
The Longitudinal Effect of Meditation on Resting-State Functional Connectivity Using Dynamic Arterial Spin Labeling: A Feasibility Study

 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Sample Size Justification
2.2. Study Population
2.3. MRI Acquisition
2.4. Image Processing
2.4.1. Quantification of Absolute CBF Maps
2.4.2. Normalization of dASL Image Time Series and CBF Maps
2.4.3. Quantification of CBF Functional Connectivity Maps
2.5. Statistical Analysis
3. Results
3.1. Basic Characteristics of the Participants
3.2. Quality of dASL Time Series
3.3. Differences in Global CBF and Relative CBF between the Baseline and Follow-Up
3.4. Association of Relative CBF with Meditation Practice Time
3.5. Differences in CBF Functional Connectivity between Baseline and Follow-Up
3.6. Association of CBF Functional Connectivity with Meditation Practice Time
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Prakash, R.S.; Whitmoyer, P.; Aldao, A.; Schirda, B. Mindfulness and emotion regulation in older and young adults. Aging Ment. Health 2015, 21, 77–87. [Google Scholar] [CrossRef]
- Hölzel, B.K.; Lazar, S.W.; Gard, T.; Schuman-Olivier, Z.; Vago, D.R.; Ott, U. How does mindfulness meditation work? Proposing mechanisms of action from a conceptual and neural perspective. Perspect. Psychol. Sci. 2011, 6, 537–559. [Google Scholar] [CrossRef]
- Zeidan, F.; Johnson, S.K.; Diamond, B.J.; David, Z.; Goolkasian, P. Mindfulness meditation improves cognition: Evidence of brief mental training. Conscious. Cogn. 2010, 19, 597–605. [Google Scholar] [CrossRef]
- Moore, A.; Gruber, T.; Derose, J.; Malinowski, P. Regular, brief mindfulness meditation practice improves elec-trophysiological markers of attentional control. Front. Hum. Neurosci. 2012, 6, 18. [Google Scholar] [CrossRef] [Green Version]
- Banks, J.B.; Welhaf, M.S.; Srour, A. The protective effects of brief mindfulness meditation training. Conscious. Cogn. 2015, 33, 277–285. [Google Scholar] [CrossRef]
- Chien, W.T.; Thompson, D. Effects of a mindfulness-based psychoeducation programme for Chinese patients with schizophrenia: 2-year follow-up. Br. J. Psychiatry 2014, 205, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-C.; Barrós-Loscertales, A.; Pinazo, D.; Ventura-Campos, N.; Borchardt, V.; Bustamante, J.-C.; Rodríguez-Pujadas, A.; Fuentes-Claramonte, P.; Balaguer, R.; Avila, C.; et al. State and training effects of mindfulness meditation on brain networks reflect neuronal mechanisms of its antidepressant effect. Neural Plast. 2016, 2016, 9504642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, D.S.; Kabat-Zinn, J. Mindfulness in medicine. JAMA 2008, 300, 1350–1352. [Google Scholar] [CrossRef]
- Janssen, L.; Kan, C.C.; Carpentier, P.J.; Sizoo, B.; Hepark, S.; Grutters, J.; Donders, R.; Buitelaar, J.K.; Speckens, A.E. Mindfulness based cognitive therapy versus treatment as usual in adults with attention deficit hyperactivity disorder (ADHD). BMC Psychiatry 2015, 15, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hölzel, B.K.; Ott, U.; Gard, T.; Hempel, H.; Weygandt, M.; Morgen, K.; Vaitl, D. Investigation of mindfulness meditation practitioners with voxel-based morphometry. Soc. Cogn. Affect. Neurosci. 2007, 3, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Vestergaard-Poulsen, P.; van Beek, M.; Skewes, J.; Bjarkam, C.R.; Stubberup, M.; Bertelsen, J.; Roepstorff, A. Long-term meditation is associated with increased gray matter density in the brain stem. Neuroreport 2009, 20, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Hernández, S.E.; Suero, J.; Barros, A.; González-Mora, J.L.; Rubia, K. Increased Grey Matter Associated with Long-Term Sahaja Yoga Meditation: A Voxel-Based Morphometry Study. PLoS ONE 2016, 11, e0150757. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.A.; Duerden, E.G.; Courtemanche, J.; Cherkasova, M.; Duncan, G.H.; Rainville, P. Cortical thickness, mental absorption and meditative practice: Possible implications for disorders of attention. Biol. Psychol. 2013, 92, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.A.; Courtemanche, J.; Duerden, E.G.; Duncan, G.H.; Rainville, P. Cortical thickness and pain sensitivity in zen meditators. Emotion 2010, 10, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.C.; Barros-Loscertales, A.; Li, M.; Pinazo, D.; Borchardt, V.; Avila, C.; Walter, M. Alterations in brain structure and amplitude of low-frequency after 8 weeks of mindfulness meditation training in meditation naive subjects. Sci. Rep. 2019, 9, 10977. [Google Scholar] [CrossRef] [PubMed]
- Lazar, S.W.; Kerr, C.E.; Wasserman, R.H.; Gray, J.R.; Greve, D.N.; Treadway, M.T.; McGarvey, M.; Quinn, B.T.; Dusek, J.A.; Benson, H.; et al. Meditation experience is associated with increased cortical thickness. Neuroreport 2005, 16, 1893–1897. [Google Scholar] [CrossRef] [Green Version]
- Holzel, B.K.; Carmody, J.; Evans, K.C.; Hoge, E.A.; Dusek, J.A.; Morgan, L.; Pitman, R.K.; Lazar, S.W. Stress reduction correlates with structural changes in the amygdala. Soc. Cogn. Affect. Neurosci. 2010, 5, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Holzel, B.K.; Carmody, J.; Vangel, M.; Congleton, C.; Yerramsetti, S.M.; Gard, T.; Lazar, S.W. Mindfulness practice leads to increases in regional brain gray matter density. Psychiatry Res. 2011, 191, 36–43. [Google Scholar] [CrossRef] [Green Version]
- Vincent, J.L.; Kahn, I.; Snyder, A.Z.; Raichle, M.E.; Buckner, R.L. Evidence for a frontoparietal control system revealed by intrinsic functional connectivity. J. Neurophysiol. 2008, 100, 3328–3342. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Luh, W.-M.; Duan, W.; Zhou, G.D.; Weinschenk, G.; Anderson, A.K.; Dai, W. Longitudinal effects of meditation on brain resting-state functional connectivity. Sci. Rep. 2021, 11, 11361. [Google Scholar] [CrossRef]
- Biswal, B.; Yetkin, F.Z.; Haughton, V.M.; Hyde, J.S. Functional connectivity in the motor cortex of resting human brain using echoplanar MRI. Magn. Reson. Med. 1995, 34, 537–541. [Google Scholar] [CrossRef]
- Mooneyham, B.W.; Mrazek, M.D.; Mrazek, A.J.; Schooler, J.W. Signal or noise: Brain network interactions underlying the experience and training of mindfulness. Ann. N. Y. Acad. Sci. 2016, 1369, 240–256. [Google Scholar] [CrossRef]
- Dai, W.; Garcia, D.; de Bazelaire, C.; Alsop, D.C. Continuous flow-driven inversion for arterial spin labeling using pulsed radio frequency and gradient fields. Magn. Reson. Med. 2008, 60, 1488–1497. [Google Scholar] [CrossRef] [Green Version]
- Dai, W.; Varma, G.; Scheidegger, R.; Alsop, D.C. Quantifying fluctuations of resting state networks using arterial spin labeling perfusion MRI. J. Cereb. Blood Flow Metab. 2016, 36, 463–473. [Google Scholar] [CrossRef] [Green Version]
- Birn, R.M.; Murphy, K.; Bandettini, P.A. The effect of respiration variations on independent component analysis results of resting state functional connectivity. Hum. Brain Mapp. 2008, 29, 740–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Power, J.D.; Barnes, K.A.; Snyder, A.Z.; Schlaggar, B.L.; Petersen, S.E. Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. Neuroimage 2012, 59, 2142–2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamame, C.M.; Vidal, J.R.; Ossandon, T.; Jerbi, K.; Dalal, S.S.; Minotti, L.; Bertrand, O.; Kahane, P.; Lachaux, J.P. Reading the mind’s eye: Online detection of visuo-spatial working memory and visual imagery in the inferior temporal lobe. Neuroimage 2012, 59, 872–879. [Google Scholar] [CrossRef] [PubMed]
- Taylor, V.A.; Grant, J.; Daneault, V.; Scavone, G.; Breton, E.; Roffe-Vidal, S.; Courtemanche, J.; Lavarenne, A.S.; Beauregard, M. Impact of mindfulness on the neural responses to emotional pictures in experienced and beginner meditators. Neuroimage 2011, 57, 1524–1533. [Google Scholar] [CrossRef]
- Berkovich-Ohana, A.; Harel, M.; Hahamy, A.; Arieli, A.; Malach, R. Alterations in task-induced activity and resting-state fluctuations in visual and DMN areas revealed in long-term meditators. Neuroimage 2016, 135, 125–134. [Google Scholar] [CrossRef]
- Froeliger, B.; Garland, E.L.; Kozink, R.V.; Modlin, L.A.; Chen, N.-K.; McClernon, F.J.; Greeson, J.M.; Sobin, P. Meditation-state Functional Connectivity (msFC): Strengthening of the dorsal attention network and beyond. Evid.-Based Complement. Altern. Med. 2012, 2012, 680407. [Google Scholar] [CrossRef]
- Dosenbach, N.U.F.; Fair, D.A.; Miezin, F.M.; Cohen, A.L.; Wenger, K.K.; Dosenbach, R.A.T.; Fox, M.D.; Snyder, A.Z.; Vincent, J.L.; Raichle, M.E.; et al. Distinct brain networks for adaptive and stable task control in humans. Proc. Natl. Acad. Sci. USA 2007, 104, 11073–11078. [Google Scholar] [CrossRef] [Green Version]
- Goldman-Rakic, P.S. Architecture of the prefrontal cortex and the central executive. Ann. N. Y. Acad. Sci. 1995, 769, 71–83. [Google Scholar] [CrossRef]
- Brewer, J.A.; Worhunsky, P.D.; Gray, J.R.; Tang, Y.-Y.; Weber, J.; Kober, H. Meditation experience is associated with differences in default mode network activity and connectivity. Proc. Natl. Acad. Sci. USA 2011, 108, 20254–20259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, J.H.; Jung, W.H.; Kang, D.H.; Byun, M.S.; Kwon, S.J.; Choi, C.H.; Kwon, J.S. Increased default mode network connectivity associated with meditation. Neurosci. Lett. 2011, 487, 358–362. [Google Scholar] [CrossRef]
- Garrison, K.A.; Scheinost, D.; Constable, R.T.; Brewer, J.A. BOLD signal and functional connectivity associated with loving kindness meditation. Brain Behav. 2014, 4, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Jao, T.; Li, C.-W.; Vértes, P.E.; Wu, C.W.; Achard, S.; Hsieh, C.-H.; Liou, C.-H.; Chen, J.-H.; Bullmore, E.T. Large-scale functional brain network reorganization during taoist meditation. Brain Connect. 2016, 6, 9–24. [Google Scholar] [CrossRef]
- King, A.P.; Block, S.R.; Sripada, R.K.; Rauch, S.; Giardino, N.; Favorite, T.; Angstadt, M.; Kessler, D.; Welsh, R.; Liberzon, I. Altered Default mode network (Dmn) resting state functional connectivity following a mindfulness based exposure therapy for Posttraumatic stress disorder (Ptsd) in combat veterans of Afghanistan and Iraq. Depress. Anxiety 2016, 33, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Duan, W.; Bolo, N.R.; Tamminga, C.; Clementz, B.A.; Pearlson, G.D.; Keshavan, M.; Alsop, D.C.; Dai, W. Sensitivity of dynamic ASL and resting-state bold in patients with bipolar disorder. In Proceedings of the 28th ISMRM, Virtual Conference, 8–14 August 2020; Volume 28, p. 3874. [Google Scholar]
- Maleki, N.; Dai, W.; Alsop, D.C. Optimization of background suppression for arterial spin labeling perfusion imaging. Magn. Reson. Mater. Phys. Biol. Med. 2012, 25, 127–133. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Alsop, D.C.; Detre, J.A.; Dai, W. Global fluctuations of cerebral blood flow indicate a global brain network independent of systemic factors. J. Cereb. Blood Flow Metab. 2019, 39, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Alsop, D.; Detre, J.A. Reduced transit-time sensitivity in noninvasive magnetic resonance imaging of human cerebral blood flow. J. Cereb. Blood Flow Metab. 1996, 16, 1236–1249. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Alsop, D.; Li, L.; Listerud, J.; Gonzalez-At, J.B.; Schnall, M.D.; Detre, J.A. Comparison of quantitative perfusion imaging using arterial spin labeling at 1.5 and 4.0 Tesla. Magn. Reson. Med. 2002, 48, 242–254. [Google Scholar] [CrossRef]
- Buxton, R.B.; Frank, L.R.; Wong, E.C.; Siewert, B.; Warach, S.; Edelman, R.R. A general kinetic model for quantitative perfusion imaging with arterial spin labeling. Magn. Reson. Med. 1998, 40, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Lacalle-Aurioles, M.; Aleman-Gomez, Y.; Guzman-De-Villoria, J.A.; Cruz-Orduna, I.; Olazaran, J.; Mateos-Perez, J.M.; Martino, M.E.; Desco, M. Is the cerebellum the optimal reference region for intensity normalization of perfusion MR studies in early Alzheimer’s disease? PLoS ONE 2013, 8, e81548. [Google Scholar]
- Kilpatrick, L.A.; Suyenobu, B.Y.; Smith, S.R.; Bueller, J.A.; Goodman, T.; Creswell, J.D.; Tillisch, K.; Mayer, E.A.; Naliboff, B.D. Impact of mindfulness-based stress reduction training on intrinsic brain connectivity. Neuroimage 2011, 56, 290–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeley, W.W.; Menon, V.; Schatzberg, A.F.; Keller, J.; Glover, G.H.; Kenna, H.; Reiss, A.L.; Greicius, M.D. Dissociable intrinsic connectivity networks for salience processing and executive control. J. Neurosci. 2007, 27, 2349–2356. [Google Scholar] [CrossRef]
- Hasenkamp, W.; Barsalou, L.W. Effects of meditation experience on functional connectivity of distributed brain networks. Front. Hum. Neurosci. 2012, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- Uddin, L. Salience processing and insular cortical function and dysfunction. Nat. Rev. Neurosci. 2015, 16, 55–61. [Google Scholar] [CrossRef]
- Menon, V.; Uddin, L.Q. Saliency, switching, attention and control: A network model of insula function. Brain Struct. Funct. 2010, 214, 655–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sridharan, D.; Levitin, D.J.; Menon, V. A critical role for the right fronto-insular cortex in switching between central-executive and default-mode networks. Proc. Natl. Acad. Sci. USA 2008, 105, 12569–12574. [Google Scholar] [CrossRef] [Green Version]
- Taren, A.A.; Gianaros, P.J.; Greco, C.M.; Lindsay, E.; Fairgrieve, A.; Brown, K.W.; Rosen, R.K.; Ferris, J.L.; Julson, E.; Marsland, A.L.; et al. Mindfulness meditation training and executive control network resting state functional connectivity: A randomized controlled trial. Psychosom. Med. 2017, 79, 674–683. [Google Scholar] [CrossRef]
- He, B.J.; Snyder, A.Z.; Vincent, J.L.; Epstein, A.; Shulman, G.L.; Corbetta, M. Breakdown of functional connec-tivity in frontoparietal networks underlies behavioral deficits in spatial neglect. Neuron 2007, 53, 905–918. [Google Scholar] [CrossRef] [Green Version]
- Kemmer, P.B.; Guo, Y.; Wang, Y.; Pagnoni, G. Network-based characterization of brain functional connectivity in Zen practitioners. Front. Psychol. 2015, 6, 603. [Google Scholar] [CrossRef] [Green Version]
- Doll, A.; Holzel, B.K.; Boucard, C.C.; Wohlschlager, A.M.; Sorg, C. Mindfulness is associated with intrinsic functional connectivity between default mode and salience networks. Front. Hum. Neurosci. 2015, 9, 461. [Google Scholar] [CrossRef] [Green Version]
- Bauer, C.C.C.; Whitfield-Gabrieli, S.; Díaz, J.L.; Pasaye, E.H.; Barrios, F.A. From state to trait meditation: Reconfiguration of central executive and default mode networks. Eneuro 2019, 6. [Google Scholar] [CrossRef] [Green Version]
- Brandmeyer, T.; Delorme, A. Reduced mind wandering in experienced meditators and associated EEG correlates. Exp. Brain Res. 2018, 236, 2519–2528. [Google Scholar] [CrossRef]
- Posner, M.; DiGirolamo, G. Executive attention: Conflict, target detection, and cognitive control. In The Attentive Brain; Parasuraman, R., Ed.; MIT Press: Cambridge, MA, USA, 1988. [Google Scholar]
- Hasenkamp, W.; Wilson-Mendenhall, C.D.; Duncan, E.; Barsalou, L.W. Mind wandering and attention during focused meditation: A fine-grained temporal analysis of fluctuating cognitive states. Neuroimage 2012, 59, 750–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robson, P.M.; Madhuranthakam, A.J.; Dai, W.; Pedrosa, I.; Rofsky, N.M.; Alsop, D. Strategies for reducing respiratory motion artifacts in renal perfusion imaging with arterial spin labeling. Magn. Reson. Med. 2009, 61, 1374–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taso, M.; Guidon, A.; Alsop, D.C. Influence of background suppression and retrospective realignment on free-breathing renal perfusion measurement using pseudo-continuous ASL. Magn. Reson. Med. 2019, 81, 2439–2449. [Google Scholar] [CrossRef] [PubMed]
- Buxton, R.B.; Uludag, K.; Dubowitz, D.J.; Liu, T. Modeling the hemodynamic response to brain activation. Neuroimage 2004, 23, S220–S233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.G.; Ugurbil, K. Comparison of blood oxygenation and cerebral blood flow effects in fMRI: Estimation of relative oxygen consumption change. Magn. Reson. Med. 1997, 38, 59–65. [Google Scholar]
- Ances, B.M.; Leontiev, O.; Perthen, J.E.; Liang, C.; Lansing, A.E.; Buxton, R.B. Regional differences in the coupling of cerebral blood flow and oxygen metabolism changes in response to activation: Implications for BOLD-fMRI. Neuroimage 2008, 39, 1510–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, T.L.; Kwong, K.K.; Weisskoff, R.M.; Rosen, B.R. Calibrated functional MRI: Mapping the dynamics of oxidative metabolism. Proc. Natl. Acad. Sci. USA 1998, 95, 1834–1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| At Follow-Up | |
|---|---|
| Total number of included subjects * | 10 |
| Age range | 19 years (8), 20 years (2) |
| Gender | Male (6), Female (4) |
| Handedness | Right (7), Left (3) |
| Total practice time (minutes) | Range: (135, 1620) 574 ± 465 |
| Time duration (days) | Range: (45, 72) 66 ± 4 |
| N Voxels |
Local Zscore | Peak-t MNI Coordinates | Mean Beta ± Std. | Anatomical Locations | % Cluster | % Region | |
|---|---|---|---|---|---|---|---|
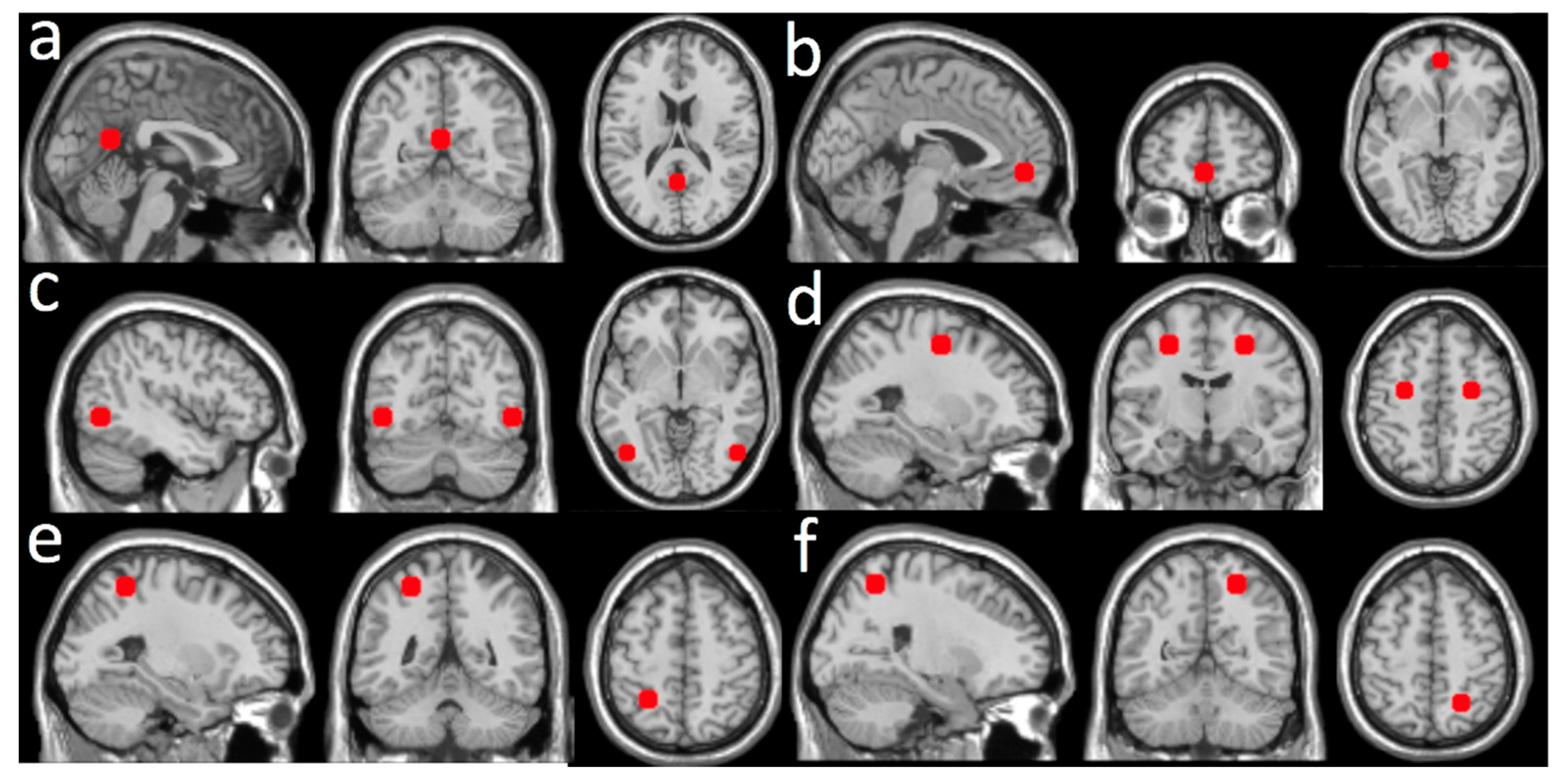
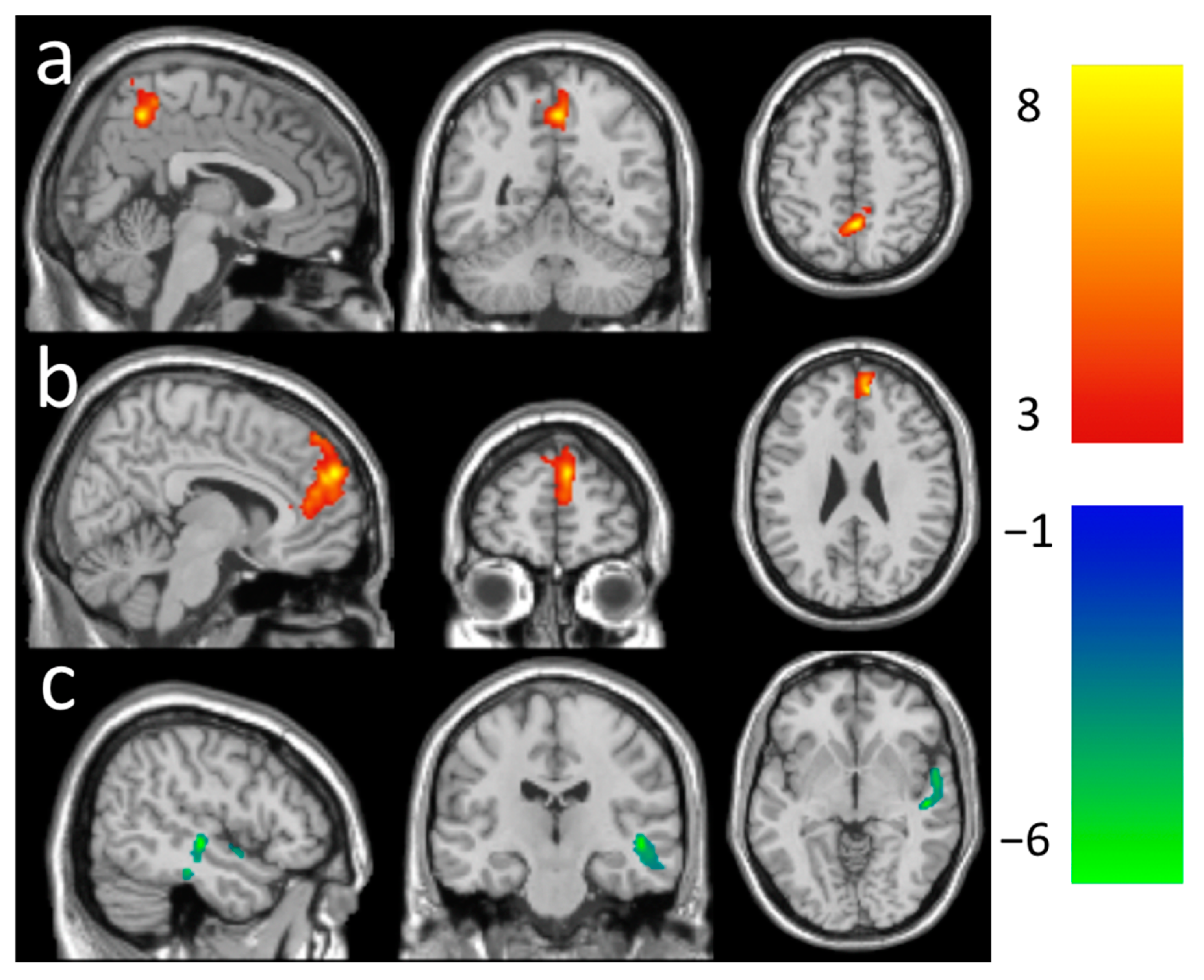
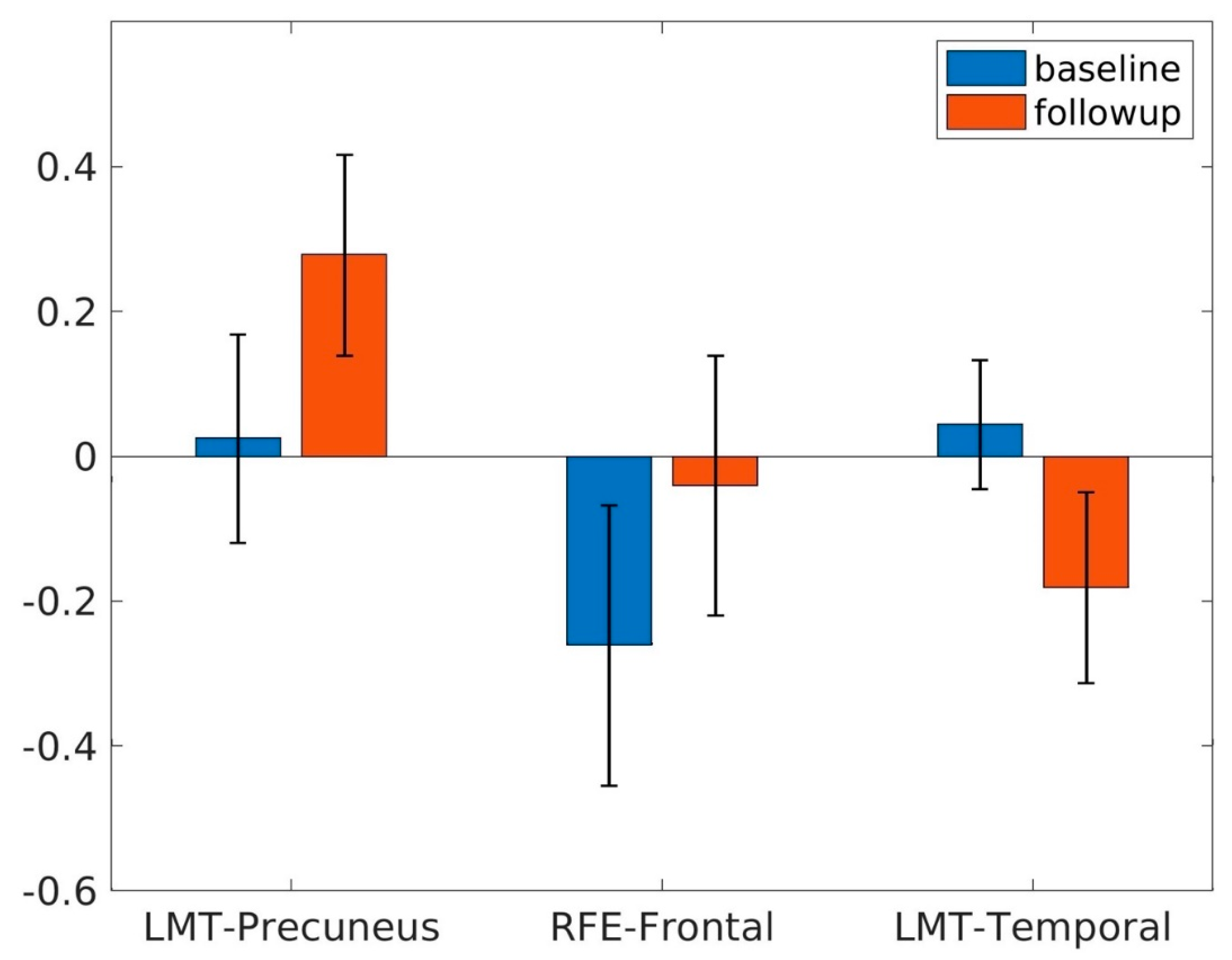
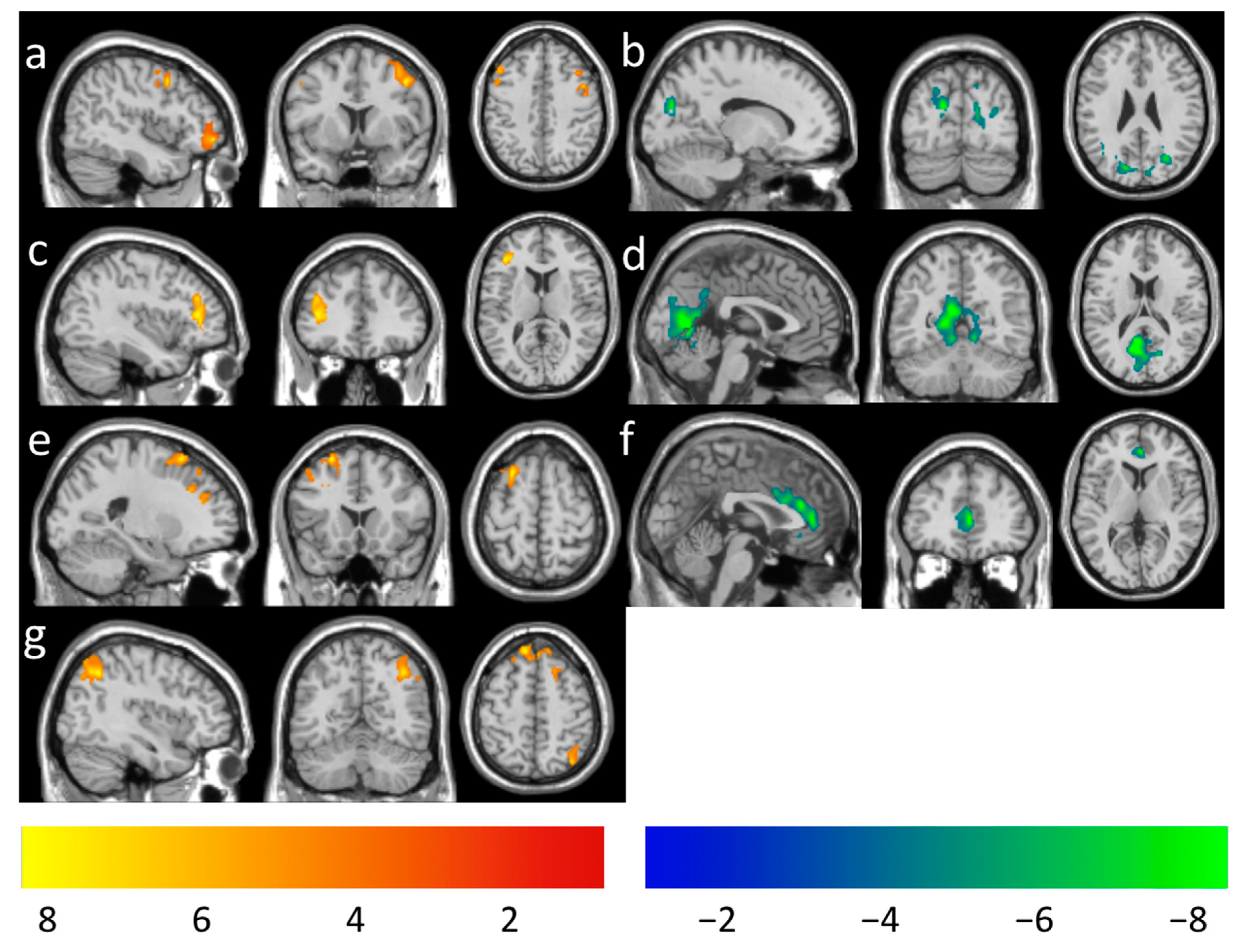
| Increased | 440 | 4.14 | 4, −52, 52 | 0.25 ± 0.03 | Parietal Lobe | ||
| LMT rsFC (Figure 2a) | Precuneus_R | 60.68 | 8.18 | ||||
| Precuneus_L | 34.77 | 4.34 | |||||
| Paracentral_Lobule_R | 2.73 | 1.44 | |||||
| Limbic System | |||||||
| Cingulum_Mid_R | 1.82 | 0.36 | |||||
| Increased | 1038 | 4.36 | 10, 50, 26 | 0.22 ± 0.04 | Frontal Lobe | ||
|
RFE rsFC (Figure 2b) | Frontal_Sup_Medial_R | 49.52 | 24.09 | ||||
| Frontal_Sup_Medial_L | 19.46 | 6.75 | |||||
| Frontal_Sup_L | 5.01 | 1.44 | |||||
| Frontal_Sup_R | 4.72 | 1.21 | |||||
| Limbic System | |||||||
| Cingulum_Ant_R | 20.81 | 16.45 | |||||
| Decreased | 499 | 3.82 | 50, −20, −6 | −0.23 ± 0.04 | Temporal Lobe | ||
| LMT rsFC (Figure 2c) | Temporal_Sup_R | 48.30 | 7.67 | ||||
| Temporal_Mid_R | 24.65 | 2.79 | |||||
| Temporal_Inf_R | 16.63 | 2.33 | |||||
| Temporal_Pole_Sup_R | 6.81 | 2.54 | |||||
| Heschl_R | 2.00 | 4.02 | |||||
| Temporal_Pole_Mid_R | 1.00 | 0.42 |
| Association with Practice Time | N Voxels |
Local Zscore | Peak-t MNI Coordinates | Mean Beta ± Std. (×10−4) (/minute) | Anatomical Locations | % Cluster | % Region |
|---|---|---|---|---|---|---|---|
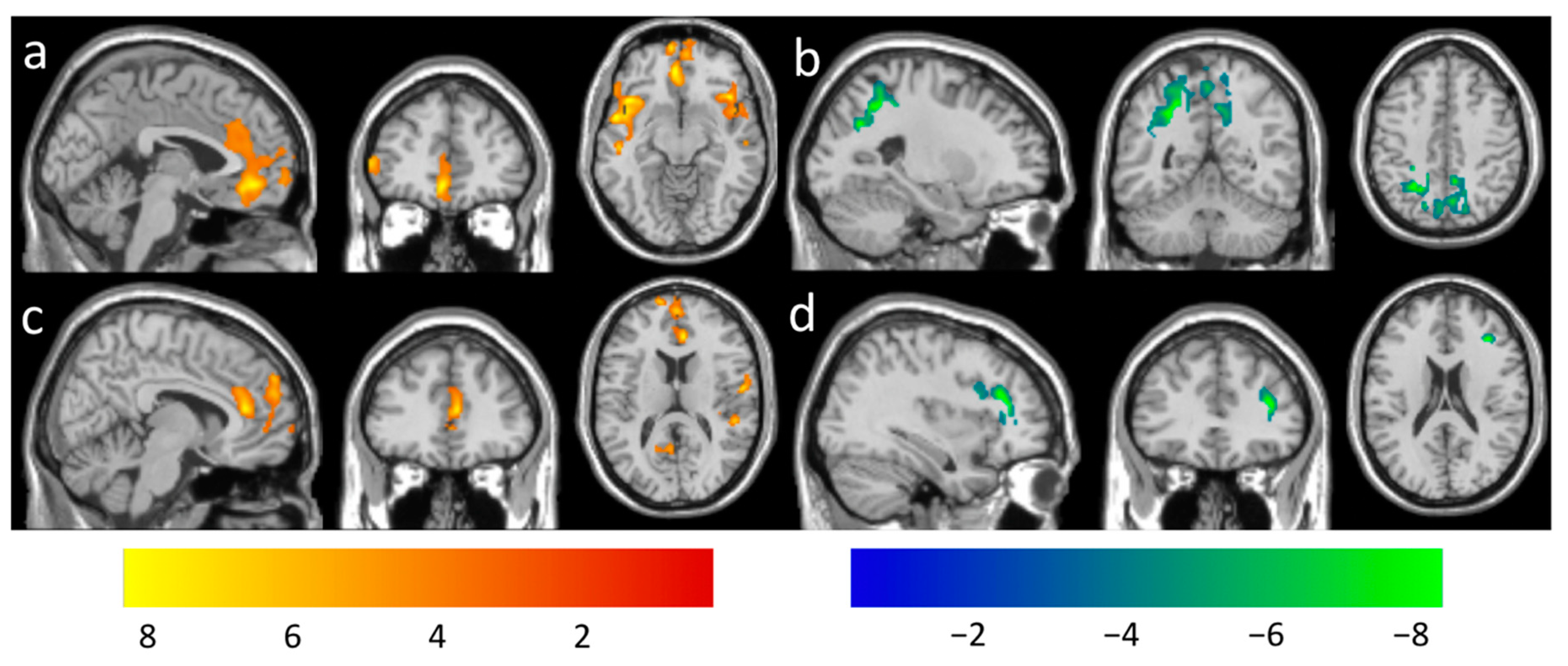
| Positive for | 2162 | 4.45 | −54, 4, −10 | 6.45 ± 1.45 | Insula | ||
| PCC rsFC | Insula_L | 22.48 | 26.16 | ||||
| (Figure 4a) | Temporal Lobe | ||||||
| Temporal_Pole_Sup_L | 17.44 | 29.34 | |||||
| Temporal_Sup_L | 15.17 | 14.29 | |||||
| Temporal_Mid_L | 5.18 | 2.17 | |||||
| Heschl_L | 2.78 | 26.67 | |||||
| Frontal Lobe | |||||||
| Frontal_Inf_Tri_L | 14.52 | 12.42 | |||||
| Frontal_Inf_Orb_L | 11.05 | 14.14 | |||||
| Frontal_Inf_Oper_L | 5.64 | 11.75 | |||||
| Rolandic_Oper_L | 3.42 | 7.47 | |||||
| Frontal_Mid_L | 1.16 | 0.51 | |||||
| 1720 | 4.10 | −4, −42, −12 | 7.14 ± 1.28 | Limbic System | |||
| Cingulum_Ant_L | 23.95 | 29.43 | |||||
| Cingulum_Mid_L | 3.84 | 3.40 | |||||
| Cingulum_Ant_R | 2.33 | 3.05 | |||||
| Frontal Lobe | |||||||
| Frontal_Sup_Medial_L | 17.44 | 10.03 | |||||
| Frontal_Med_Orb_R | 15.93 | 32.01 | |||||
| Frontal_Med_Orb_L | 15.81 | 37.83 | |||||
| Rectus_L | 6.63 | 13.38 | |||||
| Frontal_Sup_Medial_R | 6.63 | 5.34 | |||||
| Rectus_R | 4.59 | 10.60 | |||||
| Frontal_Sup_L | 2.21 | 1.06 | |||||
| 897 | 4.12 | 44, 22, −10 | 5.86 ± 1.32 | Insula | |||
| Insula_R | 34.23 | 17.34 | |||||
| Temporal Lobe | |||||||
| Temporal_Pole_Sup_R | 24.64 | 16.52 | |||||
| Temporal_Pole_Mid_R | 4.91 | 3.71 | |||||
| Temporal_Mid_R | 1.67 | 0.34 | |||||
| Frontal Lobe | |||||||
| Frontal_Inf_Orb_R | 16.50 | 8.67 | |||||
| Frontal_Inf_Oper_R | 13.60 | 8.72 | |||||
| Frontal_Inf_Tri_R | 3.68 | 1.53 | |||||
| 395 | 3.70 | 44, −24, 18 | 5.11 ± 0.86 | Temporal Lobe | |||
| Temporal_Sup_R | 42.53 | 5.35 | |||||
| Heschl_R | 14.68 | 23.29 | |||||
| Temporal_Mid_R | 1.52 | 0.14 | |||||
| Frontal Lobe | |||||||
| Rolandic_Oper_R | 30.13 | 8.94 | |||||
| Insula | |||||||
| Insula_R | 11.14 | 2.49 | |||||
| Negative for | 2892 | 5.38 | −26, −52, 42 | −6.22 ± 1.38 | Parietal Lobe | ||
| PCC rsFC (Figure 4b) | Precuneus_L | 26.38 | 21.63 | ||||
| Precuneus_R | 26.00 | 23.03 | |||||
| Parietal_Sup_L | 8.78 | 12.30 | |||||
| Parietal_Inf_L | 6.40 | 7.56 | |||||
| Angular_L | 3.63 | 8.95 | |||||
| Parietal_Sup_R | 3.56 | 4.64 | |||||
| Postcentral_L | 1.28 | 0.95 | |||||
| Occipital Lobe | |||||||
| Occipital_Mid_R | 11.45 | 15.78 | |||||
| Occipital_Sup_R | 4.88 | 9.98 | |||||
| Occipital_Mid_L | 3.67 | 3.24 | |||||
| Occipital_Sup_L | 1.38 | 2.93 | |||||
| Cuneus_R | 1.35 | 2.74 | |||||
| Positive with | 1108 | 4.14 | 8, 36, 14 | 5.66 ± 1.11 | Frontal Lobe | ||
| vmPFC rsFC (Figure 4c) | Frontal_Sup_Medial_R | 33.39 | 17.34 | ||||
| Frontal_Sup_Medial_L | 27.71 | 10.26 | |||||
| Frontal_Sup_L | 3.79 | 1.17 | |||||
| Limbic System | |||||||
| Cingulum_Ant_R | 27.8 | 23.46 | |||||
| Cingulum_Ant_L | 5.42 | 4.29 | |||||
| Cingulum_Mid_R | 1.17 | 0.59 | |||||
| 916 | 3.94 | 44, −26, 2 | 5.76 ± 1.03 | Temporal Lobe | |||
| Temporal_Sup_R | 61.68 | 17.99 | |||||
| Temporal_Mid_R | 9.28 | 1.93 | |||||
| Temporal_Inf_R | 5.02 | 1.29 | |||||
| Heschl_R | 2.84 | 10.44 | |||||
| Frontal Lobe | |||||||
| Rolandic_Oper_R | 11.90 | 8.19 | |||||
| Precentral_R | 1.20 | 0.33 | |||||
| Parietal Lobe | |||||||
| Postcentral_R | 8.08 | 1.94 | |||||
| 381 | 3.18 | −2, −58, 18 | 6.27 ± 0.96 | Parietal Lobe | |||
| Precuneus_L | 45.14 | 4.88 | |||||
| Precuneus_R | 8.14 | 0.95 | |||||
| Occipital Lobe | |||||||
| Calcarine_L | 20.73 | 3.50 | |||||
| Cuneus_L | 9.71 | 2.42 | |||||
| Lingual_L | 1.84 | 0.33 | |||||
| Limbic System | |||||||
| Cingulum_Post_L | 11.02 | 9.07 | |||||
| Cingulum_Mid_R | 1.84 | 0.32 | |||||
| Cingulum_Post_R | 1.57 | 1.79 | |||||
| Negative for | 377 | 5.16 | 36, 34, 18 | −6.13 ± 1.31 | Frontal Lobe | ||
| vmPFC rsFC (Figure 4d) | Frontal_Inf_Tri_R | 42.18 | 7.39 | ||||
| Frontal_Mid_R | 27.06 | 2.00 | |||||
| Frontal_Inf_Oper_R | 23.08 | 6.22 | |||||
| Insula | |||||||
| Insula_R | 7.69 | 1.64 | |||||
| 361 | 3.96 | 22, −68, 52 | 4.62 ± 0.86 | Parietal Lobe | |||
| Parietal_Sup_R | 72.85 | 11.84 | |||||
| Angular_R | 4.99 | 1.03 | |||||
| Postcentral_R | 1.94 | 0.18 | |||||
| Occipital Lobe | |||||||
| Occipital_Sup_R | 17.73 | 4.53 | |||||
| Occipital_Mid_R | 2.22 | 0.38 | |||||
| Positive for | 596 | 3.78 | 44, 8, 44 | 5.79 ± 1.13 | Frontal Lobe | ||
| RMT rsFC (Figure 5a) | Frontal_Mid_R | 62.58 | 7.31 | ||||
| Precentral_R | 26.01 | 4.58 | |||||
| Frontal_Sup_R | 11.41 | 1.68 | |||||
| 484 | 3.76 | −50, 28, 28 | 6.33 ± 0.98 | Frontal Lobe | |||
| Frontal_Inf_Tri_L | 47.73 | 9.13 | |||||
| Frontal_Mid_L | 40.91 | 4.06 | |||||
| Frontal_Inf_Oper_L | 7.23 | 3.37 | |||||
| Precentral_L | 4.13 | 0.57 | |||||
| 330 | 4.34 | 46, 50, −8 | 5.69 ± 1.25 | Frontal Lobe | |||
| Frontal_Inf_Orb_R | 36.36 | 7.03 | |||||
| Frontal_Mid_R | 28.28 | 1.82 | |||||
| Frontal_Mid_Orb_R | 20.61 | 6.70 | |||||
| Frontal_Inf_Tri_R | 8.48 | 1.30 | |||||
| Frontal_Sup_R | 6.36 | 0.52 | |||||
| Negative for | 724 | 3.89 | 28, −68, 22 | −6.75 ± 1.40 | Parietal Lobe | ||
| RMT rsFC (Figure 5b) | Precuneus_R | 23.07 | 5.11 | ||||
| Parietal_Sup_R | 8.29 | 2.70 | |||||
| Occipital Lobe | |||||||
| Occipital_Mid_R | 21.41 | 7.39 | |||||
| Calcarine_R | 16.44 | 6.39 | |||||
| Occipital_Sup_R | 15.88 | 8.14 | |||||
| Cuneus_R | 14.92 | 7.58 | |||||
| 571 | 4.29 | −14, −78, 24 | −6.59 ± 1.10 | Occipital Lobe | |||
| Occipital_Sup_L | 40.98 | 17.13 | |||||
| Occipital_Mid_L | 22.94 | 4.01 | |||||
| Cuneus_L | 11.38 | 4.26 | |||||
| Calarine_L | 3.33 | 0.84 | |||||
| Parietal Lobe | |||||||
| Angular_L | 16.99 | 8.27 | |||||
| Parietal_Sup_L | 1.40 | 0.39 | |||||
| Temporal Lobe | |||||||
| Temporal_Mid_L | 2.80 | 0.32 | |||||
| Positive for | 403 | 3.47 | −38, 34, 12 | 6.10 ± 1.26 | Frontal Lobe | ||
| LSPL rsFC (Figure 5c) | Frontal_Mid_L | 52.85 | 4.38 | ||||
| Frontal_Inf_Tri_L | 40.94 | 6.52 | |||||
| Frontal_Sup_L | 6.20 | 0.69 | |||||
| Negative for | 3681 | 5.55 | −2, −64, 4 | −6.49 ± 1.35 | Occipital Lobe | ||
| LSPL rsFC (Figure 5d) | Calcarine_L | 24.67 | 40.21 | ||||
| Lingual_L | 16.25 | 28.54 | |||||
| Lingual_R | 14.34 | 22.96 | |||||
| Calcarine_R | 12.85 | 25.42 | |||||
| Cuneus_L | 7.23 | 17.43 | |||||
| Parietal Lobe | |||||||
| Precuneus_L | 9.05 | 9.44 | |||||
| Precuneus_R | 1.52 | 1.72 | |||||
| Cerebelum | |||||||
| Cerebelum_4_5_L | 3.15 | 10.31 | |||||
| Vermis_4_5 | 1.98 | 10.98 | |||||
| Cerebelum_4_5_R | 1.33 | 5.69 | |||||
| Vermis_6 | 1.09 | 10.78 | |||||
| Limbic System | |||||||
| Cingulum_Post_L | 2.50 | 19.87 | |||||
| Positive for | 727 | 4.15 | −24, 18, 60 | −5.95 ± 1.29 | Frontal Lobe | ||
| RFE rsFC (Figure 5e) | Frontal_mid_L | 51.03 | 7.63 | ||||
| Frontal_Sup_L | 33.70 | 6.81 | |||||
| Frontal_Inf_Oper_L | 7.57 | 5.30 | |||||
| Precentral_L | 4.68 | 0.96 | |||||
| Frontal_Inf_Tri_L | 3.03 | 0.87 | |||||
| Negative for | 1010 | 4.33 | 0, 32, 24 | 5.68 ± 1.20 | Limbic System | ||
| RFE rsFC (Figure 5f) | Cingulum_Ant_L | 56.63 | 40.86 | ||||
| Cingulum_Mid_L | 20.69 | 10.77 | |||||
| Cingulum_Ant_R | 25.54 | 11.96 | |||||
| Cingulum_Mid_R | 2.28 | 1.04 | |||||
| Frontal Lobe | |||||||
| Frontal_sup_Medial_L | 4.16 | 1.40 | |||||
| Positive for | 546 | 3.92 | 40, −56, 46 | 5.84 ± 1.26 | Parietal Lobe | ||
| LFE rsFC (Figure 5g) | Angular_R | 70.51 | 21.97 | ||||
| Parietal_Inf_R | 17.40 | 7.06 | |||||
| Parietal_Sup_R | 10.99 | 2.70 | |||||
| 543 | 4.39 | 26, 20, 64 | 5.26 ± 0.88 | Frontal Lobe | |||
| Frontal_Sup_R | 46.04 | 6.16 | |||||
| Frontal_Sup_Medial_L | 23.39 | 4.24 | |||||
| Frontal_Sup_Medial_R | 25.65 | 3.98 | |||||
| Frontal_Sup_L | 11.42 | 1.72 | |||||
| Supp_Motor_Area_R | 1.66 | 0.38 | |||||
| Frontal_Mid_L | 1.66 | 0.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Luh, W.-M.; Duan, W.; Zhou, T.D.; Zhao, L.; Weinschenk, G.; Anderson, A.K.; Dai, W. The Longitudinal Effect of Meditation on Resting-State Functional Connectivity Using Dynamic Arterial Spin Labeling: A Feasibility Study. Brain Sci. 2021, 11, 1263. https://doi.org/10.3390/brainsci11101263
Zhang Z, Luh W-M, Duan W, Zhou TD, Zhao L, Weinschenk G, Anderson AK, Dai W. The Longitudinal Effect of Meditation on Resting-State Functional Connectivity Using Dynamic Arterial Spin Labeling: A Feasibility Study. Brain Sciences. 2021; 11(10):1263. https://doi.org/10.3390/brainsci11101263
Chicago/Turabian StyleZhang, Zongpai, Wen-Ming Luh, Wenna Duan, Tony D. Zhou, Li Zhao, George Weinschenk, Adam K. Anderson, and Weiying Dai. 2021. "The Longitudinal Effect of Meditation on Resting-State Functional Connectivity Using Dynamic Arterial Spin Labeling: A Feasibility Study" Brain Sciences 11, no. 10: 1263. https://doi.org/10.3390/brainsci11101263