
NOD2 Genotypes Affect the Symptoms and Mortality in the Porcine Circovirus 2-Spreading Pig Population

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement, Pig Population and Sampling
2.2. Comparison of Capsid Gene Sequence of PCV2
2.3. Genotypes of the NOD2 Gene
2.4. Computational Analysis
3. Results
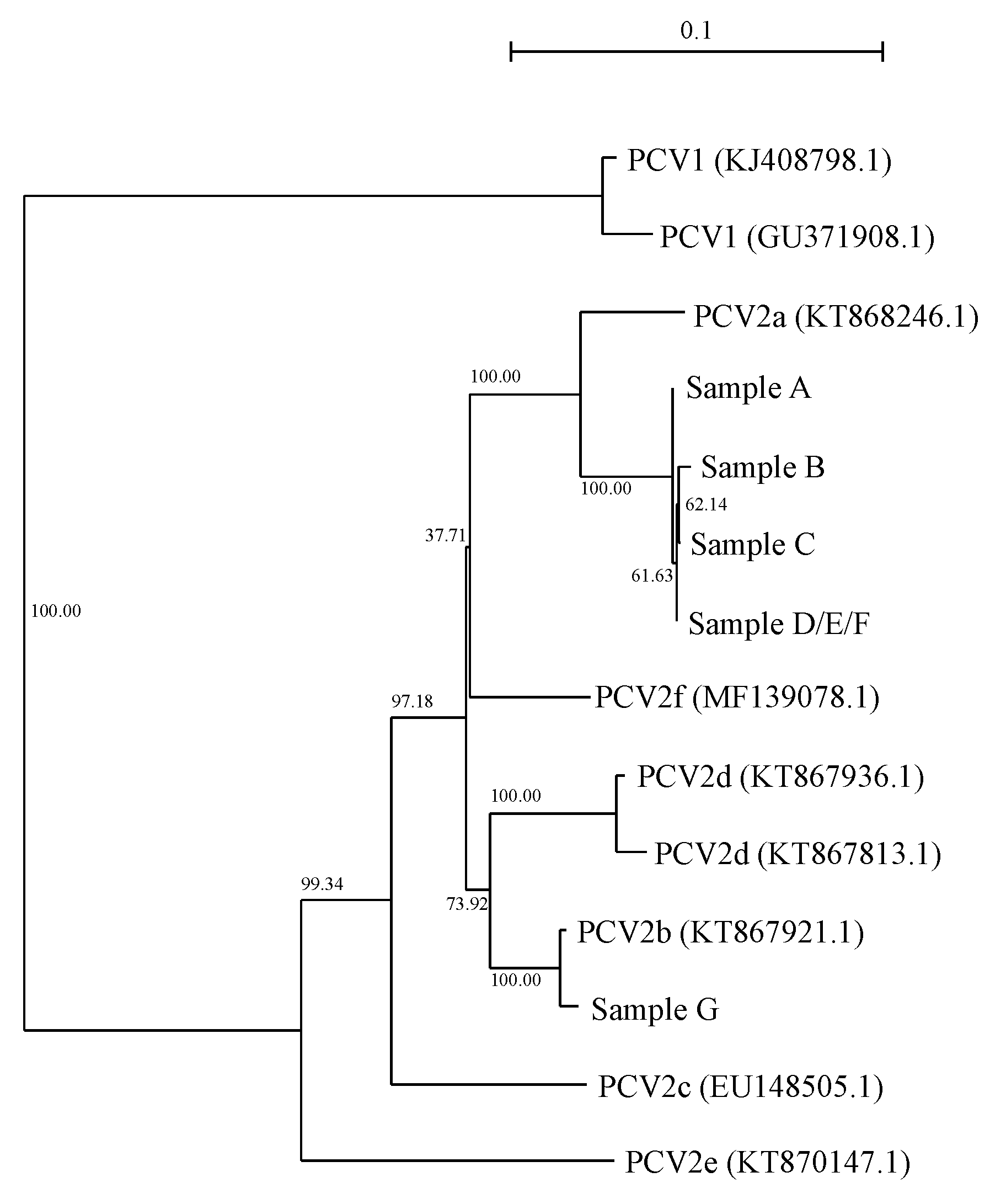
3.1. Phylogenetic Differences in PCV2 Strains before and after Wasting Symptoms and Frequent Mortality
3.2. Marked Differences in NOD2 Genotype Frequency before and after the Emergence of PCVAD
3.3. Significant Influence of NOD2 Genotypes on Pig Mortality during the Spread of PCV2b
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brubaker, S.W.; Bonham, K.S.; Zanoni, I.; Kagan, J.C. Innate Immune Pattern Recognition: A cell biological perspective. Annu. Rev. Immunol. 2015, 33, 257–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skevaki, C.; Pararas, M.; Kostelidou, K.; Tsakris, A.; Routsias, J.G. Single nucleotide polymorphisms of Toll-like receptors and susceptibility to infectious diseases. Clin. Exp. Immunol. 2015, 180, 165–177. [Google Scholar] [CrossRef] [Green Version]
- Kojima-Shibata, C.; Shinkai, H.; Morozumi, T.; Jozaki, K.; Toki, D.; Matsumoto, T.; Kadowaki, H.; Suzuki, E.; Uenishi, H. Differences in distribution of single nucleotide polymorphisms among intracellular pattern recognition receptors in pigs. Immunogenetics 2009, 61, 153–160. [Google Scholar] [CrossRef]
- Morozumi, T.; Uenishi, H. Polymorphism distribution and structural conservation in RNA-sensing Toll-like receptors 3, 7, and 8 in pigs. Biochim. Biophys. Acta 2009, 1790, 267–274. [Google Scholar] [CrossRef]
- Shinkai, H.; Tanaka, M.; Morozumi, T.; Eguchi-Ogawa, T.; Okumura, N.; Muneta, Y.; Awata, T.; Uenishi, H. Biased distribution of single nucleotide polymorphisms (SNPs) in porcine Toll-like receptor 1 (TLR1), TLR2, TLR4, TLR5, and TLR6 genes. Immunogenetics 2006, 58, 324–330. [Google Scholar] [CrossRef]
- Uenishi, H.; Shinkai, H. Porcine Toll-like receptors: The front line of pathogen monitoring and possible implications for disease resistance. Dev. Comp. Immunol. 2009, 33, 353–361. [Google Scholar] [CrossRef]
- Uenishi, H.; Shinkai, H.; Morozumi, T.; Muneta, Y. Genomic survey of polymorphisms in pattern recognition receptors and their possible relationship to infections in pigs. Vet. Immunol. Immunopathol. 2012, 148, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Shinkai, H.; Suzuki, R.; Akiba, M.; Okumura, N.; Uenishi, H. Porcine Toll-like receptors: Recognition of Salmonella enterica serovar Choleraesuis and influence of polymorphisms. Mol. Immunol. 2011, 48, 1114–1120. [Google Scholar] [CrossRef] [PubMed]
- Muneta, Y.; Arai, N.; Yakabe, Y.; Eguchi, M.; Shibahara, T.; Sakuma, A.; Shinkai, H.; Uenishi, H.; Hirose, K.; Akiba, M. In vivo effect of a TLR5 SNP (C1205T) on Salmonella enterica serovar Typhimurium infection in weaned, specific pathogen-free Landrace piglets. Microbiol. Immunol. 2018, 62, 380–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamaillard, M.; Hashimoto, M.; Horie, Y.; Masumoto, J.; Qiu, S.; Saab, L.; Ogura, Y.; Kawasaki, A.; Fukase, K.; Kusumoto, S.; et al. An essential role for NOD1 in host recognition of bacterial peptidoglycan containing diaminopimelic acid. Nat. Immunol. 2003, 4, 702–707. [Google Scholar] [CrossRef]
- Li, J.; Moran, T.; Swanson, E.; Julian, C.; Harris, J.; Bonen, D.K.; Hedl, M.; Nicolae, D.L.; Abraham, C.; Cho, J.H. Regulation of IL-8 and IL-1β expression in Crohn’s disease associated NOD2/CARD15 mutations. Hum. Mol. Genet. 2004, 13, 1715–1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.G.; Park, J.H.; Reimer, T.; Baker, D.P.; Kawai, T.; Kumar, H.; Akira, S.; Wobus, C.; Nuñez, G. Viral infection augments Nod1/2 signaling to potentiate lethality associated with secondary bacterial infections. Cell Host Microbe 2011, 9, 496–507. [Google Scholar] [CrossRef] [Green Version]
- Lupfer, C.; Thomas, P.G.; Kanneganti, T.D. Nucleotide oligomerization and binding domain 2-dependent dendritic cell activation is necessary for innate immunity and optimal CD8+ T Cell responses to influenza A virus infection. J. Virol. 2014, 88, 8946–8955. [Google Scholar] [CrossRef] [Green Version]
- Sabbah, A.; Chang, T.H.; Harnack, R.; Frohlich, V.; Tominaga, K.; Dube, P.H.; Xiang, Y.; Bose, S. Activation of innate immune antiviral responses by Nod2. Nat. Immunol. 2009, 10, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Shinkai, H.; Matsumoto, T.; Toki, D.; Okumura, N.; Terada, K.; Uenishi, H. Porcine NOD1 polymorphisms with impaired ligand recognition and their distribution in pig populations. Mol. Immunol. 2015, 63, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Jozaki, K.; Shinkai, H.; Tanaka-Matsuda, M.; Morozumi, T.; Matsumoto, T.; Toki, D.; Okumura, N.; Eguchi-Ogawa, T.; Kojima-Shibata, C.; Kadowaki, H.; et al. Influence of polymorphisms in porcine NOD2 on ligand recognition. Mol. Immunol. 2009, 47, 247–252. [Google Scholar] [CrossRef]
- Harding, J.S.; Clark, E.G. Recognition and diagnosing postweaning multisystemic wasting syndrome (PMWS). Swine Health Prod. 1997, 5, 201–203. [Google Scholar]
- Ellis, J.; Clark, E.; Haines, D.; West, K.; Krakowka, S.; Kennedy, S.; Allan, G.M. Porcine circovirus-2 and concurrent infections in the field. Vet. Microbiol. 2004, 98, 159–163. [Google Scholar] [CrossRef]
- Morozov, I.; Sirinarumitr, T.; Sorden, S.D.; Halbur, P.G.; Morgan, M.K.; Yoon, K.J.; Paul, P.S. Detection of a novel strain of porcine circovirus in pigs with postweaning multisystemic wasting syndrome. J. Clin. Microbiol. 1998, 36, 2535–2541. [Google Scholar] [CrossRef] [Green Version]
- Kawashima, K.; Katsuda, K.; Tsunemitsu, H. Epidemiological investigation of the prevalence and features of postweaning multisystemic wasting syndrome in Japan. J. Vet. Diagn. Invest. 2007, 19, 60–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.J. Porcine circovirus type 2 (PCV2): Pathogenesis and interaction with the immune system. Annu. Rev. Anim. Biosci. 2013, 1, 43–64. [Google Scholar] [CrossRef]
- Madson, D.M.; Opriessnig, T. Effect of porcine circovirus type 2 (PCV2) infection on reproduction: Disease, vertical transmission, diagnostics and vaccination. Anim. Health Res. Rev. 2011, 12, 47–65. [Google Scholar] [CrossRef] [PubMed]
- Koike, F.; Murata, S.; Oi, M.; Murakami, M. Genotypes of porcine circovirus 2 detected from healthy pigs in Japan, 2015. J. Jpn. Vet. Med. Assoc. 2017, 70, 650–654. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Nickerson, D.A.; Tobe, V.O.; Taylor, S.L. PolyPhred: Automating the detection and genotyping of single nucleotide substitutions using fluorescence-based resequencing. Nucleic Acids Res. 1997, 25, 2745–2751. [Google Scholar] [CrossRef] [Green Version]
- Gordon, D.; Abajian, C.; Green, P. Consed: A graphical tool for sequence finishing. Genome Res. 1998, 8, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Ewing, B.; Green, P. Base-calling of automated sequencer traces using Phred. II. Error probabilities. Genome Res. 1998, 8, 186–194. [Google Scholar] [CrossRef] [Green Version]
- Ewing, B.; Hillier, L.; Wendl, M.C.; Green, P. Base-calling of automated sequencer traces using Phred. I. Accuracy assessment. Genome Res. 1998, 8, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Grau-Roma, L.; Crisci, E.; Sibila, M.; Lopez-Soria, S.; Nofrarias, M.; Cortey, M.; Fraile, L.; Olvera, A.; Segales, J. A proposal on porcine circovirus type 2 (PCV2) genotype definition and their relation with postweaning multisystemic wasting syndrome (PMWS) occurrence. Vet. Microbiol. 2008, 128, 23–35. [Google Scholar] [CrossRef] [Green Version]
- Opriessnig, T.; Ramamoorthy, S.; Madson, D.M.; Patterson, A.R.; Pal, N.; Carman, S.; Meng, X.J.; Halbur, P.G. Differences in virulence among porcine circovirus type 2 isolates are unrelated to cluster type 2a or 2b and prior infection provides heterologous protection. J. Gen. Virol. 2008, 89, 2482–2491. [Google Scholar] [CrossRef]
- Groenen, M.A.; Archibald, A.L.; Uenishi, H.; Tuggle, C.K.; Takeuchi, Y.; Rothschild, M.F.; Rogel-Gaillard, C.; Park, C.; Milan, D.; Megens, H.J.; et al. Analyses of pig genomes provide insight into porcine demography and evolution. Nature 2012, 491, 393–398. [Google Scholar] [CrossRef]
- Grau-Roma, L.; Fraile, L.; Segales, J. Recent advances in the epidemiology, diagnosis and control of diseases caused by porcine circovirus type 2. Vet. J. 2011, 187, 23–32. [Google Scholar] [CrossRef]
- Rodriguez-Arrioja, G.M.; Segales, J.; Calsamiglia, M.; Resendes, A.R.; Balasch, M.; Plana-Duran, J.; Casal, J.; Domingo, M. Dynamics of porcine circovirus type 2 infection in a herd of pigs with postweaning multisystemic wasting syndrome. Am. J. Vet. Res. 2002, 63, 354–357. [Google Scholar] [CrossRef] [PubMed]
- Bao, F.; Mi, S.; Luo, Q.; Guo, H.; Tu, C.; Zhu, G.; Gong, W. Retrospective study of porcine circovirus type 2 infection reveals a novel genotype PCV2f. Transbound. Emerg. Dis. 2018, 65, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Davies, B.; Wang, X.; Dvorak, C.M.; Marthaler, D.; Murtaugh, M.P. Diagnostic phylogenetics reveals a new Porcine circovirus 2 cluster. Virus Res. 2016, 217, 32–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorr, P.M.; Baker, R.B.; Almond, G.W.; Wayne, S.R.; Gebreyes, W.A. Epidemiologic assessment of porcine circovirus type 2 coinfection with other pathogens in swine. J. Am. Vet. Med. Assoc. 2007, 230, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, T.; Zhang, X.; Liu, X.; Ren, L. Co-infection of swine with porcine circovirus type 2 and other swine viruses. Viruses 2019, 11, 185. [Google Scholar] [CrossRef] [Green Version]
- Järveots, T.; Saar, T.; Põdersoo, D.; Rüütel-Boudinot, S.; Sütt, S.; Tummeleht, L.; Suuroja, T.; Lindjärv, R. Lawsonia intracellularis and Porcine Circovirus type-2 infection in Estonia. Pol. J. Vet. Sci. 2016, 19, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Divangahi, M.; Mostowy, S.; Coulombe, F.; Kozak, R.; Guillot, L.; Veyrier, F.; Kobayashi, K.S.; Flavell, R.A.; Gros, P.; Behr, M.A. NOD2-deficient mice have impaired resistance to Mycobacterium tuberculosis infection through defective innate and adaptive immunity. J. Immunol. 2008, 181, 7157–7165. [Google Scholar] [CrossRef] [Green Version]
- Hruz, P.; Zinkernagel, A.S.; Jenikova, G.; Botwin, G.J.; Hugot, J.P.; Karin, M.; Nizet, V.; Eckmann, L. NOD2 contributes to cutaneous defense against Staphylococcus aureus through α-toxin-dependent innate immune activation. Proc. Natl. Acad. Sci. USA 2009, 106, 12873–12878. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K.S.; Chamaillard, M.; Ogura, Y.; Henegariu, O.; Inohara, N.; Nuñez, G.; Flavell, R.A. Nod2-dependent regulation of innate and adaptive immunity in the intestinal tract. Science 2005, 307, 731–734. [Google Scholar] [CrossRef]
- Opriessnig, T.; Madson, D.M.; Roof, M.; Layton, S.M.; Ramamoorthy, S.; Meng, X.J.; Halbur, P.G. Experimental reproduction of porcine circovirus type 2 (PCV2)-associated enteritis in pigs infected with PCV2 alone or concurrently with Lawsonia intracellularis or Salmonella typhimurium. J. Comp. Pathol. 2011, 145, 261–270. [Google Scholar] [CrossRef]
- Inohara, N.; Ogura, Y.; Fontalba, A.; Gutierrez, O.; Pons, F.; Crespo, J.; Fukase, K.; Inamura, S.; Kusumoto, S.; Hashimoto, M.; et al. Host recognition of bacterial muramyl dipeptide mediated through NOD2. Implications for Crohn’s disease. J. Biol. Chem. 2003, 278, 5509–5512. [Google Scholar] [CrossRef] [Green Version]
- Cooney, R.; Baker, J.; Brain, O.; Danis, B.; Pichulik, T.; Allan, P.; Ferguson, D.J.; Campbell, B.J.; Jewell, D.; Simmons, A. NOD2 stimulation induces autophagy in dendritic cells influencing bacterial handling and antigen presentation. Nat. Med. 2010, 16, 90–97. [Google Scholar] [CrossRef]
- Homer, C.R.; Richmond, A.L.; Rebert, N.A.; Achkar, J.P.; McDonald, C. ATG16L1 and NOD2 interact in an autophagy-dependent antibacterial pathway implicated in Crohn’s disease pathogenesis. Gastroenterology 2010, 139, 1630–1641.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, T.; Kitani, A.; Murray, P.J.; Wakatsuki, Y.; Fuss, I.J.; Strober, W. Nucleotide binding oligomerization domain 2 deficiency leads to dysregulated TLR2 signaling and induction of antigen-specific colitis. Immunity 2006, 25, 473–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noguchi, E.; Homma, Y.; Kang, X.; Netea, M.G.; Ma, X. A Crohn’s disease-associated NOD2 mutation suppresses transcription of human IL10 by inhibiting activity of the nuclear ribonucleoprotein hnRNP-A1. Nat. Immunol. 2009, 10, 471–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagener, J.; Malireddi, R.K.; Lenardon, M.D.; Köberle, M.; Vautier, S.; MacCallum, D.M.; Biedermann, T.; Schaller, M.; Netea, M.G.; Kanneganti, T.D.; et al. Fungal chitin dampens inflammation through IL-10 induction mediated by NOD2 and TLR9 activation. PLoS Pathog. 2014, 10, e1004050. [Google Scholar] [CrossRef] [Green Version]
- Caruso, R.; Warner, N.; Inohara, N.; Nuñez, G. NOD1 and NOD2: Signaling, host defense, and inflammatory disease. Immunity 2014, 41, 898–908. [Google Scholar] [CrossRef] [Green Version]
- Hugot, J.P.; Chamaillard, M.; Zouali, H.; Lesage, S.; Cézard, J.P.; Belaiche, J.; Almer, S.; Tysk, C.; O’Morain, C.A.; Gassull, M.; et al. Association of NOD2 leucine-rich repeat variants with susceptibility to Crohn’s disease. Nature 2001, 411, 599–603. [Google Scholar] [CrossRef]
- Ogura, Y.; Bonen, D.K.; Inohara, N.; Nicolae, D.L.; Chen, F.F.; Ramos, R.; Britton, H.; Moran, T.; Karaliuskas, R.; Duerr, R.H.; et al. A frameshift mutation in NOD2 associated with susceptibility to Crohn’s disease. Nature 2001, 411, 603–606. [Google Scholar] [CrossRef] [PubMed]
- Waites, K.B.; Talkington, D.F. Mycoplasma pneumoniae and its role as a human pathogen. Clin. Microbiol. Rev. 2004, 17, 697–728, table of contents. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinkai, H.; Terada, K.; Toki, D.; Tohno, M.; Uenishi, H. Q969R polymorphism in NLRP3 is associated with immune responses to vaccination against bacterial infections in pigs. Anim. Sci. J. 2018, 89, 1043–1050. [Google Scholar] [CrossRef]
- Tohno, M.; Shinkai, H.; Toki, D.; Okumura, N.; Tajima, K.; Uenishi, H. Identification of the Q969R gain-of-function polymorphism in the gene encoding porcine NLRP3 and its distribution in pigs of Asian and European origin. Immunogenetics 2016, 68, 693–701. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Birth Year | Male | Female | Total | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Total | Dead | Dead/ Total (%) | Total | Dead | Dead/ Total (%) | Total | Dead | Dead/Total (%) | |
| 2008 | 128 | 10 | 7.8 | 130 | 7 | 5.4 | 258 | 17 | 6.6 |
| 2009 | 136 | 30 | 22.1 | 129 | 23 | 17.8 | 265 | 53 | 20.0 |
| 2010 | 116 | 39 | 33.6 | 101 | 34 | 33.7 | 217 | 73 | 33.6 |
| 2011 | 86 | 10 | 11.6 | 78 | 7 | 9.0 | 164 | 17 | 10.4 |
| 2012 | 96 (71) | 12 (5) | 12.5 (7.0) | 100 (80) | 6 (5) | 6.0 (6.3) | 196 (151) | 18 (10) | 9.2 (6.6) |
| 2013 | 142 (142) | 8 (8) | 5.6 (5.6) | 153 (153) | 9 (9) | 5.9 (5.9) | 295 (295) | 17 (17) | 5.8 (5.8) |
| Birth Year | AA | AC | CC | Frequency of a Allele (%) |
|---|---|---|---|---|
| 2008 | 20 | 75 | 29 | 46.4 |
| 2009 | 23 | 65 | 44 | 42.9 |
| 2010 | 10 | 58 | 31 | 39.4 |
| 2011 | 22 | 36 * | 23 | 49.4 |
| 2012 | 8 | 30 ** | 62 †† | 23.0 ‡‡ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, K.; Shinkai, H.; Yoshioka, G.; Matsumoto, T.; Tanaka, J.; Hayashi, N.; Kitazawa, H.; Uenishi, H. NOD2 Genotypes Affect the Symptoms and Mortality in the Porcine Circovirus 2-Spreading Pig Population. Genes 2021, 12, 1424. https://doi.org/10.3390/genes12091424
Suzuki K, Shinkai H, Yoshioka G, Matsumoto T, Tanaka J, Hayashi N, Kitazawa H, Uenishi H. NOD2 Genotypes Affect the Symptoms and Mortality in the Porcine Circovirus 2-Spreading Pig Population. Genes. 2021; 12(9):1424. https://doi.org/10.3390/genes12091424
Chicago/Turabian StyleSuzuki, Kasumi, Hiroki Shinkai, Gou Yoshioka, Toshimi Matsumoto, Junji Tanaka, Noboru Hayashi, Haruki Kitazawa, and Hirohide Uenishi. 2021. "NOD2 Genotypes Affect the Symptoms and Mortality in the Porcine Circovirus 2-Spreading Pig Population" Genes 12, no. 9: 1424. https://doi.org/10.3390/genes12091424