
Whole-Genome Sequencing of Corallococcus sp. Strain EGB Reveals the Genetic Determinants Linking Taxonomy and Predatory Behavior

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culturing and DNA Isolation of Corallococcus sp. Strain EGB
2.2. Genome Sequencing, Assembly, and Annotation
2.3. Comparative Genomics Analysis
2.4. Phylogenetic Analysis
2.5. Bioinformatics Analyses
3. Results
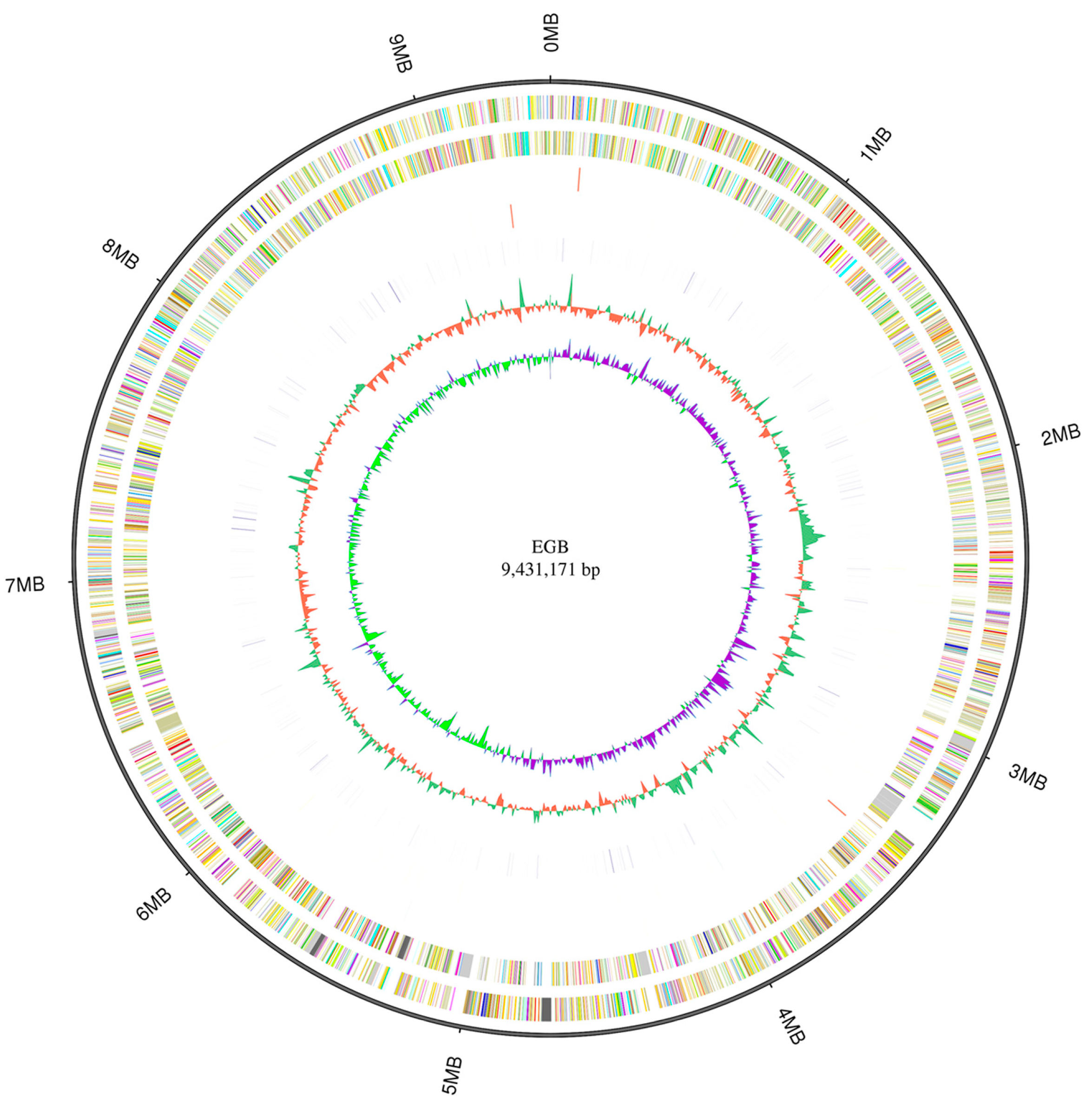
3.1. Genomic Characterization of Corallococcus sp. Strain EGB
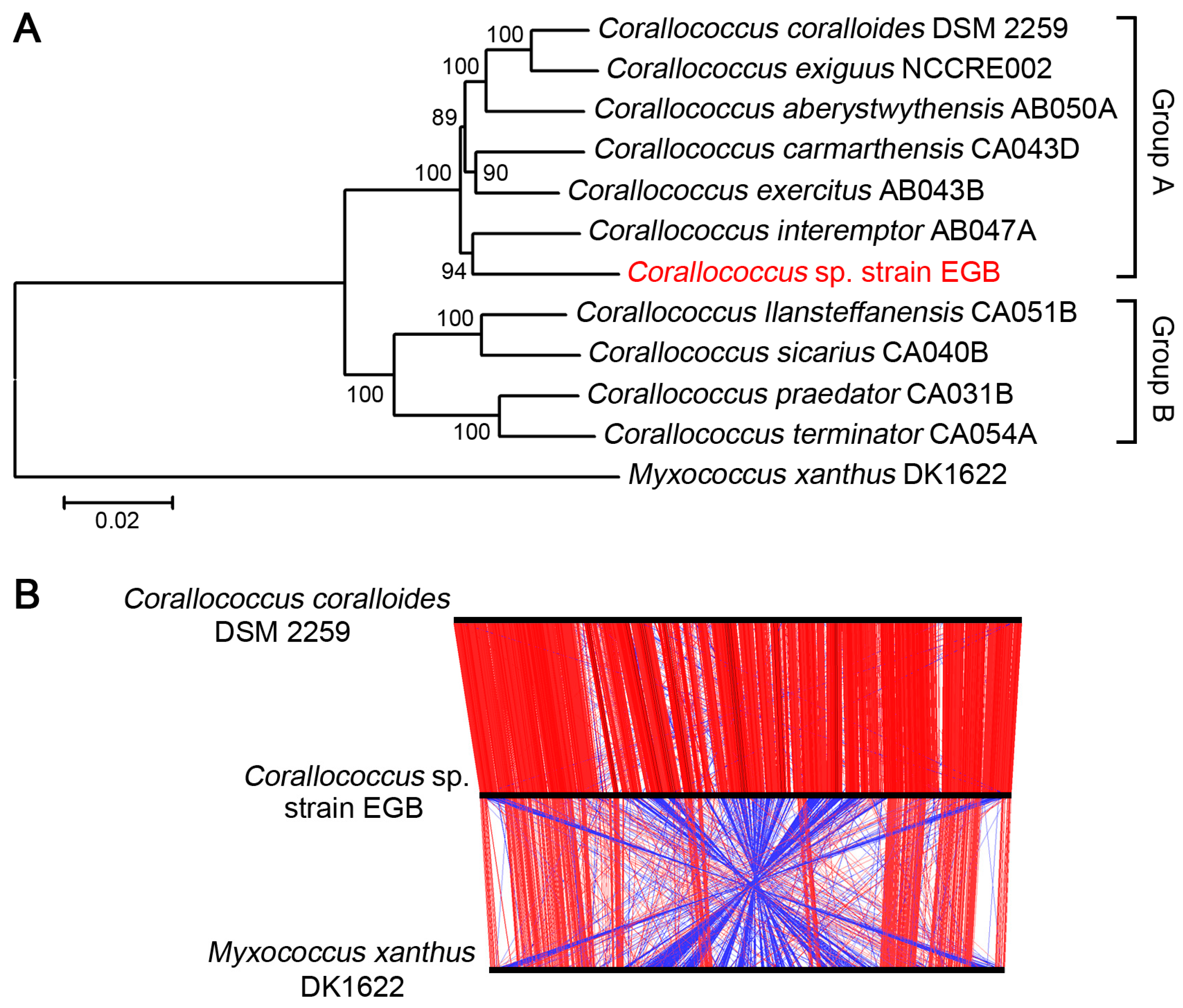
3.2. Strain EGB was Inferred as A Novel Species
3.3. Corallococcus Pan-Genome Was Large and Open
3.4. Genes Involved in Secretion
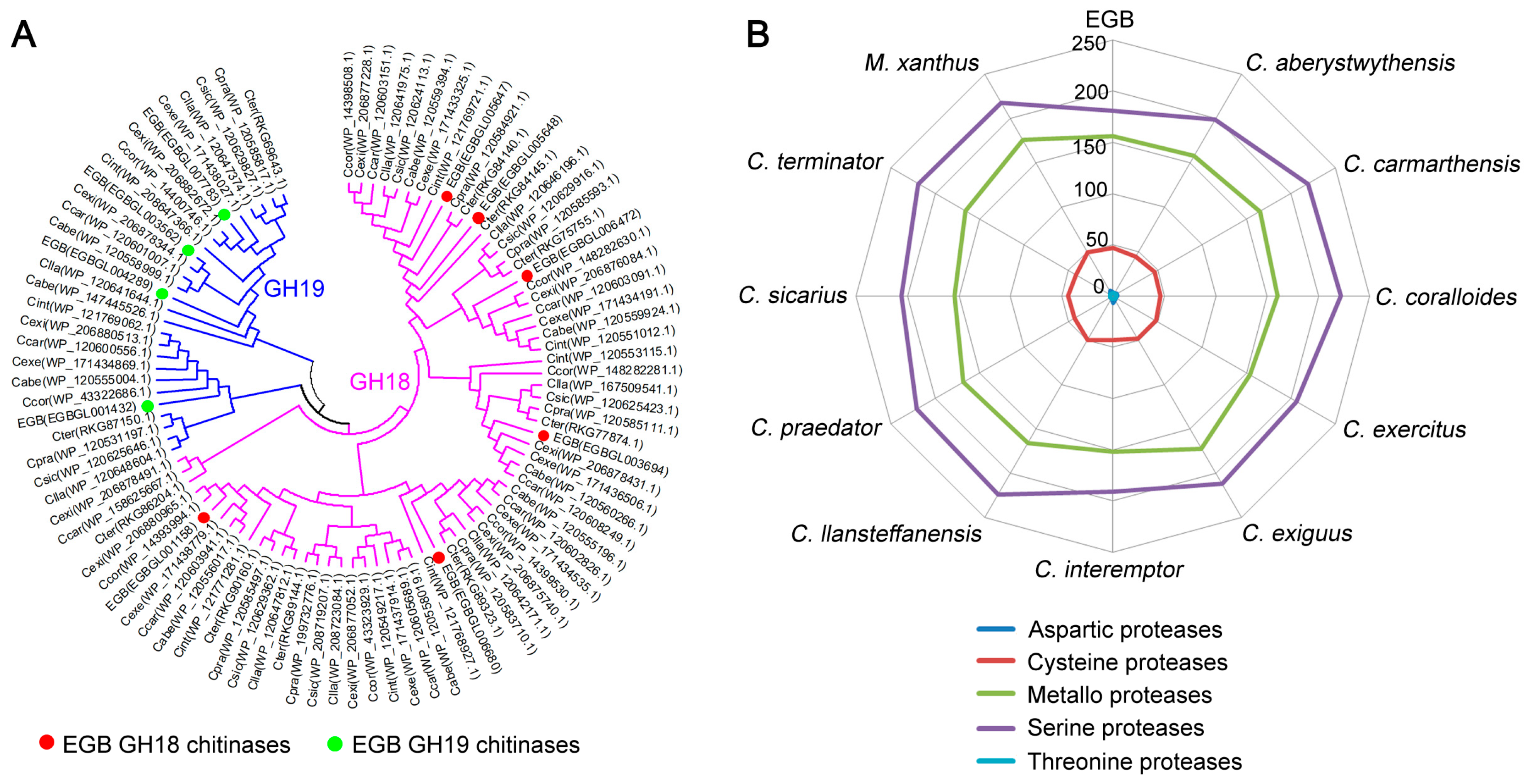
3.5. Expansion of Microorganism Surface Degrading Enzymes
3.6. Secondary Metabolite Gene Clusters Are Enriched in the Genome
3.7. Genomic Evidence for Signal Transduction and Regulation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reichenbach, H. The ecology of the myxobacteria. Environ. Microbiol. 1999, 1, 15–21. [Google Scholar] [CrossRef]
- Konovalova, A.; Petters, T.; Sogaard-Andersen, L. Extracellular biology of Myxococcus xanthus. FEMS Microbiol. Rev. 2010, 34, 89–106. [Google Scholar] [CrossRef] [Green Version]
- Whitworth, D.E.; Zwarycz, A. A genomic survey of signalling in the Myxococcaceae. Microorganisms 2020, 8, 1739. [Google Scholar] [CrossRef]
- Morgan, A.D.; MacLean, R.C.; Hillesland, K.L.; Velicer, G.J. Comparative analysis of Myxococcus predation on Soil Bacteria. Appl. Environ. Microb. 2010, 76, 6920–6927. [Google Scholar] [CrossRef] [Green Version]
- Livingstone, P.G.; Morphew, R.M.; Whitworth, D.E. Myxobacteria are able to prey broadly upon clinically-relevant pathogens, exhibiting a prey range which cannot be explained by phylogeny. Front. Microbiol. 2017, 8, 1593. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Wei, X.M.; Ebright, R.; Wall, D. Antibiotic production by myxobacteria plays a role in predation. J. Bacteriol. 2011, 193, 4626–4633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hug, J.J.; Panter, F.; Krug, D.; Muller, R. Genome mining reveals uncommon alkylpyrones as type III PKS products from myxobacteria. J. Ind. Microbiol. Biotechnol. 2019, 46, 319–334. [Google Scholar] [CrossRef]
- Chambers, J.; Sparks, N.; Sydney, N.; Livingstone, P.G.; Cookson, A.R.; Whitworth, D.E. Comparative genomics and pan-genomics of the Myxococcaceae, including a description of five novel species: Myxococcus eversor sp. nov., Myxococcus IlanfairpwlIg-wyngyllgogetychwymdrobwIlllantysiliogogogochensis sp. nov., Myxococcus vastator sp. nov., Pyxidicoccus caerfyrddinensis sp. nov., and Pyxidicoccus trucidator sp. nov. Genome Biol. Evol. 2020, 12, 2279–2302. [Google Scholar]
- Perez-Burgos, M.; Garcia-Romero, I.; Jung, J.; Schander, E.; Valvano, M.A.; Sogaard-Andersen, L. Characterization of the exopolysaccharide biosynthesis pathway in Myxococcus xanthus. J. Bacteriol. 2020, 202, e00335-20. [Google Scholar] [CrossRef]
- Li, Z.K.; Ye, X.F.; Chen, P.L.; Ji, K.; Zhou, J.; Wang, F.; Dong, W.L.; Huang, Y.; Zhang, Z.G.; Cui, Z.L. Antifungal potential of Corallococcus sp strain EGB against plant pathogenic fungi. Biol. Control 2017, 110, 10–17. [Google Scholar] [CrossRef]
- Zhou, J.; Li, Z.K.; Wu, J.L.; Li, L.F.; Li, D.; Ye, X.F.; Luo, X.; Huang, Y.; Cui, Z.L.; Cao, H. Functional analysis of a novel beta-(1,3)-glucanase from Corallococcus sp strain EGB containing a fascin-like module. Appl. Environ. Microb. 2017, 83, e01016-17. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Chen, J.H.; Li, Z.K.; Ye, X.F.; Dong, W.L.; Jiang, M.; Huang, Y.; Cui, Z.L. Enzymatic properties of a multi-specific beta-(1,3)-glucanase from Corallococcus sp. EGB and its potential antifungal applications. Protein Expres Purif 2019, 164, 105481. [Google Scholar] [CrossRef]
- Li, Z.K.; Xia, C.Y.; Wang, Y.X.; Li, X.; Qiao, Y.; Li, C.Y.; Zhou, J.; Zhang, L.; Ye, X.F.; Huang, Y.; et al. Identification of an endo-chitinase from Corallococcus sp. EGB and evaluation of its antifungal properties. Int. J. Biol. Macromol. 2019, 132, 1235–1243. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.F.; Chen, Y.; Ma, S.Y.; Yuan, T.; Wu, Y.X.; Li, Y.X.; Zhao, Y.Q.; Chen, S.Y.; Zhang, Y.W.; Li, L.Y.; et al. Biocidal effects of volatile organic compounds produced by the myxobacterium Corrallococcus sp. EGB against fungal phytopathogens. Food Microbiol. 2020, 91, 103502. [Google Scholar] [CrossRef]
- Livingstone, P.G.; Morphew, R.M.; Whitworth, D.E. Genome sequencing and pan-genome analysis of 23 Corallococcus spp. strains reveal unexpected diversity, with particular plasticity of predatory gene sets. Front. Microbiol. 2018, 9, 3187. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.J.; Han, K.; Feng, J.; Zhuo, L.; Li, Y.J.; Li, Y.Z. The complete genome sequence and analysis of a plasmid-bearing myxobacterial strain Myxococcus fulvus 124B02 (M 206081). Stand Genom. Sci. 2016, 11, 1. [Google Scholar] [CrossRef] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatusov, R.L.; Galperin, M.Y.; Natale, D.A.; Koonin, E.V. The COG database: A tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 2000, 28, 33–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enright, A.J.; Van Dongen, S.; Ouzounis, C.A. An efficient algorithm for large-scale detection of protein families. Nucleic Acids Res. 2002, 30, 1575–1584. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carver, T.J.; Rutherford, K.M.; Berriman, M.; Rajandream, M.A.; Barrell, B.G.; Parkhill, J. ACT: The Artemis comparison tool. Bioinformatics 2005, 21, 3422–3423. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.H.; Ha, S.M.; Lim, J.; Kwon, S.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Anton Leeuw Int. J. G 2017, 110, 1281–1286. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Goker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sonderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Zhang, H.; Yohe, T.; Huang, L.; Entwistle, S.; Wu, P.; Yang, Z.; Busk, P.K.; Xu, Y.; Yin, Y. dbCAN2: A meta server for automated carbohydrate-active enzyme annotation. Nucleic Acids Res. 2018, 46, W95–W101. [Google Scholar] [CrossRef] [Green Version]
- Rawlings, N.D.; Waller, M.; Barrett, A.J.; Bateman, A. MEROPS: The database of proteolytic enzymes, their substrates and inhibitors. Nucleic Acids Res. 2014, 42, D503–D509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.W.; De Meyer, S.; et al. Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Micr. 2018, 68, 461–466. [Google Scholar] [CrossRef]
- Natale, P.; Bruser, T.; Driessen, A.J. Sec- and Tat-mediated protein secretion across the bacterial cytoplasmic membrane--distinct translocases and mechanisms. Biochim. Biophys. Acta 2008, 1778, 1735–1756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, E.R.; Mecsas, J. Bacterial Secretion Systems: An Overview. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Berleman, J.E.; Allen, S.; Danielewicz, M.A.; Remis, J.P.; Gorur, A.; Cunha, J.; Hadi, M.Z.; Zusman, D.R.; Northen, T.R.; Witkowska, H.E.; et al. The lethal cargo of Myxococcus xanthus outer membrane vesicles. Front. Microbiol. 2014, 5, 474. [Google Scholar] [CrossRef]
- Botella, J.A.; Murillo, F.J.; Ruizvazquez, R. A cluster of structural and regulatory genes for light-induced carotenogenesis in Myxococcus xanthus. Eur. J. Biochem. 1995, 233, 238–248. [Google Scholar] [CrossRef]
- Lorenzen, W.; Ahrendt, T.; Bozhuyuk, K.A.J.; Bode, H.B. A multifunctional enzyme is involved in bacterial ether lipid biosynthesis. Nat. Chem. Biol. 2014, 10, 425–427. [Google Scholar] [CrossRef]
- Gaitatzis, N.; Kunze, B.; Muller, R. In vitro reconstitution of the myxochelin biosynthetic machinery of Stigmatella aurantiaca Sg a15: Biochemical characterization of a reductive release mechanism from nonribosomal peptide synthetases. Proc. Natl. Acad. Sci. USA 2001, 98, 11136–11141. [Google Scholar] [CrossRef] [Green Version]
- Giglio, S.; Jiang, J.Y.; Saint, C.P.; Cane, D.E.; Monis, P.T. Isolation and characterization of the gene associated with geosmin production in cyanobacteria. Environ. Sci. Technol. 2008, 42, 8027–8032. [Google Scholar] [PubMed] [Green Version]
- Lang, E.; Stackebrandt, E. Emended descriptions of the genera Myxococcus and Corallococcus, typification of the species Myxococcus stipitatus and Myxococcus macrosporus and a proposal that they be represented by neotype strains. Request for an Opinion. Int. J. Syst. Evol. Microbiol. 2009, 59, 2122–2128. [Google Scholar] [CrossRef]
- Garcia, R.; Gerth, K.; Stadler, M.; Dogma, I.J., Jr.; Muller, R. Expanded phylogeny of myxobacteria and evidence for cultivation of the ‘unculturables’. Mol. Phylogenet Evol. 2010, 57, 878–887. [Google Scholar] [CrossRef] [PubMed]
- Hinger, I.; Ansorge, R.; Mussmann, M.; Romano, S. Phylogenomic analyses of members of the widespread marine heterotrophic genus pseudovibrio suggest distinct evolutionary trajectories and a novel genus, Polycladidibacter gen. nov. Appl. Environ. Microbiol. 2020, 86, e02395-19. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Genome Size (Mb) | %GC | Contigs | CDS * Number | Accession |
|---|---|---|---|---|---|---|
| Corallococcus sp. | EGB | 9.4 | 70.4 | 1 | 7916 | CP079946 |
| C. coralloides | DSM 2259 | 10.08 | 69.9 | 1 | 7952 | CP003389.1 |
| C. exiguus | NCCRE002 | 10.54 | 69.7 | 8 | 8283 | GCA_017302975.1 |
| C. interemptor | AB047A | 9.47 | 70 | 459 | 7569 | GCA_003668875.1 |
| C. praedator | CA031B | 10.51 | 69.7 | 1491 | 8176 | GCA_003612125.1 |
| C. aberystwythensis | AB050A | 9.98 | 70 | 625 | 8319 | GCA_003612165.1 |
| C. llansteffanensis | CA051B | 10.53 | 70.3 | 1244 | 8140 | GCA_003612055.1 |
| C. sicarius | CA040B | 10.39 | 70.2 | 802 | 8410 | GCA_003611735.1 |
| C. carmarthensis | CA043D | 10.79 | 69.9 | 530 | 8523 | GCA_003611695.1 |
| C. exercitus | AB043B | 10.26 | 70.2 | 690 | 8102 | GCA_013116705.1 |
| C. terminator | CA054A | 10.35 | 69.5 | 863 | 8454 | GCA_003611635.1 |
| dDDH\ANI | EGB | Cint | Cexe | Ccar | Cabe | Cexi | Ccor | Clla | Csic | Cpra | Cter | Mxan |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EGB | 100 | 91.19 | 91.11 | 90.88 | 90.79 | 91.34 | 91.51 | 86.63 | 86.21 | 86.17 | 85.83 | 78.37 |
| Cint | 42.4 | 100 | 91.29 | 91.12 | 91.31 | 91.72 | 92.27 | 86.53 | 86.3 | 86.18 | 85.9 | 78.35 |
| Cexe | 41.9 | 42.7 | 100 | 92.73 | 92.6 | 91.56 | 91.83 | 87.37 | 87.03 | 86.87 | 86.52 | 78.43 |
| Ccar | 41.5 | 42.1 | 47.5 | 100 | 92.57 | 91.62 | 91.78 | 87.18 | 86.87 | 86.81 | 86.38 | 78.41 |
| Cabe | 41.2 | 42.7 | 47.1 | 47.6 | 100 | 91.55 | 91.84 | 87.04 | 86.73 | 86.7 | 86.2 | 78.41 |
| Cexi | 42.4 | 43.1 | 43.1 | 43.6 | 43.2 | 100 | 94.32 | 86.61 | 86.13 | 86.12 | 85.82 | 78.15 |
| Ccor | 43.7 | 46.1 | 43.9 | 44.4 | 44.3 | 54.1 | 100 | 86.82 | 86.46 | 86.38 | 85.95 | 78.25 |
| Clla | 30.9 | 30.8 | 32.4 | 32.1 | 31.7 | 30.7 | 31.1 | 100 | 93.42 | 89.27 | 88.82 | 78.78 |
| Csic | 29.9 | 30 | 31.6 | 31.3 | 31 | 29.8 | 30.2 | 50.4 | 100 | 88.87 | 88.45 | 78.65 |
| Cpra | 29.9 | 30.1 | 31.5 | 31.4 | 31 | 30 | 30.5 | 36.6 | 35.9 | 100 | 93.23 | 78.68 |
| Cter | 29.3 | 29.7 | 30.8 | 30.6 | 30.2 | 29.4 | 29.8 | 35.2 | 35 | 49.6 | 100 | 78.27 |
| Mxan | 21.2 | 21.3 | 21.6 | 21.6 | 21.5 | 21.2 | 21.2 | 21.7 | 21.5 | 21.6 | 21.2 | 100 |
| EGB | Cabe | Ccar | Ccor | Cexe | Cexi | Cint | Clla | Cpra | Csic | Cter | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ser/Thr kinases | 96 | 106 | 123 | 103 | 112 | 105 | 108 | 113 | 101 | 116 | 106 |
| Two-component system proteins | 289 | 299 | 307 | 303 | 318 | 297 | 284 | 322 | 323 | 312 | 311 |
| Histidine kinases | 146 | 156 | 157 | 160 | 167 | 153 | 151 | 170 | 162 | 167 | 163 |
| Phosphotransfer proteins | 5 | 5 | 3 | 2 | 4 | 2 | 2 | 5 | 7 | 4 | 4 |
| Response regulators | 138 | 138 | 147 | 141 | 147 | 142 | 131 | 147 | 154 | 141 | 144 |
| Transcription factors | 320 | 322 | 380 | 351 | 353 | 378 | 306 | 344 | 352 | 303 | 334 |
| One-component system proteins | 60 | 68 | 86 | 79 | 78 | 84 | 61 | 75 | 69 | 61 | 68 |
| Response regulators | 51 | 42 | 54 | 48 | 47 | 52 | 42 | 46 | 51 | 44 | 50 |
| sigma factors | 53 | 58 | 58 | 56 | 57 | 56 | 51 | 58 | 56 | 53 | 50 |
| Transcriptional regulators | 156 | 154 | 182 | 168 | 171 | 186 | 152 | 165 | 176 | 145 | 166 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Wang, Y.; Xia, C.; Li, X.; Ye, X.; Fan, Q.; Huang, Y.; Li, Z.; Zhu, C.; Cui, Z. Whole-Genome Sequencing of Corallococcus sp. Strain EGB Reveals the Genetic Determinants Linking Taxonomy and Predatory Behavior. Genes 2021, 12, 1421. https://doi.org/10.3390/genes12091421
Zhao Y, Wang Y, Xia C, Li X, Ye X, Fan Q, Huang Y, Li Z, Zhu C, Cui Z. Whole-Genome Sequencing of Corallococcus sp. Strain EGB Reveals the Genetic Determinants Linking Taxonomy and Predatory Behavior. Genes. 2021; 12(9):1421. https://doi.org/10.3390/genes12091421
Chicago/Turabian StyleZhao, Yuqiang, Yanxin Wang, Chengyao Xia, Xu Li, Xianfeng Ye, Qiwen Fan, Yan Huang, Zhoukun Li, Cancan Zhu, and Zhongli Cui. 2021. "Whole-Genome Sequencing of Corallococcus sp. Strain EGB Reveals the Genetic Determinants Linking Taxonomy and Predatory Behavior" Genes 12, no. 9: 1421. https://doi.org/10.3390/genes12091421