
Morphometric Maps of Bilateral Asymmetry in the Human Humerus: An Implementation in the R Package Morphomap

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Preparation and Processing
2.2. Asymmetry and Cross-Sectional Geometry
2.3. Description of the Function MorphomapAsymmetry
- Load the output of the first long bone processed with morphomapShape.
- Load the output of the second long bone processed with morphomapShape (Figure 1).
- Specify if one of the two input objects needs to be mirrored (Figure 2A).
- Calculate the cortical thickness map of the entire diaphysis in both long bones (Figure 2B).
- Standardize the cortical thicknesses by dividing the matrices of cortical thickness by the biomechanical length (optional).
- Choose the method of visualization by setting the argument type to:
- type = “diff”Calculate the differences between the cortical thickness maps of the two long bones (Figure 2C).
- type = “onMean”Calculate the differences between the two cortical maps and their mean (Figure 2E).
- type = “relChange”Compute a cortical map as the percentage change of one side (target) with respect to the other one (reference) (Figure 2D).
- 2D Plot the map of differences in cortical thickness between the selected specimens. The difference map is displayed after “unrolling” the long-bone shaft to produce a 2D plot, starting and ending at the anterior (A) border passing through the lateral (L), posterior (P), and medial (M) borders (Figure 2C).
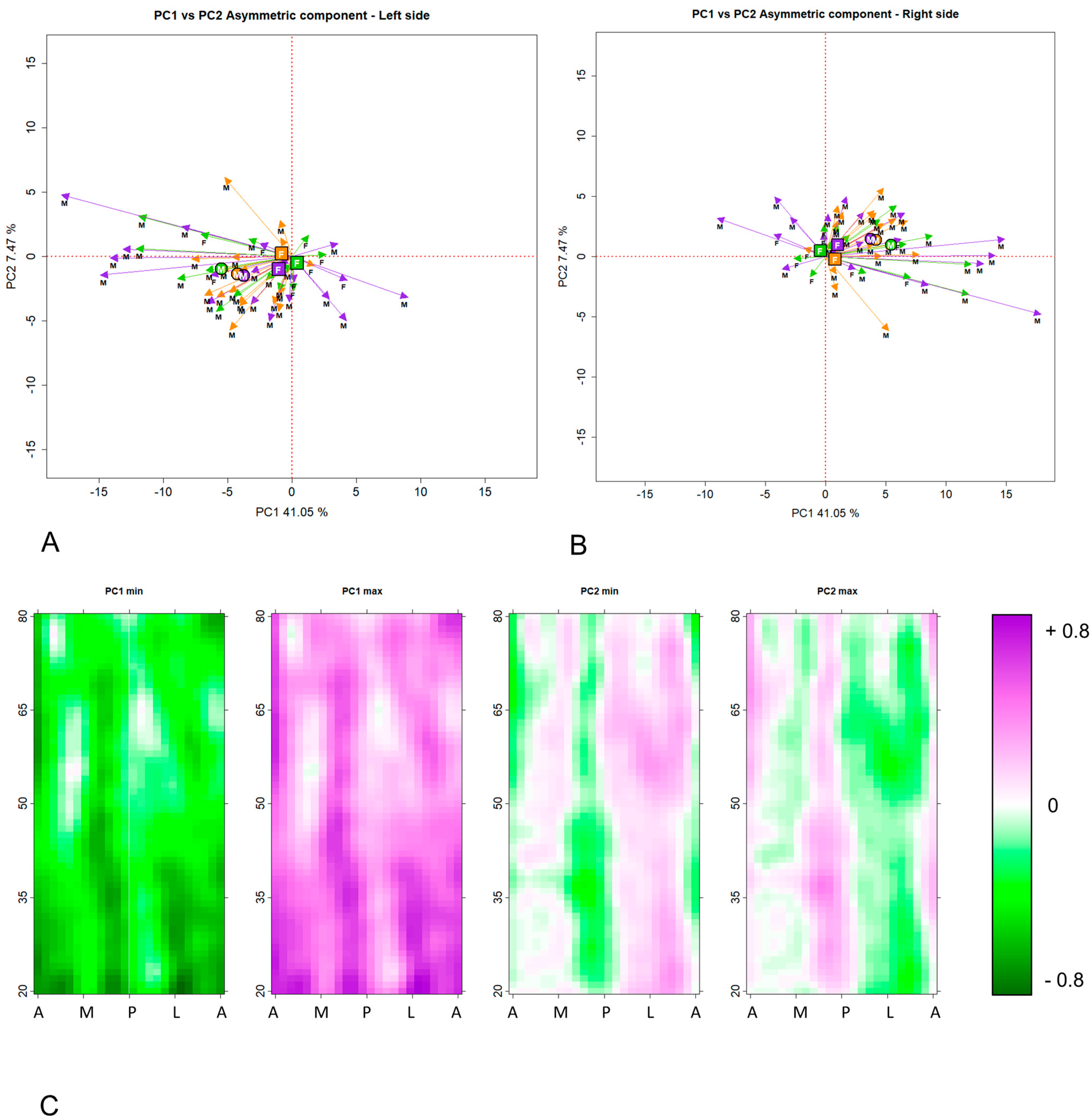
2.4. Description of the Function MorphomapMapPCA
- PCA of the mean morphometric maps calculated by averaging left and mirrored right side (symmetric component).
- PCA of the matrices obtained by in each individual subtracting the mean matrix of cortical thicknesses from the matrices of cortical thicknesses of the left and mirrored right sides (asymmetric component).
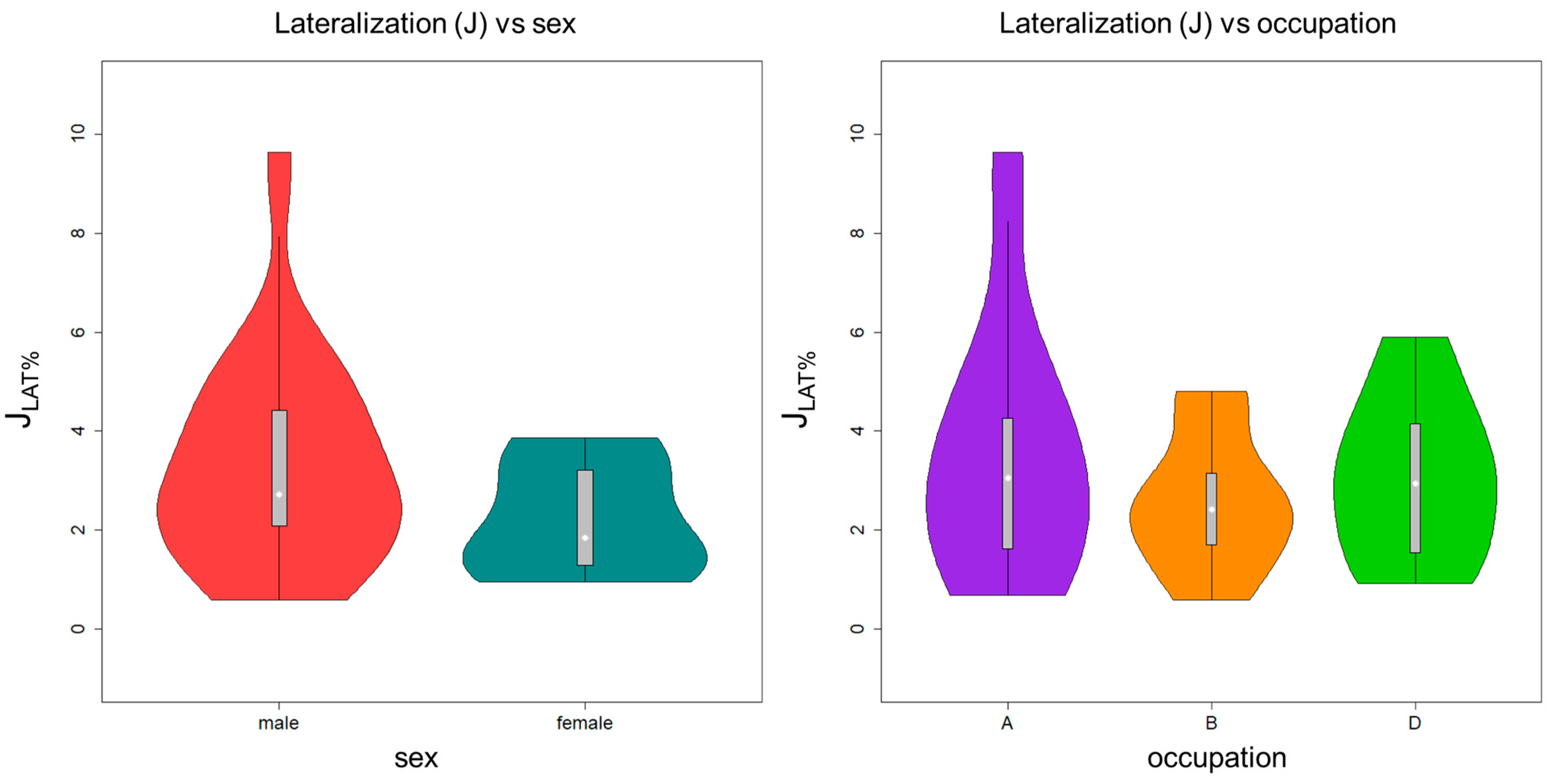
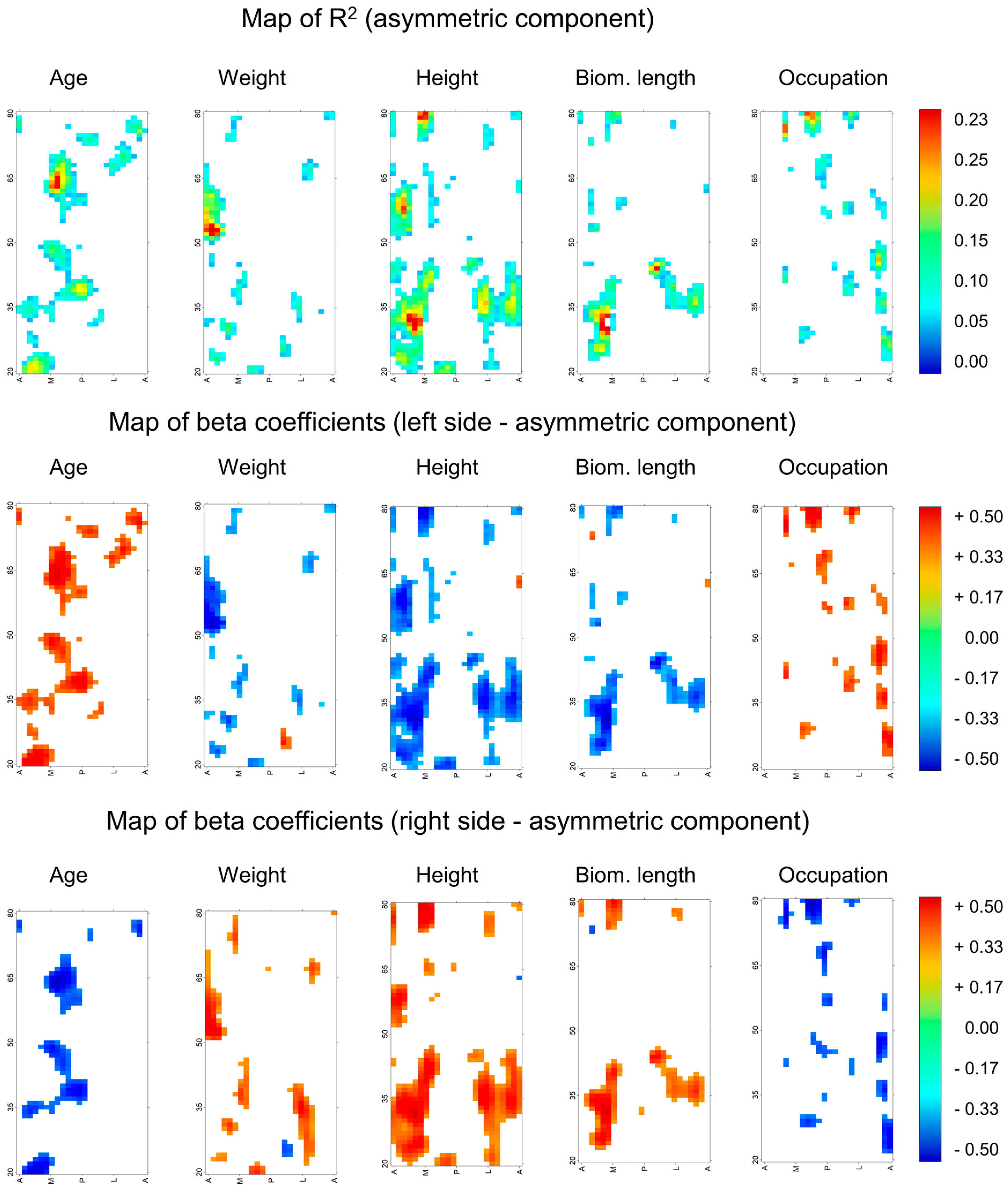
2.5. Relation between Cortical Thickness Asymmetry Humerus and Biological Variables
3. Results
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jurmain, R.; Cardoso, F.A.; Henderson, C.; Villotte, S. Bioarchaeology’s holy grail: The reconstruction of activity. In A Companion to Paleopathology; Grauer, A.L., Ed.; Wiley-Blackwell: Oxford, UK, 2011; pp. 531–552. [Google Scholar]
- Larsen, C.S. Bioarchaeology: Interpreting Behavior from the Human Skeleton, 2nd ed.; Cambridge Studies in Biological and Evolutionary Anthropology; Cambridge University Press: Cambridge, UK, 2015. [Google Scholar]
- Ruff, C.; Holt, B.; Trinkaus, E. Who’s afraid of the big bad Wolff? “Wolff’s law” and bone functional adaptation. Am. J. Phys. Anthr. 2006, 129, 484–498. [Google Scholar] [CrossRef]
- Ruff, C.B. Biomechanical analyses of archaeological human skeletons. In Biological Anthropology of the Human Skeleton, 2nd ed.; Katzenberg, M.A., Saunders, S.R., Eds.; Alan R. Liss: New York, NY, USA, 2008; pp. 183–206. [Google Scholar]
- Trinkaus, E.; Churchill, S.E.; Ruff, C.B. Postcranial robusticity inHomo. II: Humeral bilateral asymmetry and bone plasticity. Am. J. Phys. Anthr. 1994, 93, 1–34. [Google Scholar] [CrossRef]
- Miller, B.F.; Hamilton, K.L.; Majeed, Z.R.; Abshire, S.M.; Confides, A.L.; Hayek, A.M.; Hunt, E.R.; Shipman, P.; Peelor, F.F.; Butterfield, T.A.; et al. Enhanced skeletal muscle regrowth and remodelling in massaged and contralateral non-massaged hindlimb. J. Physiol. 2017, 596, 83–103. [Google Scholar] [CrossRef] [PubMed]
- Trinkaus, E.; Churchill, S.E. Diaphyseal cross-sectional geometry of near eastern middle palaeolithic humans: The humerus. J. Archaeol. Sci. 1999, 26, 173–184. [Google Scholar] [CrossRef]
- Frost, H.M. Orthopaedic Biomechanics; Charles C. Thomas: Springfield, IL, USA, 1973. [Google Scholar]
- Currey, J. The many adaptations of bone. J. Biomech. 2003, 36, 1487–1495. [Google Scholar] [CrossRef]
- Oxnard, C.E. Thoughts on bone biomechanics. Folia Primatol. 2004, 75, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, F.A.; Henderson, C. Enthesopathy formation in the humerus: Data from known age-at-death and known occupation skeletal collections. Am. J. Phys. Anthr. 2009, 141, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Weiss, E. Humeral cross-sectional morphology from 18th century Quebec prisoners of war: Limits to activity reconstruction. Am. J. Phys. Anthr. 2005, 126, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Bertram, J.E.A.; Swartz, S.M. The ‘law of bone transformation’: A case of crying wolff? Biol. Rev. 1991, 66, 245–273. [Google Scholar] [CrossRef]
- Melcher, A.H.; Accursi, G.E. Transmission of an “osteogenic message” through intact bone after wounding. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1972, 173, 265–275. [Google Scholar] [CrossRef]
- Pearson, O.M.; Lieberman, D.E. The aging of Wolff’s “law”: Ontogeny and responses to mechanical loading in cortical bone. Am. J. Phys. Anthr. 2004, 125, 63–99. [Google Scholar] [CrossRef] [PubMed]
- Ruff, C.B.; Jones, H.H. Bilateral asymmetry in cortical bone of the humerus and tibia—Sex and age factors. Hum. Biol. 1981, 53, 69–86. [Google Scholar] [PubMed]
- Frost, H.M. An overview: Spinal tissue vital biomechanics for clinicians. In Spinal Disorders in Growth and Aging; Takahashi, H.E., Ed.; Springer: Tokyo, Japan, 1995; pp. 95–126. [Google Scholar]
- Sparacello, V.; Pearson, O.; Coppa, A.; Marchi, D. Changes in skeletal robusticity in an iron age agropastoral group: The samnites from the Alfedena necropolis (Abruzzo, Central Italy). Am. J. Phys. Anthr. 2010, 144, 119–130. [Google Scholar] [CrossRef]
- Rhodes, J.A.; Knüsel, C.J. Activity-related skeletal change in medieval humeri: Cross-sectional and architectural alterations. Am. J. Phys. Anthr. 2005, 128, 536–546. [Google Scholar] [CrossRef] [Green Version]
- Tur, S. Bilateral asymmetry of long bones in bronze and early iron age pastoralists of the altai. Archaeol. Ethnol. Anthr. Eurasia 2014, 42, 141–156. [Google Scholar] [CrossRef]
- Sládek, V.; Ruff, C.B.; Berner, M.; Holt, B.; Niskanen, M.; Schuplerová, E.; Hora, M. The impact of subsistence changes on humeral bilateral asymmetry in Terminal Pleistocene and Holocene Europe. J. Hum. Evol. 2016, 92, 37–49. [Google Scholar] [CrossRef] [Green Version]
- Kubicka, A.M.; Nowaczewska, W.; Balzeau, A.; Piontek, J. Bilateral asymmetry of the humerus in Neandertals, Australian aborigines and medieval humans. Am. J. Phys. Anthr. 2018, 167, 46–60. [Google Scholar] [CrossRef] [PubMed]
- Churchill, S.E.; Formicola, V. A case of marked bilateral asymmetry in the Upper Limbs of an Upper Palaeolithic Male from Barma Grande (Liguria). Italy Int. J. Osteoarchaeol. 1997, 7, 18–38. [Google Scholar] [CrossRef]
- Perchalski, B.; Placke, A.; Sukhdeo, S.M.; Shaw, C.N.; Gosman, J.H.; Raichlen, D.A.; Ryan, T.M. Asymmetry in the cortical and trabecular bone of the human humerus during development. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2018, 301, 1012–1025. [Google Scholar] [CrossRef] [Green Version]
- Ben-Itzhak, S.; Smith, P.; Bloom, R.A. Radiographic study of the humerus in Neandertals and Homo sapiens sapiens. Am. J. Phys. Anthr. 1988, 77, 231–242. [Google Scholar] [CrossRef]
- Shaw, C.N.; Hofmann, C.L.; Petraglia, M.D.; Stock, J.T.; Gottschall, J.S. Neandertal humeri may reflect adaptation to scraping tasks, but not spear thrusting. PLoS ONE 2012, 7, e40349. [Google Scholar] [CrossRef] [Green Version]
- Sparacello, V.S.; Villotte, S.; Shackelford, L.L.; Trinkaus, E. Patterns of humeral asymmetry among Late Pleistocene humans. C. R. Palevol 2017, 16, 680–689. [Google Scholar] [CrossRef]
- Vandermeersch, B.; Trinkaus, E. The postcranial remains of the Régourdou 1 Neandertal: The shoulder and arm remains. J. Hum. Evol. 1995, 28, 439–476. [Google Scholar] [CrossRef]
- Pérez-Criado, L.; Rosas, A.; Bastir, M.; Pastor, F. Humeral laterality in modern humans and Neanderthals: A 3D geometric morphometric analysis. Anthr. Sci. 2017, 125, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Uomini, N.T. Handedness in Neanderthals. In Neanderthal Lifeways, Subsistence and Technology: One Hundred Fifty Years of Neanderthal Study; Conard, N.J., Richter, J., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 139–154. [Google Scholar]
- Ireland, A.; Maden-Wilkinson, T.; Mcphee, J.; Cooke, K.; Narici, M.; Degens, H.; Rittweger, J. Upper limb muscle-bone asymmetries and bone adaptation in elite youth tennis players. Med. Sci. Sports Exerc. 2013, 45, 1749–1758. [Google Scholar] [CrossRef] [Green Version]
- Haapasalo, H.; Kontulainen, S.; Sievänen, H.; Kannus, P.; Järvinen, M.; Vuori, I. Exercise-induced bone gain is due to enlargement in bone size without a change in volumetric bone density: A peripheral quantitative computed tomography study of the upper arms of male tennis players. Bone 2000, 27, 351–357. [Google Scholar] [CrossRef] [Green Version]
- Kontulainen, S.; Sievänen, H.; Kannus, P.; Pasanen, M.; Vuori, I. Effect of long-term impact-loading on mass, size, and estimated strength of humerus and radius of female racquet-sports players: A peripheral quantitative computed tomography study between young and old starters and controls. J. Bone Miner. Res. 2003, 18, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Nikander, R.; Sievänen, H.; Uusi-Rasi, K.; Heinonen, A.; Kannus, P. Loading modalities and bone structures at nonweight-bearing upper extremity and weight-bearing lower extremity: A pQCT study of adult female athletes. Bone 2006, 39, 886–894. [Google Scholar] [CrossRef]
- Shaw, C.N.; Stock, J.T. Habitual throwing and swimming correspond with upper limb diaphyseal strength and shape in modern human athletes. Am. J. Phys. Anthr. 2009, 140, 160–172. [Google Scholar] [CrossRef]
- Qu, X. Morphological effects of mechanical forces on the human humerus. Br. J. Sports Med. 1992, 26, 51–53. [Google Scholar] [CrossRef] [Green Version]
- Ruff, C.B.; Walker, A.; Trinkaus, E. Postcranial robusticity inHomo. III: Ontogeny. Am. J. Phys. Anthr. 1994, 93, 35–54. [Google Scholar] [CrossRef]
- Niinimäki, S. The relationship between musculoskeletal stress markers and biomechanical properties of the humeral diaphysis. Am. J. Phys. Anthr. 2012, 147, 618–628. [Google Scholar] [CrossRef]
- Ibáñez-Gimeno, P.; De Esteban-Trivigno, S.; Jordana, X.; Manyosa, J.; Malgosa, A.; Galtés, I. Functional plasticity of the human humerus: Shape, rigidity, and muscular entheses. Am. J. Phys. Anthr. 2013, 150, 609–617. [Google Scholar] [CrossRef]
- Wang, X.; Liu, H.; Zhao, L.; Fei, C.; Feng, X.; Chen, S.; Wang, Y. Structural properties characterized by the film thickness and annealing temperature for La2O3 films grown by atomic layer deposition. Nanoscale Res. Lett. 2017, 12, 233. [Google Scholar] [CrossRef] [Green Version]
- Profico, A.; Bondioli, L.; Raia, P.; O’Higgins, P.; Marchi, D. Morphomap: An R package for long bone landmarking, cortical thickness, and cross-sectional geometry mapping. Am. J. Phys. Anthr. 2020, 174, 129–139. [Google Scholar] [CrossRef]
- Edgar, H.J.H.; Daneshvari-Berry, S.; Moes, E.; Adolphi, N.L.; Bridges, P.; Nolte, K.B. New Mexico Decedent Image Database; Office of the Medical Investigator; University of New Mexico: Albuquerque, NM, USA, 2020. [Google Scholar]
- Montoye, H.J. Energy costs of exercise and sport. In Nutrition in Sport; Maughan, R.J., Ed.; Blackwell Science Ltd: Oxford, UK, 2000; pp. 53–72. [Google Scholar]
- Ruff, C.B. Long bone articular and diaphyseal structure in old world monkeys and apes. I: Locomotor effects. Am. J. Phys. Anthr. 2002, 119, 305–342. [Google Scholar] [CrossRef]
- Borcard, D.; Legendre, P.; Drapeau, P. Partialling out the spatial component of ecological variation. Ecology 1992, 73, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Mardia, K.; Bookstein, F.; Moreton, I. Statistical assessment of bilateral symmetry of shapes. Biometrika 2000, 87, 285–300. [Google Scholar] [CrossRef]
- Perelle, I.B.; Ehrman, L. An international study of human handedness: The data. Behav. Genet. 1994, 24, 217–227. [Google Scholar] [CrossRef]
- McManus, I.C. The history and geography of human handedness. In Language Lateralization and Psychosis; Sommer, I.E.C., Kahn, R.S., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 37–58. [Google Scholar]
- Shaw, C.N. Is ‘hand preference’ coded in the hominin skeleton? An in-vivo study of bilateral morphological variation. J. Hum. Evol. 2011, 61, 480–487. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Argument | Definition |
|---|---|
| mshape1 | First long bone processed with morphomapShape |
| mshape2 | Second long bone processed with morphomapShape |
| standandize | If TRUE, the matrices of cortical thickness are standardized using the average biomechanical length between sides. |
| plot | If TRUE, the map of cortical thickness asymmetry is returned. |
| type | Defines the method to calculate the differences in cortical thickness between the two long bones: “diff” a map of arithmetic difference between reference and target is computed; “onMean” the morphometric map of asymmetry is defined by computing the differences from the mean for each long bone; “relChange” the morphometric map is computed by calculating the relative change in cortical thickness, expressed as percentage difference between reference and target long bones |
| reference | If set to 1, mshape1 is defined as reference; if set on 2, mshape2 is defined as reference. |
| rem.out | If TRUE, outliers are removed from the matrices of cortical thickness. |
| scale | If TRUE, the matrices of cortical thickness are scaled from 0 to 1. |
| gamMap | If TRUE, gam smoothing is applied. |
| ID | Sex | Age | Weight | Height | Occupation | JL | JR | JLAT% |
|---|---|---|---|---|---|---|---|---|
| 100221 | male | 34 | 91 | 188 | D | 2.84 | 3.60 | 5.90 |
| 101358 | male | 21 | 70 | 168 | B | 1.50 | 1.57 | 1.07 |
| 101510 | male | 26 | 86 | 195 | A | 2.39 | 2.96 | 5.33 |
| 102253 | male | 46 | 100 | 183 | D | 1.62 | 1.92 | 4.26 |
| 102436 | male | 37 | 83 | 183 | D | 3.10 | 3.22 | 0.91 |
| 102602 | male | 32 | 86 | 193 | B | 2.26 | 2.40 | 1.56 |
| 103530 | male | 22 | 73 | 168 | B | 1.51 | 1.72 | 3.26 |
| 103862 | male | 31 | 91 | 191 | A | 2.11 | 2.26 | 1.76 |
| 104373 | male | 34 | 86 | 170 | B | 2.47 | 2.56 | 0.86 |
| 108039 | male | 38 | 82 | 188 | A | 1.65 | 1.92 | 3.77 |
| 114405 | male | 27 | 79 | 178 | B | 1.62 | 1.97 | 4.81 |
| 116546 | male | 25 | 86 | 184 | A | 2.82 | 2.43 | 3.72 |
| 116833 | male | 45 | 109 | 183 | A | 2.40 | 2.61 | 2.08 |
| 117662 | female | 24 | 68 | 173 | A | 1.29 | 1.50 | 3.85 |
| 118646 | male | 26 | 84 | 180 | A | 1.92 | 2.84 | 9.63 |
| 121289 | male | 25 | 102 | 184 | B | 2.74 | 2.98 | 2.10 |
| 123096 | male | 24 | 75 | 184 | B | 2.81 | 3.14 | 2.78 |
| 123240 | male | 31 | 82 | 180 | D | 1.64 | 2.05 | 5.61 |
| 125527 | female | 26 | 50 | 157 | D | 0.69 | 0.79 | 3.35 |
| 127137 | female | 54 | 59 | 158 | D | 1.38 | 1.32 | 1.20 |
| 129131 | male | 33 | 79 | 173 | A | 1.80 | 1.87 | 0.93 |
| 129352 | male | 30 | 82 | 180 | A | 1.56 | 1.86 | 4.29 |
| 130388 | male | 37 | 68 | 168 | A | 1.67 | 1.82 | 2.16 |
| 130964 | male | 25 | 68 | 175 | D | 2.87 | 3.42 | 4.42 |
| 132233 | male | 29 | 80 | 166 | B | 1.77 | 2.13 | 4.64 |
| 132433 | male | 27 | 70 | 185 | D | 2.06 | 2.30 | 2.72 |
| 139871 | female | 49 | 61 | 157 | B | 1.49 | 1.56 | 1.27 |
| 140368 | male | 34 | 80 | 185 | B | 3.17 | 3.83 | 4.74 |
| 141318 | male | 32 | 77 | 178 | B | 2.04 | 2.24 | 2.32 |
| 143365 | female | 47 | 72 | 165 | A | 1.16 | 1.10 | 1.32 |
| 143984 | male | 30 | 91 | 183 | D | 2.51 | 2.63 | 1.17 |
| 144071 | female | 35 | 57 | 160 | A | 1.04 | 1.08 | 0.94 |
| 144977 | male | 24 | 75 | 175 | D | 2.06 | 2.24 | 2.09 |
| 146626 | male | 27 | 89 | 163 | A | 0.98 | 1.16 | 4.24 |
| 147949 | male | 29 | 91 | 178 | A | 1.82 | 2.63 | 9.15 |
| 150608 | male | 34 | 63 | 158 | B | 1.81 | 2.01 | 2.64 |
| 152567 | male | 24 | 82 | 185 | A | 2.46 | 2.74 | 2.70 |
| 156886 | male | 29 | 77 | 178 | B | 1.78 | 1.97 | 2.51 |
| 158402 | male | 35 | 109 | 185 | B | 2.32 | 2.37 | 0.58 |
| 162065 | male | 28 | 64 | 170 | B | 1.56 | 1.88 | 4.76 |
| 166116 | female | 36 | 56 | 162 | B | 1.07 | 1.17 | 2.22 |
| 171170 | male | 32 | 106 | 182 | A | 2.96 | 3.75 | 5.90 |
| 171479 | male | 27 | 100 | 185 | D | 2.50 | 2.82 | 3.06 |
| 173218 | male | 22 | 84 | 163 | D | 1.95 | 2.06 | 1.36 |
| 175725 | male | 28 | 86 | 196 | B | 3.12 | 3.46 | 2.54 |
| 176660 | female | 24 | 68 | 173 | A | 0.79 | 0.84 | 1.47 |
| 177679 | male | 38 | 77 | 168 | A | 2.75 | 2.82 | 0.68 |
| 178078 | male | 31 | 73 | 168 | B | 2.10 | 2.31 | 2.32 |
| 180030 | male | 40 | 82 | 180 | A | 2.06 | 2.33 | 3.05 |
| 188902 | female | 26 | 77 | 165 | D | 1.25 | 1.12 | 2.82 |
| 190756 | female | 24 | 48 | 162 | D | 0.84 | 0.98 | 3.82 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Profico, A.; Zeppilli, C.; Micarelli, I.; Mondanaro, A.; Raia, P.; Marchi, D.; Manzi, G.; O’Higgins, P. Morphometric Maps of Bilateral Asymmetry in the Human Humerus: An Implementation in the R Package Morphomap. Symmetry 2021, 13, 1711. https://doi.org/10.3390/sym13091711
Profico A, Zeppilli C, Micarelli I, Mondanaro A, Raia P, Marchi D, Manzi G, O’Higgins P. Morphometric Maps of Bilateral Asymmetry in the Human Humerus: An Implementation in the R Package Morphomap. Symmetry. 2021; 13(9):1711. https://doi.org/10.3390/sym13091711
Chicago/Turabian StyleProfico, Antonio, Carlotta Zeppilli, Ileana Micarelli, Alessandro Mondanaro, Pasquale Raia, Damiano Marchi, Giorgio Manzi, and Paul O’Higgins. 2021. "Morphometric Maps of Bilateral Asymmetry in the Human Humerus: An Implementation in the R Package Morphomap" Symmetry 13, no. 9: 1711. https://doi.org/10.3390/sym13091711