
Opposing Actions of Octopamine and Tyramine on Honeybee Vision


1
Biocenter, Department of Behavioural Physiology and Sociobiology, Julius-Maximilians-Universität of Würzburg, Am Hubland, 97074 Würzburg, Germany
2
Department of Biological Cybernetics and Theoretical Biology, University of Bielefeld, 25, 33615 Bielefeld, Germany
*
Author to whom correspondence should be addressed.
Biomolecules 2021, 11(9), 1374; https://doi.org/10.3390/biom11091374
Submission received: 21 August 2021
/
Revised: 14 September 2021
/
Accepted: 15 September 2021
/
Published: 17 September 2021
(This article belongs to the Special Issue Insect Receptors: Biochemical, Physiological and Molecular Studies)
Abstract
:The biogenic amines octopamine and tyramine are important neurotransmitters in insects and other protostomes. They play a pivotal role in the sensory responses, learning and memory and social organisation of honeybees. Generally, octopamine and tyramine are believed to fulfil similar roles as their deuterostome counterparts epinephrine and norepinephrine. In some cases opposing functions of both amines have been observed. In this study, we examined the functions of tyramine and octopamine in honeybee responses to light. As a first step, electroretinography was used to analyse the effect of both amines on sensory sensitivity at the photoreceptor level. Here, the maximum receptor response was increased by octopamine and decreased by tyramine. As a second step, phototaxis experiments were performed to quantify the behavioural responses to light following treatment with either amine. Octopamine increased the walking speed towards different light sources while tyramine decreased it. This was independent of locomotor activity. Our results indicate that tyramine and octopamine act as functional opposites in processing responses to light.
Keywords:
biogenic amines; neurotransmitter; phototaxis; Apis mellifera; ERG; behaviour; modulation; visual system; octopamine; tyramine1. Introduction
The biogenic amines octopamine (OA) and tyramine (TA) play decisive roles in the modulation of honeybee behaviour. They can act as neurohormones, neurotransmitters and neuromodulators [1,2,3,4,5,6,7]. Octopamine is present in the honeybee brain in a concentration of up to 900 pg/brain [8,9]. Tyramine is present in the honeybee brain in much smaller quantities, ranging between 50 and 65 pg/brain [10]. Five different OA receptor genes and two TA receptor genes have been characterized in the honeybee. The activation of the respective receptor proteins can turn on very different intracellular second messenger cascades, which differentially affect physiology and behaviour. AmOctαR1 leads to a change in intracellular calcium ([Ca2+]i) when activated [11]. The remaining receptors are coupled with the adenylyl cyclase and lead to changes in intracellular cAMP ([cAMP]i) when activated. AmOctαR2 and AmTAR1 decrease [cAMP]i [12,13], while AmOctβR1/2/3-4 and AmTAR2 increase [cAMP]i [14,15].
Based on the structural similarities of the receptor subtypes and intracellular signalling it is generally assumed that the insect OA/TA system is comparable to the deuterostome epinephrine/norepinephrine system [4,5,7,16,17]. Furthermore, some studies show that OA and TA can have opposite effects, indicating that both amines may act functionally antagonistically [18,19]. Octopamine has frequently been shown to increase responsiveness to different stimulus modalities in honeybees and other insects [1,4,5,6,7,20]. Drosophila melanogaster mutants lacking a functional tyramine β hydroxylase, which mediates the final step in OA synthesis, showed a significantly reduced responsiveness to sucrose due to the lack of OA or an increase in TA titres [21]. Additionally, OA plays further important roles in associative learning and memory [1,4,22] by mediating and modulating the reward in appetitive learning [7,23,24]. In contrast to OA, the role of TA has been studied less intensively. The Drosophila mutant honoka displays a reduced expression of the TAR1 receptor [25]. These mutant flies are slightly hyperactive, have defects in olfactory perception and display a reduced TA-induced muscle contraction. Furthermore, TA was shown to rescue cocaine sensitization defects in Drosophila iav/TRPV channel mutants [26].
In the honeybee, OA and TA have been shown to have functionally similar effects in different taste-related behaviours. Both amines accelerate the rate of habituation of the proboscis extension response (PER) [27] and increase responsiveness to sucrose in honeybees [7,28,29]. They can both improve appetitive learning performance in the PER assay, most likely by increasing responsiveness to the reward [10,30]. Further, both OA and TA are involved in the social organisation of honeybees. Octopamine speeds up the adult behavioural maturation [31] and is associated with the transition from hive work to foraging [32]. Additionally, expression of the OA receptor gene AmOctαR1 is higher in foragers compared to nurse bees [33] and the levels of both biogenic amines are higher in foragers compared to nurse bees as well [9,10]. The TA receptor gene AmTAR1 is located on a quantitative trait locus linked to foraging behaviour [34]. However, how OA and TA affect division of labour in honeybees is unclear. They assumedly modulate sensory responses to sensory stimuli, i.e., gustatory, visual and olfactory cues, as has been shown for the gustatory system.
While some behavioural evidence suggests that OA plays in honeybee vision, little is known about the role of TA in the visual system. Octopamine was shown to enhance the direction-specific antennal responses during presentation of moving stripe patterns [35]. Whether TA affects this behaviour is unknown. Interestingly, in honeybee nectar foragers we showed earlier that TA enhances phototaxis without affecting locomotor behaviour. Octopamine had the opposite effect, i.e., reducing the walking speed towards a light source [20].
To differentiate effects of both amines on the periphery and on the decision-making processes in the central nervous system, we performed comparative studies using electroretinography (ERG) and phototaxis. To investigate whether the substances have a similar effect on honeybees of different ages or behavioural groups, we used young nurse-aged bees and forager bees.
2. Materials and Methods
2.1. Honeybees
Honeybees (Apis mellifera carnica) were reared at the departmental apiary and collected individually in uncoated bottles with snap-on caps. After collection, the honeybees were placed into cages and maintained in an incubator (30 °C—constant darkness) overnight with access to 50% sugar solution ad libitum. Bees were tested for behaviour and/or sensory responses on the following day. Different honeybee groups were chosen for the two experiments. For the ERG experiments, returning non-pollen foragers were caught directly at the hive entrance. For the phototaxis experiments, two groups of honeybees were collected. Foragers were sampled during winter and were therefore collected inside a glasshouse at a feeder containing a 50% sucrose solution. Young honeybees were sampled in spring. Here, we marked newly emerged honeybees and inserted them into an existing colony. They were collected 6 to 14 days later to be used for the experiments, thus representing hive bees. Due to the experimental design and a timespan of 6 months during the experiments, different bee groups had to be used for the different experiments. However, during the same experiment, the two treatment groups were identical.
2.2. Electroretinography
For the ERG, a honeybee was removed from the cage, immobilized on ice and mounted on a small acrylic glass block. After mounting, the honeybee’s mandibles and neck were fixed with low melting dental wax. A tiny window was cut into the head capsule between the antenna base and the ocelli. The window could be folded backwards to apply the substances. The glands and trachea dorsal to the honeybee´s brain were removed. A reference electrode (silver wire, diameter: 25 µm; Nilaco, Tokyo, Japan) was placed into one eye and connected to a common ground, whilst the recording electrode, located inside a glass capillary (1B100F-3, WPI, Sarasota, FL, USA) was pulled with a DMZ-Universal Puller as done before [36] and filled with 0.1 M KCl; it was then inserted into the contralateral eye. The signal was amplified 50x (Neuroprobe Amplifier 1600, A-M Systems, Inc, Sequim, WA, USA) and high-pass filtered by an acquisition board (Labtrax 4/16, WPI, Sarasota, FL, USA) and recorded using LabScribe 2 (iWorx Systems Inc., Dover, NH, USA).
A xenon light source (Perkin Elmer optoelectronics, XL2000 Fiber Optic Illumination System) emitting a daylight spectrum between 350 nm and 800 nm was used in combination with three greyscale intensity filters (36%–, 59%– and 100% light intensity) to stimulate the compound eye with a maximum light intensity of 4.95 × 1016 photons/cm2. To open the light beam we used a manual shutter with a shutter time of 85 ± 6 ms. After placing a honeybee into the ERG setup, it was allowed to rest in constant darkness for 15 min before starting the stimulation with the three intensities in ascending order. Each intensity was applied four times with an inter-trial interval and an inter-stimulus interval of 1 min (Pre-test). Next, the head window was opened and 1 µL of Ringer (270 mM NaCl, 3.2 mM KCl, 1.2 mM CaCl2, 10 mM MgCl2, 10 mM 3-(N-morpholino) propanesulfonic acid, pH 7.4) or 1 µL of either OA or TA in one of three concentrations (10−3 mol/L, 10−4 mol/L or 10−5 mol/L—diluted in Ringer) was applied directly onto the honeybee brain. After another adaptation period of 15 min, the intensities were applied in the same way as in the Pre-test (Post-test). All concentrations for one substance as well as one Ringer control were tested each day using a new honeybee for each substance. The testing order of the substances was distributed randomly. Individual raw ERG data are shown for OA and TA respectively (Figure S1).
2.3. Phototaxis Assay
For the phototaxis experiment, a honeybee was removed from the cage, immobilized on ice and mounted in a small plastic tube. Thereafter, the median ocellus was punctured using a small micro dissecting needle. Afterwards, it was injected with 300 nL of Ringer (see above), tyramine (10−2 mol/L—diluted in Ringer) or octopamine (10−2 mol/L—diluted in Ringer) using a micro manipulator and glass capillaries (1B100F-4, WPI, Sarasota, FL, USA), pulled as described above. Each honeybee was allowed to rest in a Petri dish (diameter = 85 mm) for 15 min in constant darkness. Then it was placed inside the phototaxis arena to measure light responsiveness and walking parameters. The phototaxis assay was analysed as described before [20,36,37]. In short, dark-adapted honeybees were individually placed inside the phototaxis arena and their mean velocity in darkness was recorded for 2 min to measure their locomotor activity. Afterwards, the arena was illuminated with different green light-emitting diodes (LEDs, wavelength = 527 nm) of different light intensities (3%, 6%, 12%, 25%, 50% and 100% light intensity) with a maximum light intensity of 2.61 × 1014 photons/cm2. For technical reasons, we used non-modifiable LEDs in the phototaxis assay and not the same white light source as in the ERG setup. Two LEDs with the same light intensity were placed opposite each other. Once a honeybee reached a light source, the LED was switched off, and the opposite LED with the same light intensity was switched on. This was repeated four times for each light intensity. Each experiment was started by switching on the LED with the lowest light intensity. The other LEDs were turned on in ascending order. The honeybee´s walking time towards each light source was recorded using a computer stopwatch (Comfort Software Group) [37]. Honeybees for one treatment and the corresponding Ringer controls were tested each day in a pseudo random order from 9 am until 4 pm. Honeybees were kept in the cage with ad libitum access to 50% sucrose solution under constant darkness until injection with the treatment or Ringer.
2.4. Data Analysis
For the ERG experiment, data were recorded using LabScribe 2 (iWorx Systems, Inc.). We extracted the maximal response amplitude of the ERG response to compare them before (pre) and after (post) treatment using a one-way ANOVA. Post hoc analyses were conducted using Dunnett’s multiple comparison test. Differences in the ERG responses elicited due to the three light intensity filters were analysed with a repeated measures (RM) one-way ANOVA followed by a Bonferroni´s multiple comparison test. For the phototaxis experiment, the mean velocity during the dark-runs was analysed using a Student’s t-test, since data were distributed normally. The means of one honeybee of four trials for each light intensity were calculated, compared and analysed with a RM 2-way ANOVA using GraphPad PRISM (GraphPad Software Inc., V7, San Diego, CA, USA). Post hoc analyses were conducted using Bonferroni´s multiple comparison test.
3. Results
3.1. The Effect of Octopamine and Tyramine on the ERG Response
Prior to treatment with either amine or the control solution, we investigated whether the three light intensities tested elicited differential receptor responses. In general, the more transparent the filter, the higher the receptor responses independent of the treatment (RM one way ANOVA, p < 0.001; for details see Table 1).
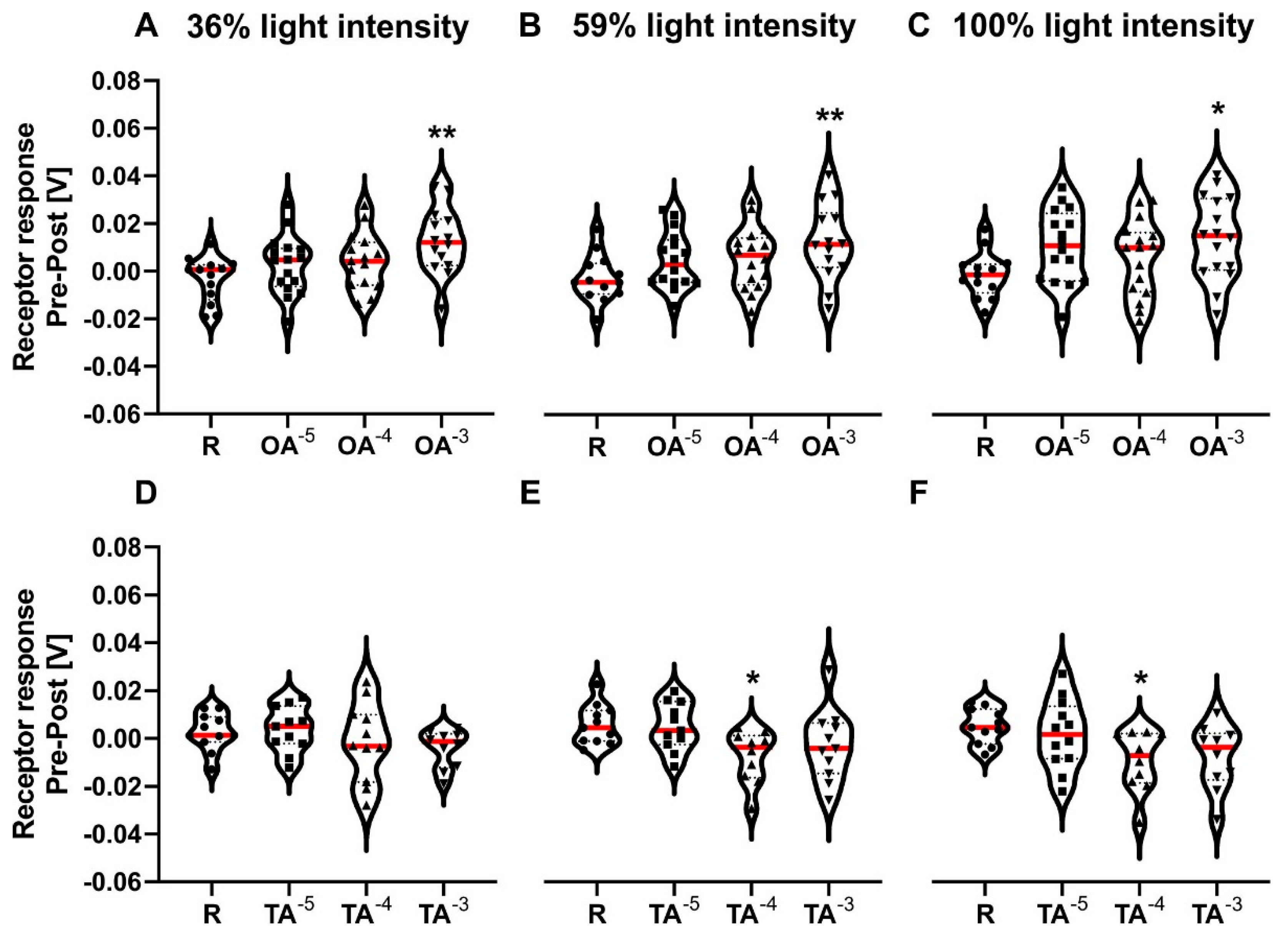
Afterwards, we measured the ERG responses of honeybees following treatment with OA, TA or Ringer. Octopamine had a significant overall effect on the ERG response compared to the Ringer control at all three light intensities (Figure 1A–C; Table 2). It significantly increased the ERG response at all three light intensities for a concentration of 10−3 mol/L (Table 2). Tyramine had a significant overall effect on the pre-post response compared to the control at two out of three light intensities (Figure 1E–F; Table 2). It significantly decreased the ERG response at 59% light intensity and 100% light intensity for a concentration of 10−4 mol/L (Table 2).
3.2. Tyramine and Octopamine Have Opposite Effects on the Phototaxis of In-Hive Bees and Foragers
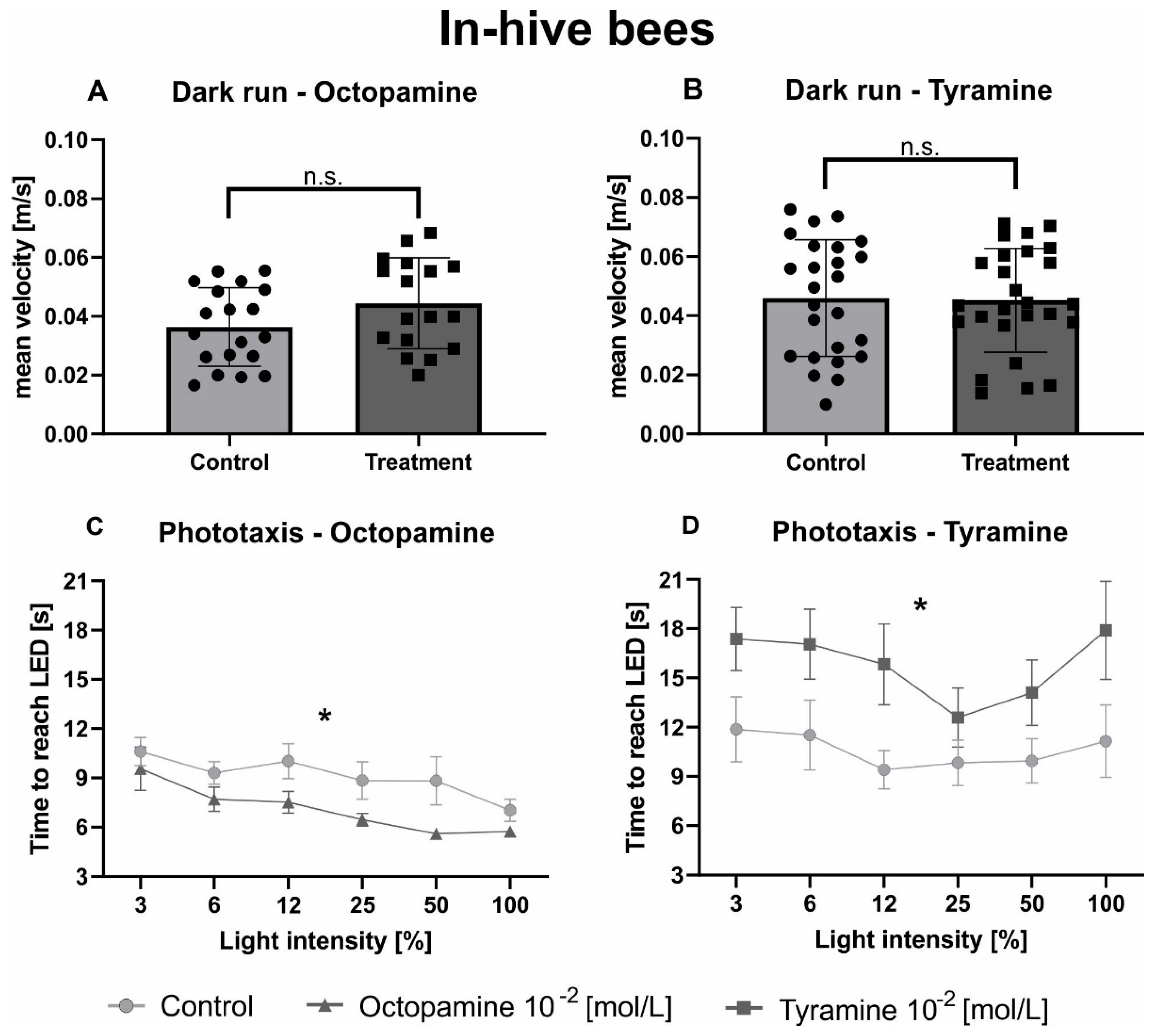
Neither OA (Figure 2A, Table 3) nor TA (Figure 2B, Table 3) affected locomotor behaviour of in-hive bees in the dark arena. Light intensity significantly influenced phototaxis of in-hive bees in both experiments (Figure 2C,D; Table 3). Octopamine significantly decreased the time in-hive bees took to walk towards the switched-on light source compared to the control group (Figure 2C, Table 3), while TA significantly increased the time in-hive bees needed to reach the different switched-on light sources (Figure 2D, Table 3).
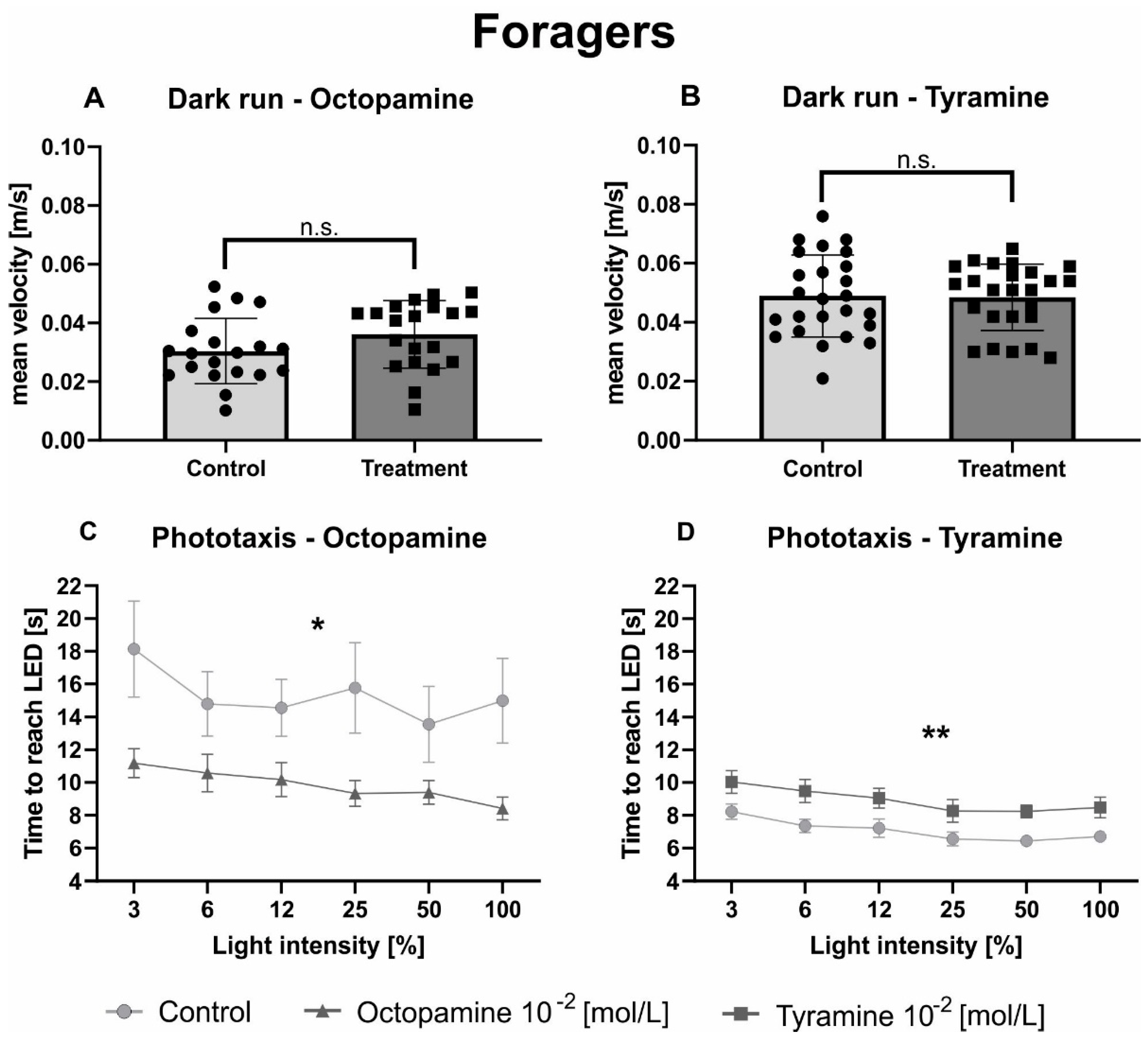
Similar to hive bees, no effect of OA (Figure 3A; Table 3) nor TA (Figure 3B; Table 3) could be observed on the mean velocity of foragers in the dark arena. As expected, the factor light intensity significantly influenced phototactic behaviour of foragers in both experiments (Figure 3C,D; Table 3). Octopamine significantly decreased the walking time of foragers towards the switched-on light sources compared to the control group (Figure 3C, Table 3), while TA significantly increased the time foragers needed to reach the different switched-on light sources (Figure 3D, Table 3).
4. Discussion
In this study we investigated the influence of octopamine and tyramine on honeybee responses to light. One major goal was to separate the effects of both amines at the sensory input level (ERG) and the behavioural output level (phototaxis). To understand whether the stronger attraction to light induced by OA was based on a higher perception of light at the sensory periphery we quantified ERG responses. In the ERG, OA (10−3 mol/L) elicited stronger receptor responses compared to controls. Tyramine (10−4 mol/L) had the opposite effect. The same pattern could be observed on the behavioural level. While OA elicited faster walking behaviour to light, indicating a stronger incentive value of the light, TA had the opposite effect. Neither amine affected the velocity during the dark runs and presumably also during the phototaxis assay. Our findings are in line with earlier studies showing that OA mainly has arousing functions in insects [5,7,32] and that it increases the perceived value of a food source of honeybee foragers [38]. Whether TA has similar or opposing effects is little-known so far. Only a few behavioural experiments were conducted including both biogenic amines. It has been shown that OA can decrease the walking speed towards different light sources when applied chronically, while TA can increase the walking speed in foragers [20]. However, TA also influenced the general locomotor activity in those experiments, so it is impossible to state whether the change in walking times due to TA treatment was a locomotor effect, an effect of increased light perception or a mixture of both in those experiments. The present study differs mainly in the drug application. While we injected both monoamines locally, the authors in the earlier study [20] fed them over three consecutive days before conducting the experiments, possibly resulting in opposite effects. Other studies could also show an arousing effect of OA on the phototaxis. Feeding formamidines, toxins reported to inhibit the OA-stimulated adenylate cyclase, to Drosophila reduced phototactic behaviour, indicating a positive effect of OA on the phototaxis [39]. Different application methods could possibly lead to different target receptors. Octopamine and tyramine are known to elicit different effects depending on their targeted receptors [7]. However, due to the lack of suitable antibodies, the location of most OA/TA receptors in the adult honeybee is still unknown. Only the spatial distribution of the octopamine receptor AmOctαR1 and of the tyramine receptor AmTAR1 have been described in the honeybee brain. While strong labelling of AmOctαR1 can be observed for the optic lobes [40], AmTAR1 is not present here [41]. This indicates that AmOctαR1 is a strong candidate for the observed effects in this study. In Drosophila, Kholy et al. [42] showed expression of Oamb and the TyrRIII in the optic lobes of Drosophila melanogaster. Similar to the AmOctαR1, the Oamb increases [cAMP]i [43]. However, a TyrRIII honeybee homologue, which decreases [cAMP]i [44] is currently unknown. Similar to flies, different OA/TA receptors should be present in the optic lobes of honeybees which might explain our results. Yet, our current experiments do not allow us to specify which receptors are activated by OA or TA. Here we suggest targeted knockout of individual receptor genes using CRISPR/Cas9 [45] or RNA interference [46] in future studies.
Both neurotransmitters are known to modulate not only the central nervous system (CNS) but also peripheral organs expressing respective receptors [47]. It has been shown that OA can target receptors in the CNS, as well as in the periphery, independently of the application method [30,48,49]. Therefore, application duration might be more important than the application itself. Application over three consecutive days might target the CNS and the periphery, while a local injection might preferably target receptors in the honeybee brain. Furthermore, TA is the metabolic precursor of octopamine [4]. Over time, TA might be converted into OA by the enzyme tyramine β hydroxylase. Thus, feeding TA over three consecutive days could lead to an OA effect rather than the expected TA effect. Additionally, a constant treatment of honeybees with either amine might lead to an internalization of the respective receptors which in turn could also lead to the opposite effect [49]. Here, it would be important in future investigations to quantify OA and TA brain titres directly after the phototaxis experiment. In addition to applying the CRISPR/Cas9 technique for a targeted knockout, RNA interference (RNAi) might be an interesting option to reduce receptor gene expression [50,51]. One could argue that an ocellus injection might have a strong negative impact on bee’s behaviour, which might lead to different results. However, we did show that the ocellus injection, does not negatively affect honeybee behaviour during the assay (Figure S2).
To find out whether the opposing effects of OA and TA on walking speed towards light were related to perception, we performed ERG experiments. Octopamine increased the photoreceptor response, while TA decreased it. These results show the same pattern as the phototaxis experiments, leading to the conclusion that sensory input and behavioural output might be directly linked. Little is known about the influence of biogenic amines on the ERG response of honeybees. Lim and Wasserman [52] showed that washing OA-containing seawater over the eye of Limulus polyphemus increases the receptor potential in ERG experiments, while Battelle et al. [53] showed that OA increased the ERG amplitude of a Limulus polyphemus eye. Erber et al. [54] demonstrated that OA could increase the visual antennal response. However, Chyb et al. [55] found OA to be decreasing the ERG response in Drosophila melanogaster. As seen in the previously mentioned behavioural studies, results of biogenic amine experiments can be contradicting. Another explanation might be that OA- and TA receptors are very similar. Tyramine does not only bind to TA receptors but also to OA receptors when applied in high concentrations, although TA has a much higher affinity towards TA receptors [14,15]. If TA bound to all TA receptors present, it might also have activated OA receptors. OA receptors could then elicit different or even opposite effects.
It seems likely that the results obtained in our study reflect the short-term modulation of the visual system in honeybees by OA and TA. Studies that obtained different results also differed in their application methods, indicating a difference between short- and long-term modulation of perception by OA and TA. This is supported by a study from Scheiner et al. [29]. They showed that injecting TA into the abdomen of honeybees leads to an increase in OA and TA in the honeybee brain. This effect is most likely a result of the tyramine β hydroxylase converting TA into OA. While the exact time point of the OA and TA titre quantification is not stated, PER experiments were conducted prior to the amine quantification of the same honeybees. This indicates that at least 1.5 hours passed between the injection and the quantification of both biogenic amines. This time seems sufficient for metabolizing TA into OA.
As stated before, the phototaxis and the ERG results both show the same opposing pattern for octopamine and tyramine. This coincides with the proposed hypothesis by Roeder et al. [4,5,6] stating that the OA/TA system in insects can be compared to the epinephrine/norepinephrine system in deuterostomes. This is also supported by other studies. Saraswati et al. [18] showed the opposing functions of OA and TA in the locomotion of Drosophila melanogaster larvae. Furthermore, Fussnecker et al. [56] showed that honeybees spent a significantly increased time flying when treated with OA, whereas those treated with TA spent a significantly decreased time flying compared to controls. Yet, when feeding or injecting OA and TA into the thorax, both increased the PER of honeybees [28]. This indicates that both substances fulfil complementary roles in some sensory systems but opposing functions in other systems. This study clearly shows the opposing functions of OA and TA on the visual system of honeybees. However, further experiments need to be conducted into differentiating long-term and short-term modulation as well as turning off or blocking single receptors to fully understand the modulating effects of both important biogenic amines.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/biom11091374/s1, Figure S1: Raw data of eight example bees for the ERG experiments, Figure S2: Puncturing the median ocellus does not affect mean walking velocity in the dark arena or walking time towards the switched-on LEDs.
Author Contributions
Conceptualization, M.T. and R.S.; methodology, F.S.; data analysis, F.S.; writing—original draft preparation, F.S.; writing—review and editing, F.S., M.T., M.S.-B. and R.S.; visualization, F.S.; supervision, M.S.-B. and R.S.; project administration, R.S.; funding acquisition, R.S. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Deutsche Forschungsgemeinschaft (DFG) through grant SCHE 1573/9-1 to R.S. This publication was supported by the Open Access Publication Fund of the University of Wuerzburg.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Correspondence and requests for materials should be addressed to F.S. and R.S.
Acknowledgments
We thank Josephine Gabler for help with the phototaxis experiments and Fabian Schmalz for help with the ERG experiments. Additionally, we thank the three anonymous reviewers for their constructive feedback.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Blenau, W.; Baumann, A. Molecular and pharmacological properties of insect biogenic amine receptors: Lessons from Drosophila melanogaster and Apis mellifera. Arch. Insect Biochem. Physiol. 2001, 48, 13–38. [Google Scholar] [CrossRef]
- Blenau, W.; Baumann, A. Aminergic signal transduction in invertebrates: Focus on tyramine and octopamine receptors. Recent Res. Dev. Neurochem. 2003, 6, 225–240. [Google Scholar]
- Evans, P.D. Biogenic Amines in the Insect Nervous System. Adv. Insect Phys. 1980, 15, 317–473. [Google Scholar] [CrossRef]
- Roeder, T.; Seifert, M.; Kähler, C.; Gewecke, M. Tyramine and octopamine: Antagonistic modulators of behavior and metabolism. Arch. Insect Biochem. Physiol. 2003, 54, 1–13. [Google Scholar] [CrossRef]
- Roeder, T. Tyramine and octopamine: Ruling behavior and metabolism. Annu. Rev. Entomol. 2005, 50, 447–477. [Google Scholar] [CrossRef]
- Roeder, T. The control of metabolic traits by octopamine and tyramine in invertebrates. J. Exp. Biol. 2020, 223, jeb194282. [Google Scholar] [CrossRef]
- Scheiner, R.; Baumann, A.; Blenau, W. Aminergic Control and Modulation of Honeybee Behaviour. Curr. Neuropharmacol. 2006, 4, 259–276. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.W.; Woodring, J. Effects of stress, age, season, and source colony on levels of octopamine, dopamine and serotonin in the honey bee (Apis mellifera L.) brain. J. Insect Physiol. 1992, 38, 29–35. [Google Scholar] [CrossRef]
- Wagener-Hulme, C.; Kuehn, J.C.; Schulz, D.J.; Robinson, G.E. Biogenic amines and division of labor in honey bee colonies. J. Comp. Physiol. A 1999, 184, 471–479. [Google Scholar] [CrossRef]
- Scheiner, R.; Reim, T.; Søvik, E.; Entler, B.V.; Barron, A.B.; Thamm, M. Learning, gustatory responsiveness and tyramine differences across nurse and forager honeybees. J. Exp. Biol. 2017, 220, 1443–1450. [Google Scholar] [CrossRef] [Green Version]
- Grohmann, L.; Blenau, W.; Erber, J.; Ebert, P.R.; Strünker, T.; Baumann, A. Molecular and functional characterization of an octopamine receptor from honeybee (Apis mellifera) brain. J. Neurochem. 2003, 86, 725–735. [Google Scholar] [CrossRef] [Green Version]
- Blenau, W.; Balfanz, S.; Baumann, A. Amtyr1: Characterization of a Gene from Honeybee (Apis mellifera) Brain Encoding a Functional Tyramine Receptor. J. Neurochem. 2000, 74, 900–908. [Google Scholar] [CrossRef]
- Blenau, W.; Wilms, J.A.; Balfanz, S.; Baumann, A. AmOctα2R: Functional characterization of a honeybee octopamine receptor inhibiting adenylyl cyclase activity. Int. J. Mol. Sci. 2020, 21, 9334. [Google Scholar] [CrossRef]
- Balfanz, S.; Jordan, N.; Langenstück, T.; Breuer, J.; Bergmeier, V.; Baumann, A. Molecular, pharmacological, and signaling properties of octopamine receptors from honeybee (Apis mellifera) brain. J. Neurochem. 2014, 129, 284–296. [Google Scholar] [CrossRef]
- Reim, T.; Balfanz, S.; Baumann, A.; Blenau, W.; Thamm, M.; Scheiner, R. AmTAR2: Functional characterization of a honeybee tyramine receptor stimulating adenylyl cyclase activity. Insect Biochem. Mol. Biol. 2017, 80, 91–100. [Google Scholar] [CrossRef]
- Behrends, A.; Scheiner, R. Octopamine improves learning in newly emerged bees but not in old foragers. J. Exp. Biol. 2012, 215, 1076–1083. [Google Scholar] [CrossRef] [Green Version]
- Bauknecht, P.; Jékely, G. Ancient coexistence of norepinephrine, tyramine, and octopamine signaling in bilaterians. BMC Biol. 2017, 15, 6. [Google Scholar] [CrossRef] [Green Version]
- Saraswati, S.; Fox, L.E.; Soll, D.R.; Wu, C.-F. Tyramine and octopamine have opposite effects on the locomotion of Drosophila larvae. J. Neurobiol. 2004, 58, 425–441. [Google Scholar] [CrossRef]
- Uzzan, A.; Dudai, Y. Aminergic Receptors in Drosophila melanogaster: Responsiveness of Adenylate Cyclase to Putative Neurotransmitters. J. Neurochem. 1982, 38, 1542–1550. [Google Scholar] [CrossRef]
- Scheiner, R.; Toteva, A.; Reim, T.; Søvik, E.; Barron, A.B. Differences in the phototaxis of pollen and nectar foraging honey bees are related to their octopamine brain titers. Front. Physiol. 2014, 5, 116. [Google Scholar] [CrossRef] [Green Version]
- Scheiner, R.; Steinbach, A.; Claßen, G.; Strudthoff, N.; Scholz, H. Octopamine indirectly affects proboscis extension response habituation in Drosophila melanogaster by controlling sucrose responsiveness. J. Insect Physiol. 2014, 69, 107–117. [Google Scholar] [CrossRef]
- Menzel, R.; Hammer, M.; Müller, U.; Rosenboom, H. Behavioral, neural and cellular components underlying olfactory learning in the honeybee. Proc. J. Physiol. Paris 1996, 90, 395–398. [Google Scholar] [CrossRef]
- Hammer, M.; Menzel, R. Learning and memory in the honeybee. J. Neurosci. 1995, 15, 1617–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.C.; Lee, H.G.; Lim, J.; Han, K.A. Appetitive learning requires the alpha1-like octopamine receptor oamb in the drosophila mushroom body neurons. J. Neurosci. 2013, 33, 1672–1677. [Google Scholar] [CrossRef] [Green Version]
- Kutsukake, M.; Komatsu, A.; Yamamoto, D.; Ishiwa-Chigusa, S. A tyramine receptor gene mutation causes a defective olfactory behavior in Drosophila melanogaster. Gene 2000, 245, 31–42. [Google Scholar] [CrossRef]
- McClung, C.; Hirsh, J. The trace amine tyramine is essential for sensitization to cocaine in Drosophila. Curr. Biol. 1999, 9, 853–860. [Google Scholar] [CrossRef] [Green Version]
- Braun, G.; Bicker, G. Habituation of an appetitive reflex in the honeybee. J. Neurophysiol. 1992, 67, 588–598. [Google Scholar] [CrossRef]
- Scheiner, R.; Plückhahn, S.; Öney, B.; Blenau, W.; Erber, J. Behavioural pharmacology of octopamine, tyramine and dopamine in honey bees. Behav. Brain Res. 2002, 136, 545–553. [Google Scholar] [CrossRef]
- Scheiner, R.; Entler, B.V.; Barron, A.B.; Scholl, C.; Thamm, M. The Effects of Fat Body Tyramine Level on Gustatory Responsiveness of Honeybees (Apis mellifera) Differ between Behavioral Castes. Front. Syst. Neurosci. 2017, 11, 55. [Google Scholar] [CrossRef] [Green Version]
- Scheiner, R.; Kuritz-Kaiser, A.; Menzel, R.; Erber, J. Sensory responsiveness and the effects of equal subjective rewards on tactile learning and memory of honeybees. Learn. Mem. 2005, 12, 626–635. [Google Scholar] [CrossRef] [Green Version]
- Barron, A.B.; Robinson, G.E. Selective modulation of task performance by octopamine in honey bee (Apis mellifera) division of labour. J. Comp. Physiol. A Neuroethol. 2005, 191, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Schulz, D.J.; Robinson, G.E. Octopamine influences division of labor in honey bee colonies. J. Comp. Physiol. A Sens. Neural Behav. Physiol. 2001, 187, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Reim, T.; Scheiner, R. Division of labour in honey bees: Age- and task-related changes in the expression of octopamine receptor genes. Insect Mol. Biol. 2014, 23, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Hunt, G.J.; Amdam, G.V.; Schlipalius, D.; Emore, C.; Sardesai, N.; Williams, C.E.; Rueppell, O.; Guzmán-Novoa, E.; Arechavaleta-Velasco, M.; Chandra, S.; et al. Behavioral genomics of honeybee foraging and nest defense. Naturwissenschaften 2007, 94, 247–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kloppenburg, P.; Erber, J. The modulatory effects of serotonin and octopamine in the visual system of the honey bee (Apis mellifera L.). J. Comp. Physiol. A 1995, 176, 119–129. [Google Scholar] [CrossRef]
- Thamm, M.; Balfanz, S.; Scheiner, R.; Baumann, A.; Blenau, W. Characterization of the 5-HT1A receptor of the honeybee (Apis mellifera) and involvement of serotonin in phototactic behavior. Cell. Mol. Life Sci. 2010, 67, 2467–2479. [Google Scholar] [CrossRef]
- Erber, J.; Hoormann, J.; Scheiner, R. Phototactic behaviour correlates with gustatory responsiveness in honey bees (Apis mellifera L.). Behav. Brain Res. 2006, 174, 174–180. [Google Scholar] [CrossRef]
- Barron, A.B.; Maleszka, R.; Vander Meer, R.K.; Robinson, G.E. Octopamine modulates honey bee dance behavior. Proc. Natl. Acad. Sci. USA 2007, 104, 1703–1707. [Google Scholar] [CrossRef] [Green Version]
- Dudai, Y.; Buxbaum, J.; Corfas, G.; Ofarim, M. Formamidines interact with Drosophila octopamine receptors, alter the flies’ behavior and reduce their learning ability. J. Comp. Physiol. A 1987, 161, 739–746. [Google Scholar] [CrossRef]
- Sinakevitch, I.; Mustard, J.A.; Smith, B.H. Distribution of the octopamine receptor AmOA1 in the honey bee brain. PLoS ONE 2011, 6, e14536. [Google Scholar] [CrossRef]
- Thamm, M.; Scholl, C.; Reim, T.; Grübel, K.; Möller, K.; Rössler, W.; Scheiner, R. Neuronal distribution of tyramine and the tyramine receptor AmTAR1 in the honeybee brain. J. Comp. Neurol. 2017, 525, 2615–2631. [Google Scholar] [CrossRef]
- El-Kholy, S.; Stephano, F.; Li, Y.; Bhandari, A.; Fink, C.; Roeder, T. Expression analysis of octopamine and tyramine receptors in Drosophila. Cell Tissue Res. 2015, 361, 669–684. [Google Scholar] [CrossRef] [PubMed]
- Balfanz, S.; Strünker, T.; Frings, S.; Baumann, A. A family of octopamine [corrected] receptors that specifically induce cyclic AMP production or Ca2+ release in Drosophila melanogaster. J. Neurochem. 2005, 93, 440–451. [Google Scholar] [CrossRef]
- Bayliss, A.; Roselli, G.; Evans, P.D. A comparison of the signalling properties of two tyramine receptors from Drosophila. J. Neurochem. 2013, 125, 37–48. [Google Scholar] [CrossRef]
- Değirmenci, L.; Geiger, D.; Rogé Ferreira, F.L.; Keller, A.; Krischke, B.; Beye, M.; Steffan-Dewenter, I.; Scheiner, R. CRISPR/Cas9 mediated mutations as a new tool for studying taste in honeybees. bioRxiv 2020. [Google Scholar] [CrossRef]
- Scheiner, R.; Kulikovskaja, L.; Thamm, M. The honey bee tyramine receptor AmTYR1 and division of foraging labour. J. Exp. Biol. 2014, 217, 1215–1217. [Google Scholar] [CrossRef] [Green Version]
- Degen, J.; Gewecke, M.; Roeder, T. Octopamine receptors in the honey bee and locust nervous system: Pharmacological similarities between homologous receptors of distantly related species. Br. J. Pharmacol. 2000, 130, 587–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barron, A.B.; Maleszka, J.; Vander Meer, R.K.; Robinson, G.E.; Maleszka, R. Comparing injection, feeding and topical application methods for treatment of honeybees with octopamine. J. Insect Physiol. 2007, 53, 187–194. [Google Scholar] [CrossRef]
- Wanka, L.; Behr, V.; Beck-Sickinger, A.G. Arrestin-dependent internalization of rhodopsin-like G protein-coupled receptors. Biol. Chem. 2021. [Google Scholar] [CrossRef]
- Farooqui, T.; Robinson, K.; Vaessin, H.; Smith, B.H. Modulation of early olfactory processing by an octopaminergic reinforcement pathway in the honeybee. J. Neurosci. Off. J. Soc. Neurosci. 2003, 23, 5370–5380. [Google Scholar] [CrossRef]
- Scholl, C.; Kübert, N.; Muenz, T.S.; Rössler, W. CaMKII knockdown affects both early and late phases of olfactory long-term memory in the honeybee. J. Exp. Biol. 2015, 218, 3788–3796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, C.C.M.; Wasserman, G.S. Categorical and Prolonged Potentials Are Evoked when Brief, Intermediate-Intensity Flashes Stimulate Horseshoe Crab Lateral Eye Photoreceptors during Octopamine Neuromodulation. Neurosignals 2001, 10, 399–415. [Google Scholar] [CrossRef]
- Battelle, B.A.; Edwards, S.C.; Kass, L.; Maresch, H.M.; Pierce, S.K.; Wishart, A.C. Identification and Function of Octopamine and Tyramine Conjugates in the Limulus Visual System. J. Neurochem. 1988, 51, 1240–1251. [Google Scholar] [CrossRef] [PubMed]
- Erber, J.; Kloppenburg, P.; Scheidler, A. Neuromodulation by serotonin and octopamine in the honeybee: Behaviour, neuroanatomy and electrophysiology. Experientia 1993, 49, 1073–1083. [Google Scholar] [CrossRef]
- Chyb, S.; Hevers, W.; Forte, M.; Wolfgang, W.J.; Selinger, Z.; Hardie, R.C. Modulation of the light response by cAMP in Drosophila photoreceptors. J. Neurosci. Off. J. Soc. Neurosci. 1999, 19, 8799–8807. [Google Scholar] [CrossRef] [Green Version]
- Fussnecker, B.L.; Smith, B.H.; Mustard, J.A. Octopamine and tyramine influence the behavioral profile of locomotor activity in the honey bee (Apis mellifera). J. Insect Physiol. 2006, 52, 1083–1092. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
ERG−amplitude receptor response pre-post for three concentrations of octopamine (x-axis in A–C) and three concentrations of tyramine (x-axis in D–F) and the respective Ringer control (R). The different light intensities are shown at the top. The median is marked in red. Dots, squares, upper triangles and lower triangles represent individual data points for Ringer, OA/TA 10−5 mol/L, OA/TA 10−4 mol/L, OA/TA 10−3 mol/L, respectively. (A–C): Pre-post response after OA and Ringer were applied using 36%, 59% and 100% light intensity filters for A, B, and C respectively. A significant overall effect was found for all three light intensities. OA 10−3 mol/L significantly increased the amplitude (for statistics, see Table 2). (D–F): Pre-post responses after TA or Ringer were applied using 36%, 59% and 100% light intensity filters for D, E, and F respectively. No significant differences were found for the 36% light intensity filter. A significant overall effect was found for 59% and 100% light intensity. TA 10−4 mol/L significantly decreased the amplitude (for statistics, see Table 2). Significant differences between Ringer and either treatment are indicated by asterisks (* p < 0.05, ** p < 0.01).
Figure 1.
ERG−amplitude receptor response pre-post for three concentrations of octopamine (x-axis in A–C) and three concentrations of tyramine (x-axis in D–F) and the respective Ringer control (R). The different light intensities are shown at the top. The median is marked in red. Dots, squares, upper triangles and lower triangles represent individual data points for Ringer, OA/TA 10−5 mol/L, OA/TA 10−4 mol/L, OA/TA 10−3 mol/L, respectively. (A–C): Pre-post response after OA and Ringer were applied using 36%, 59% and 100% light intensity filters for A, B, and C respectively. A significant overall effect was found for all three light intensities. OA 10−3 mol/L significantly increased the amplitude (for statistics, see Table 2). (D–F): Pre-post responses after TA or Ringer were applied using 36%, 59% and 100% light intensity filters for D, E, and F respectively. No significant differences were found for the 36% light intensity filter. A significant overall effect was found for 59% and 100% light intensity. TA 10−4 mol/L significantly decreased the amplitude (for statistics, see Table 2). Significant differences between Ringer and either treatment are indicated by asterisks (* p < 0.05, ** p < 0.01).

Figure 2.
Octopamine and tyramine have opposite effects on the phototaxis of in-hive bees. Control bees are shown in light grey. Those injected with OA (10−2 mol/L) or TA (10−2 mol/L) are shown in dark grey. (A,B): Average velocity (mean + standard deviation) of honeybees during one minute of constant movement in the dark arena. Neither OA nor TA differed significantly from the Ringer control in their mean velocity in the dark (Table 3). (C,D): Average walking time (mean + standard error) towards the different switched-on light sources. The factor light intensity significantly influenced phototactic behaviour (Table 3). Octopamine decreased the time honeybees needed to reach the switched-on LEDs significantly compared to the control solution, while TA increased it (Table 3). Significant differences between Ringer and OA/TA are indicated by asterisks (n.s. p > 0.05, * p < 0.05).
Figure 2.
Octopamine and tyramine have opposite effects on the phototaxis of in-hive bees. Control bees are shown in light grey. Those injected with OA (10−2 mol/L) or TA (10−2 mol/L) are shown in dark grey. (A,B): Average velocity (mean + standard deviation) of honeybees during one minute of constant movement in the dark arena. Neither OA nor TA differed significantly from the Ringer control in their mean velocity in the dark (Table 3). (C,D): Average walking time (mean + standard error) towards the different switched-on light sources. The factor light intensity significantly influenced phototactic behaviour (Table 3). Octopamine decreased the time honeybees needed to reach the switched-on LEDs significantly compared to the control solution, while TA increased it (Table 3). Significant differences between Ringer and OA/TA are indicated by asterisks (n.s. p > 0.05, * p < 0.05).

Figure 3.
Octopamine and tyramine have opposite effects on the phototaxis of foragers. Control bees are shown in light grey. Those injected with OA (10−2 mol/L) or TA (10−2 mol/L) are shown in dark grey. (A,B): Average velocity (mean + standard deviation) of honeybees during one minute of constant movement in the dark arena. Neither OA nor TA did differ significantly from Ringer in their mean velocity in the dark (Table 3). (C,D): Average walking time (mean + standard error) towards the different switched-on light sources. The light intensity factor significantly influenced phototacic behavior (Table 3). Octopamine decreased the time honeybees needed to reach the switched-on LEDs significantly compared to the control solution, while TA increased it (Table 3). Significant differences between Ringer and OA/TA are indicated by asterisks (n.s. p > 0.05, * p < 0.05, ** p < 0.01).
Figure 3.
Octopamine and tyramine have opposite effects on the phototaxis of foragers. Control bees are shown in light grey. Those injected with OA (10−2 mol/L) or TA (10−2 mol/L) are shown in dark grey. (A,B): Average velocity (mean + standard deviation) of honeybees during one minute of constant movement in the dark arena. Neither OA nor TA did differ significantly from Ringer in their mean velocity in the dark (Table 3). (C,D): Average walking time (mean + standard error) towards the different switched-on light sources. The light intensity factor significantly influenced phototacic behavior (Table 3). Octopamine decreased the time honeybees needed to reach the switched-on LEDs significantly compared to the control solution, while TA increased it (Table 3). Significant differences between Ringer and OA/TA are indicated by asterisks (n.s. p > 0.05, * p < 0.05, ** p < 0.01).


{kind=link}
{kind=link}
{kind=link}
Table 1.
Analysis of the ERG response of three light intensity filters before and after the application of octopamine/tyramine (treatment) or Ringer (control).
Table 1.
Analysis of the ERG response of three light intensity filters before and after the application of octopamine/tyramine (treatment) or Ringer (control).
| OA—Pre-Response | Friedmann Test | χ2(2) | p Value | ||
|---|---|---|---|---|---|
| Ringer—Control | 14.63 | <0.001 | *** | ||
| Dunn´s test | n (Ringer) | n (Treatment) | |||
| 36% vs. 59% | 16 | 16 | 0.024 | * | |
| 36% vs. 100% | 16 | 16 | <0.001 | *** | |
| 59% vs. 100% | 16 | 16 | 0.867 | n.s. | |
| OA—post response | Friedmann test | χ2(2) | p value | ||
| Ringer—Control | 16.63 | <0.001 | *** | ||
| Dunn´s test | n (Ringer) | n (Treatment) | |||
| 36% vs. 59% | 16 | 16 | 0.008 | ** | |
| 36% vs. 100% | 16 | 16 | <0.001 | *** | |
| 59% vs. 100% | 16 | 16 | 0.99 | n.s. | |
| OA—pre-response | Friedmann test | χ2(2) | p value | ||
| OA—Treatment | 26.79 | <0.001 | *** | ||
| Dunn´s test | n (Ringer) | n (Treatment) | |||
| 36% vs. 59% | 48 | 48 | 0.001 | ** | |
| 36% vs. 100% | 48 | 48 | <0.001 | *** | |
| 59% vs. 100% | 48 | 48 | 0.99 | n.s. | |
| OA—post response | Friedmann test | χ2(2) | p value | ||
| OA—Treatment | 38.17 | <0.001 | *** | ||
| Dunn´s test | n (Ringer) | n (Treatment) | |||
| 36% vs. 59% | 48 | 48 | <0.001 | *** | |
| 36% vs. 100% | 48 | 48 | <0.001 | *** | |
| 59% vs. 100% | 48 | 48 | 0.459 | n.s. | |
| TA—pre-response | Friedmann test | χ2(2) | p value | ||
| Ringer—Control | 24 | <0.001 | *** | ||
| Dunn´s test | n (Ringer) | n (Treatment) | |||
| 36% vs. 59% | 12 | 12 | 0.043 | * | |
| 36% vs. 100% | 12 | 12 | <0.001 | *** | |
| 59% vs. 100% | 12 | 12 | 0.043 | * | |
| TA—post response | Friedmann test | χ2(2) | p value | ||
| Ringer—Control | 24 | <0.001 | *** | ||
| Dunn´s test | n (Ringer) | n (Treatment) | |||
| 36% vs. 59% | 12 | 12 | 0.043 | * | |
| 36% vs. 100% | 12 | 12 | <0.001 | *** | |
| 59% vs. 100% | 12 | 12 | 0.043 | * | |
| TA—pre-response | Friedmann test | χ2(2) | p value | ||
| TA—Treatment | 70.06 | <0.001 | *** | ||
| Dunn´s test | n (Ringer) | n (Treatment) | |||
| 36% vs. 59% | 36 | 36 | <0.001 | *** | |
| 36% vs. 100% | 36 | 36 | <0.001 | *** | |
| 59% vs. 100% | 36 | 36 | <0.001 | *** | |
| TA—post response | Friedmann test | χ2(2) | p value | ||
| TA—Treatment | 62.39 | <0.001 | *** | ||
| Dunn´s test | n (Ringer) | n (Treatment) | |||
| 36% vs. 59% | 36 | 36 | <0.001 | *** | |
| 36% vs. 100% | 36 | 36 | <0.001 | *** | |
| 59% vs. 100% | 36 | 36 | <0.001 | *** |
Significant differences are indicated by asterisks (n.s. p > 0.05, * p < 0.05, ** p < 0.01, *** p < 0.001).
Table 2.
Statistical comparison of the octopamine/tyramine (treatment) or Ringer (control) pre-post receptor response in the honeybee retina.
Table 2.
Statistical comparison of the octopamine/tyramine (treatment) or Ringer (control) pre-post receptor response in the honeybee retina.
| OA—Pre-Post Response | 1way ANOVA | F (3, 54) | p Value | ||||
|---|---|---|---|---|---|---|---|
| - 36% light intensity | 4091 | 0.011 | * | ||||
| Dunnett´s test | n (Ringer) | n (Treatment) | q | DF | |||
| R vs. OA 10-3 | 13 | 14 | 3457 | 54 | 0.003 | ** | |
| R vs. OA 10-4 | 13 | 15 | 1667 | 54 | 0.237 | n.s. | |
| R vs. OA 10-5 | 13 | 16 | 1.37 | 54 | 0.386 | n.s. | |
| OA—pre-post response | 1way ANOVA | F (3, 55) | p value | ||||
| - 59% light intensity | 3176 | 0.031 | * | ||||
| Dunnett´s test | n (Ringer) | n (Treatment) | q | DF | |||
| R vs. OA 10-3 | 13 | 14 | 3078 | 55 | 0.009 | ** | |
| R vs. OA 10-4 | 13 | 16 | 1768 | 55 | 0.197 | n.s. | |
| R vs. OA 10-5 | 13 | 16 | 1544 | 55 | 0.293 | n.s. | |
| OA—pre-post response | 1way ANOVA | F (3, 57) | p value | ||||
| - 100% light intensity | 2821 | 0.0469 | * | ||||
| Dunnett´s test | n (Ringer) | n (Treatment) | q | DF | |||
| R vs. OA 10-3 | 13 | 16 | 2744 | 57 | 0.022 | * | |
| R vs. OA 10-4 | 13 | 16 | 1232 | 57 | 0.471 | n.s. | |
| R vs. OA 10-5 | 13 | 16 | 2118 | 57 | 0.098 | n.s. | |
| TA—pre-post response | 1way ANOVA | F (3, 39) | p value | ||||
| - 36% light intensity | 1.16 | 0.337 | n.s. | ||||
| Dunnett´s test | n (Ringer) | n (Treatment) | statistics | DF | |||
| R vs. TA 10-3 | 11 | 10 | 1404 | 39 | 0.37 | n.s. | |
| R vs. TA 10-4 | 11 | 11 | 0.892 | 39 | 0.703 | n.s. | |
| R vs. TA 10-5 | 11 | 11 | 0.232 | 39 | 0.991 | n.s. | |
| TA—pre-post response | 1way ANOVA | F (3, 39) | p value | ||||
| - 59% light intensity | 3304 | 0.03 | * | ||||
| Dunnett´s test | n (Ringer) | n (Treatment) | q | DF | |||
| R vs. TA 10-3 | 11 | 11 | 2 | 39 | 0.211 | n.s. | |
| R vs. TA 10-4 | 11 | 10 | 2677 | 39 | 0.029 | * | |
| R vs. TA 10-5 | 11 | 11 | 0.162 | 39 | 0.997 | n.s. | |
| TA—pre-post response | 1way ANOVA | F (3, 39) | p value | ||||
| - 100% light intensity | 3492 | 0.025 | * | ||||
| Dunnett´s test | n (Ringer) | n (Treatment) | q | DF | |||
| R vs. TA 10-3 | 11 | 10 | 2373 | 39 | 0.059 | n.s. | |
| R vs. TA 10-4 | 11 | 10 | 2702 | 39 | 0.027 | * | |
| R vs. TA 10-5 | 11 | 12 | 0.6678 | 39 | 0.844 | n.s. |
Significant differences are indicated by asterisks (n.s. p > 0.05, * p < 0.05, ** p < 0.01).
Table 3.
Statistical comparison of the dark runs and the phototaxis of honeybees (either in-hive bees or foragers) treated with octopamine/tyramine (treatment) or Ringer (control).
Table 3.
Statistical comparison of the dark runs and the phototaxis of honeybees (either in-hive bees or foragers) treated with octopamine/tyramine (treatment) or Ringer (control).
| Octopamine (In-Hive) | Unpaired t Test | Statistic | DF | n (Ringer) | n (Treatment) | p Value | |
|---|---|---|---|---|---|---|---|
| Dark run | t = 1.679 | 34 | 19 | 17 | 0.102 | n.s. | |
| Phototaxis | 2way ANOVA | ||||||
| Intensity | F(5, 170) = 6.131 | <0.001 | *** | ||||
| Treatment | F(1, 34) = 5.750 | 0.022 | * | ||||
| Interaction | F(5, 170) = 0.625 | 0.681 | n.s. | ||||
| Bonferroni test | |||||||
| 3% | 0.824 | 204 | 19 | 17 | 0.999 | n.s. | |
| 6% | 1258 | 204 | 19 | 17 | 0.999 | n.s. | |
| 12% | 1960 | 204 | 19 | 17 | 0.308 | n.s. | |
| 25% | 1874 | 204 | 19 | 17 | 0.374 | n.s. | |
| 50% | 2531 | 204 | 19 | 17 | 0.073 | n.s. | |
| 100% | 1017 | 204 | 19 | 17 | 0.999 | n.s. | |
| Tyramine (in-hive) | unpaired t test | statistic | DF | n (Ringer) | n (Treatment) | p value | |
| Dark run | t = 0.139 | 49 | 25 | 26 | 0.890 | n.s. | |
| Phototaxis | 2way ANOVA | ||||||
| Intensity | F(5, 245) = 2.564 | 0.028 | * | ||||
| Treatment | F(1, 49) = 4.919 | 0.031 | * | ||||
| Interaction | F(5, 245) = 0.669 | 0.647 | n.s. | ||||
| Bonferroni test | |||||||
| 3% | 1916 | 294 | 25 | 26 | 0.338 | n.s. | |
| 6% | 1931 | 294 | 25 | 26 | 0.327 | n.s. | |
| 12% | 2236 | 294 | 25 | 26 | 0.157 | n.s. | |
| 25% | 0.964 | 294 | 25 | 26 | 0.999 | n.s. | |
| 50% | 1452 | 294 | 25 | 26 | 0.886 | n.s. | |
| 100% | 2352 | 294 | 25 | 26 | 0.116 | n.s. | |
| Octopamine (forager) | unpaired t test | statistic | DF | n (Ringer) | n (Treatment) | p value | |
| Dark run | t = 1.595 | 38 | 20 | 20 | 0.119 | n.s. | |
| Phototaxis | 2way ANOVA | ||||||
| Intensity | F(5, 190) = 4.342 | < 0.001 | *** | ||||
| Treatment | F(1, 38) = 5.223 | 0.028 | * | ||||
| Interaction | F(5, 190) = 1.514 | 0.187 | n.s. | ||||
| Bonferroni test | |||||||
| 3% | 2695 | 228 | 20 | 20 | 0.045 | * | |
| 6% | 1630 | 228 | 20 | 20 | 0.627 | n.s. | |
| 12% | 1700 | 228 | 20 | 20 | 0.543 | n.s. | |
| 25% | 2492 | 228 | 20 | 20 | 0.081 | n.s. | |
| 50% | 1610 | 228 | 20 | 20 | 0.653 | n.s. | |
| 100% | 2545 | 228 | 20 | 20 | 0.070 | n.s. | |
| Tyramine (forager) | unpaired t test | statistic | DF | n (Ringer) | n (Treatment) | p value | |
| Dark run | t = 0.123 | 48 | 25 | 25 | 0.903 | n.s. | |
| Phototaxis | 2way ANOVA | ||||||
| Intensity | F(5, 245) = 5.986 | < 0.001 | *** | ||||
| Treatment | F(1, 49) = 11.29 | 0.002 | ** | ||||
| Interaction | F(5, 245) = 0.072 | 0.996 | n.s. | ||||
| Bonferroni test | |||||||
| 3% | 2412 | 294 | 26 | 25 | 0.099 | n.s. | |
| 6% | 2838 | 294 | 26 | 25 | 0.029 | * | |
| 12% | 2435 | 294 | 26 | 25 | 0.093 | n.s. | |
| 25% | 2268 | 294 | 26 | 25 | 0.144 | n.s. | |
| 50% | 2377 | 294 | 26 | 25 | 0.109 | n.s. | |
| 100% | 2334 | 294 | 26 | 25 | 0.122 | n.s. |
Significant differences are indicated by asterisks (n.s. p > 0.05, * p < 0.05, ** p < 0.01, *** p < 0.001).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Schilcher, F.; Thamm, M.; Strube-Bloss, M.; Scheiner, R. Opposing Actions of Octopamine and Tyramine on Honeybee Vision. Biomolecules 2021, 11, 1374. https://doi.org/10.3390/biom11091374
AMA Style
Schilcher F, Thamm M, Strube-Bloss M, Scheiner R. Opposing Actions of Octopamine and Tyramine on Honeybee Vision. Biomolecules. 2021; 11(9):1374. https://doi.org/10.3390/biom11091374
Chicago/Turabian StyleSchilcher, Felix, Markus Thamm, Martin Strube-Bloss, and Ricarda Scheiner. 2021. "Opposing Actions of Octopamine and Tyramine on Honeybee Vision" Biomolecules 11, no. 9: 1374. https://doi.org/10.3390/biom11091374
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.