
Piglet Gut and in-Barn Manure from Farms on a Raised without Antibiotics Program Display Reduced Antimicrobial Resistance but an Increased Prevalence of Pathogens

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
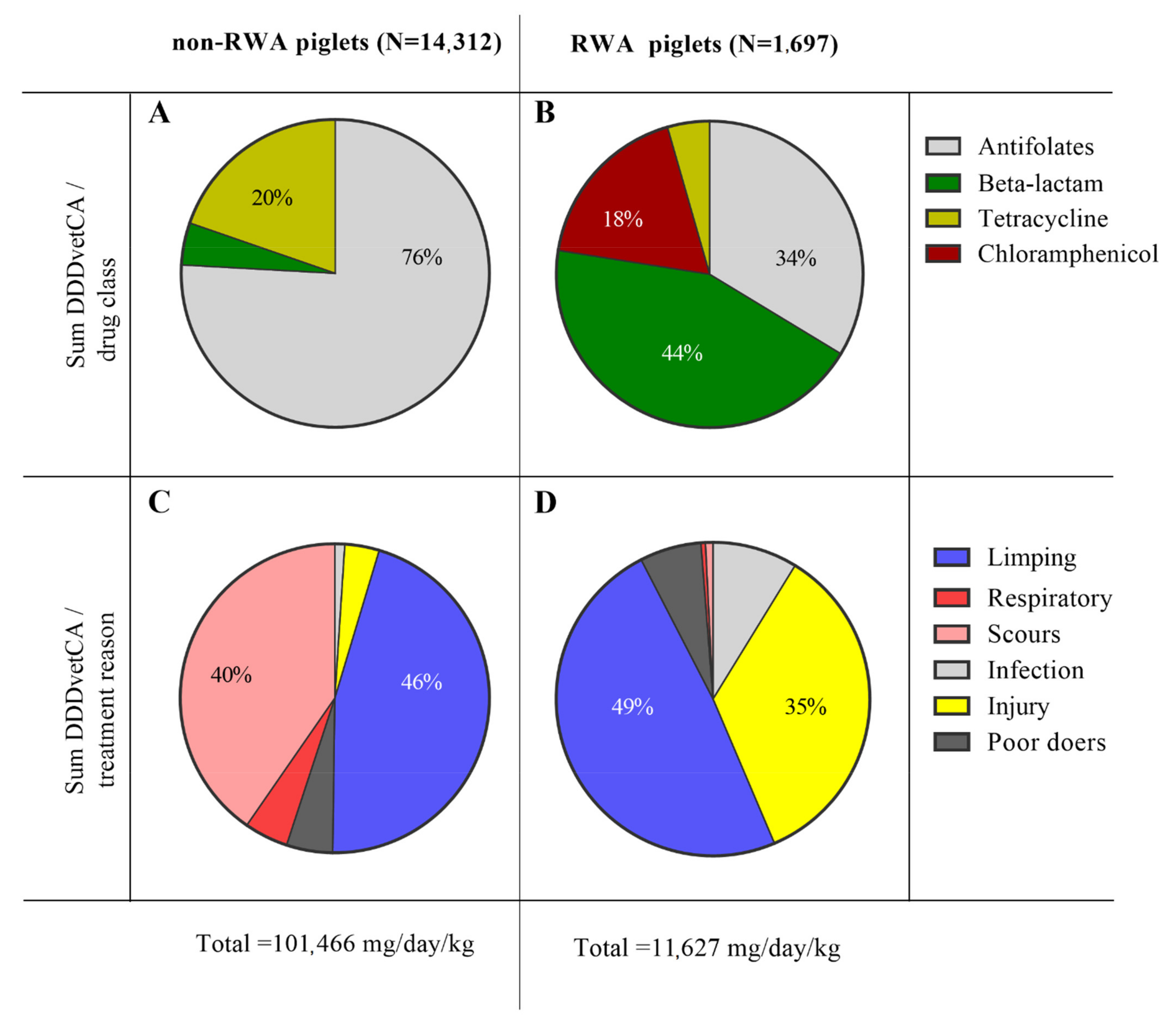
2.1. Decreased Antibiotic Utilization Changed the Occurrence of Scours and Injury in RWA Piglets
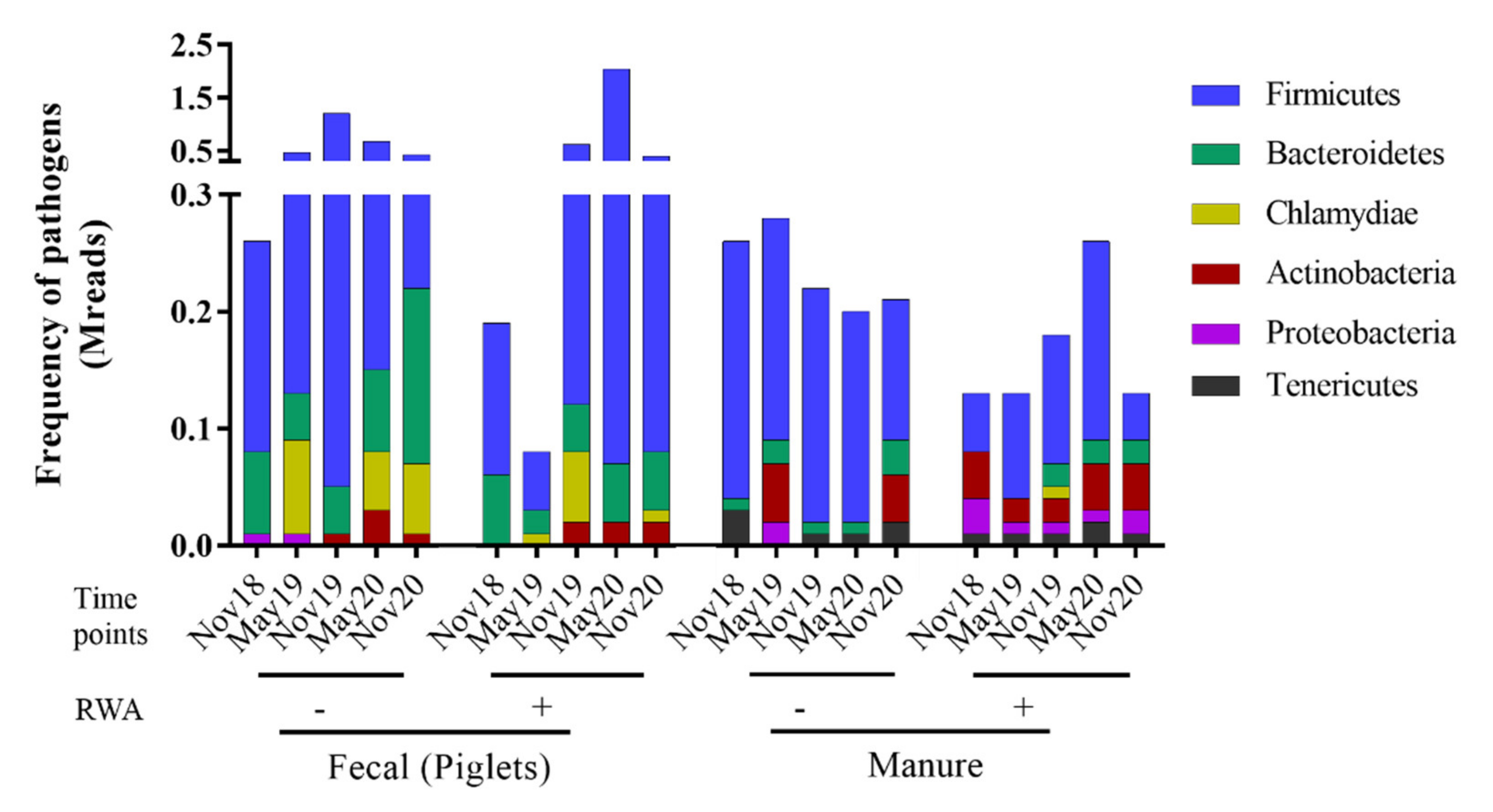
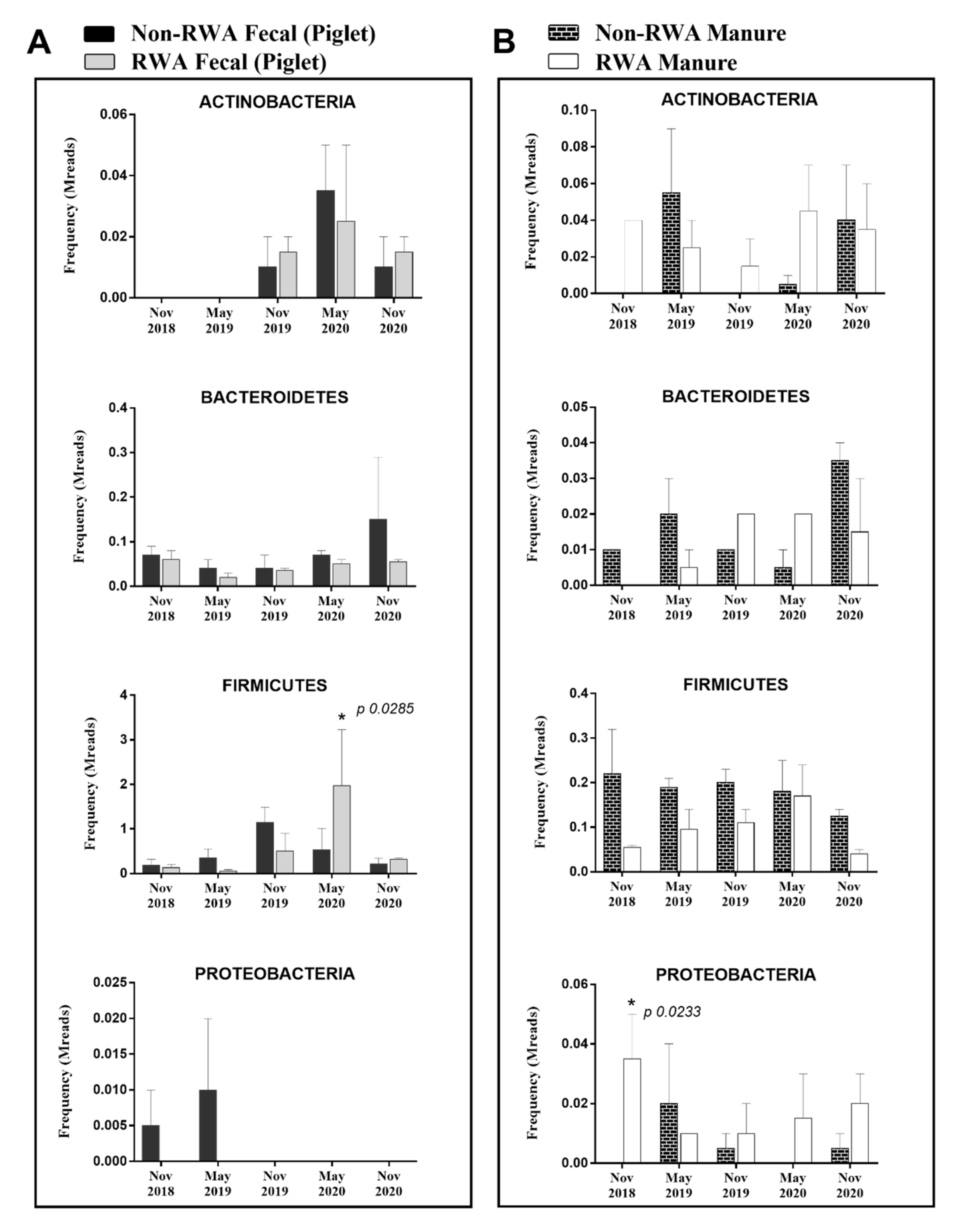
2.2. Pathogenic Firmicutes and Proteobacteria were More Prevalent in Barn Manure and Piglet Fecal Samples from RWA Barns
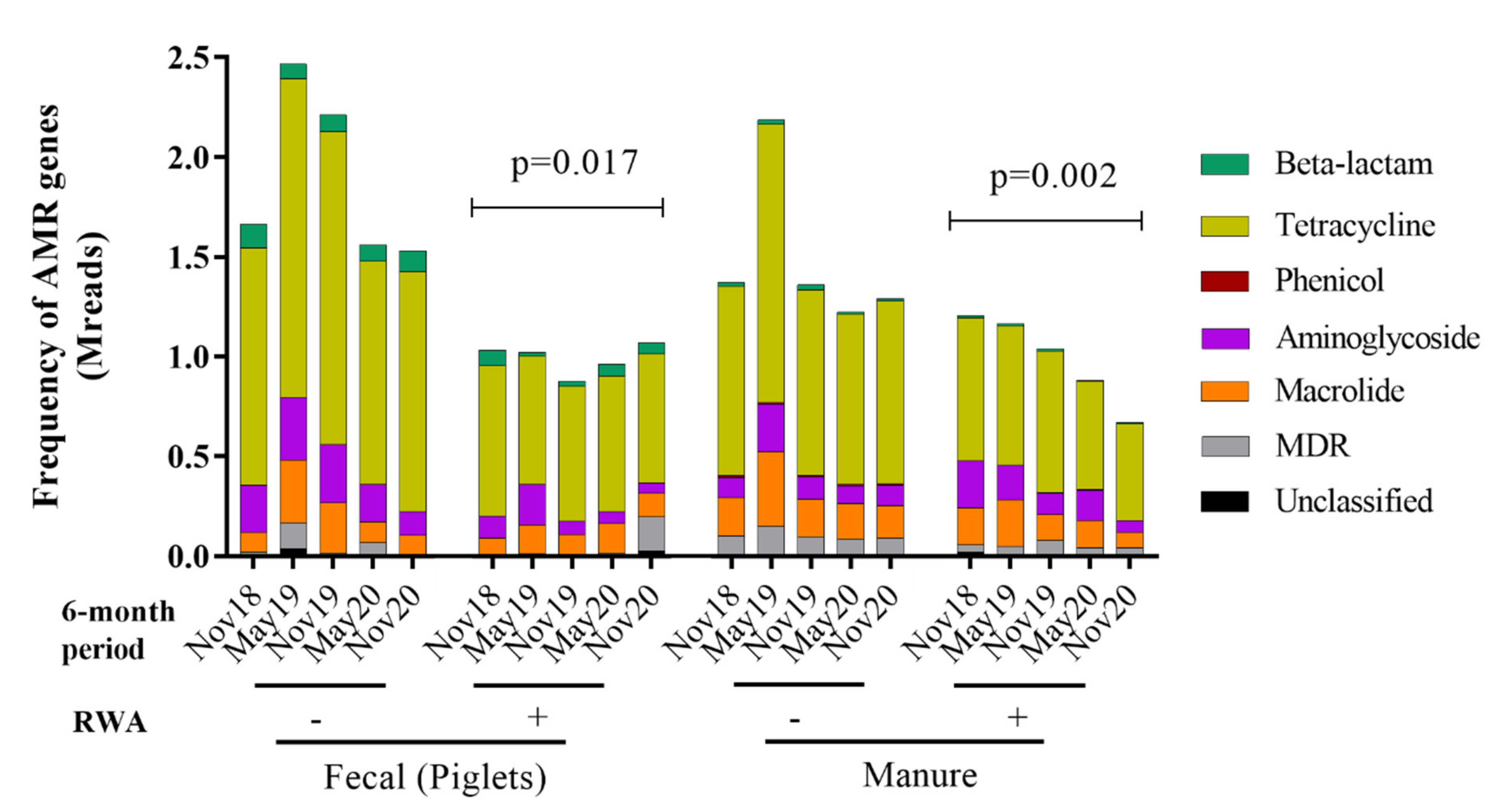
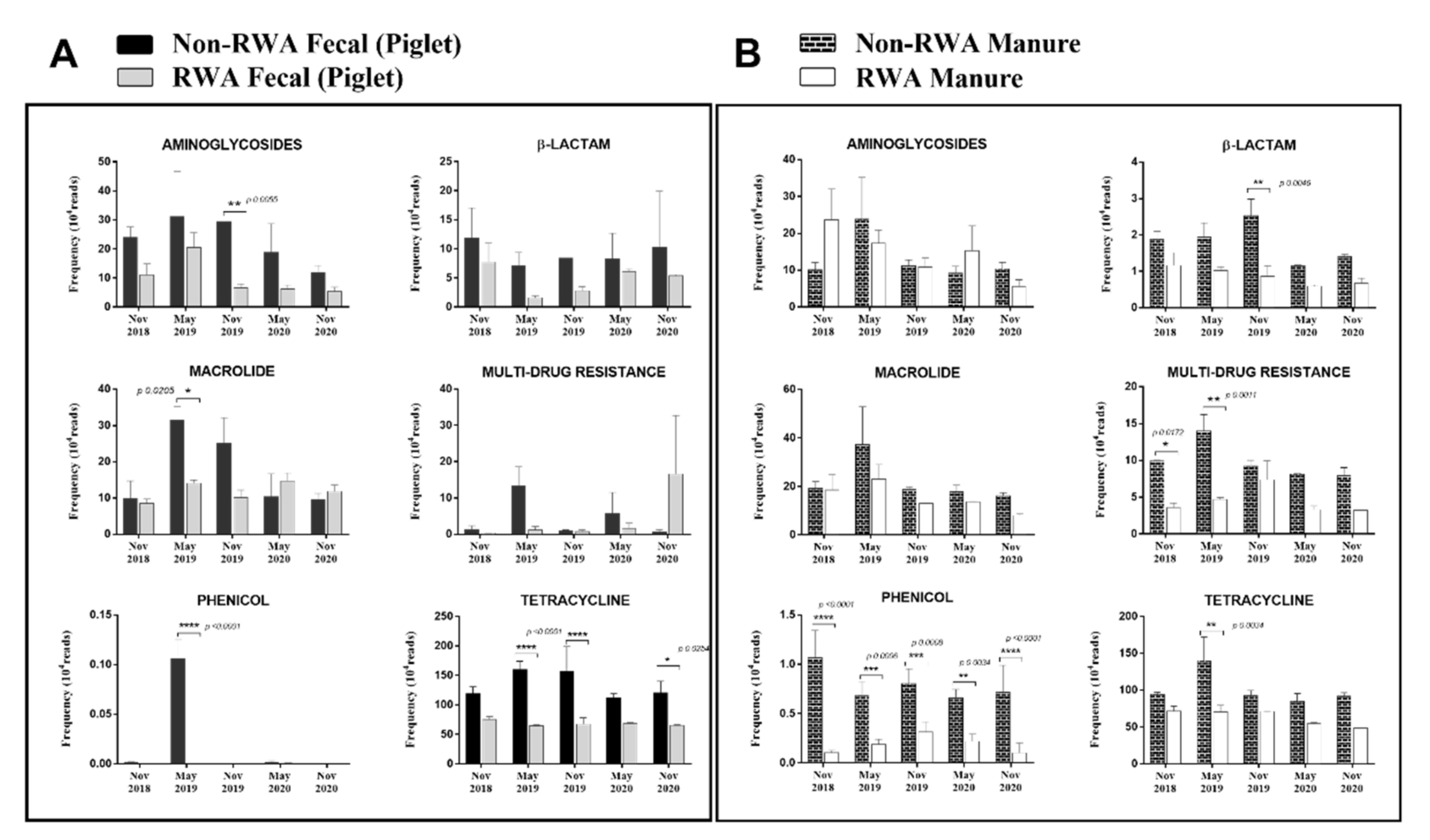
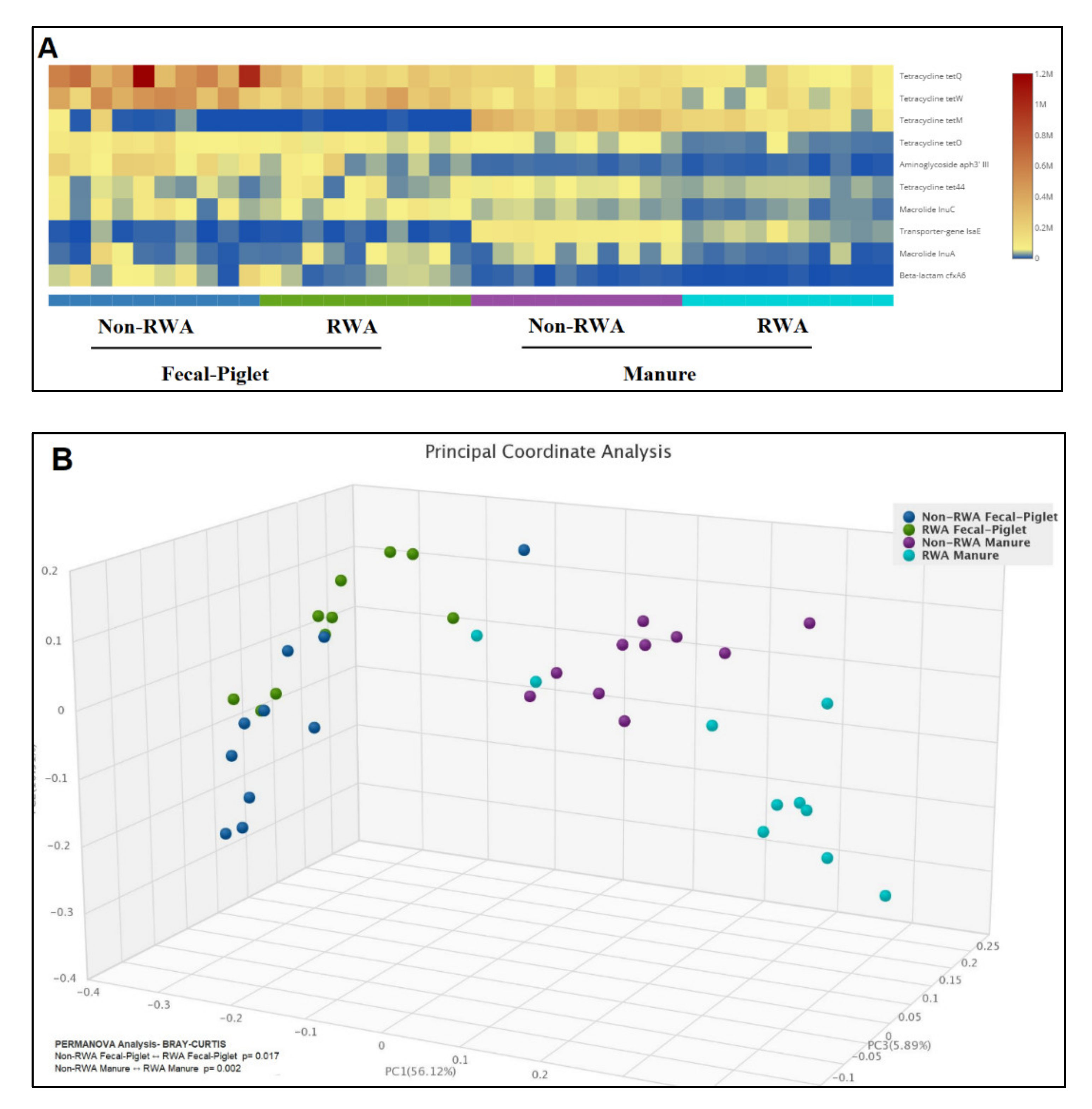
2.3. RWA Practices Reduced AMR in Piglet Gut and Barn Manure
3. Discussion
4. Materials and Methods
4.1. Experimental Design and Sample Collection
4.2. Whole-Genome Sequencing and Sequence Analyses
4.3. Profiling the Prevalence of Pathogens
4.4. Profiling the Resistome
4.5. Statistical Analysis
4.6. Data Availability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Danner, M.C.; Robertson, A.; Behrends, V.; Reiss, J. Antibiotic pollution in surface fresh waters: Occurrence and effects. Sci. Total Environ. 2019, 664, 793–804. [Google Scholar] [CrossRef]
- ESVAC. Sales of Veterinary Antimicrobial Agents in 31 European Countries in 2017: Trends from 2010 to 2017; Centers for Disease Control and Prevention, Amsterdam, The Netherlands: 2019. Available online: https://www.ema.europa.eu/en (accessed on 15 October 2019).
- PHAC. Canadian Antimicrobial Resistance Surveillance System Report 2016; Public Health Agency of Canada: Ottawa, ON, Canada, 2016. Available online: https://www.canada.ca/en/public-health/services/publications (accessed on 12 September 2016).
- Van Boeckel, T.P.; Pires, J.; Silvester, R.; Zhao, C.; Song, J.; Criscuolo, N.G.; Gilbert, M.; Bonhoeffer, S.; Laxminarayan, R. Global trends in antimicrobial resistance in animals in low- and middle-income countries. Science 2019, 365, eaaw1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castanon, J.I. History of the use of antibiotic as growth promoters in European poultry feeds. Poult. Sci. 2007, 86, 2466–2471. [Google Scholar] [CrossRef] [PubMed]
- Escudeiro, P.; Pothier, J.; Dionisio, F.; Nogueira, T. Antibiotic Resistance Gene Diversity and Virulence Gene Diversity Are Correlated in Human Gut and Environmental Microbiomes. mSphere 2019, 4, e00135-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dibner, J.J.; Richards, J.D. Antibiotic growth promoters in agriculture: History and mode of action. Poult. Sci. 2005, 84, 634–643. [Google Scholar] [CrossRef] [PubMed]
- CIPARS. The Canadian Integrated Program for Antimicrobial Resistance Surveillance (CIPARS) 2015 Annual Report; Public Health Agency of Canada: Ottawa, ON, Canada, 2015. Available online: https://www.canada.ca/en/public-health/services/surveillance/canadian-integrated-program-antimicrobial-resistance-surveillance-cipars.html (accessed on 2 September 2019).
- PHAC. Responsible use of Medically Important Antimicrobials in Animals. 2017. Available online: https://www.canada.ca/en/public-health/services/antibiotic-antimicrobial-resistance/animals/actions/responsible-use-antimicrobials.html (accessed on 2 September 2019).
- Cromwell, G.L. Why and how antibiotics are used in swine production. Anim. Biotechnol. 2002, 13, 7–27. [Google Scholar] [CrossRef]
- CIPARS-b. Canadian Integrated Program for Antimicrobial Resistance Surveillance (CIPARS) Annual Report; Public Health Agency of Canada: Ottawa, ON, Canada, 2016. Available online: https://www.canada.ca/en/public-health/services/surveillance/canadian-integrated-program-antimicrobial-resistance-surveillance-cipars/cipars-reports/2016-annual-report-summary.html (accessed on 2 September 2019).
- Chekabab, S.M.; Lawrence, J.R.; Alvarado, A.; Predicala, B.; Korber, D.R. A health metadata-based management approach for comparative analysis of high-throughput genetic sequences for quantifying antimicrobial resistance reduction in Canadian hog barns. Comput. Struct. Biotechnol. J. 2020, 18, 2629–2638. [Google Scholar] [CrossRef]
- StatCan. Hogs Statistics, Number of Hogs on Farms at End of Semi-Annual Period. 2021. Available online: https://www150.statcan.gc.ca/t1/tbl1/en/cv.action?pid=3210016001 (accessed on 1 March 2021).
- Davies, P.R.; Singer, R.S. Antimicrobial use in wean to market pigs in the United States assessed via voluntary sharing of proprietary data. Zoonoses Public Health 2020, 67 (Suppl. 1), 6–21. [Google Scholar] [CrossRef]
- Bergspica, I.; Kaprou, G.; Alexa, E.A.; Prieto, M.; Alvarez-Ordonez, A. Extended Spectrum beta-Lactamase (ESBL) Producing Escherichia coli in Pigs and Pork Meat in the European Union. Antibiotics 2020, 9, 678. [Google Scholar] [CrossRef]
- Hayer, S.S.; Rovira, A.; Olsen, K.; Johnson, T.J.; Vannucci, F.; Rendahl, A.; Perez, A.; Alvarez, J. Prevalence and trend analysis of antimicrobial resistance in clinical Escherichia coli isolates collected from diseased pigs in the USA between 2006 and 2016. Transbound. Emerg. Dis. 2020, 67, 1930–1941. [Google Scholar] [CrossRef]
- Hayer, S.S.; Rovira, A.; Olsen, K.; Johnson, T.J.; Vannucci, F.; Rendahl, A.; Perez, A.; Alvarez, J. Prevalence and time trend analysis of antimicrobial resistance in respiratory bacterial pathogens collected from diseased pigs in USA between 2006–2016. Res. Vet. Sci. 2020, 128, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Pollock, J.; Muwonge, A.; Hutchings, M.R.; Mainda, G.; Bronsvoort, B.M.; Gally, D.L.; Corbishley, A. Resistance to change: AMR gene dynamics on a commercial pig farm with high antimicrobial usage. Sci. Rep. 2020, 10, 1708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takemura, Y.; Kobayashi, H.; Miyachi, H. Cellular and molecular mechanisms of resistance to antifolate drugs: New analogues and approaches to overcome the resistance. Int. J. Hematol. 1997, 66, 459–477. [Google Scholar] [CrossRef]
- Visentin, M.; Zhao, R.; Goldman, I.D. The Antifolates. Hematol. Oncol. Clin. N. Am. 2012, 26, 629–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, A.R.; Paranhos, A.G.O.; de Aquino, S.F.; Silva, S.Q. Distribution of genetic elements associated with antibiotic resistance in treated and untreated animal husbandry waste and wastewater. Environ. Sci. Pollut. Res. Int. 2021, 28, 26380–26403. [Google Scholar] [CrossRef] [PubMed]
- Kunz, A.; Miele, M.; Steinmetz, R.L. Advanced swine manure treatment and utilization in Brazil. Bioresour. Technol. 2009, 100, 5485–5489. [Google Scholar] [CrossRef]
- Oliver, J.P.; Gooch, C.A.; Lansing, S.; Schueler, J.; Hurst, J.J.; Sassoubre, L.; Crossette, E.M.; Aga, D.S. Invited review: Fate of antibiotic residues, antibiotic-resistant bacteria, and antibiotic resistance genes in US dairy manure management systems. J. Dairy Sci. 2020, 103, 1051–1071. [Google Scholar] [CrossRef]
- Pruden, A.; Larsson, D.G.; Amezquita, A.; Collignon, P.; Brandt, K.K.; Graham, D.W.; Lazorchak, J.M.; Suzuki, S.; Silley, P.; Snape, J.R.; et al. Management options for reducing the release of antibiotics and antibiotic resistance genes to the environment. Environ. Health Perspect. 2013, 121, 878–885. [Google Scholar] [CrossRef]
- Assaraf, Y.G. Molecular basis of antifolate resistance. Cancer Metastasis Rev. 2007, 26, 153–181. [Google Scholar] [CrossRef]
- Nikaido, H.; Pages, J.M. Broad-specificity efflux pumps and their role in multidrug resistance of Gram-negative bacteria. FEMS Microbiol. Rev. 2012, 36, 340–363. [Google Scholar] [CrossRef] [Green Version]
- Garofalo, C.; Vignaroli, C.; Zandri, G.; Aquilanti, L.; Bordoni, D.; Osimani, A.; Clementi, F.; Biavasco, F. Direct detection of antibiotic resistance genes in specimens of chicken and pork meat. Int. J. Food Microbiol. 2007, 113, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Gundogan, N.; Citak, S.; Yucel, N.; Devren, A. A note on the incidence and antibiotic resistance of Staphylococcus aureus isolated from meat and chicken samples. Meat Sci. 2005, 69, 807–810. [Google Scholar] [CrossRef]
- Ramchandani, M.; Manges, A.R.; DebRoy, C.; Smith, S.P.; Johnson, J.R.; Riley, L.W. Possible animal origin of human-associated, multidrug-resistant, uropathogenic Escherichia coli. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2005, 40, 251–257. [Google Scholar] [CrossRef] [PubMed]
- CDC-BMBL. Biosafety in Microbiological and Biomedical Laboratories (BMBL), 5th ed.; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2018. Available online: https://www.cdc.gov/labs/BMBL.html (accessed on 24 October 2018).
- Al-Hebshi, N.N.; Baraniya, D.; Chen, T.; Hill, J.; Puri, S.; Tellez, M.; Hasan, N.A.; Colwell, R.R.; Ismail, A. Metagenome sequencing-based strain-level and functional characterization of supragingival microbiome associated with dental caries in children. J. Oral Microbiol. 2019, 11, 1557986. [Google Scholar] [CrossRef] [PubMed]
- Hourigan, S.K.; Subramanian, P.; Hasan, N.A.; Ta, A.; Klein, E.; Chettout, N.; Huddleston, K.; Deopujari, V.; Levy, S.; Baveja, R.; et al. Comparison of Infant Gut and Skin Microbiota, Resistome and Virulome Between Neonatal Intensive Care Unit (NICU) Environments. Front. Microbiol. 2018, 9, 1361. [Google Scholar] [CrossRef]
- Connelly, S.; Bristol, J.A.; Hubert, S.; Subramanian, P.; Hasan, N.A.; Colwell, R.R.; Kaleko, M. SYN-004 (ribaxamase), an oral beta-lactamase, mitigates antibiotic-mediated dysbiosis in a porcine gut microbiome model. J. Appl. Microbiol. 2017, 123, 66–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chekabab, S.M.; Lawrence, J.R.; Alvarado, A.C.; Predicala, B.Z.; Korber, D.R. Piglet Gut and in-Barn Manure from Farms on a Raised without Antibiotics Program Display Reduced Antimicrobial Resistance but an Increased Prevalence of Pathogens. Antibiotics 2021, 10, 1152. https://doi.org/10.3390/antibiotics10101152
Chekabab SM, Lawrence JR, Alvarado AC, Predicala BZ, Korber DR. Piglet Gut and in-Barn Manure from Farms on a Raised without Antibiotics Program Display Reduced Antimicrobial Resistance but an Increased Prevalence of Pathogens. Antibiotics. 2021; 10(10):1152. https://doi.org/10.3390/antibiotics10101152
Chicago/Turabian StyleChekabab, Samuel M., John R. Lawrence, Alvin C. Alvarado, Bernardo Z. Predicala, and Darren R. Korber. 2021. "Piglet Gut and in-Barn Manure from Farms on a Raised without Antibiotics Program Display Reduced Antimicrobial Resistance but an Increased Prevalence of Pathogens" Antibiotics 10, no. 10: 1152. https://doi.org/10.3390/antibiotics10101152