Delta area in East China. Quat. Sci. 2010, 30, 233–244. [Google Scholar]
- Wei, R.-P.; Yang, R.; Wei, Q. Effect of Lime Application to Acidic Soils on Oleoresin Yield Tapped from Pine Plantations in South China. Open J. For. 2014, 4, 390–397. [Google Scholar] [CrossRef] [Green Version]
- Anjum, J.; Lone, R.; Wani, K.A. Lychee (Litchi chinensis): Biochemistry, Panacea, and Nutritional Value. In Lychee Disease Management; Kumar, M., Kumar, V., Bhalla-Sarin, N., Varma, A., Eds.; Springer Singapore: Singapore, 2017; pp. 237–256. [Google Scholar]
- Liu, G. Soil Physical and Chemical Analysis, Description of Soil Profiles; Standard Press of China: Beijing, China, 1996. [Google Scholar]
- Bremner, J.M.; Mulvaney, C.S. Nitrogen—Total. In Methods of Soil Analysis; American Society of Agronomy: Madison, WI, USA, 1983; pp. 595–624. [Google Scholar]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Xu, H.W.; Qu, Q.; Li, P.; Guo, Z.Q.; Wulan, E.; Xue, S. Stocks and Stoichiometry of Soil Organic Carbon, Total Nitrogen, and Total Phosphorus after Vegetation Restoration in the Loess Hilly Region, China. Forests 2019, 10, 27. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.Y.; Yang, Q.; Sun, D.X.; Ding, X.G.; Meng, F.R. Extended model prediction of high-resolution soil organic matter over a large area using limited number of field samples. Comput. Electron. Agric. 2020, 169, 12. [Google Scholar] [CrossRef]
- de Blecourt, M.; Brumme, R.; Xu, J.C.; Corre, M.D.; Veldkamp, E. Soil Carbon Stocks Decrease following Conversion of Secondary Forests to Rubber (Hevea brasiliensis) Plantations. PLoS ONE 2013, 8, e69357. [Google Scholar]
- Yang, Y.S.; Xie, J.S.; Sheng, H.; Chen, G.S.; Li, X.; Yang, Z.J. The impact of land use/cover change on storage and quality of soil organic carbon in midsubtropical mountainous area of southern China. J. Geogr. Sci. 2009, 19, 49–57. [Google Scholar] [CrossRef]
- Li, Y.; Xu, M.; Zou, X.M.; Shi, P.J.; Zhang, Y.Q. Comparing soil organic carbon dynamics in plantation and secondary forest in wet tropics in Puerto Rico. Glob. Chang. Biol. 2005, 11, 239–248. [Google Scholar] [CrossRef]
- Chevasco, E.D.; Minogue, P.J.; Mackowiak, C.; Comerford, N.B. Fertilization and pine straw raking in slash pine plantations: P removals and effects on total and mobile soil, foliage and litter P pools. For. Ecol. Manag. 2016, 376, 310–320. [Google Scholar] [CrossRef]
- Bui, E.N.; Henderson, B.L. C:N:P stoichiometry in Australian soils with respect to vegetation and environmental factors. Plant Soil 2013, 373, 553–568. [Google Scholar] [CrossRef]
- Schapel, A.; Marschner, P.; Churchman, J. Clay amount and distribution influence organic carbon content in sand with subsoil clay addition. Soil Tillage Res. 2018, 184, 253–260. [Google Scholar] [CrossRef]
- Jin, S.F.; Wang, H.S. Relationships between soil pH and soil carbon in China’s carbonate soils. Fresenius Environ. Bull. 2018, 27, 605–611. [Google Scholar]
- Brevik, E.; Fenton, T.; Moran, L. Effect of soil compaction on organic carbon amounts and distribution, South-Central Iowa. Environ. Pollut. 2002, 116, S137–S141. [Google Scholar] [CrossRef]
- Cheng, Y.; Li, P.; Xu, G.; Li, Z.; Cheng, S.; Gao, H. Spatial distribution of soil total phosphorus in Yingwugou watershed of the Dan River, China. CATENA 2016, 136, 175–181. [Google Scholar] [CrossRef]
- Kooijman, A.M.; Jongejans, J.; Sevink, J. Parent material effects on Mediterranean woodland ecosystems in NE Spain. CATENA 2005, 59, 55–68. [Google Scholar] [CrossRef]
- Tian, H.; Chen, G.; Zhang, C.; Melillo, J.M.; Hall, C.A.S. Pattern and variation of C:N:P ratios in China’s soils: A synthesis of observational data. Biogeochemistry 2010, 98, 139–151. [Google Scholar] [CrossRef]
- Zhu, J.; Wu, A.C.; Zhou, G.Y. Spatial distribution patterns of soil total phosphorus influenced by climatic factors in China’s forest ecosystems. Sci. Rep. 2021, 11, 5357. [Google Scholar] [CrossRef]
- Garcia-Velazquez, L.; Rodriguez, A.; Gallardo, A.; Maestre, F.T.; Dos Santos, E.; Lafuente, A.; Fernandez-Alonso, M.J.; Singh, B.K.; Wang, J.T.; Duran, J. Climate and soil micro-organisms drive soil phosphorus fractions in coastal dune systems. Funct. Ecol. 2020, 34, 1690–1701. [Google Scholar] [CrossRef]
- Wang, Y.P.; Law, R.M.; Pak, B. A global model of carbon, nitrogen and phosphorus cycles for the terrestrial biosphere. Biogeosciences 2010, 7, 2261–2282. [Google Scholar] [CrossRef] [Green Version]
- Li, X.P.; Chang, S.X.; Liu, J.T.; Zheng, Z.M.; Wang, X.H. Topography-soil relationships in a hilly evergreen broadleaf forest in subtropical China. J. Soils Sediments 2017, 17, 1101–1115. [Google Scholar] [CrossRef]
- Zhang, S.; Yan, L.; Huang, J.; Mu, L.; Huang, Y.; Zhang, X.; Sun, Y. Spatial Heterogeneity of Soil C:N Ratio in a Mollisol Watershed of Northeast China. Land Degrad. Dev. 2016, 27, 295–304. [Google Scholar] [CrossRef]
- Yang, S.B.; Feng, C.; Ma, Y.H.; Wang, W.J.; Huang, C.; Qi, C.J.; Fu, S.L.; Chen, H.Y.H. Transition from N to P limited soil nutrients over time since restoration in degraded subtropical broadleaved mixed forests. For. Ecol. Manag. 2021, 494, 119298. [Google Scholar] [CrossRef]
- Sun, J.; Li, S.; Wei, Z.; Wang, D.; Qi, Z. Distribution of soil nutrients in the litchi orchard in the midwest Hainan. Guangdong Agric. Sci. 2013, 40, 62–64. [Google Scholar]
- Yao, L.; Zhou, C.; He, Z.; Li, G.; Yang, B.; Bai, C. Nutrient accumulation characteristics of main litchi cultivars and their relationships with soil nutrients. J. South China Agric. Univ. 2020, 41, 40–47. [Google Scholar]
- Menzel, C.M.; Haydon, G.F.; Simpson, D.R. Growth of Lychee (Litchi-chinensis sonn) in sand culture under variable nitrogen supply. J. Horticult. Sci. 1995, 70, 757–767. [Google Scholar]
- Zhang, J.E.; Luo, S.M.; Duan, S.S. The eco-environment of a young litchi orchard interplanted with forage grass in south China. In Proceedings of the First International Symposium on Litchi and Longan, Guangzhou, China, 16–19 June 2000; Huang, H., Menzel, C., Eds.; International Society for Horticultural Science: Leuven, Belgium, 2001; pp. 311–313. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | SOC Conc. | TN Conc. | TP Conc. | |||
| F value | P value | F value | P value | F value | P value | |
| F | 5.16 | 0.0004 | 1.19 | 0.3122 | 1.47 | 0.2093 |
| S | 41.75 | <0.0001 | 3.84 | 0.0044 | 0.56 | 0.6911 |
| F × S | 0.65 | 0.8456 | 0.49 | 0.9511 | 0.15 | 1.0000 |
| SOC Stock | TN Stock | TP Stock | ||||
| F value | P value | F value | P value | F value | P value | |
| F | 6.12 | <0.0001 | 0.60 | 0.6635 | 0.99 | 0.4128 |
| S | 35.35 | <0.0001 | 2.51 | 0.0409 | 0.41 | 0.8049 |
| F × S | 0.67 | 0.8221 | 0.53 | 0.9343 | 0.14 | 1.0000 |
| C:N | C:P | N:P | ||||
| F value | P value | F value | P value | F value | P value | |
| F | 1.57 | 0.1806 | 3.51 | 0.0077 | 0.39 | 0.8171 |
| S | 5.82 | 0.0001 | 18.27 | <0.0001 | 2.72 | 0.0290 |
| F × S | 0.49 | 0.9540 | 0.35 | 0.9915 | 0.31 | 0.9955 |
| Variable | Parameter Estimate | SE | Model R2 | F Value | p Value |
|---|---|---|---|---|---|
| 0–20 cm | |||||
| Intercept | −624.42 | 292.62 | - | 4.55 | 0.0353 |
| Longitude | 5.90 | 2.59 | 0.1396 | 5.19 | 0.0248 |
| Elevation | 0.02 | 0.01 | 0.0954 | 4.40 | 0.0384 |
| pH | −5.67 | 2.51 | 0.0564 | 6.15 | 0.0147 |
| 20–40 cm | |||||
| Intercept | 16.23 | 3.92 | - | 17.09 | <0.0001 |
| Elevation | 0.02 | 0.01 | 0.1583 | 19.37 | <0.0001 |
| BD | −6.33 | 2.65 | 0.2029 | 5.71 | 0.0187 |
| 40–60 cm | |||||
| Intercept | 153.60 | 68.07 | - | 5.09 | 0.0262 |
| Latitude | −6.03 | 3.00 | 0.2162 | 4.04 | 0.0472 |
| Slope | 0.14 | 0.05 | 0.1848 | 6.43 | 0.0127 |
| BD | −8.28 | 2.46 | 0.1334 | 15.86 | 0.0001 |
| 60–80 cm | |||||
| Intercept | −321.37 | 116.60 | - | 7.60 | 0.0069 |
| Longitude | 2.91 | 1.04 | 0.2239 | 7.82 | 0.0062 |
| Elevation | 0.02 | 0.01 | 0.1645 | 20.27 | <0.0001 |
| 80–100 cm | |||||
| Intercept | 198.09 | 51.37 | - | 14.87 | 0.0002 |
| Latitude | −8.25 | 2.30 | 0.1245 | 14.65 | 0.0002 |
| Slope | 0.09 | 0.04 | 0.1892 | 8.13 | 0.0053 |
| BD | −4.81 | 2.04 | 0.2317 | 5.59 | 0.0200 |
| Variable | Parameter Estimate | SE | Model R2 | F Value | p Value |
|---|---|---|---|---|---|
| Intercept | 22.61 | 11.02 | - | 4.21 | 0.0429 |
| Latitude | −0.97 | 0.49 | 0.1172 | 4.04 | 0.0472 |
| Elevation | 0.01 | 0.01 | 0.0823 | 9.24 | 0.0030 |
| Variable | Parameter Estimate | SE | Model R2 | F Value | p Value |
|---|---|---|---|---|---|
| 0–20 cm | |||||
| Intercept | −17.96 | 5.55 | - | 10.48 | 0.0016 |
| Longitude | 0.16 | 0.05 | 0.0952 | 10.83 | 0.0014 |
| 20–40 cm | |||||
| Intercept | −14.99 | 4.57 | - | 10.76 | 0.0014 |
| Longitude | 0.14 | 0.04 | 0.0979 | 11.18 | 0.0012 |
| 40–60 cm | |||||
| Intercept | −11.39 | 4.16 | - | 7.48 | 0.0074 |
| Longitude | 0.11 | 0.04 | 0.0640 | 7.04 | 0.0092 |
| BD | −0.16 | 0.08 | 0.0991 | 3.98 | 0.0488 |
| 60–80 cm | |||||
| Intercept | 0.29 | 0.03 | - | 125.68 | <0.0001 |
| 80–100 cm | |||||
| Intercept | −11.62 | 4.19 | - | 7.69 | 0.0066 |
| Longitude | 0.11 | 0.04 | 0.0758 | 8.44 | 0.0045 |
| BD | −0.20 | 0.09 | 0.1214 | 5.29 | 0.0235 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, X.; Li, X.; Qi, Y.; Zhao, Z.; Sun, D.; Wei, H. Depth-Dependent C-N-P Stocks and Stoichiometry in Ultisols Resulting from Conversion of Secondary Forests to Plantations and Driving Forces. Forests 2021, 12, 1300. https://doi.org/10.3390/f12101300
Ding X, Li X, Qi Y, Zhao Z, Sun D, Wei H. Depth-Dependent C-N-P Stocks and Stoichiometry in Ultisols Resulting from Conversion of Secondary Forests to Plantations and Driving Forces. Forests. 2021; 12(10):1300. https://doi.org/10.3390/f12101300
Chicago/Turabian StyleDing, Xiaogang, Xiaochuan Li, Ye Qi, Zhengyong Zhao, Dongxiao Sun, and Hongxu Wei. 2021. "Depth-Dependent C-N-P Stocks and Stoichiometry in Ultisols Resulting from Conversion of Secondary Forests to Plantations and Driving Forces" Forests 12, no. 10: 1300. https://doi.org/10.3390/f12101300