
Selenite Downregulates STAT3 Expression and Provokes Lymphocytosis in the Liver of Chronically Exposed Syrian Golden Hamsters

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
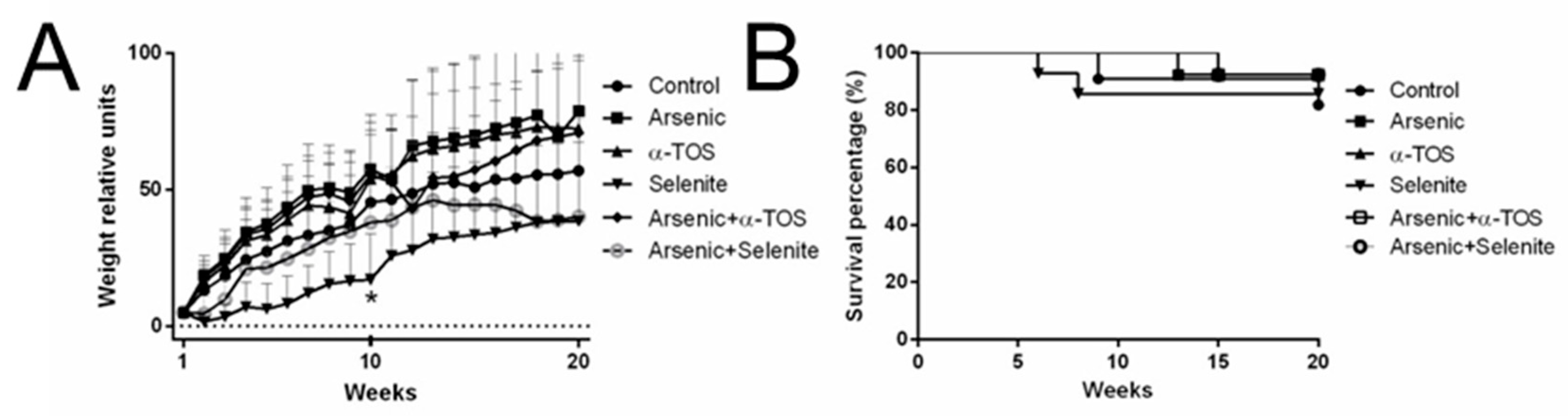
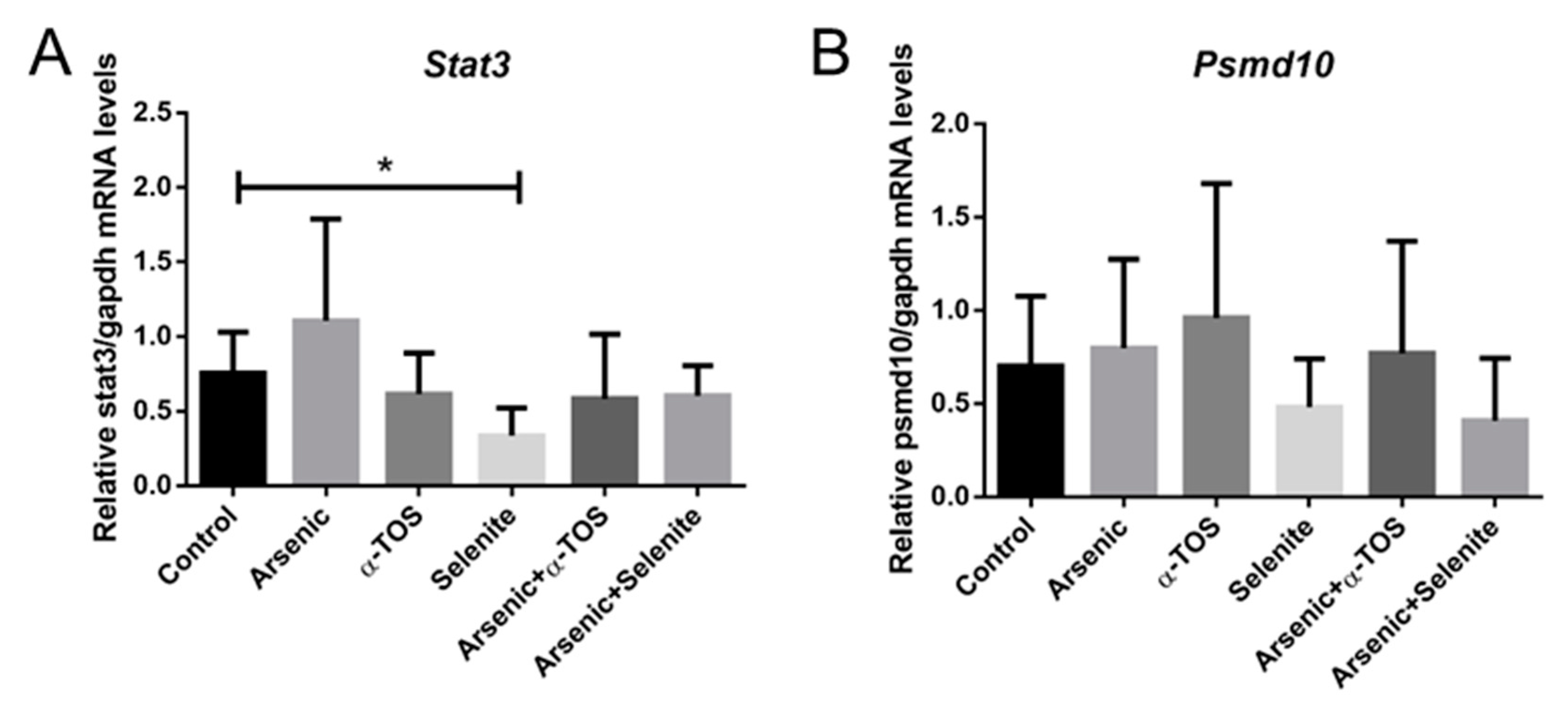
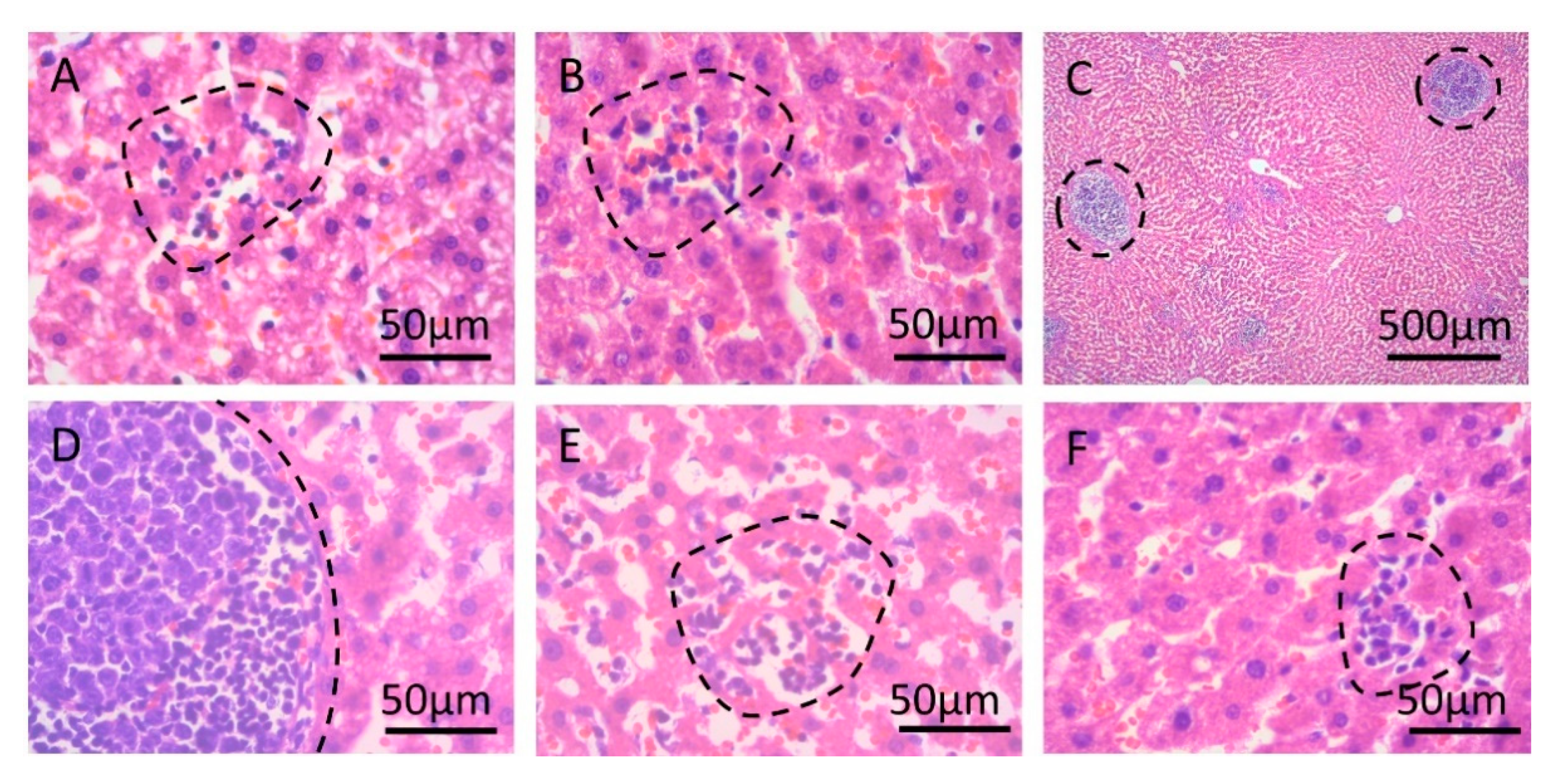
2. Results
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animals and Treatments
4.3. Measurement of Arsenic and Arsenic Species
4.4. RNA Extraction and cDNA Synthesis
4.5. SYBR Green Quantitative Real-Time Reverse Transcriptase (RT)-PCR
4.6. Hematoxylin and Eosin Staining
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Cullen, W.R. Is Arsenic an Aphrodisiac?: The Sociochemistry of an Element; RSC Publishing: Cambridge, UK, 2008. [Google Scholar]
- Bjorklund, G.; Oliinyk, P.; Lysiuk, R.; Rahaman, M.S.; Antonyak, H.; Lozynska, I.; Lenchyk, L.; Peana, M. Arsenic intoxication: General aspects and chelating agents. Arch. Toxicol. 2020, 94, 1879–1897. [Google Scholar] [CrossRef]
- Singh, M.K.; Dwivedi, S.; Yadav, S.S.; Sharma, P.; Khattri, S. Arsenic-Induced Hepatic Toxicity and Its Attenuation by Fruit Extract of Emblica officinalis (Amla) in Mice. Indian J. Clin. Biochem. 2014, 29, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Enterline, P.E.; Day, R.; Marsh, G.M. Cancers related to exposure to arsenic at a copper smelter. Occup. Environ. Med. 1995, 52, 28–32. [Google Scholar] [CrossRef] [Green Version]
- Lubin, J.H.; Pottern, L.M.; Stone, B.J.; Fraumeni, J.F., Jr. Respiratory cancer in a cohort of copper smelter workers: Results from more than 50 years of follow-up. Am. J. Epidemiol. 2000, 151, 554–565. [Google Scholar] [CrossRef] [Green Version]
- Hurtado-Jimenez, R.; Gardea-Torresdey, J.L. Arsenic in drinking water in the Los Altos de Jalisco region of Mexico. Rev. Panam. Salud. Publica. 2006, 20, 236–247. [Google Scholar] [CrossRef]
- Del Razo, L.M.; Garcia-Vargas, G.G.; Valenzuela, O.L.; Castellanos, E.H.; Sanchez-Pena, L.C.; Currier, J.M.; Drobna, Z.; Loomis, D.; Styblo, M. Exposure to arsenic in drinking water is associated with increased prevalence of diabetes: A cross-sectional study in the Zimapan and Lagunera regions in Mexico. Environ. Health 2011, 10, 73. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Patel, M.; Singh, P.; Bundschuh, J.; Pittman, C.U., Jr.; Trakal, L.; Mohan, D. Emerging technologies for arsenic removal from drinking water in rural and peri-urban areas: Methods, experience from, and options for Latin America. Sci. Total Environ. 2019, 694, 133427. [Google Scholar] [CrossRef]
- Souza, A.C.F.; Marchesi, S.C.; de Almeida Lima, G.D.; Machado-Neves, M. Effects of Arsenic Compounds on Microminerals Content and Antioxidant Enzyme Activities in Rat Liver. Biol. Trace Elem. Res. 2018, 183, 305–313. [Google Scholar] [CrossRef]
- Chakraborty, A.K.; Saha, K.C. Arsenical dermatosis from tubewell water in West Bengal. Indian. J. Med. Res. 1987, 85, 326–334. [Google Scholar]
- Guha Mazumder, D.N.; Chakraborty, A.K.; Ghose, A.; Gupta, J.D.; Chakraborty, D.P.; Dey, S.B.; Chattopadhyay, N. Chronic arsenic toxicity from drinking tubewell water in rural West Bengal. Bull. World Health Organ. 1988, 66, 499–506. [Google Scholar]
- Li, S.; Ding, Y.; Niu, Q.; Xu, S.; Pang, L.; Ma, R.; Jing, M.; Feng, G.; Tang, J.X.; Zhang, Q.; et al. Lutein has a protective effect on hepatotoxicity induced by arsenic via Nrf2 signaling. Biomed. Res. Int. 2015, 2015, 315205. [Google Scholar] [CrossRef] [PubMed]
- Khairul, I.; Wang, Q.Q.; Jiang, Y.H.; Wang, C.; Naranmandura, H. Metabolism, toxicity and anticancer activities of arsenic compounds. Oncotarget 2017, 8, 23905–23926. [Google Scholar] [CrossRef] [Green Version]
- Sampayo-Reyes, A.; Tamez-Guerra, R.S.; Bermudez de Leon, M.; Vargas-Villarreal, J.; Lozano-Garza, H.G.; Rodriguez-Padilla, C.; Cortes, C.; Marcos, R.; Hernandez, A. Tocopherol and selenite modulate the transplacental effects induced by sodium arsenite in hamsters. Reprod. Toxicol. 2017, 74, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ahsan, H. Cancer burden from arsenic in drinking water in Bangladesh. Am. J. Public Health 2004, 94, 741–744. [Google Scholar] [CrossRef]
- Sakurai, T.; Yada, N.; Hagiwara, S.; Arizumi, T.; Minaga, K.; Kamata, K.; Takenaka, M.; Minami, Y.; Watanabe, T.; Nishida, N.; et al. Gankyrin induces STAT3 activation in tumor microenvironment and sorafenib resistance in hepatocellular carcinoma. Cancer Sci. 2017, 108, 1996–2003. [Google Scholar] [CrossRef] [PubMed]
- Adebayo, A.O.; Zandbergen, F.; Kozul-Horvath, C.D.; Gruppuso, P.A.; Hamilton, J.W. Chronic exposure to low-dose arsenic modulates lipogenic gene expression in mice. J. Biochem. Mol. Toxicol. 2015, 29, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bailey, K.A.; Wu, M.C.; Ward, W.O.; Smeester, L.; Rager, J.E.; Garcia-Vargas, G.; Del Razo, L.M.; Drobna, Z.; Styblo, M.; Fry, R.C. Arsenic and the epigenome: Interindividual differences in arsenic metabolism related to distinct patterns of DNA methylation. J. Biochem. Mol. Toxicol. 2013, 27, 106–115. [Google Scholar] [CrossRef]
- Wang, Z.; Li, J.; Xiao, W.; Long, J.; Zhang, H. The STAT3 inhibitor S3I-201 suppresses fibrogenesis and angiogenesis in liver fibrosis. Lab. Investig. 2018, 98, 1600–1613. [Google Scholar] [CrossRef]
- Qin, Y.; Shembrey, C.; Smith, J.; Paquet-Fifield, S.; Behrenbruch, C.; Beyit, L.M.; Thomson, B.N.J.; Heriot, A.G.; Cao, Y.; Hollande, F. Laminin 521 enhances self-renewal via STAT3 activation and promotes tumor progression in colorectal cancer. Cancer Lett. 2020, 476, 161–169. [Google Scholar] [CrossRef]
- Aguirre-Vazquez, A.; Sampayo-Reyes, A.; Gonzalez-Escalante, L.; Hernandez, A.; Marcos, R.; Castorena-Torres, F.; Lozano-Garza, G.; Tamez-Guerra, R.; de Leon, M.B. Selenite restores Pax6 expression in neuronal cells of chronically arsenic-exposed Golden Syrian hamsters. Acta Biochim. Pol. 2017, 64, 635–639. [Google Scholar] [CrossRef] [Green Version]
- Elshenawy, O.H.; Abdelhamid, G.; Soshilov, A.A.; Denison, M.S.; El-Kadi, A.O. Down-regulation of cytochrome P450 1A1 by monomethylarsonous acid in human HepG2 cells. Toxicol. Lett. 2017, 270, 34–50. [Google Scholar] [CrossRef]
- Dong, L.W.; Yang, G.Z.; Pan, Y.F.; Chen, Y.; Tan, Y.X.; Dai, R.Y.; Ren, Y.B.; Fu, J.; Wang, H.Y. The oncoprotein p28GANK establishes a positive feedback loop in beta-catenin signaling. Cell Res. 2011, 21, 1248–1261. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.Y.; Wang, H.Y.; Tan, L.; Liu, S.Q.; Cao, H.F.; Wu, M.C. Overexpression of p28/gankyrin in human hepatocellular carcinoma and its clinical significance. World J. Gastroenterol. 2002, 8, 638–643. [Google Scholar] [CrossRef]
- Higashitsuji, H.; Liu, Y.; Mayer, R.J.; Fujita, J. The oncoprotein gankyrin negatively regulates both p53 and RB by enhancing proteasomal degradation. Cell Cycle 2005, 4, 1335–1337. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Li, J.; Zhang, C. STAT3 promotes the proliferation and migration of hepatocellular carcinoma cells by regulating AKT2. Oncol. Lett. 2018, 15, 3333–3338. [Google Scholar] [CrossRef]
- Beems, R.B.; van Beek, L. Short-term (6-week) oral toxicity study of selenium in Syrian hamsters. Food Chem. Toxicol. 1985, 23, 945–947. [Google Scholar] [CrossRef]
- Julius, A.D.; Davies, M.H.; Birt, D.F. Toxic effects of dietary selenium in the Syrian hamster. Ann. Nutr. Metab. 1983, 27, 296–305. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, C.W.; Wu, M.M.; Kuo, T.L. Cancer potential in liver, lung, bladder and kidney due to ingested inorganic arsenic in drinking water. Br. J. Cancer 1992, 66, 888–892. [Google Scholar] [CrossRef] [Green Version]
- Majumdar, K.K.; Guha Mazumder, D.N. Effect of drinking arsenic-contaminated water in children. Indian J. Public Health 2012, 56, 223–226. [Google Scholar] [PubMed]
- Tsuji, P.A.; Carlson, B.A.; Anderson, C.B.; Seifried, H.E.; Hatfield, D.L.; Howard, M.T. Dietary Selenium Levels Affect Selenoprotein Expression and Support the Interferon-gamma and IL-6 Immune Response Pathways in Mice. Nutrients 2015, 7, 6529–6549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwolak, I. The Role of Selenium in Arsenic and Cadmium Toxicity: An Updated Review of Scientific Literature. Biol. Trace Elem. Res. 2020, 193, 44–63. [Google Scholar] [CrossRef] [Green Version]
- Chi, L.; Bian, X.; Gao, B.; Ru, H.; Tu, P.; Lu, K. Sex-Specific Effects of Arsenic Exposure on the Trajectory and Function of the Gut Microbiome. Chem. Res. Toxicol. 2016, 29, 949–951. [Google Scholar] [CrossRef] [Green Version]
- Lindberg, A.L.; Ekstrom, E.C.; Nermell, B.; Rahman, M.; Lonnerdal, B.; Persson, L.A.; Vahter, M. Gender and age differences in the metabolism of inorganic arsenic in a highly exposed population in Bangladesh. Environ. Res. 2008, 106, 110–120. [Google Scholar] [CrossRef]
- Liu, J.; Xie, Y.; Merrick, B.A.; Shen, J.; Ducharme, D.M.; Collins, J.; Diwan, B.A.; Logsdon, D.; Waalkes, M.P. Transplacental arsenic plus postnatal 12-O-teradecanoyl phorbol-13-acetate exposures associated with hepatocarcinogenesis induce similar aberrant gene expression patterns in male and female mouse liver. Toxicol. Appl. Pharmacol. 2006, 213, 216–223. [Google Scholar] [CrossRef]
- Torres-Sanchez, L.; Lopez-Carrillo, L.; Rosado, J.L.; Rodriguez, V.M.; Vera-Aguilar, E.; Kordas, K.; Garcia-Vargas, G.G.; Cebrian, M.E. Sex differences in the reduction of arsenic methylation capacity as a function of urinary total and inorganic arsenic in Mexican children. Environ. Res. 2016, 151, 38–43. [Google Scholar] [CrossRef]
- Waalkes, M.P.; Ward, J.M.; Diwan, B.A. Induction of tumors of the liver, lung, ovary and adrenal in adult mice after brief maternal gestational exposure to inorganic arsenic: Promotional effects of postnatal phorbol ester exposure on hepatic and pulmonary, but not dermal cancers. Carcinogenesis 2004, 25, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Drobna, Z.; Styblo, M.; Thomas, D.J. An Overview of Arsenic Metabolism and Toxicity. Curr. Protoc. Toxicol. 2009, 42, 4–31. [Google Scholar] [CrossRef] [Green Version]
- Chung, C.J.; Pu, Y.S.; Chen, Y.T.; Su, C.T.; Wu, C.C.; Shiue, H.S.; Huang, C.Y.; Hsueh, Y.M. Protective effects of plasma alpha-tocopherols on the risk of inorganic arsenic-related urothelial carcinoma. Sci. Total Environ. 2011, 409, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- Johnson, V.J.; Tsunoda, M.; Sharma, R.P. Increased production of proinflammatory cytokines by murine macrophages following oral exposure to sodium selenite but not to seleno-L-methionine. Arch. Environ. Contam. Toxicol. 2000, 39, 243–250. [Google Scholar] [CrossRef]
- Benko, I.; Nagy, G.; Tanczos, B.; Ungvari, E.; Sztrik, A.; Eszenyi, P.; Prokisch, J.; Banfalvi, G. Subacute toxicity of nano-selenium compared to other selenium species in mice. Environ. Toxicol. Chem. 2012, 31, 2812–2820. [Google Scholar] [CrossRef]
- Jacevic, V.; Jokic, G.; Dragojevic-Simic, V.; Bokonjic, D.; Vucinic, S.; Vuksa, M. Acute toxicity of sodium selenite in rodents: Pathomorphological study. MMSL 2011, 80, 90–96. [Google Scholar] [CrossRef]
- Sun, H.J.; Rathinasabapathi, B.; Wu, B.; Luo, J.; Pu, L.P.; Ma, L.Q. Arsenic and selenium toxicity and their interactive effects in humans. Environ. Int. 2014, 69, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, E.M.; Ringer, L.; Bulut, G.; Sajwan, K.P.; Hall, M.D.; Lee, Y.C.; Peaceman, D.; Ozdemirli, M.; Rodriguez, O.; Macdonald, T.J.; et al. Arsenic trioxide inhibits human cancer cell growth and tumor development in mice by blocking Hedgehog/GLI pathway. J. Clin. Investig. 2011, 121, 148–160. [Google Scholar] [CrossRef] [Green Version]
- Miller, W.H., Jr.; Schipper, H.M.; Lee, J.S.; Singer, J.; Waxman, S. Mechanisms of action of arsenic trioxide. Cancer Res. 2002, 62, 3893–3903. [Google Scholar]
- Zhang, P. On arsenic trioxide in the clinical treatment of acute promyelocytic leukemia. Leuk. Res. Rep. 2017, 7, 29–32. [Google Scholar] [CrossRef]
- Hoonjan, M.; Jadhav, V.; Bhatt, P. Arsenic trioxide: Insights into its evolution to an anticancer agent. J. Biol. Inorg. Chem. 2018, 23, 313–329. [Google Scholar] [CrossRef]
- Lohner, T.W.; Reash, R.J.; Willet, V.E.; Rose, L.A. Assessment of tolerant sunfish populations (Lepomis sp.) inhabiting selenium-laden coal ash effluents. 1. Hematological and population level assessment. Ecotoxicol. Environ. Saf. 2001, 50, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, F.W.; Hashimoto, A.C.; Shafer, L.A.; Dow, S.; Berry, M.J.; Hoffmann, P.R. Dietary selenium modulates activation and differentiation of CD4+ T cells in mice through a mechanism involving cellular free thiols. J. Nutr. 2010, 140, 1155–1161. [Google Scholar] [CrossRef]
- Matsushita, M.; Freigang, S.; Schneider, C.; Conrad, M.; Bornkamm, G.W.; Kopf, M. T cell lipid peroxidation induces ferroptosis and prevents immunity to infection. J. Exp. Med. 2015, 212, 555–568. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.; Fan, Y.; Zhang, Z.; Chen, C.; Chen, C.; Wang, X.; Deng, J.; Peng, G.; Hu, Y.; Cao, S.; et al. Sodium selenite inhibits deoxynivalenol-induced injury in GPX1-knockdown porcine splenic lymphocytes in culture. Sci. Rep. 2018, 8, 17676. [Google Scholar] [CrossRef]
- Di Napoli, M.; Maresca, V.; Sorbo, S.; Varcamonti, M.; Basile, A.; Zanfardino, A. Proteins of the fruit pulp of Acca sellowiana have antimicrobial activity directed against the bacterial membranes. Nat. Prod. Res. 2019, 35, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Tortora, F.; Notariale, R.; Maresca, V.; Good, K.V.; Sorbo, S.; Basile, A.; Piscopo, M.; Manna, C. Phenol-Rich Feijoa sellowiana (Pineapple Guava) Extracts Protect Human Red Blood Cells from Mercury-Induced Cellular Toxicity. Antioxidants 2019, 8, 220. [Google Scholar] [CrossRef] [Green Version]
- Kane, A.; Deenick, E.K.; Ma, C.S.; Cook, M.C.; Uzel, G.; Tangye, S.G. STAT3 is a central regulator of lymphocyte differentiation and function. Curr. Opin. Immunol. 2014, 28, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.O.; Panopoulos, A.D.; Nurieva, R.; Chang, S.H.; Wang, D.; Watowich, S.S.; Dong, C. STAT3 regulates cytokine-mediated generation of inflammatory helper T cells. J. Biol. Chem. 2007, 282, 9358–9363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browning, L.; Patel, M.R.; Horvath, E.B.; Tawara, K.; Jorcyk, C.L. IL-6 and ovarian cancer: Inflammatory cytokines in promotion of metastasis. Cancer Manag. Res. 2018, 10, 6685–6693. [Google Scholar] [CrossRef] [Green Version]
- Jung, I.H.; Choi, J.H.; Chung, Y.Y.; Lim, G.L.; Park, Y.N.; Park, S.W. Predominant Activation of JAK/STAT3 Pathway by Interleukin-6 Is Implicated in Hepatocarcinogenesis. Neoplasia 2015, 17, 586–597. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, N.; Kannan, G.M.; Pant, B.P.; Jaiswal, D.K.; Malhotra, P.R.; Flora, S.J. Arsenic-induced changes in certain neurotransmitter levels and their recoveries following chelation in rat whole brain. Toxicol. Lett. 1997, 92, 201–208. [Google Scholar] [CrossRef]
- Hernandez-Zavala, A.; Matousek, T.; Drobna, Z.; Paul, D.S.; Walton, F.; Adair, B.M.; Jiri, D.; Thomas, D.J.; Styblo, M. Speciation analysis of arsenic in biological matrices by automated hydride generation-cryotrapping-atomic absorption spectrometry with multiple microflame quartz tube atomizer (multiatomizer). J. Anal. At. Spectrom. 2008, 23, 342–351. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | n | Treatment |
|---|---|---|
| Control | 9 | Tap water |
| Arsenic | 10 | Sodium arsenite, 100 ppm |
| α-TOS | 9 | α-TOS, 6 ppm |
| Arsenic + α-TOS | 9 | Sodium arsenite and α-TOS |
| Selenite | 10 | Sodium selenite, 8.5 ppm |
| Arsenic + Selenite | 7 | Sodium arsenite and sodium selenite |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camacho-Moll, M.E.; Sampayo-Reyes, A.; Castorena-Torres, F.; Lozano-Garza, G.; Alarcón-Galván, G.; Hernández, A.; Marcos, R.; Alcocer-González, J.M.; Tamez-Guerra, R.; Bermúdez de León, M. Selenite Downregulates STAT3 Expression and Provokes Lymphocytosis in the Liver of Chronically Exposed Syrian Golden Hamsters. Molecules 2021, 26, 5614. https://doi.org/10.3390/molecules26185614
Camacho-Moll ME, Sampayo-Reyes A, Castorena-Torres F, Lozano-Garza G, Alarcón-Galván G, Hernández A, Marcos R, Alcocer-González JM, Tamez-Guerra R, Bermúdez de León M. Selenite Downregulates STAT3 Expression and Provokes Lymphocytosis in the Liver of Chronically Exposed Syrian Golden Hamsters. Molecules. 2021; 26(18):5614. https://doi.org/10.3390/molecules26185614
Chicago/Turabian StyleCamacho-Moll, María Elena, Adriana Sampayo-Reyes, Fabiola Castorena-Torres, Gerardo Lozano-Garza, Gabriela Alarcón-Galván, Alba Hernández, Ricard Marcos, Juan Manuel Alcocer-González, Reyes Tamez-Guerra, and Mario Bermúdez de León. 2021. "Selenite Downregulates STAT3 Expression and Provokes Lymphocytosis in the Liver of Chronically Exposed Syrian Golden Hamsters" Molecules 26, no. 18: 5614. https://doi.org/10.3390/molecules26185614