
Ray Traits of Juvenile Wood and Mature Wood: Pinus massonia and Cunninghamia lanceolata
by
Qiulu Meng
1,2,3, 
Feng Fu
2,
Jie Wang
2,3,
Tuo He
2,3,
Xiaomei Jiang
2,3,
Yonggang Zhang
2,3,
Yafang Yin
2,3,
Ning Li
1,* and
Juan Guo
2,3,* 1
School of Resources, Environment and Materials, Guangxi University, No. 100 Daxue East Road, Nanning 530004, China
2
Research Institute of Wood Industry, Chinese Academy of Forestry, No. 1 Dongxiaofu, Haidian District, Beijing 100091, China
3
Wood Collection, Chinese Academy of Forestry, No. 1 Dongxiaofu, Haidian District, Beijing 100091, China
*
Authors to whom correspondence should be addressed.
Forests 2021, 12(9), 1277; https://doi.org/10.3390/f12091277
Submission received: 3 September 2021
/
Revised: 11 September 2021
/
Accepted: 14 September 2021
/
Published: 17 September 2021
(This article belongs to the Special Issue Wood Formation and Environmental Constraints: Multiscale Approach)
Abstract
:Ray traits affect secondary xylem development and wood properties. Pinus massonia and Cunninghamia lanceolata, commercially important timber species, were chosen to study the differences in wood ray traits of juvenile versus mature wood. Seven ray traits, i.e., percentage of rays, ray spacing, ray number, uniseriate ray height, fusiform ray height, ray parenchyma cell length and ray tracheid length, as well as eight wood axial tissue traits, were investigated quantitatively. Intraspecific variations in ray traits and axial tissue traits between juvenile wood and mature wood were displayed in violin plots. The results showed that anatomical differences between juvenile wood and mature wood were significant for both ray traits and axial tissue traits. Juvenile wood generally possessed the larger percentage of rays, higher ray spacing and ray number, smaller ray height and shorter ray cells than mature wood. A positive correlation was present between the ray parenchyma cell length and ray tracheid length. Negative correlations of the ray number and ray spacing with uniseriate ray height were found. Additionally, the axial tracheid cell wall thickness all had Pearson’s correlations with ray spacing, ray number and ray parenchyma cell length.
1. Introduction
Wood is a heterogeneous assemblage of cell walls deposited by the cell cytoplasm during differentiation [1]. The diversity and extent of the cambium determine the wood structure. Because the cambial age has an important effect on the structure of cambial cells and their derivatives [2], xylem is artificially defined by juvenile wood and mature wood, which are produced during the early years of cambial growth and the mature cambium, respectively. In China, domestic commercial timber is mainly from the plantation forest. At present, the average diameter at the breast height of domestic plantation timber is only about 12 cm, and the proportion of juvenile wood reaches to 40.72% [3]. Among them, Pinus massonia (Masson pine) and Cunninghamia lanceolata (Chinese fir) are very important coniferous commercial timbers, whose domestic timber stock exceeds 943 million m3 and their proportion is above 27% [3]. Thus, deeply illustrating differences in the wood anatomical properties between juvenile wood and mature wood for these two species is urgently needed.
Until now, differences in wood properties between juvenile wood and mature wood have mainly been paid attention to regarding the longitudinally organized xylem axial tissues, which are developed from fusiform cambial initials, such as axial tracheids, vessel elements, fibres and axial parenchyma cells. It is reported that, compared with mature wood, juvenile wood has the characteristics of wider growth rings, lower latewood percentage, lower density, shorter axial tracheids (fibers), thinner cell walls, higher microfibril angle, greater longitudinal shrinkage and higher incidence of reaction wood, in general [4,5,6].
However, to date, research studying wood ray traits between juvenile wood and mature wood remains very limited [7,8,9,10]. Differences in wood ray traits between juvenile wood and mature wood is not clear. Wood rays consist of ray parenchyma cell and ray tracheids. Ray tracheids are present normally in some Gymnospermae, i.e., Pinus sp. [11]. Rays produced by ray cambial initials play very important roles in xylem development. They organize radially and function as the radial transport of reserve materials, water and gases to provide the solute demands for cambial cell division and proliferation as well as wood differentiation [12]. Ray parenchyma cells also function as a radial route for nutrient distribution, which connect with axial parenchyma cells to form a three-dimensional storage system. Moreover, the ray traits affect the wood performance in such aspects as shrinkage [13], pulping properties [14], sound velocity [15], stiffness and radial tensile strength [16], because it is perpendicular to stem and the ray restraint theory. Therefore, a deeper understanding of the differences in wood ray traits between juvenile wood and mature wood could contribute to better the cultivation and utilization of forests.
Herein, juvenile wood and mature wood were collected respectively from the fast-grown plantation of Pinus massonia (Pinaceae) and Cunninghamia lanceolata (Cupressaceae). The differences in wood ray traits of juvenile versus mature wood were studied. The wood anatomical traits were investigated quantitatively. Seven ray traits, including the percentage of rays, ray spacing, ray number, uniseriate ray height, fusiform ray height, ray parenchyma cell length and ray tracheid length, were measured, enabling us to understand the intraspecific ray trait variation between juvenile wood and mature wood. Moreover, eight xylem axial tissue traits, such as axial tracheid cell wall thickness, percentage of axial resin canals and axial parenchyma cell length, were also measured for studying their relationships with ray traits by the correlation analysis. Based on the analysis above, this work could offer detailed anatomical insight into the ray traits of juvenile wood and mature wood in conifers as well as the correlation between rays and wood axial tissues. The knowledge presented will help in future to understand juvenile wood and mature wood for better cultivation and utilization of the forest.
2. Materials and Methods
2.1. Materials
A total of 5 Masson pine trees and 4 Chinese fir trees were harvested in Chongzuo City, Guangxi Province (21°59′50″ N, 107°2′2″ E) and Chengde City, Hunan Province (28°41′28″ N, 111°33′61″ E), respectively. Disks with a thickness of 7–10 cm were collected at the breast height of 1.3 m from each tree in 2019 and stored for further measurements. Juvenile wood and mature wood were selected at the annual ring number of 6th and 19th from the pith, on the basis of their threshold cambial ages differentiating juvenile wood and mature wood at about 16 years and 12 years [8] for Masson pine and Chinese fir, respectively.
2.2. Quantitative Wood Anatomy
The 10 × 10 × 10 mm3 wood samples blocks were taken from the 6th and 19th growth rings in each wood disk, respectively. Transverse, radial and tangential wood sections with thicknesses of 18 µm were prepared with a sliding microtome (Leica SM 2010R, Leica Biosystems Inc., Wetzlar, Germany). The wood sections were stained with a mixture of safranin (0.2 w/v%, in 50% ethanol) and Alcian blue (1.0 w/v%, in distilled water) for about 2.5 min, thoroughly washed with distilled water, dehydrated with ethanol series (30%, 50%, 75%, 95%, 100%, 100%) for a few seconds, washed with xylene, and mounted with a drop of Canada balsam.
Quantitative anatomical observations were conducted by the optical microscope (Olympus DP71, Olympus, Tokyo, Japan) and measured, according to the IAWA list [17] by ImageJ software (ImageJ 1.44p, National Institute of Mental Health, Bethesda, MD, USA). Table 1 presents the feature abbreviations and their statistical analysis methods.
2.3. Scanning Electron Microscopy Observation
Scanning electron microscope (SEM) was used to analyze the ray parenchyma cell length of the Masson pine because it was difficult to be measured in the presence of window-like cross-field pitting under the optical microscope. Samples were split radially into small blocks (10 × 5 × 2 mm3), trimmed with the microtome and oven-dried at 60 °C for 24 h. After being coated with gold at 15 mA in an anion sputter coater (Quorum SC7620, East Sussex, England) for 15 min, they were observed by SEM (ZEISS Gemini 300, Oberkochen, Germany) under high vacuum (3 kV accelerate voltage, 8.1 mm working distance, Inlens detector).
2.4. Data Analysis
One-way analysis of variance (ANOVA) was used to compare each wood anatomical feature between juvenile wood and mature wood using SPSS software (SPSS Statistics 17.0, IBM, New York, NY, USA). Violin plot and Pearson’s correlation matrix were performed in GraphPad Prism9.0 (GraphPad Software, San Diego, CA, USA). A violin plot was used to visually compare the distribution of each wood anatomical feature between the juvenile wood and mature wood based on the samples collected for this study. Pearson’s correlation matrix was used to analyze the relationships between the wood ray features and other anatomical features, as well as the relationships amongst wood anatomical features between juvenile wood and mature wood.
3. Results and Discussion
3.1. Percentages of Different Cell Types
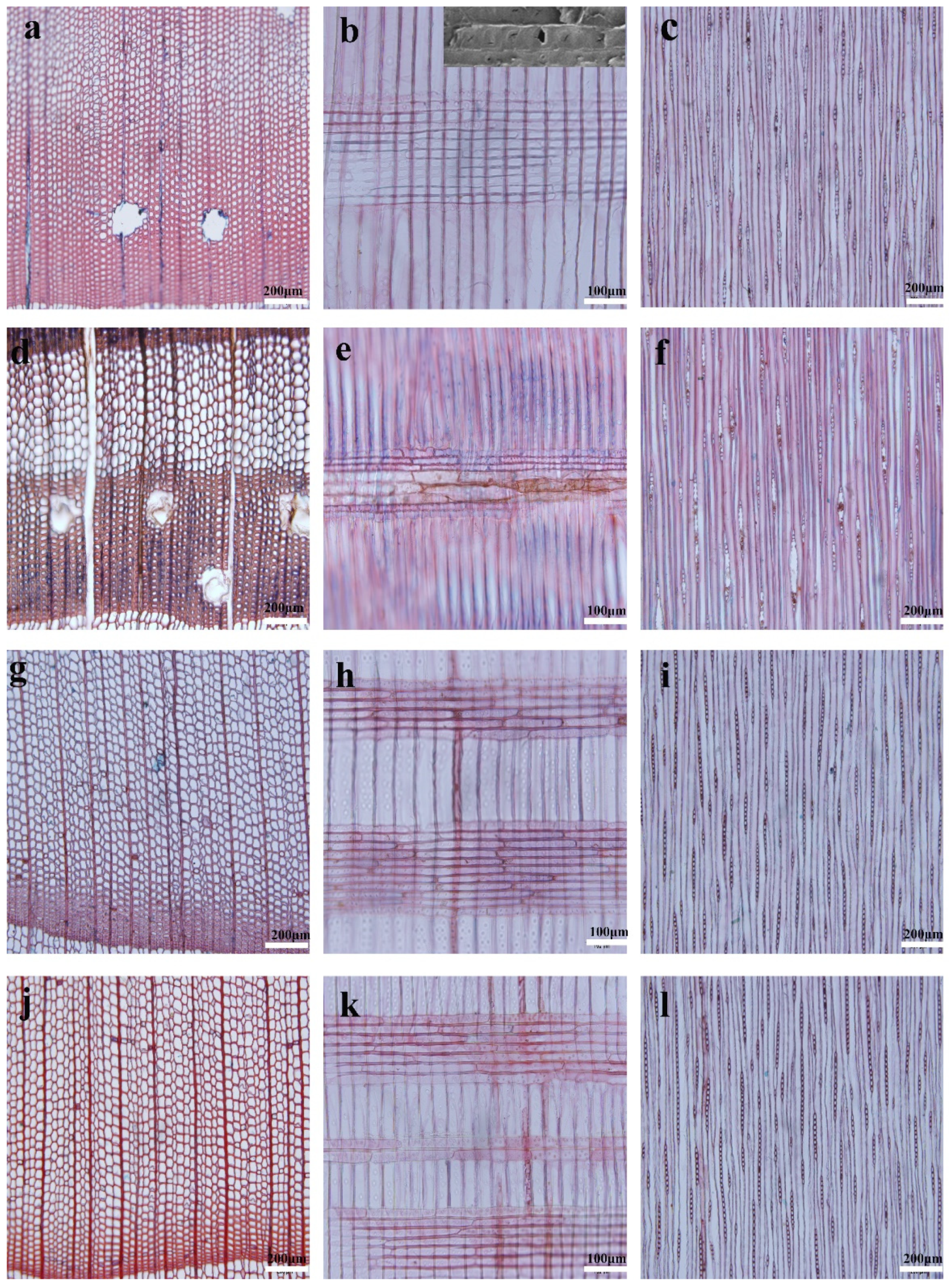
Figure 1 shows the representative microscopic images of juvenile wood and mature wood of Masson pine and Chinese fir. The distribution of resin canals is dependent upon species [27]. Regarding the studied Masson pine, axial resin canals are mainly distributed in the earlywood/latewood transition zone for the juvenile wood (Figure 1a), as reported in Pinus radiata [27], but become more frequent in mature latewood (Figure 1d), similar to Pinus krempeii [28]. Furthermore, juvenile wood has a similar diameter of the axial resin canal as mature wood. Fusiform rays are bi- or triseriate throughout the central portion with a single radial resin canal, and taper above and below to uniseriate margins (Figure 1c,f). The window-like cross-field pitting in the Masson pine makes observing the end walls of the ray parenchyma cells difficult under the optical microscopy (Figure 1b,e), compared to the Chinese fir (Figure 1h,k), which has taxodioid cross-field pittings. Thus, the ray parenchyma cell length was measured by SEM observation instead, as shown in the top right corner of Figure 1b.
Results of the comprehensive wood anatomical features of Masson pine and Chinese fir are listed in Table 2. The average percentages of axial tracheids, rays and axial resin canals of juvenile Masson pine are 95.3%, 3.9% and 0.8%, respectively. Those of mature Masson pine are 93.4%, 3.4% and 3.2%, respectively. The percentage of rays between juvenile Masson pine and mature Masson pine is insignificant, at a 95% confidence level. It is consistent with the little variation among consecutive annual rings for the percentage of rays in Larix gmelinii of Pinaceae [29]. However, the average percentage of axial resin canals significantly increases to 3.2% for mature Masson pine, at a 99% confidence level (Table 1). The percentage of resin canals found in softwood can be influenced by external conditions, e.g., climate, fertilizer application and insect attack [27]. It is reported that the percentage of resin canals in Picea abies is positively and significantly correlated with the summer temperature [30]. The increase in the percentage of axial resin canals of studied Masson pine might be also due to the increased annual average temperature, which was changed from 21.7 °C in 1967 to 24.0 °C in 2015 in Chongzuo City [31]. Consequently, the significant increase in the percentage of axial resin canals from juvenile wood to mature wood apparently caused the decrease in the percentage of axial tracheids for Masson pine.
Regarding the Chinese fir, the average percentage of axial tracheids, percentage of rays and percentage of axial parenchyma are 89.9%, 9.3% and 0.8% for the juvenile wood, and 91.6%, 8.1% and 0.3% for the mature wood, respectively (Table 2). A significant decrease in the average percentage of axial parenchyma was present from juvenile wood to mature wood, at a 99% confidence level. Moreover, the percentage of rays of mature Chinese fir was also smaller than that of juvenile wood, as previously reported for Chinese fir by Bao et al. [32]. Decreases in percentage of rays and percentage of axial parenchyma caused the increased percentage of axial tracheids for mature Chinese fir, consequently.
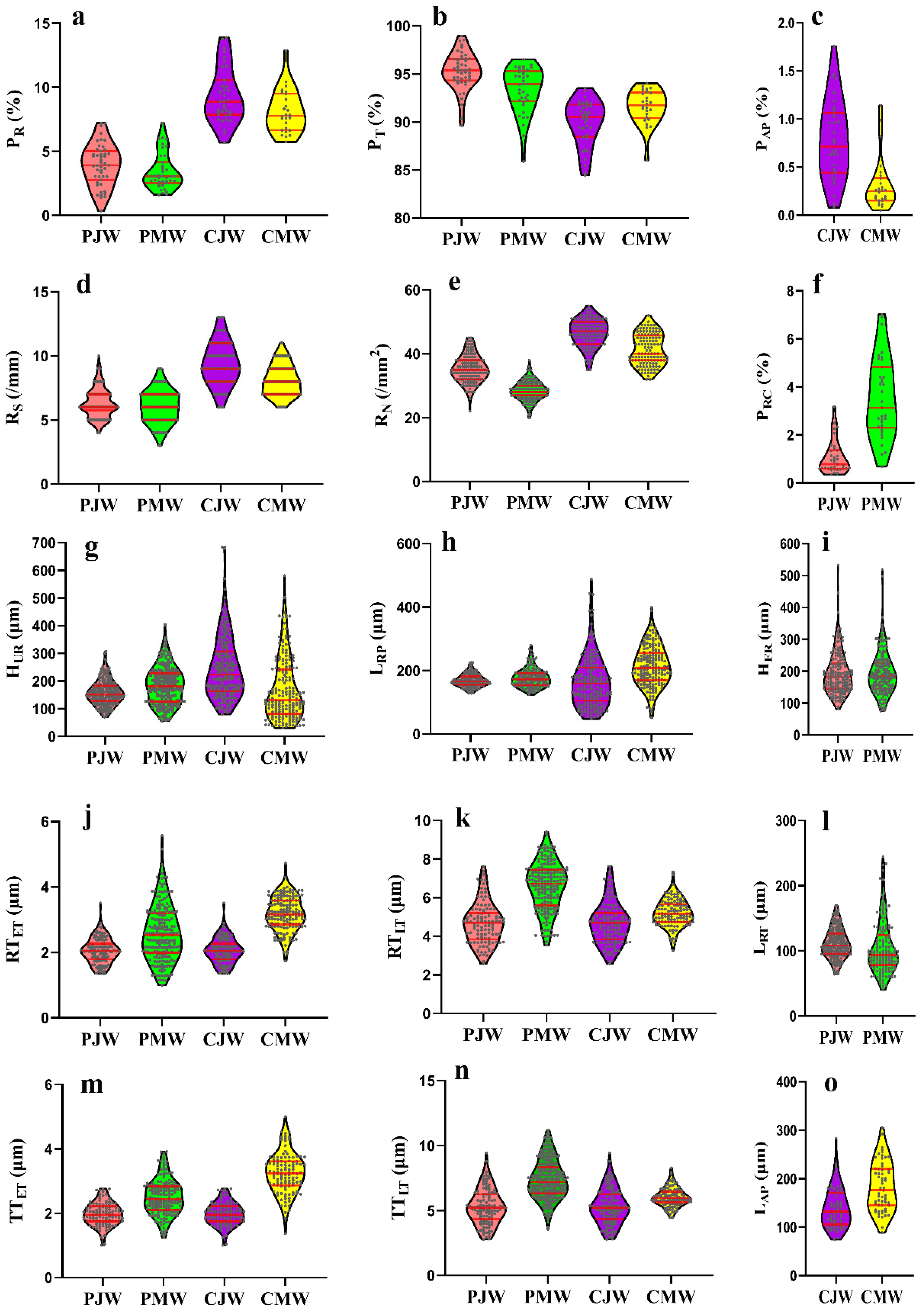
Violin plots were further used to display the overall distribution of each wood anatomical feature (Figure 2). Red lines inside the violin plot show the first quartile, median, and third quartile. Violin plots confirm the variations in percentages of different cell types between juvenile wood and mature wood. It is vividly shown that Masson pine has significant differences in the percentage of axial tracheids and axial resin canals, but insignificant difference in the percentage of rays between juvenile wood and mature wood (Figure 2a,c,f). However, there are significant variations in the percentage of axial tracheids, percentage of rays and percentage of axial parenchyma between juvenile wood and mature wood in Chinese fir.
3.2. Ray Traits
Six ray traits, including ray spacing, ray number, uniseriate ray height, fusiform ray height, ray parenchyma cell length and ray tracheid length, were measured. Their differences between juvenile wood and mature wood were investigated by the assistance of ANOVA.
Average ray spacing values of juvenile wood and mature wood were 6.3/mm and 6.0/mm in Masson pine, and 9.4/mm and 8.1/mm in Chinese fir, respectively (Table 2). It is consistent with the average ray spacing of coniferous wood, ranging 6 to 9/mm [33]. As shown in Figure 2d, the violin plot of ray spacing presents a fluctuation around the 6/mm for Masson pine. An obvious decrease is shown in Chinese fir from juvenile wood to mature wood, in accordance with the decrease in the ray spacing of Larix gmelinii and Larix kaempferi as the growth ring number increased [10]. It should be emphasized that, according to the hypothesis proposed by Carmi et al. [34] and Lev-Yadun and Aloni [12], ray spacing is controlled by excess stimuli, such as the hormone ethylene, which has a negative effect on polar auxin transport, and in turn, affects the formation of ray initials and the enlargement of existing ones. Therefore, different radial variations of the ray spacing within the stems have been observed in the published literature [10,21].
The average ray number of juvenile wood and mature wood was 35.3/mm2 and 28.4/mm2 for Masson pine, 46.5/mm2 and 41.3/mm2 for Chinese fir, respectively. Masson pine had the similar average ray number value as Pinus koraiensis, Larix gmelinii and Larix kaempferi [10]. Chinese fir had a much higher average ray number value than Masson pine. For both species, the average ray numbers of juvenile wood were significantly higher than those of mature wood (p < 0.01, Table 2). This is consistent with the previous studies; the ray number in coniferous wood decreased with increasing the number of the growth ring and became a constant in mature wood [10,35,36]. The violin plot confirmed that juvenile wood of both species has higher first, median, and third quartile values than their corresponding mature wood (Figure 2e) because the largest numbers of the ray cambial initials are formed during the early juvenile years when peripheral expansion of the cambium is greatest [21].
Most rays increase in height either by transverse division of ray cambial initials, by the merger of adjacent rays, or by the addition of segments from declining fusiform initials [21]. In gymnosperms, uniseriate rays commonly increase in height by simple, transverse anticlinal divisions of the terminal cells. As a general rule, ray heights increase with the age of the tree. Accordingly, the studied Masson pine presented an obvious increase in average uniseriate ray height from 157.3 μm of juvenile wood to 183.3 μm of mature wood (p < 0.05, Table 2), similar to the radial variation of ray heights of Pinus halepensis and Pinus pinea [37]. However, a decline in the average uniseriate ray height from juvenile wood to mature wood was present in Chinese fir. This is probably the result of the influence of species differences and growth conditions [21]. The decline in ray height could be due to the elimination of ray initials in the cambium, as reported in Taxus baccata [38]. The elimination of ray initials further led to a concomitant decrease in the percentage of rays. Thus, the studied mature Chinese fir had a shorter uniseriate ray height and lower percentage of rays than juvenile wood.
Fusiform ray, a special wood anatomical feature of Pinaceae, displayed an insignificant difference between juvenile wood and mature wood (Table 2). The fusiform ray height appeared to be less affected by variations between juvenile wood and mature wood than uniseriate rays (Figure 2i), as previously reported in some Pinus sp., such as Pinus ponderosa, Pinus contorta, Pinus resinosa and Pinus strobus [39]. Different variations in uniseriate ray height and fusiform ray height have been also reported previously [10].
Significant differences between juvenile wood and mature wood were also observed in the ray parenchyma cell length and ray tracheid length for both species (p < 0.05, Table 2). In most of the arborescent species with cylindrical cambium, the length of the ray cells usually corresponded to the radial dimension of fusiform-cell pockets, which were formed via percilinal divisions of derivatives of fusiform initials [40]. The ray length could be thus affected by the cambial age, causing a varied length between juvenile wood and mature wood. In agreement, the average ray parenchyma cell length of mature wood was significantly longer than that of juvenile wood for both species (Table 2). Moreover, the ray parenchyma cell length of the studied conifer was similar to that of Pinus sp., at 167 μm to 214 μm [41]. Regarding the ray tracheid, it was 111.3 μm and 104.6 μm for juvenile Masson pine and mature Masson pine, respectively. This is similar to Pinus rigida and Pinus kaempferi [41]. Furthermore, the ray tracheids were shorter than ray parenchyma cells in Masson pine, as reported previously [42].
In general, both Masson pine and Chinese fir possessed a larger percentage of rays, higher ray spacing and ray number, smaller ray heights as well as the shorter ray cells in juvenile wood, compared with mature wood. This is probably resulted from the necessity of ray parenchyma cells for transferring statistic and dynamic loads, and transporting water and nutrients during initial wood formation, especially for juvenile wood [43].
3.3. Wood Axial Tissue Traits
Wood axial tissue traits, such as axial tracheid cell wall thickness and axial parenchyma cell length, were studied. Their variations between juvenile wood and mature wood for both species were also discussed (Figure 2g,k,m–o). As shown in Table 2, between juvenile wood and mature wood, significant differences were present in both the radial wall thickness and tangential wall thickness of earlywood axial tracheid and latewood axial tracheid for both species (Table 2). The axial tracheid cell wall thickness of mature wood was always thicker than that of juvenile wood in both Masson pine and Chinese fir (p < 0.01), as reported previously [32,44]. This confirmed that the studied wood samples were characteristic of juvenile wood and mature wood. Moreover, the axial tracheids of Masson pine were always thicker than those of Chinese fir.
Axial parenchyma cells were also studied (Figure 2o). Generally, in coniferous wood, axial parenchyma cells are sparse or totally absent [45]. However, scattered axial parenchyma cells were observed in transverse sections of Chinese fir (Figure 1g,j). The average axial parenchyma cell length of juvenile Chinese fir and mature Chinese fir was 138.9 μm and 183.5 μm, respectively (p < 0.01, Table 2). The average axial parenchyma cell length of mature wood was longer than the juvenile wood (Figure 2o).
The results of the wood anatomical traits showed that the obvious intraspecific variation about ray traits and wood axial tissue traits are present between juvenile wood and mature wood for Masson pine and Chinese fir. Ray initials grow in the radial direction symplastically, in consonance with the rate of radial growth of neighboring fusiform cells, and also divide percilinally with a frequency similar to that of fusiform initials [46]. The cambium initials growth, dividing the fusiform initials growth and ray initials growth, is affected by the cambium age. Thus, there are concerned variations in wood anatomical features between juvenile wood and mature wood.
3.4. Relationship of Wood Anatomical Traits in Relation to Ray Traits
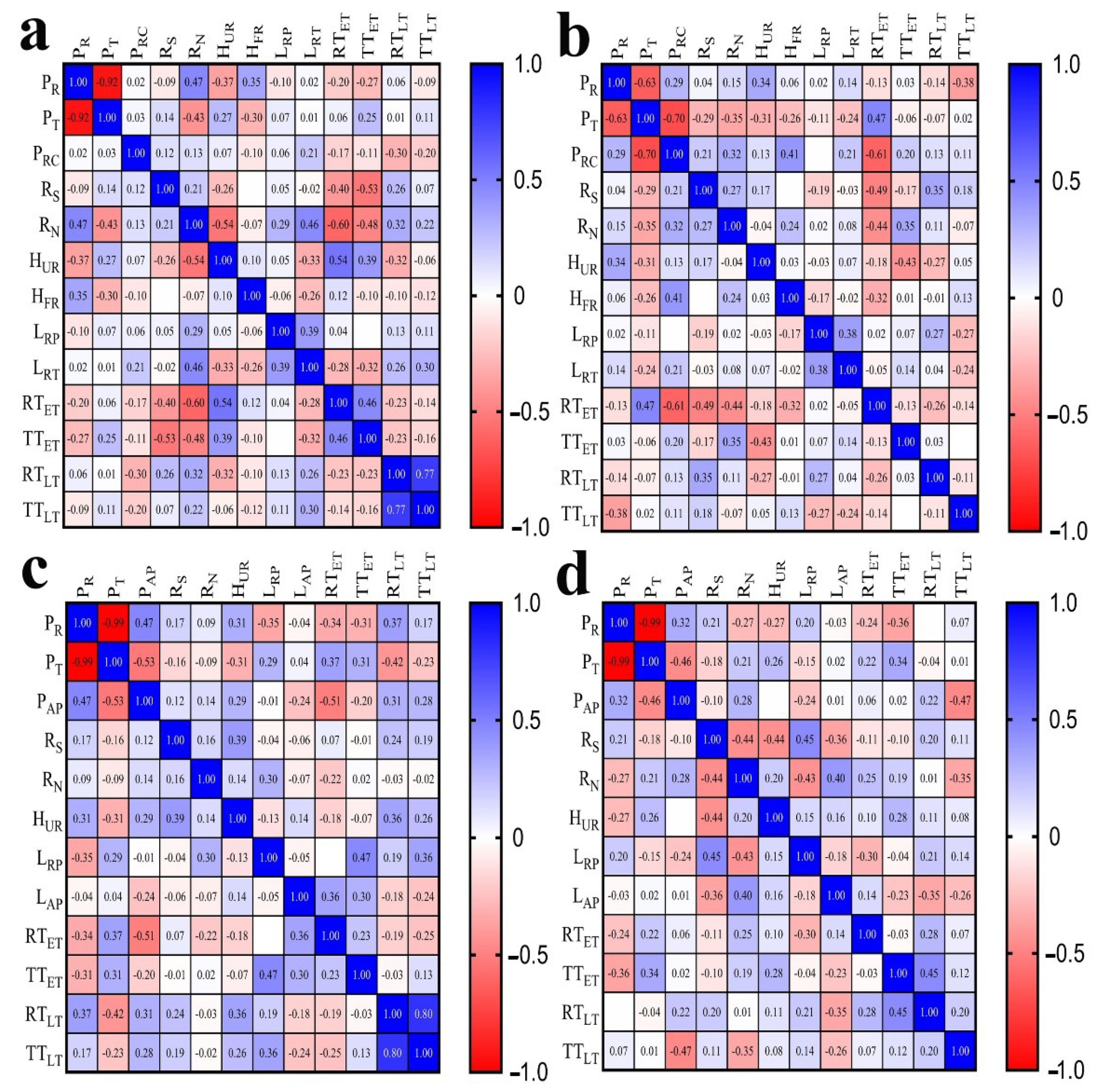
A correlation analysis was conducted to study the relationship of wood anatomical traits with ray traits. The Pearson correlation coefficient (r) was used to quantify the relationship. The r values below 0.30 were not involved in the discussion, because the low Pearson correlation coefficient generally meant a statistically insignificant correlation. Shown in the following, the relationships amongst ray traits and the relationships between ray traits and wood axial tissue traits were studied separately.
Figure 3 shows the Pearson correlation coefficients for wood anatomical traits. Strong negative correlations (r < −0.60) were found between percentage of rays and percentage of axial tracheids for both juvenile wood and mature wood in Masson pine and Chinese fir. Furthermore, the percentage of axial tracheids also had a negative correlation with the percentage of axial resin canals (r = −0.7) towards mature Masson pine. The percentage of rays and percentage of axial parenchyma had positive correlations for juvenile wood and mature wood in Chinese fir (r = 0.47; r = 0.32). This positive relationship was probably because ray and axial parenchyma interconnect commonly and contribute together as the conjunctive tissue in the majority of woods for the acquisition of physiological functions [47]. The percentage of axial tracheids and percentage of axial parenchyma represented negative correlations (r = −0.53; r = −0.46) in both juvenile wood and mature wood in Chinese fir.
The relationships amongst ray traits were also present. Positive correlations between ray parenchyma cell length and ray tracheid length were revealed in both juvenile wood and mature wood of Masson pine (r = 0.39; r = 0.38). As found by Wodzicki and Brown [46], two distinct types of ray initials were produced in Pinaceae; (1) marginal ray initials with their radial diameter equal to, or smaller than, their axial length, and (2) radially elongated initials of the ray interior. Ray parenchyma cells were produced by ray initials in the second group, whereas the ray tracheid may be produced from ray initials in the first group, commonly, and also from ray initials in the second group. Either type could arise from the same initial but proceed in different secondary differentiation processes. Therefore, there is a concerned correlation between ray parenchyma cell length and ray tracheid length. In addition, the negative correlation of ray number and ray spacing with uniseriate ray height revealed in Pinus halepensis and Pinus pinea [37], Larix gmelinii and Larix kaempferi [48] were also present in Masson pine (r = −0.54, Figure 3a) and Chinese fir (r = −0.44, Figure 3d).
Moreover, correlations of axial tracheid cell wall thickness in relation to ray traits were present in the studied Masson pine and Chinese fir. The cell wall thickness of earlywood axial tracheids was negatively correlated with ray spacing (r = −0.4, r = −0.53) for juvenile Masson pine, Figure 3a; r = −0.49 for mature Masson pine, (Figure 3b), negatively correlated with ray number (r = −0.60, r = −0.48 for juvenile Masson pine, Figure 3a; r = −0.44 for mature Masson pine, Figure 3b), correlated with uniseriate ray height (r = 0.54, r = 0.39 for juvenile Masson pine, Figure 3a; (r = −0.43 for mature Masson pine, Figure 3b) and correlated with the ray parenchyma cell length (r = 0.47, r = 0.36 for juvenile Chinese fir, Figure 3c; r = −0.30 for mature Chinese fir, Figure 3d). A weak correlation was previously revealed for cell wall thickness between fibers and rays (r = 0.294, p < 0.05) in 50 Japanese hardwoods [49]. Rays and wood axial tissues, such as axial tracheids, form a complex and highly interconnected functional network throughout the xylem [50]. Ray cells host critical metabolic processes during wood formation [51], produce monolignols that contribute to lignification of axial tracheid cell walls [52], function in storage and transport of nonstructural carbohydrates (NSCs) to develop wood axial tissues [53], cause the thickening of axial tracheid walls [54,55], and need to fit the axial tracheid size to provide functional integration [29]. Thus, concerned relationships between axial tracheid cell wall thickness and ray traits for studied two conifers confirmed that rays could influence the development of wood axial tissue.
4. Conclusions
The ray traits of Conifer Masson pine and Chinese fir were investigated. ANOVA analysis results indicated that anatomical differences between juvenile wood and mature wood were present in seven ray traits and eight wood axial tissue traits for Masson pine and Chinese fir. In general, juvenile wood had the larger percentage of rays, higher ray spacing and ray number, smaller ray heights as well as shorter ray cells than mature wood. Masson pine had the higher percentage of rays, taller uniseriate rays, longer ray parenchyma and thicker axial tracheids than Chinese fir. Furthermore, the ray traits had Pearson’s correlations with the xylem axial tissue traits. Pearson’s correlation results indicated that the rays may influence the formation of axial tissue. Generally, it was shown that the percentage of rays was negatively correlated with percentage of axial tracheids. Ray spacing, ray number and ray parenchyma cell length all had Pearson’s correlations with the axial tracheid cell wall thickness. A positive correlation was present between the ray parenchyma cell length and ray tracheid length. Negative correlations of ray number and ray spacing with uniseriate ray height were also found. Our results suggest that, between the juvenile wood and mature wood, the rays present significant anatomical differences.
Author Contributions
Conceptualization, Q.M.; methodology, J.G.; software, Q.M.; validation, J.G., Q.M.; formal analysis, Q.M.; investigation, Q.M. and N.L; resources, J.W. and F.F.; data curation, J.G. and N.L.; writing—original draft preparation, Q.M.; writing—review and editing, F.F., T.H., Y.Y., N.L., X.J. and J.G.; funding acquisition, F.F. and J.G. field curation. Y.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Nature Science Foundation of China (No. 31890772), and the department of Science and Technology, National Forestry and Grassland Administration (2020132601).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Butterfield, B.G.; Meylan, B.A. Three-Dimensional Structure of Wood. An Ultrastructural Approach. British Library Cataloguing in Publication Data; British Library: New York, NY, USA, 1980; p. 84. [Google Scholar]
- Fromm, J. Plant Cell Monographs; Springer: New York, NY, USA, 2013; pp. 3–39. [Google Scholar]
- Zhang, J.Q.; Li, S.M.; Xu, J.D. China Forest Resources Report; China Forestry Publishing House: Beijing, China, 1992; pp. 15, 28–29. [Google Scholar]
- Yin, Y.; Bian, M.; Song, K.; Xiao, F.; Jiang, X. Influence of microfibril angle on within-tree variations in the mechanical properties of Chinese fir (Cunninghamia lanceolata). IAWA J. 2011, 32, 431–442. [Google Scholar] [CrossRef]
- Moore, J.R.; Cown, D.J. Corewood (juvenile wood) and its impact on wood utilization. Curr. For. Rep. 2017, 3, 107–118. [Google Scholar]
- Luo, X.Q.; Jiang, X.M.; Yin, Y.F.; Li, D.Q.; Wang, B.G. Variations in wood properties of Masson Pine (Pinus massoniana L.) plantation. For. Res. 2002, 1, 28–33. [Google Scholar]
- Davies, F.T., Jr.; Lazarte, J.E.; Joiner, J.N. Initiation and development of roots in juvenile and mature leaf bud cuttings of Ficus pumila L. Botany 1982, 69, 804–811. [Google Scholar] [CrossRef]
- Bao, F.C.; Jiang, Z.H. Wood Properties of Main Tree Species from Plantation in China; China forestry publishing house: Beijing, China, 1998; pp. 5–36. [Google Scholar]
- Song, K.L. Changes of Structure and Performance of Cell Walls during the Transformation from Sapwood to Heartwood of Cunninghamia lanceolata. Master’s Thesis, Chinese Academy of Forestry, Beijing, China, 2012. [Google Scholar]
- Kim, D.H.; Kim, S.H.; Jo, J.I.; Kim, J.H.; Purusatama, B.D.; Lee, S.H.; Kim, N.H. Ray properties in the stems of Dahurian larch (Larix gmelinii) and Japanese larch (Larix kaempferi). IAWA J. 2020, 42, 1–9. [Google Scholar] [CrossRef]
- Holden, R. Ray tracheids in the Coniferales. Int. J. Plant Sci. 1913, 55, 56–65. [Google Scholar] [CrossRef]
- Lev-Yadun, S.; Aloni, R. Differentiation of the ray system in woody plants. Bot. Rev. 1995, 61, 45–84. [Google Scholar] [CrossRef]
- Xue, Q.; Sun, W.; Fagerstedt, K.; Guo, X.; Dong, M.; Wang, W.; Cao, H. Effects of wood rays on the shrinkage of wood during the drying process. Bio-Resources 2018, 13, 7086–7095. [Google Scholar]
- Ona, T.; Sonoda, T.; Ito, K.; Shibata, M.; Tamai, Y.; Kojima, Y.; Ohshima, J.; Yokota, S.; Yoshizawa, N. Investigation of relationships between cell and pulp properties in Eucalyptus by examination of within-tree property variations. Wood Sci. Technol. 2001, 35, 229–243. [Google Scholar] [CrossRef]
- Baar, J.; Tippner, J.; Gryc, V. The relation of fiber length and ray dimensions to sound propagation velocity in wood of selected tropical hardwoods. IAWA J. 2013, 34, 49–60. [Google Scholar] [CrossRef]
- Dória, L.C.; Podadera, D.S.; Lima, R.S.; Lens, F.; Marcati, C.R. Axial sampling height outperforms site as predictor of wood trait variation. IAWA J. 2019, 40, 191–214. [Google Scholar] [CrossRef] [Green Version]
- IAWA Committee. IAWA List of Microscopic Features for Softwood Identification; Brill Publisher: Leiden, The Netherlands, 2004. [Google Scholar]
- Wilkins, A.P.; Papassotiriou, S. Wood anatomical variation of acacia melanoxylon in relation to latitude. IAWA J. 1989, 10, 201–207. [Google Scholar] [CrossRef]
- Pirralho, M.; Flores, D.; Sousa, V.B.; Quilhó, T.; Knapic, S.; Pereira, H. Evaluation on paper making potential of nine Eucalyptus species based on wood anatomical features. Ind. Crops Prod. 2014, 54, 327–334. [Google Scholar] [CrossRef]
- Martín, J.A.; Esteban, L.G.; Palacios, P.; Fernández, F.G. Variation in wood anatomical traits of Pinus sylvestris L. between Spanish regions of provenance. Trees 2010, 24, 1017–1028. [Google Scholar] [CrossRef]
- Larson, P.R. The Vascular Cambium Development and Structure; Springer: New York, NY, USA, 1994; pp. 363–451. [Google Scholar]
- Gregory, R.A.; Romberger, J.A. Cambium activity and height of uniseriate vascular rays in conifers. Int. J. Plant Sci. 1975, 136, 246–253. [Google Scholar]
- Yang, K.C.; Chen, Y.S.; Benson, C.A. Vertical and radial variation of nuclear elongation index of using sapwood ray parenchyma cells in a plantation tree of Cryptomeria Japonica. IAWA J. 1994, 15, 323–327. [Google Scholar] [CrossRef]
- Thompson, W.P. Ray tracheids in abies. Int. J. Plant Sci. 1912, 53, 331–338. [Google Scholar] [CrossRef]
- Thayn, G.F.; Tidwell, W.D. Flora of the lower cretaceous cedar Mountain Formation of Utah and Colorado, Part II, Mesembrioxylon stokesi. Great Basin Nat. 1984, 44, 257–262. [Google Scholar]
- Guo, J.; Guo, X.; Xiao, F.; Xiong, C.; Yin, Y. Influences of provenance and rotation age on heartwood ratio, stem diameter and radial variation in tracheid dimension of Cunninghamia lanceolata. Eur. J. Wood Prod. 2018, 76, 669–677. [Google Scholar] [CrossRef]
- Cown, D.J.; Donaldson, L.A.; Downes, G.M. A review of resin features in radiata pine. N. Z. J. For. Sci. 2011, 41, 41–61. [Google Scholar]
- Ickert-Bond, S.M. Reexamination of wood anatomical features in Pinus krempfii (Pinaceae). IAWA J. 2001, 22, 355–365. [Google Scholar] [CrossRef] [Green Version]
- Fonti, P.; Tabakova, M.A.; Kirdyanov, A.V.; Bryukhanova, M.V.; von Arx, G. Variability of ray anatomy of Larix gmelinii along a forest productivity gradient in Siberia. Trees 2015, 29, 1165–1175. [Google Scholar] [CrossRef]
- Wimmer, R.; Grabner, M. A comparison of tree-ring features in Picea Abies as correlated with climate. IAWA J. 2020, 21, 403–416. [Google Scholar] [CrossRef]
- Mao, J.S.; Huang, X.T.; Yan, L. Temperature change characteristics of Chongzuo City from 1957 to 2017. Mod. Agric. Sci. Technol. 2019, 9, 93–197. [Google Scholar]
- Bao, F.C.; Jiang, Z.H.; Jiang, X.M.; Lu, X.X.; Luo, X.Q.; Zhang, S.Y. Differences in wood properties between juvenile wood and mature wood in 10 species grown in China. Wood Sci. Technol. 2001, 35, 363–375. [Google Scholar] [CrossRef]
- Panshin, A.J.; Zeeuw, C. Textbook of Wood Technology, 4th ed.; McGraw-Hill: New York, NY, USA, 1980. [Google Scholar]
- Carmi, A.; Sachs, T.; Fahn, A. The relation of ray spacing to cambial growth. New Phytol. 1972, 71, 349–353. [Google Scholar] [CrossRef]
- Kim, N.H.; Kwon, S.M.; Chun, K.W. Radial variation of rays in two commercial softwoods grown in Korea. Wood Fiber Sci. 2009, 41, 138–144. [Google Scholar]
- Kim, D.W.; Hwang, S.W.; Lee, W.H. Property of Uljin-Geumgangsong wood (Pinus densiflora forma erecta Uyeki): Appearance pattern of resin canal and ray. J. Korean Wood Sci. Technol. 2014, 42, 78–87. [Google Scholar] [CrossRef]
- Lev-Yadun, S. The relationship between growth-ring width and ray density and ray height in cell number in the earlywood of Pinus Halepensis and Pinus Pinea. IAWA J. 1998, 19, 131–139. [Google Scholar] [CrossRef]
- Klinken, F. Über das gleitende wachstum der initialen im kambium der koniferen und den markstrahlenverlauf in ihrer sekundären rinde. Bibl. Bot. 1914, 19, 1–37. [Google Scholar]
- Bannan, M.W. Ray contacts and rate of anticlinal division in fusiform cambial cells of some Pinaceae. Can. J. Bot. 1965, 43, 487–508. [Google Scholar] [CrossRef]
- Wilczek, A.; Jura-Morawiec, J.; Kojs, P.; Iqbal, M.; Włoch, W. Correlation of intrusive growth of cambial initials to rearrangement of rays in the vascular cambium. IAWA J. 2011, 32, 313–331. [Google Scholar] [CrossRef]
- Ahmed, S.A.; Chong, S.H.; Chun, S.K.; Park, B.S. Ray parenchyma and ray tracheid structure of four Korean pine wood species. J. Korea Furnit. Soc. 2006, 17, 101–107. [Google Scholar]
- Nakaba, S.; Kubo, T.; Funada, R. Differences in patterns of cell death between ray parenchyma cells and ray tracheids in the conifers Pinus densiflora and Pinus rigida. Trees 2008, 22, 623–630. [Google Scholar] [CrossRef]
- Tomczak, A. Relationships between selected tree characteristics and the properties of juvenile wood in Scots pine (Pinus sylvestris L.). For. Res. Pap. 2014, 75, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, M.D.; Denne, M.P. Variation in density of Picea sitchensis in relation to within-tree trends in tracheid diameter and wall thickness. Forestry 1997, 70, 47–60. [Google Scholar] [CrossRef] [Green Version]
- Gričar, J.; Eler, K. The frequency of ray and axial parenchyma versus tree ring width in silver fir (Abies alba Mill.). Trees 2015, 29, 1023–1027. [Google Scholar] [CrossRef]
- Wodzicki, T.J.; Brown, C.L. Cellular differentiation of the cambium in the Pinaceae. Bot. Gaz. 1973, 134, 139–146. [Google Scholar] [CrossRef]
- Calquist, S. Living cells in wood. 1. Absence, scarcity and histology of axial parenchyma as keys to function. Bot. J. Linn. Soc. 2015, 177, 291–321. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.Y. Studies on the wood rays in coniferous woods mutual relation on the indicators of wood rays in coniferous woods. J. Korean Wood Sci. Technol. 1987, 15, 90–98. [Google Scholar]
- Fujiwara, S. Anatomy and properties of Japanese hardwoods II. Variation of dimensions of ray cells and their relation to basic density. IAWA J. 1992, 13, 397–402. [Google Scholar] [CrossRef]
- Spicer, R. Symplasmic networks in secondary vascular tissues: Parenchyma distribution and activity supporting long-distance transport. J. Exp. Bot. 2014, 65, 1829–1848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abreu, I.N.; Johansson, A.I.; Sokołowska, K.; Niittylä, T.; Sundberg, B.; Hvidsten, T.R.; Street, N.R.; Moritz, T. A metabolite roadmap of the wood-forming tissue in Populus tremula. New Phytol. 2020, 228, 1559–1572. [Google Scholar] [CrossRef] [PubMed]
- Blokhina, O.; Laitinen, T.; Hatakeyama, Y.; Delhomme, N.; Paasela, T.; Zhao, L.; Street, N.R.; Wada, H.; Kärkönen, A.; Fagerstedt, K. Ray parenchymal cells contribute to lignification of tracheids in developing xylem of Norway spruce. Plant Physiol. 2019, 181, 1552–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, H.; Plavcova, L.; Cvecko, P.; Fichtler, E.; Gillingham, M.A.F.; Martínez-Cabrera, H.I.; McGlinn, D.J.; Wheeler, E.; Zheng, J.; Zieminska, K.; et al. A global analysis of parenchyma tissue fractions in secondary xylem of seed plants. New Phytol. 2016, 209, 1553–1565. [Google Scholar] [CrossRef] [PubMed]
- Ladell, J.L. Ray thickenings in the walls of Conifer tracheids. Nature 1967, 213, 470–473. [Google Scholar] [CrossRef]
- Keith, C.T. Tangential wall thickenings in Conifer tracheids at ray contact areas. Wood Fiber Sci. 1975, 7, 129–135. [Google Scholar]
Figure 1.
Optical micrographs of the transverse, radial and tangential sections from Masson pine: juvenile wood (a–c) and mature wood (d–f); Chinese fir: juvenile wood (g–i) and mature wood (j–l). A SEM graph for the schematic measurement of ray parenchyma cell length is inserted in the top right corner of Figure 1b.
Figure 1.
Optical micrographs of the transverse, radial and tangential sections from Masson pine: juvenile wood (a–c) and mature wood (d–f); Chinese fir: juvenile wood (g–i) and mature wood (j–l). A SEM graph for the schematic measurement of ray parenchyma cell length is inserted in the top right corner of Figure 1b.

Figure 2.
Violin plots of wood anatomical features in juvenile wood and mature wood of Masson pine and Chinese fir. PJW: juvenile Masson pine, PMW: mature Masson pine, CJW: juvenile Chinese fir, CMW: mature Chinese fir. (a) PR, the percentage of rays, (b) PT, the percentage of axial tracheids, (c) PAP, the percentage of axial parenchyma, (d) RS, the ray spacing, (e) RN, the ray number, (f) PRC, the percentage of axial resin canals, (g) HUR, the uniseriate ray height, (h) LRP, the ray parenchyma cell length, (i) HFR, the fusiform ray height, (j) LRT, the ray tracheid length, (k) LAP, the axial parenchyma cell length, (l) RTET, the radial wall thickness of earlywood axial tracheid, (m) RTLT, the radial wall thickness of latewood axial tracheid, (n) TTET, the tangential wall thickness of earlywood axial tracheid and (o) TTLT, the tangential wall thickness of latewood axial tracheid.
Figure 2.
Violin plots of wood anatomical features in juvenile wood and mature wood of Masson pine and Chinese fir. PJW: juvenile Masson pine, PMW: mature Masson pine, CJW: juvenile Chinese fir, CMW: mature Chinese fir. (a) PR, the percentage of rays, (b) PT, the percentage of axial tracheids, (c) PAP, the percentage of axial parenchyma, (d) RS, the ray spacing, (e) RN, the ray number, (f) PRC, the percentage of axial resin canals, (g) HUR, the uniseriate ray height, (h) LRP, the ray parenchyma cell length, (i) HFR, the fusiform ray height, (j) LRT, the ray tracheid length, (k) LAP, the axial parenchyma cell length, (l) RTET, the radial wall thickness of earlywood axial tracheid, (m) RTLT, the radial wall thickness of latewood axial tracheid, (n) TTET, the tangential wall thickness of earlywood axial tracheid and (o) TTLT, the tangential wall thickness of latewood axial tracheid.

Figure 3.
Correlation coefficients calculated amongst wood anatomical features for Masson pine and Chinese fir. (a) Juvenile Masson pine, (b) mature Masson pine, (c) juvenile Chinese fir, (d) mature Chinese fir.
Figure 3.
Correlation coefficients calculated amongst wood anatomical features for Masson pine and Chinese fir. (a) Juvenile Masson pine, (b) mature Masson pine, (c) juvenile Chinese fir, (d) mature Chinese fir.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Measured wood anatomical features of Masson pine and Chinese fir.
| Wood Anatomical Features | Abbreviation | Unit | N 1 | Measurement 3 |
|---|---|---|---|---|
| Percentage of rays [18] | PR | % | 10 | On ten transverse sections. Rays area as a percentage of measured transverse section area |
| Percentage of axial tracheids [18] | PT | % | 10 | Masson pine: 100% − (percentage of rays + percentage of axial resin canals) Chinese fir: 100% − (percentage of rays + percentage of axial parenchyma) |
| Percentage of axial parenchyma [19] | PAP | % | 10 | On ten transverse sections. Axial parenchyma area as a percentage of measured transverse section area |
| Percentage of axial resin canals [20] | PRC | % | 10 | On ten transverse sections. Axial resin canals area as a percentage of measured transverse section area |
| Ray spacing [21] | RS | /mm | 30 | On ten tangential sections. Count the number of rays in 1 mm |
| Ray number [10] | RN | /mm2 | 30 | On ten tangential sections. Count the number of rays in 1 mm2 |
| Uniseriate ray height [22] | HUR | μm | 50 | On ten tangential sections. Individual uniseriate ray height; tails were included in the measurement |
| Fusiform ray height [10] | HFR | μm | 50 | On the ten tangential sections. Individual fusiform ray height; tails were included in the measurement |
| Ray parenchyma cell length 2 [23] | LRP | μm | 30 | On ten radial sections. Measure individual ray parenchyma cell length |
| Ray tracheid length [24] | LRT | μm | 50 | On ten radial sections. Measure individual ray tracheid length |
| Axial parenchyma cell length [25] | LAP | μm | 30 | On ten radial sections. Measure individual axial parenchyma cell length |
| Radial wall thickness of earlywood axial tracheid [26] | RTET | μm | 30 | On ten transverse sections. The thickness of the single radial cell wall of earlywood axial tracheid on transverse section |
| Tangential wall thickness of earlywood axial tracheid [26] | TTET | μm | 30 | On ten transverse sections. The thickness of the single tangential cell wall of earlywood axial tracheid on transverse section |
| Radial wall thickness of latewood axial tracheid [26] | RTLT | μm | 30 | On ten transverse sections. The thickness of the single radial cell wall of latewood axial tracheid on transverse section |
| Tangential wall thickness of latewood axial tracheid [26] | TTLT | μm | 30 | On ten transverse sections. The thickness of the single tangential cell wall of latewood axial tracheid on transverse section |
Notes: 1: The N presents the measurement repetition of each feature. 2: The ray parenchyma cell length of Masson pine was measured by SEM. 3: For quantitative wood anatomy, 10 transverse, radial and tangential wood slices were prepared from the 6th and 19th growth rings of 5 Masson pine wood disks and Chinese fir wood disks, respectively.
Table 2.
Wood anatomical features measured in Masson pine and Chinese fir as well as ANOVA results of juvenile wood and mature wood, respectively 1.
Table 2.
Wood anatomical features measured in Masson pine and Chinese fir as well as ANOVA results of juvenile wood and mature wood, respectively 1.
| Percentage of Rays (%) | Percentage of Axial Tracheids (%) | Percentage of Axial Parenchyma (%) | Percentage of Axial Resin Canals (%) | Ray Spacing (/mm) | Ray Number (/mm2) | Uniseriate Ray Height (μm) | Fusiform Ray Height (μm) | Ray Parenchyma Cell Length 2 (μm) | Ray Tracheid Length (μm) | Axial Parenchyma Cell Length (μm) | Cell wall Thickness of Earlywood Axial Tracheid | Cell Wall Thickness of Latewood Axial Tracheid | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Radial (μm) | Tangential (μm) | Radial (μm) | Tangential (μm) | |||||||||||||
| Masson pine | Juvenile wood | 3.9 ± 1.6 (0.3–7.2) | 95.3 ± 1.9 (89.6–99.0) | / | 0.8 ± 0.7 (0.3–3.2) | 6.3 ± 1.1 (4–10) | 35.3 ± 4.2 (22–45) | 157.3 ± 43.8 (69.4–306.7) | 192.4 ± 65.7 (81.7–532.5) | 168.5 ± 19.8 (128.2–225.3) | 111.3 ± 22.3 (64.1–169.6) | / | 2.5 ± 0.4 (1.2–4.4) | 2.2 ± 0.3 (1.2–4.2) | 6.8 ± 1.1 (3.7–9.8) | 6.3 ± 1.3 (4.1–8.2) |
| Mature wood | 3.4 ± 1.4 (1.6–7.2) | 93.4 ± 2.3 (85.9–96.5) | / | 3.2 ± 1.6 (0.7–7.0) | 6.0 ± 1.3 (3–9) | 28.4 ± 3.0 (20–38) | 183.3 ± 67.0 (56.1–403.6) | 193.7 ± 67.8 (75.5–518.9) | 176.0 ± 28.6 (124.4–279.7) | 104.6 ± 40.2 (40.2–244.7) | / | 2.6 ± 0.8 (1.0–5.6) | 2.4 ± 0.5 (1.2–3.9) | 7.2 ± 1.2 (5.1–9.4) | 7.4 ± 1.5 (3.5–11.2) | |
| p value 3 | 0.117 | 0.000 ** | / | 0.000 ** | 0.089 | 0.000 ** | 0.000 ** | 0.292 | 0.026 * | 0.034 * | / | 0.002 ** | 0.000 ** | 0.000 ** | 0.000 ** | |
| Chinese fir | Juvenile wood | 9.3 ± 2.1 (5.7–13.9) | 89.9 ± 2.4 (84.5–93.5) | 0.8 ± 0.4 (0.1–1.8) | / | 9.4 ± 1.7 (6–13) | 46.5 ± 4.2 (35–55) | 244.8 ± 109.5 (79.4–684.6) | / | 164.6 ± 79.2 (47.6–488.1) | / | 138.9 ± 40.5 (74.5–283.2) | 2.1 ± 0.4 (1.4–3.5) | 2.0 ± 0.3 (1.0–4.7) | 4.7 ± 1.1 (2.6–7.6) | 5.3 ± 1.3 (2.8–9.4) |
| Mature wood | 8.1 ± 1.6 (5.7–12.9) | 91.6 ± 1.7 (86.0–94.0) | 0.3 ± 0.3 (0.1–1.1) | / | 8.1 ± 1.2 (6–11) | 41.3 ± 4.8 (32–52) | 167.0 ± 109.2 (30.0–581.4) | / | 214.1 ± 61.3 (51.6–399.3) | / | 183.5 ± 49.7 (88.6–304.5) | 3.2 ± 0.5 (1.7–4.3) | 3.2 ± 0.6 (1.4–5.0) | 5.2 ± 0.7 (3.2–7.3) | 6.0 ± 0.7 (4.5–8.3) | |
| p value 3 | 0.016 * | 0.004 ** | 0.000 ** | / | 0.000 ** | 0.000 ** | 0.000 ** | / | 0.000 ** | / | 0.000 ** | 0.000 ** | 0.000 ** | 0.000 ** | 0.000 ** | |
Notes: 1: Numbers in parentheses indicate the range for each of the wood anatomical features. 2: The ray parenchyma cell length of Masson pine was measured by SEM. 3: ** statistically significant at p < 0.01, * statistically significant at p < 0.05.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Meng, Q.; Fu, F.; Wang, J.; He, T.; Jiang, X.; Zhang, Y.; Yin, Y.; Li, N.; Guo, J. Ray Traits of Juvenile Wood and Mature Wood: Pinus massonia and Cunninghamia lanceolata. Forests 2021, 12, 1277. https://doi.org/10.3390/f12091277
AMA Style
Meng Q, Fu F, Wang J, He T, Jiang X, Zhang Y, Yin Y, Li N, Guo J. Ray Traits of Juvenile Wood and Mature Wood: Pinus massonia and Cunninghamia lanceolata. Forests. 2021; 12(9):1277. https://doi.org/10.3390/f12091277
Chicago/Turabian StyleMeng, Qiulu, Feng Fu, Jie Wang, Tuo He, Xiaomei Jiang, Yonggang Zhang, Yafang Yin, Ning Li, and Juan Guo. 2021. "Ray Traits of Juvenile Wood and Mature Wood: Pinus massonia and Cunninghamia lanceolata" Forests 12, no. 9: 1277. https://doi.org/10.3390/f12091277
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.