
Characterization of a Primate Blood-Brain Barrier Co-Culture Model Prepared from Primary Brain Endothelial Cells, Pericytes and Astrocytes

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Materials and Reagents
2.3. Primary Cell Cultures
2.4. Construction of the BBB Model
2.5. Immunohistochemistry
2.6. Reverse Transcription Polymerase CHAIN Reaction
2.7. Evaluation of the Barrier Integrity of the BBB Models
2.8. Functional Assay for Efflux Transport
2.9. Drug Permeability Experiments
2.10. Western Blotting
2.11. Statistical Analysis
3. Results
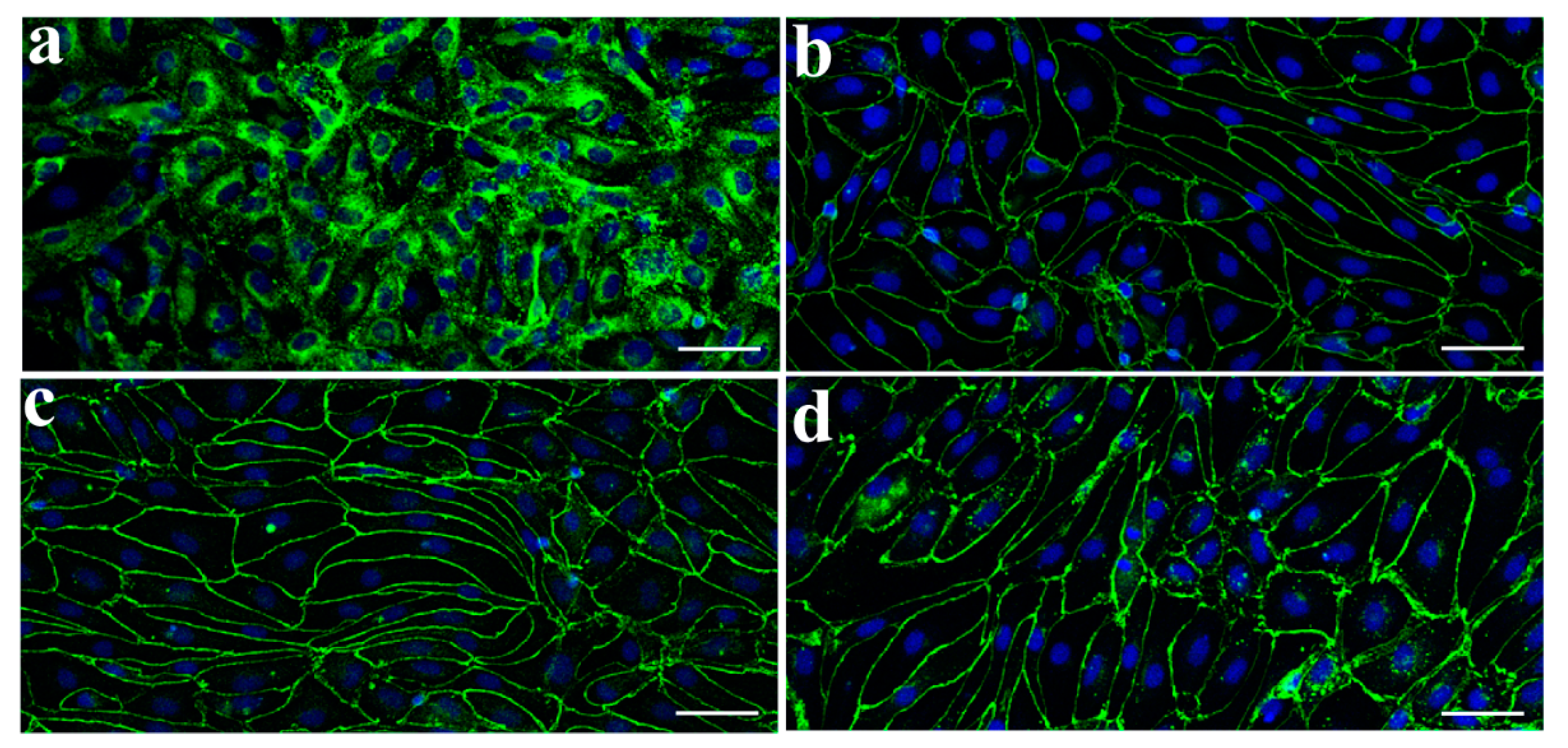
3.1. Characterization of Monkey Brain Endothelial Cells
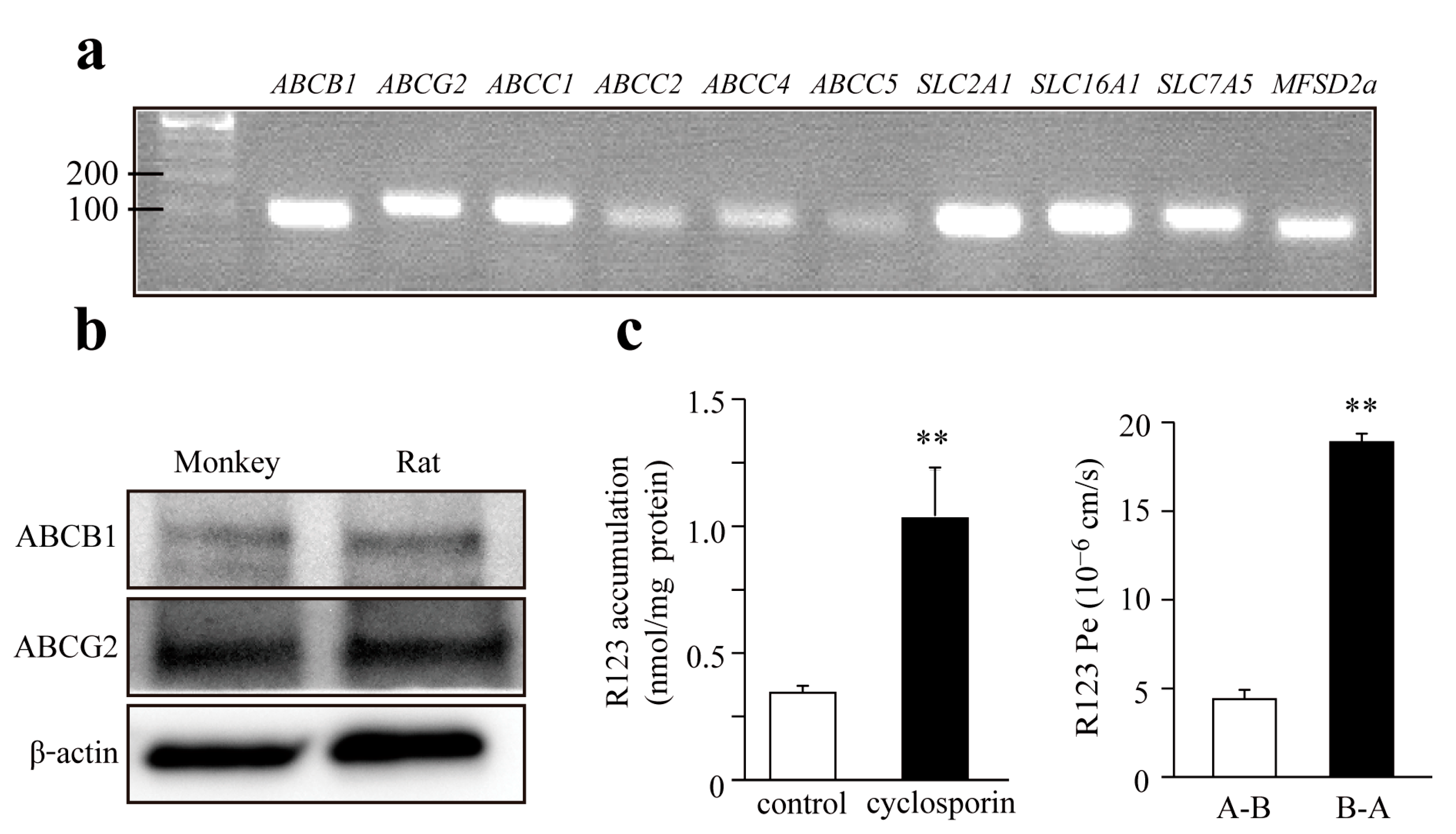
3.2. Expression of Transporters in Monkey Brain Endothelial Cells
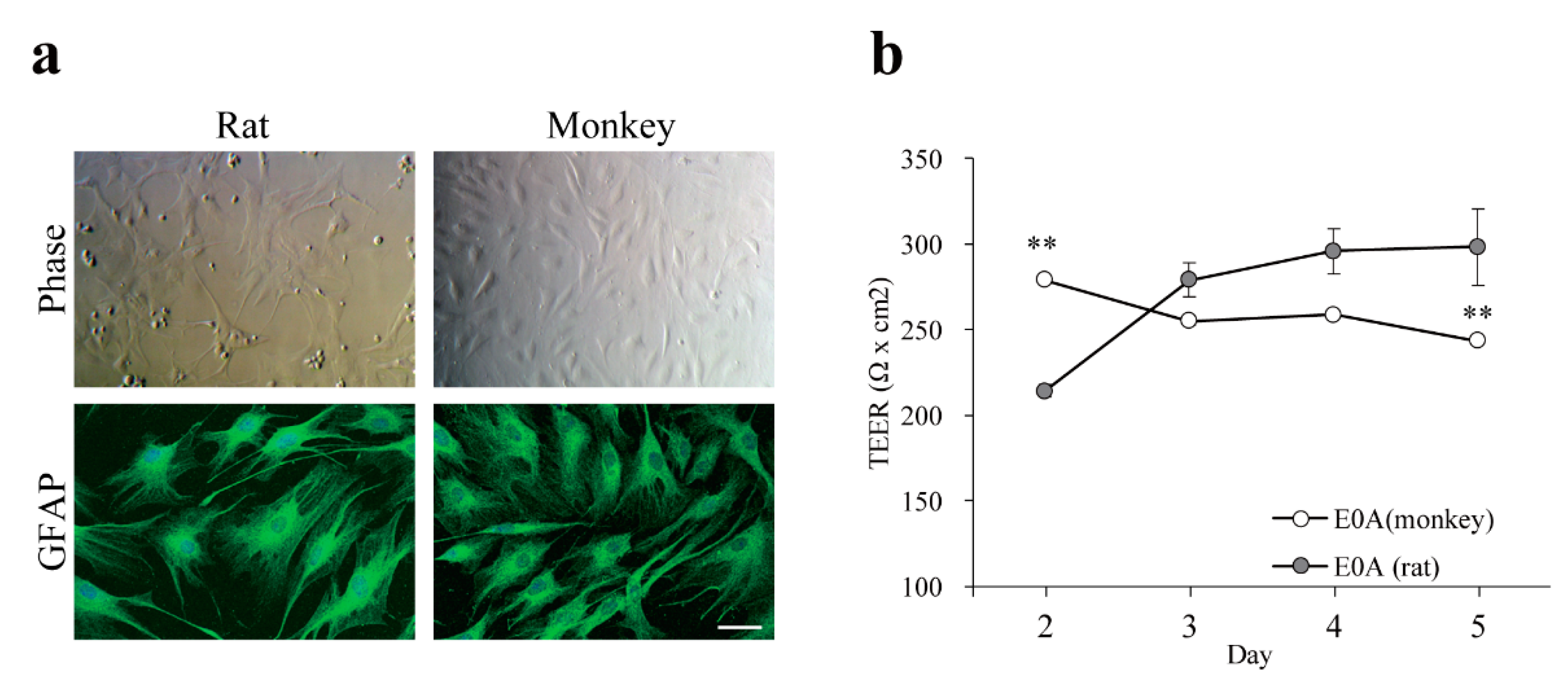
3.3. Effect of Astrocytes on Barrier Function of Monkey Brain Endothelial Cells
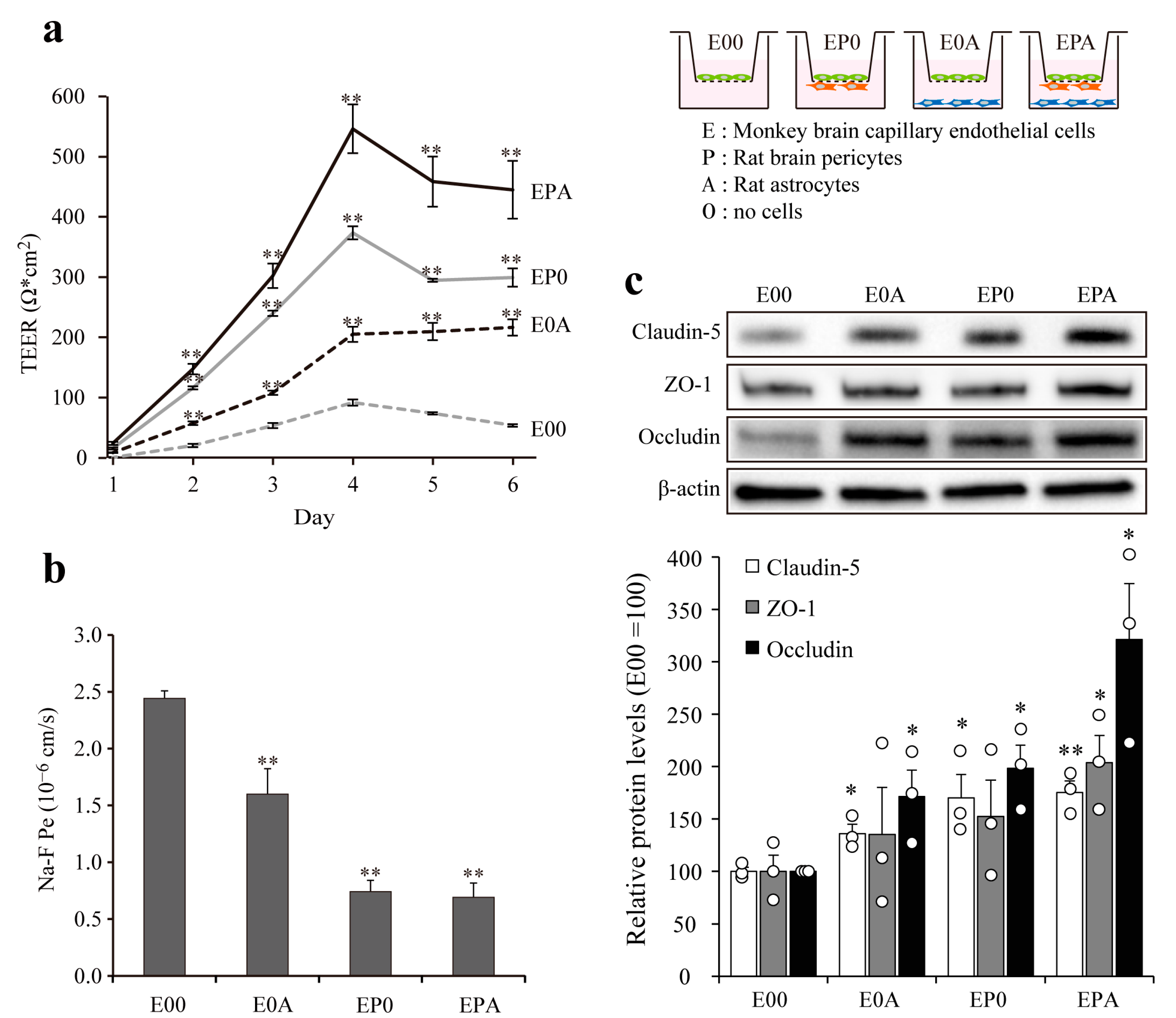
3.4. Barrier Integrity of Different BBB Models
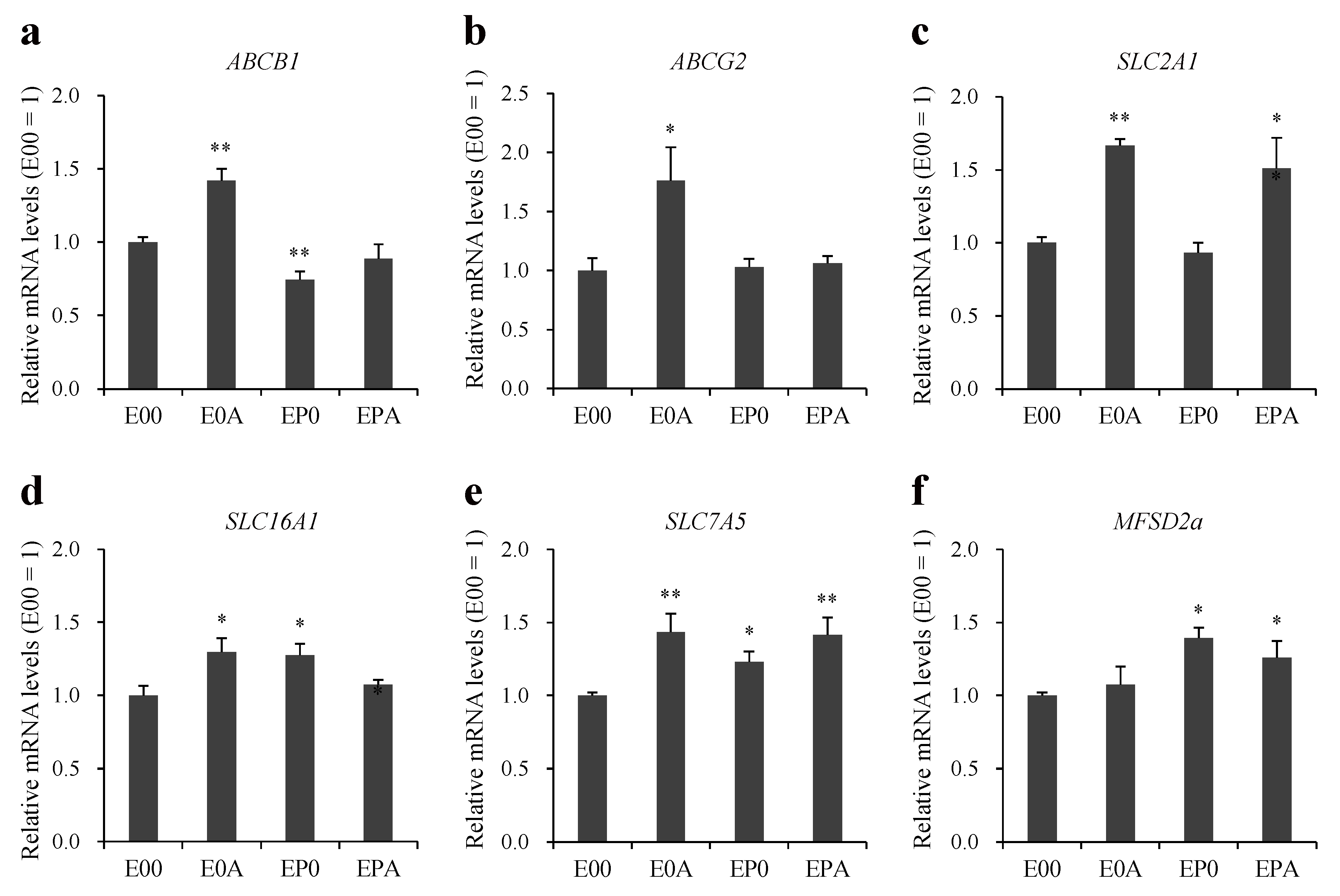
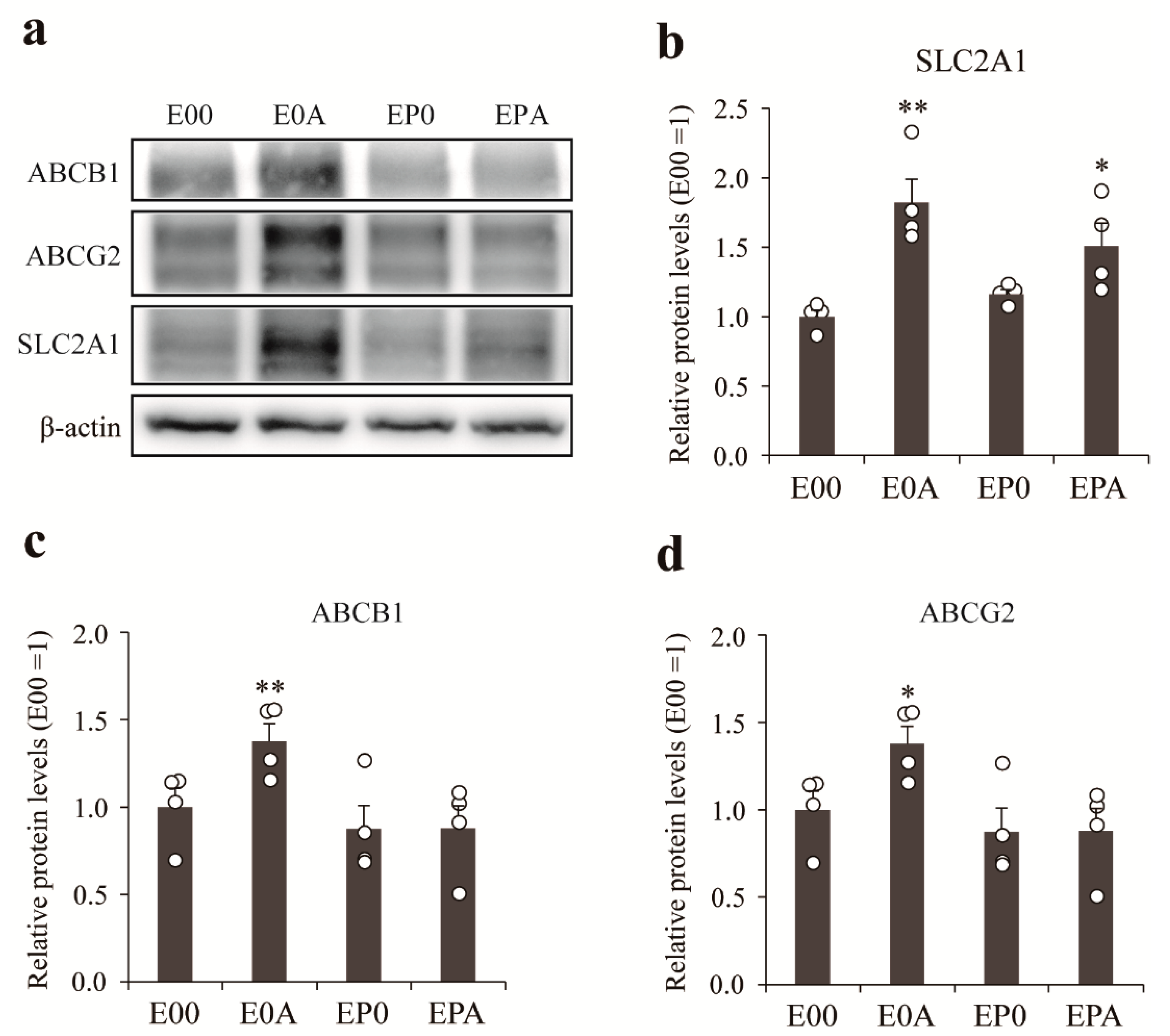
3.5. Expression of Transporters on Different BBB Models
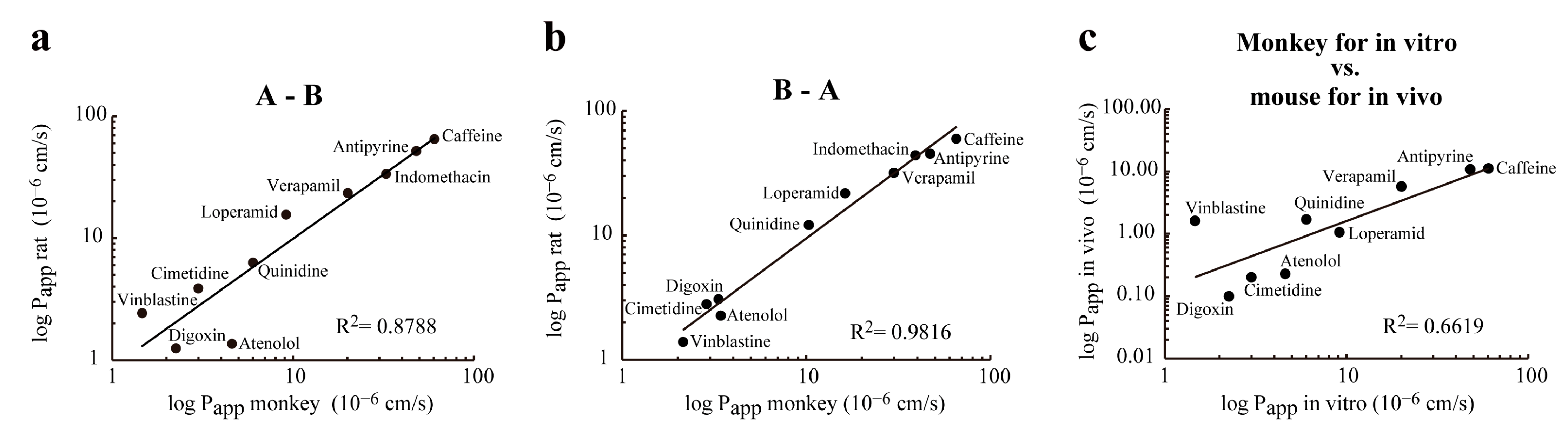
3.6. Correlation between Permeability of Drugs Tested in Rat and Monkey BBB Co-Culture Models, and Mouse In Vivo Model
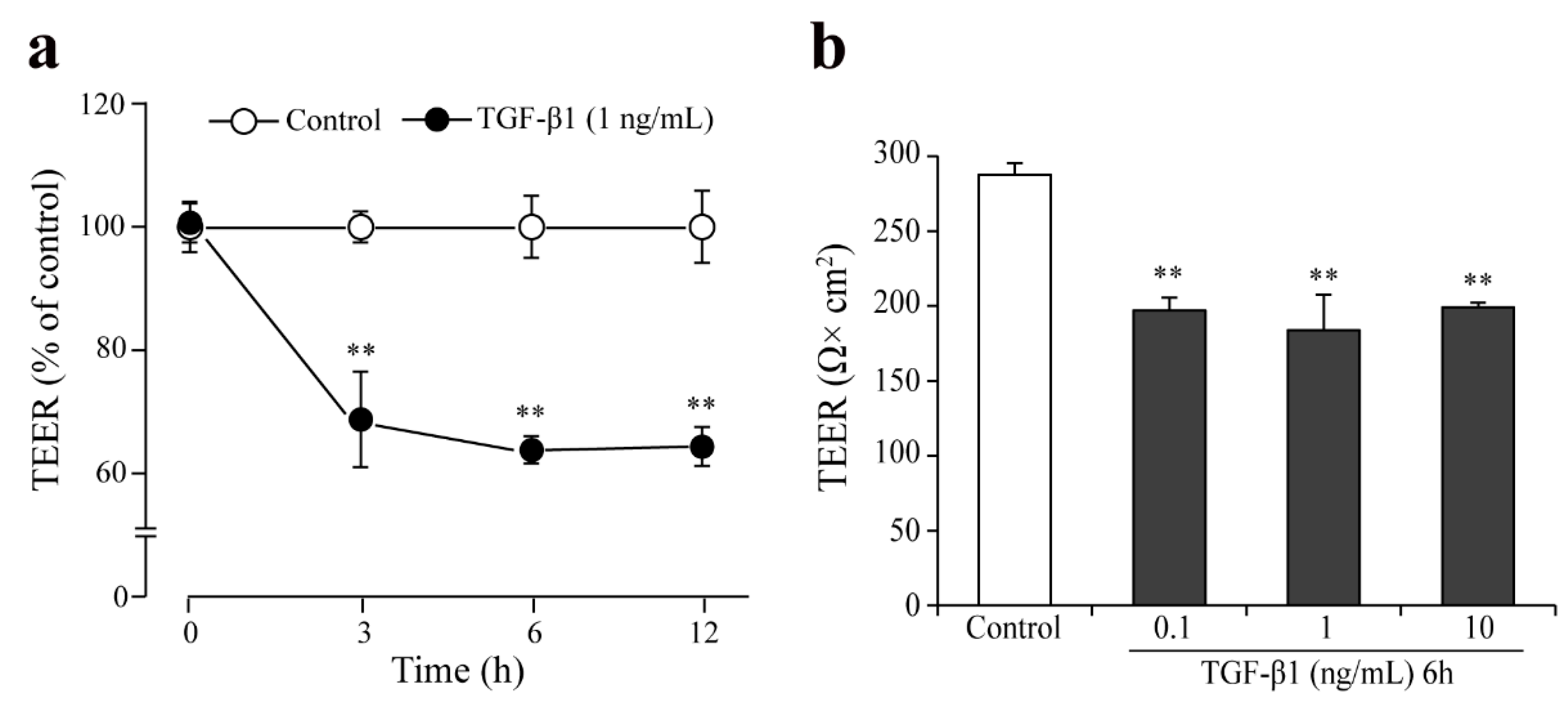
3.7. Effect of TGF-β1 on Barrier Integrity in the Monkey Co-Culture BBB Model
4. Discussion
4.1. Characterization of the Primary Triple-Co-Culture Primate BBB Model
4.2. Application of the Primary Triple-Co-Culture Primate BBB Model
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Deli, M.A.; Ábrahám, C.S.; Kataoka, Y.; Niwa, M. Permeability studies on in vitro blood-brain barrier models: Physiology, pathology, and pharmacology. Cell. Mol. Neurobiol. 2005, 25, 59–127. [Google Scholar] [CrossRef]
- Cecchelli, R.; Berezowski, V.; Lundquist, S.; Culot, M.; Renftel, M.; Dehouck, M.P.; Fenart, L. Modelling of the blood-brain barrier in drug discovery and development. Nat. Rev. Drug Discov. 2007, 6, 650–661. [Google Scholar] [CrossRef]
- Helms, H.C.; Abbott, N.J.; Burek, M.; Cecchelli, R.; Couraud, P.O.; Deli, M.A.; Förster, C.; Galla, H.J.; Romero, I.A.; Shusta, E.V.; et al. In vitro models of the blood-brain barrier: An overview of commonly used brain endothelial cell culture models and guidelines for their use. J. Cereb. Blood Flow Metab. 2016, 36, 862–890. [Google Scholar] [CrossRef]
- Tóth, A.; Veszelka, S.; Nakagawa, S.; Niwa, M.; Deli, M.A. Patented in vitro blood-brain barrier models in CNS drug discovery. Recent Pat. CNS Drug Discov. 2011, 6, 107–118. [Google Scholar] [CrossRef]
- Avdeef, A.; Deli, M.A.; Neuhaus, W. In vitro assays for assessing BBB permeability: Artificial membrane and cell culture models. In Blood-Brain Barrier in Drug Discovery: Optimizing Brain Exposure of CNS Drugs and Minimizing Brain Side Effects; Di, L., Kerns, E.H., Eds.; John Wiley & Sons, Inc: Hoboken, NJ, USA, 2015; pp. 188–237. ISBN 978-1-118-78835-6. [Google Scholar] [CrossRef]
- Veszelka, S.; Tóth, A.; Walter, F.R.; Tóth, A.E.; Gróf, I.; Mészáros, M.; Bocsik, A.; Hellinger, E.; Vastag, M.; Rákhely, G.; et al. Comparison of a rat primary cell-based blood-brain barrier model with epithelial and brain endothelial cell lines: Gene expression and drug transport. Front. Mol. Neurosci. 2018, 11, 166. [Google Scholar] [CrossRef] [PubMed]
- Banks, W.A. From blood-brain barrier to blood-brain interface: New opportunities for CNS drug delivery. Nat. Rev. Drug Discov. 2016, 15, 275–292. [Google Scholar] [CrossRef]
- Nakagawa, S.; Deli, M.A.; Nakao, S.; Honda, M.; Hayashi, K.; Nakaoke, R.; Kataoka, Y.; Niwa, M. Pericytes from brain microvessels strengthen the barrier integrity in primary cultures of rat brain endothelial cells. Cell. Mol. Neurobiol. 2007, 27, 687–694. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, S.; Deli, M.A.; Kawaguchi, H.; Shimizudani, T.; Shimono, T.; Kittel, Á.; Tanaka, K.; Niwa, M. A new blood-brain barrier model using primary rat brain endothelial cells, pericytes and astrocytes. Neurochem. Int. 2009, 54, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Sweeney, M.D.; Zhao, Z.; Montagne, A.; Nelson, A.R.; Zlokovic, B.V. Blood-brain barrier: From physiology to disease and back. Physiol. Rev. 2019, 99, 21–78. [Google Scholar] [CrossRef]
- Ito, K.; Uchida, Y.; Ohtsuki, S.; Aizawa, S.; Kawakami, H.; Katsukura, Y.; Kamiie, J.; Terasaki, T. Quantitative membrane protein expression at the blood-brain barrier of adult and younger cynomolgus monkeys. J. Pharm. Sci. 2011, 100, 3939–3950. [Google Scholar] [CrossRef]
- Ohtsuki, S.; Uchida, Y.; Kubo, Y.; Terasaki, T. Quantitative targeted absolute proteomics-based ADME research as a new path to drug discovery and development: Methodology, advantages, strategy, and prospects. J. Pharm. Sci. 2011, 100, 3547–3559. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.M.; Houghton, S.; Magdeldin, T.; Durán, J.G.B.; Minotti, A.P.; Snead, A.; Sproul, A.; Nguyen, D.T.; Xiang, J.; Fine, H.A.; et al. Pluripotent stem cell-derived epithelium misidentified as brain microvascular endothelium requires ETS factors to acquire vascular fate. Proc. Natl. Acad. Sci. USA 2021, 118, e2016950118. [Google Scholar] [CrossRef]
- Perrière, N.; Demeuse, P.; Garcia, E.; Regina, A.; Debray, M.; Andreux, J.P.; Couvreur, P.; Scherrmann, J.M.; Temsamani, J.; Couraud, P.O.; et al. Puromycin-based purification of rat brain capillary endothelial cell cultures. Effect on the expression of blood-brain barrier-specific properties. J. Neurochem. 2005, 93, 279–289. [Google Scholar] [CrossRef]
- Hellinger, É.; Veszelka, S.; Tóth, A.E.; Walter, F.; Kittel, Á.; Bakk, M.L.; Tihanyi, K.; Háda, V.; Nakagawa, S.; Duy, T.D.H.; et al. Comparison of brain capillary endothelial cell-based and epithelial (MDCK-MDR1, Caco-2, and VB-Caco-2) cell-based surrogate blood-brain barrier penetration models. Eur. J. Pharm. Biopharm. 2012, 82, 340–351. [Google Scholar] [CrossRef]
- Campos-Bedolla, P.; Walter, F.R.; Veszelka, S.; Deli, M.A. Role of the blood-brain barrier in the nutrition of the central nervous system. Arch. Med. Res. 2014, 45, 610–638. [Google Scholar] [CrossRef]
- Morofuji, Y.; Nakagawa, S. Drug development for central nervous system diseases using in vitro blood-brain barrier models and drug repositioning. Curr. Pharm. Des. 2020, 26, 1466–1485. [Google Scholar] [CrossRef]
- Syvänen, S.; Lindhe, O.; Palner, M.; Kornum, B.R.; Rahman, O.; Långström, B.; Knudsen, G.M.; Hammarlund-Udenaes, M. Species differences in blood-brain barrier transport of three positron emission tomography radioligands with emphasis on P-glycoprotein transport. Drug Metab. Dispos. 2009, 37, 635–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves, C.; Do, T.M.; Cegarra, C.; Roudières, V.; Tolou, S.; Thill, G.; Rocher, C.; Didier, M.; Lesuisse, D. Non-human primate blood-brain barrier and in vitro brain endothelium: From transcriptome to the establishment of a new model. Pharmaceutics 2020, 12, 967. [Google Scholar] [CrossRef]
- MacLean, A.G.; Orandle, M.S.; Alvarez, X.; Williams, K.C.; Lackner, A.A. Rhesus macaque brain microvessel endothelial cells behave in a manner phenotypically distinct from umbilical vein endothelial cells. J. Neuroimmunol. 2001, 118, 223–232. [Google Scholar] [CrossRef]
- MacLean, A.G.; Orandle, M.S.; MacKey, J.; Williams, K.C.; Alvarez, X.; Lackner, A.A. Characterization of an in vitro rhesus macaque blood-brain barrier. J. Neuroimmunol. 2002, 131, 98–103. [Google Scholar] [CrossRef] [Green Version]
- MacLean, A.G.; Rasmussen, T.A.; Bieniemy, D.N.; Alvarez, X.; Lackner, A.A. SIV-induced activation of the blood-brain barrier requires cell-associated virus and is not restricted to endothelial cell activation. J. Med. Primatol. 2004, 33, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Sansing, H.A.; Renner, N.A.; MacLean, A.G. An inverted blood-brain barrier model that permits interactions between glia and inflammatory stimuli. J. Neurosci. Methods 2012, 207, 91–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, T.; Schwartz, J.P. Gene expression patterns in in vivo normal adult astrocytes compared with cultured neonatal and normal adult astrocytes. Neurochem. Int. 2004, 45, 203–242. [Google Scholar] [CrossRef] [PubMed]
- Toyoda, K.; Tanaka, K.; Nakagawa, S.; Thuy, D.H.; Ujifuku, K.; Kamada, K.; Hayashi, K.; Matsuo, T.; Nagata, I.; Niwa, M. Initial contact of glioblastoma cells with existing normal brain endothelial cells strengthen the barrier function via fibroblast growth factor 2 secretion: A new in vitro blood-brain barrier model. Cell. Mol. Neurobiol. 2013, 33, 489–501. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, L.B.; Burkhart, A.; Moos, T. A triple culture model of the blood-brain barrier using porcine brain endothelial cells, astrocytes and pericytes. PLoS ONE 2015, 10, e0134765. [Google Scholar] [CrossRef] [Green Version]
- Saaby, L.; Helms, H.C.; Brodin, B. IPEC-J2 MDR1, a Novel High-Resistance Cell Line with Functional Expression of Human P-glycoprotein (ABCB1) for Drug Screening Studies. Mol. Pharm. 2016, 13, 640–652. [Google Scholar] [CrossRef]
- Nakagawa, S.; Deli, M.A.; Thuy, D.H.D.; Sagara, M.; So, G.; Yamada, N.; Tatsumi, R.; Tanaka, K.; Niwa, M. An in vitro monkey BBB model. J. Pharmacol. Sci. 2010, 112, 91. [Google Scholar]
- Watanabe, S.; Nakagawa, S.; Deli, M.A.; Izawa, K.; Aruga, J.; Niwa, M. A new primate in vitro blood-brain barrier model using primary monkey capillary cells. J. Pharmacol. Sci. 2017, 133, S236. [Google Scholar]
- BBB Kit™ (MBT-24H)/(MBT-24F). Available online: https://www.pharmacocell.co.jp/en/products/mbt24h_e.html (accessed on 16 June 2021).
- Salcedo, G.P.M.; Amano, M.; Yashchuk, S.; Mizuno, A.; Das, D.; Ghosh, A.K.; Mitsuya, H. GRL-04810 and GRL-05010, difluoride-containing nonpeptidic HIV-1 protease inhibitors (PIs) that inhibit the replication of multi-PI-resistant HIV-1 in vitro and possess favorable lipophilicity that may allow blood-brain barrier penetration. Antimicrob. Agents Chemother. 2013, 57, 6110–6121. [Google Scholar] [CrossRef] [Green Version]
- Amano, M.; Tojo, Y.; Salcedo-Gómez, P.M.; Parham, G.L.; Nyalapatla, P.R.; Das, D.; Ghosh, A.K.; Mitsuya, H. A novel tricyclic ligand-containing nonpeptidic HIV-1 protease inhibitor, GRL-0739, effectively inhibits the replication of multidrug-resistant HIV-1 variants and has a desirable central nervous system penetration property in vitro. Antimicrob. Agents Chemother. 2015, 59, 2625–2635. [Google Scholar] [CrossRef] [Green Version]
- Tominaga, N.; Kosaka, N.; Ono, M.; Katsuda, T.; Yoshioka, Y.; Tamura, K.; Lötvall, J.; Nakagama, H.; Ochiya, T. Brain metastatic cancer cells release microRNA-181c-containing extracellular vesicles capable of destructing blood-brain barrier. Nat. Commun. 2015, 6, 6716. [Google Scholar] [CrossRef] [Green Version]
- Amano, M.; Salcedo-Gómez, P.M.; Zhao, R.; Yedidi, R.S.; Das, D.; Bulut, H.; Delino, N.S.; Sheri, V.R.; Ghosh, A.K.; Mitsuya, H. A modified P1 moiety enhances in vitro antiviral activity against various multidrug-resistant HIV-1 variants and in vitro central nervous system penetration properties of a novel nonpeptidic protease inhibitor, GRL-10413. Antimicrob. Agents Chemother. 2016, 60, 7046–7059. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, Y.; Shirakura, K.; Okada, Y.; Takeda, H.; Endo, K.; Tamura, M.; Watari, A.; Sadamura, Y.; Sawasaki, T.; Doi, T.; et al. Claudin-5-binders enhance permeation of solutes across the blood-brain barrier in a mammalian model. J. Pharmacol. Exp. Ther. 2017, 363, 275–283. [Google Scholar] [CrossRef]
- Amano, M.; Salcedo-Gómez, P.M.; Yedidi, R.S.; Zhao, R.; Hayashi, H.; Hasegawa, K.; Nakamura, T.; Martyr, C.D.; Ghosh, A.K.; Mitsuya, H. Novel central nervous system (CNS)-targeting protease inhibitors for drug-resistant HIV infection and HIV-associated CNS complications. Antimicrob. Agents Chemother. 2019, 63, e00466-19. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Liang, M.; Munesue, S.; Deguchi, K.; Harashima, A.; Furuhara, K.; Yuhi, T.; Zhong, J.; Akther, S.; GGoto, H.; et al. Vascular RAGE transports oxytocin into the brain to elicit its maternal bonding behaviour in mice. Commun. Biol. 2019, 2, 76. [Google Scholar] [CrossRef]
- Kimura, A.M.; Tsuji, M.; Yasumoto, T.; Mori, Y.; Oguchi, T.; Tsuji, Y.; Umino, M.; Umino, A.; Nishikawa, T.; Nakamura, S.; et al. Myricetin prevents high molecular weight Aβ1-42 oligomer-induced neurotoxicity through antioxidant effects in cell membranes and mitochondria. Free Radic. Biol. Med. 2021, 171, 232–244. [Google Scholar] [CrossRef]
- Mochizuki, T.; Mizuno, T.; Kurosawa, T.; Yamaguchi, T.; Higuchi, K.; Tega, Y.; Nozaki, Y.; Kawabata, K.; Deguchi, Y.; Kusuhara, H. Functional investigation of solute carrier family 35, member F2, in three cellular models of the primate blood-brain barrier. Drug Metab. Dispos. 2021, 49, 3–11. [Google Scholar] [CrossRef]
- Tachibana, K.; Hashimoto, Y.; Shirakura, K.; Okada, Y.; Hirayama, R.; Iwashita, Y.; Nishino, I.; Ago, Y.; Takeda, H.; Kuniyasu, H.; et al. Safety and efficacy of an anti-claudin-5 monoclonal antibody to increase blood-brain barrier permeability for drug delivery to the brain in a non-human primate. J. Control Release 2021, 336, 105–111. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | Size (bp) |
|---|---|---|
| ABCB1 (P-gp) | F: GGCCTAAAGCCGAACACATTG R: CTGAAGCACTGGGATGTCCA | 90 |
| ABCG2 (BCRP) | F: GCCACGGAGATCATAGAGCC R: TCACCCCCGGAAAGTTGATG | 125 |
| ABCC1 (MRP-1) | F: CAAGGGATTGCCGTGTTTGG R: AAGAAGCTCATGGGTGACCG | 116 |
| ABCC2 (MRP-2) | F: GCACAAGCAACTGCTCAACA R: CCGTGGAAATATCACCGGCA | 104 |
| ABCC4 (MRP-4) | F: TCGCAATACCCTTGGTTCCC R: CACTGGGCTCCGAGTTGTAG | 110 |
| ABCC5 (MRP-5) | F: CTTTGTCAAGGGCACACTGC R: CCTGTGGGGGTTGTGTCAAA | 102 |
| SLC2A1 (GLUT1) | F: GAACTCTTCAGCCAGGGTCC R: GGACCACATAGTTGCTCCAC | 116 |
| SLC16A1 (MCT1) | F: ACAAGTAAACGAGGCAGCGA R: ACAAATATCGTTATAAGCGCGGA | 123 |
| SLC7A5 (LAT1) | F: CGTGAACTGCTACAGCGTGA R: TTGGACACATCACCCTTCCC | 126 |
| MFSD2a | F: GCCCAGGTGAAGAAAGAACC R: CACAGCCTGTCACCTGGTAG | 103 |
| GAPDH | F: CTCAAGATCGTCAGCAACGC R: TCTTCTGGGTGGCAGTGATG | 130 |
| Compound | Primary Transport Mechanism | Monkey BBB | Rat BBB * | Mouse In Vivo * | ||||
|---|---|---|---|---|---|---|---|---|
| Papp A-B (×10−6 cm/s) | Papp B-A (×10−6 cm/s) | EffluxRatio | Papp A-B (×10−6 cm/s) | Papp B-A (×10−6 cm/s) | EffluxRatio | Papp (×10−6 cm/s) | ||
| Caffeine | PT/CMT | 60.53 | 65.25 | 1.08 | 64.95 | 59.91 | 0.92 | 11.11 |
| Antipyrin | PT | 48.06 | 47.07 | 0.98 | 51.78 | 45.38 | 0.88 | 10.75 |
| Indomethacin | PT | 32.70 | 39.11 | 1.20 | 33.63 | 44.07 | 1.31 | 0.13 |
| Verapamil | PT/E | 20.10 | 29.86 | 1.49 | 23.38 | 31.82 | 1.38 | 5.69 |
| Loperamide | PT/E | 9.16 | 16.26 | 1.78 | 15.57 | 21.76 | 1.47 | 1.05 |
| Quinidine | PT/E | 6.01 | 10.33 | 1.74 | 6.28 | 12.11 | 1.93 | 1.69 |
| Cimetidine | E | 3.00 | 2.88 | 0.97 | 3.86 | 2.80 | 0.75 | 0.20 |
| Digoxin | E | 2.25 | 3.34 | 1.49 | 1.25 | 3.07 | 2.47 | 0.10 |
| Vinblastine | E | 1.47 | 2.14 | 1.46 | 2.42 | 1.39 | 0.69 | 1.43 |
| Atenolol | PP/E | 4.60 | 3.43 | 0.83 | 1.36 | 2.26 | 1.71 | 0.23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watanabe, D.; Nakagawa, S.; Morofuji, Y.; Tóth, A.E.; Vastag, M.; Aruga, J.; Niwa, M.; Deli, M.A. Characterization of a Primate Blood-Brain Barrier Co-Culture Model Prepared from Primary Brain Endothelial Cells, Pericytes and Astrocytes. Pharmaceutics 2021, 13, 1484. https://doi.org/10.3390/pharmaceutics13091484
Watanabe D, Nakagawa S, Morofuji Y, Tóth AE, Vastag M, Aruga J, Niwa M, Deli MA. Characterization of a Primate Blood-Brain Barrier Co-Culture Model Prepared from Primary Brain Endothelial Cells, Pericytes and Astrocytes. Pharmaceutics. 2021; 13(9):1484. https://doi.org/10.3390/pharmaceutics13091484
Chicago/Turabian StyleWatanabe, Daisuke, Shinsuke Nakagawa, Yoichi Morofuji, Andrea E. Tóth, Monika Vastag, Jun Aruga, Masami Niwa, and Mária A. Deli. 2021. "Characterization of a Primate Blood-Brain Barrier Co-Culture Model Prepared from Primary Brain Endothelial Cells, Pericytes and Astrocytes" Pharmaceutics 13, no. 9: 1484. https://doi.org/10.3390/pharmaceutics13091484