
Genetic Subtypes and Natural Resistance Mutations in HCV Genotype 4 Infected Saudi Arabian Patients

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Population
2.2. Serum Samples
2.3. Amplification and NS3, NS5A, and NS5B Sequencing
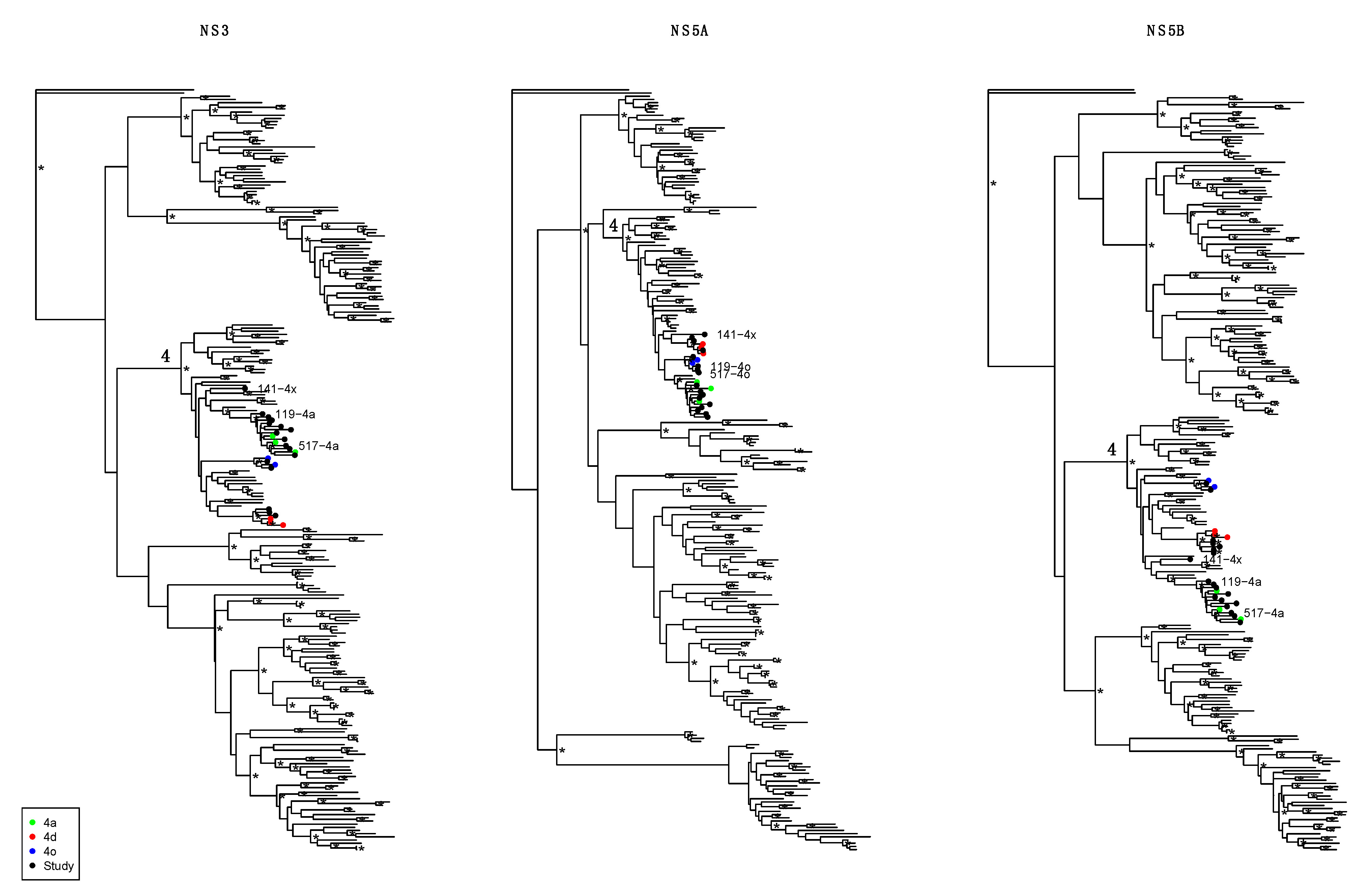
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Word Health Organization. Global Hepatitis Reports. 2017. Available online: https://apps.who.int/iris/handle/10665/255016 (accessed on 10 September 2021).
- Mohd Hanafiah, K.; Groeger, J.; Flaxman, A.D.; Wiersma, S.T. Global epidemiology of hepatitis C virus infection: New estimates of age-specific antibody to HCV seroprevalence. Hepatology 2013, 57, 1333–1342. [Google Scholar] [CrossRef]
- Messina, J.P.; Humphreys, I.; Flaxman, A.; Brown, A.; Cooke, G.S.; Pybus, O.G.; Barnes, E. Global distribution and prevalence of hepatitis C virus genotypes. Hepatology 2015, 61, 77–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borgia, S.M.; Hedskog, C.; Parhy, B.; Hyland, R.H.; Stamm, L.M.; Brainard, D.M.; Subramanian, M.G.; McHutchison, J.G.; Mo, H.; Svarovskaia, E.; et al. Identification of a Novel Hepatitis C Virus Genotype From Punjab, India: Expanding Classification of Hepatitis C Virus Into 8 Genotypes. J. Infect. Dis. 2018, 218, 1722–1729. [Google Scholar] [CrossRef] [Green Version]
- Gower, E.; Estes, C.; Blach, S.; Razavi-Shearer, K.; Razavi, H. Global epidemiology and genotype distribution of the hepatitis C virus infection. J. Hepatol. 2014, 61, S45–S57. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Moneim, A.S.; Bamaga, M.S.; Shehab, G.M.; Abu-Elsaad, A.A.; Farahat, F.M. HCV infection among Saudi popula-tion: High prevalence of genotype 4 and increased viral clearance rate. PLoS ONE 2012, 7, e29781. [Google Scholar]
- Vallet, S.; The ANRS AC11 HCV Group; Viron, F.; Henquell, C.; Le Guillou-Guillemette, H.; Lagathu, G.; Abravanel, F.; Trimoulet, P.; Soussan, P.; Schvoerer, E.; et al. NS3 protease polymorphism and natural resistance to protease inhibitors in French patients infected with HCV genotypes 1–5. Antivir. Ther. 2011, 16, 1093–1102. [Google Scholar] [CrossRef] [Green Version]
- Bertoli, A.; Sorbo, M.C.; Aragri, M.; Lenci, I.; Teti, E.; Polilli, E.; Di Maio, V.C.; Gianserra, L.; Biliotti, E.; Masetti, C.; et al. Prevalence of Single and Multiple Natural NS3, NS5A and NS5B Resistance-Associated Substitutions in Hepatitis C Virus Genotypes 1–4 in Italy. Sci. Rep. 2018, 8, 8988. [Google Scholar] [CrossRef]
- Andrade, E.; Rocha, D.; Fontana-Maurell, M.; Costa, E.; Ribeiro, M.; De Godoy, D.T.; Ferreira, A.G.P.; Tanuri, A.; Brindeiro, R.; Alvarez, P. Resistance mutations of NS3 and NS5b in treatment-naïve patients infected with hepatitis C virus in Santa Catarina and Rio Grande do Sul states, Brazil. Genet. Mol. Biol. 2020, 43, e20180237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Childs, K.; Davis, C.; Cannon, M.; Montague, S.; Filipe, A.C.D.S.; Tong, L.; Simmonds, P.; Smith, D.; Thomson, E.; Dusheiko, G.; et al. Suboptimal SVR rates in African patients with atypical genotype 1 subtypes: Implications for global elimination of hepatitis C. J. Hepatol. 2019, 71, 1099–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, R.; Ahovegbe, L.; Niebel, M.; Shepherd, J.; Thomson, E.C. Non-epidemic HCV genotypes in low- and middle-income countries and the risk of resistance to current direct-acting antiviral regimens. J. Hepatol. 2021, 75, 462–473. [Google Scholar] [CrossRef]
- Fourati, S.; Rodriguez, C.; Hézode, C.; Soulier, A.; Ruiz, I.; Poiteau, L.; Chevaliez, S.; Pawlotsky, J. Frequent Antiviral Treatment Failures in Patients Infected with Hepatitis C Virus Genotype 4, Subtype 4r. Hepatology 2019, 69, 513–523. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.; Smith, D.; Vaughan-Jackson, A.; Magri, A.; Barnes, E.; Simmonds, P. Efficacy of NS5A inhibitors against unusual and potentially difficult-to-treat HCV subtypes commonly found in sub-Saharan Africa and South East Asia. J. Hepatol. 2020, 73, 794–799. [Google Scholar] [CrossRef] [PubMed]
- Castera, L.; Forns, X.; Alberti, A. Non-invasive evaluation of liver fibrosis using transient elastography. J. Hepatol. 2008, 48, 835–847. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, A.; Giordanengo, V.; Dunais, B.; De Salvador-Guillouet, F.; Perbost, I.; Durant, J.; Pugliese, P.; Joulié, A.; Roger, P.M.; Rosenthal, E. Virological response and resistance mutations to NS3/4A inhibitors in hepatitis C virus-human immunodeficiency virus coinfection. World J. Hepatol. 2015, 7, 2177–2183. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, M.; Faleo, G.; Mohamed, A.M.F.; Morella, S.; Bruno, S.R.; Tundo, P.; Fiore, J.R.; Santantonio, T.A. Resistance Associated Mutations in HCV Patients Failing DAA Treatment. New Microbiol. 2021, 44, 12–18. [Google Scholar]
- Di Maio, V.C.; Cento, V.; Lenci, I.; Aragri, M.; Rossi, P.; Barbaliscia, S.; Melis, M.; Verucchi, G.; Magni, C.F.; Teti, E.; et al. HCV Italian Resistance Network Study Group. Mul-ticlass HCV resistance to direct-acting antiviral failure in real-life patients advocates for tailored second-line therapies. Liver Int. 2017, 37, 514–528. [Google Scholar] [CrossRef] [PubMed]
- Sarrazin, C.; Dvory-Sobol, H.; Svarovskaia, E.S.; Doehle, B.P.; Pang, P.S.; Chuang, S.-M.; Ma, J.; Ding, X.; Afdhal, N.; Kowdley, K.V.; et al. Prevalence of Resistance-Associated Substitutions in HCV NS5A, NS5B, or NS3 and Outcomes of Treatment With Ledipasvir and Sofosbuvir. Gastroenterology 2016, 151, 501–512.e1. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Lethiec, F.; Duroux, P.; Gascuel, O. PHYML Online--a web server for fast maximum likelihood-based phylogenetic inference. Nucleic Acids Res. 2005, 33, W557–W559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2018, 35, 526–528. [Google Scholar] [CrossRef]
- Sorbo, M.C.; Cento, V.; Di Maio, V.C.; Howe, A.Y.; Garcia, F.; Perno, C.F.; Ceccherini-Silberstein, F. Hepatitis C virus drug resistance associated substitutions and their clinical relevance: Update 2018. Drug Resist. Updates 2018, 37, 17–39. [Google Scholar] [CrossRef] [PubMed]
- Gaballah, A.; Metwally, D.E.; Elsheredy, A.G.; Shawki, M.A.; Mahmoud, O.A.E.K. Characterization of Drug–resistance Profiles to Directly Acting Agents in Hepatitis C virus Naive Patients. Egypt. J. Med. Microbiol. 2018, 27, 29–39. [Google Scholar]
- Peres-da-Silva, A.; de Almeida, A.J.; Lampe, E. Mutations in hepatitis C virus NS3 protease domain associated with re-sistance to specific protease inhibitors in antiviral therapy naïve patients. Arch. Virol. 2010, 155, 807–811. [Google Scholar] [CrossRef] [PubMed]
- Di Biagio, A.; Taramasso, L.; Cenderello, G. Treatment of hepatitis C virus genotype 4 in the DAA era. Virol. J. 2018, 15, 180. [Google Scholar] [CrossRef] [PubMed]
- Paolucci, S.; Fiorina, L.; Piralla, A.; Gulminetti, R.; Novati, S.; Barbarini, G.; Sacchi, P.; Gatti, M.; Dossena, L.; Baldanti, F. Naturally occurring mutations to HCV protease inhibitors in treatment-naïve patients. Virol. J. 2012, 9, 245. [Google Scholar] [CrossRef] [Green Version]
- Paolucci, S.; Fiorina, L.; Mariani, B.; Gulminetti, R.; Novati, S.; Barbarini, G.; Bruno, R.; Baldanti, F. Naturally occurring resistance mutations to inhibitors of HCV NS5A region and NS5B polymerase in DAA treatment-naïve patients. Virol. J. 2013, 10, 355. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Patients | Geographical Area | Sex | Age | Risk Factors | Fibrosis | Viral Load (IU/mL) |
|---|---|---|---|---|---|---|
| 710 | Saudi | F | 61 | Blood Transfusion | F1 | 2,410,000 |
| 959 | Egypt | M | 65 | Unknown | F0 | 207,000 |
| 506 | Sudan | M | 32 | Unknown | F2 | 181,000 |
| 585 | Egypt | M | 64 | Unknown | Not available | 302,000 |
| 908 | Egypt | M | 42 | Intrafamiliar Transmission | F1 | 80,000 |
| 611 | Saudi | F | 66 | Blood Transfusion | F4 | 743,000 |
| 517 | Saudi | F | 41 | Blood Transfusion | F0 | 683,000 |
| 119 | Saudi | M | 36 | Blood Transfusion | F0 | 130,000 |
| 307 | Saudi | M | 36 | Blood Transfusion | F4 | 478,000 |
| 532 | Saudi | M | 35 | Blood Transfusion | F3 | 1,940,000 |
| 263 | Saudi | F | 65 | Surgery | F0 | 82,000 |
| 460 | Saudi | M | 32 | Unknown | F0 | 153,000 |
| 589 | Saudi | M | 34 | Blood Transfusion | F0 | 12,000 |
| 792 | Palestine | M | 73 | Unknown | F4 | 239,000 |
| 141 | Saudi | M | 64 | Unknown | F4 | 750,000 |
| 082 | Saudi | F | 59 | Unknown | F4 | 495,000 |
| 234 | Saudi | F | 33 | Blood Transfusion | F0 | 134,000 |
| Patients | GT4 Subtype | NS3 | NS5A | NS5B |
|---|---|---|---|---|
| 710 | GT 4a | A61S; S101A; A102S; | K44R; V53M; K56T; I99V; | N213T |
| I114V; I134T; R150A | L28M | |||
| 959 | GT 4a | A61S; S101A; A102S; | K44R; V53M; K56T; I99V | N213T |
| I114V; I134T; R150A | ||||
| 506 | GT 4a | A61S; S101A; A102S; | K44R; V53M; K56T; I99V | N213T |
| I114V; I134T; R150A | ||||
| 585 | GT 4a | A61S; S101A; A102S; | K44R; V53M; K56T; I99V | N213T |
| I114V; I134T; R150A | ||||
| 908 | GT 4a | A61S; S101A; 102S; I114V; | K44R; V53M; K56T; I99V | N213T |
| I134T; R150A; V170I | ||||
| 611 | GT 4o | L14F; V28A; T95A; S98T; | M56I; E62N; K107E; | R100K; A130T |
| R149H | I121V; S127F; L158I; | |||
| C174S | ||||
| 517 | GT 4a/GT 4o/GT 4a | A61S; S101A; A102S; | M56I; E62N; I121V; S127F | N213T |
| I114V; I134T; R150A; | ||||
| 119 | GT 4a/GT 4o/GT 4a | A61S; S101A; A102S; | M56I; E62N; I121V; S127F | N213T; |
| I114V; I134T; R150A; V170I | ||||
| 307 | GT 4o | A61S; S101A; A102S; | M56I; E62N; I121V; S127F | R100K; A130T |
| I114V; I134T; R150A; V170I | ||||
| 532 | GT 4d | T95S; | D105N; D126E, A164T; | R127L; T130N |
| L168M | ||||
| 263 | GT 4a | A61S; S101A; A102S; | K44R; V53M; K56T; I99V | N213S |
| I114V; I134T; R150V | ||||
| 460 | GT 4a | A61S; S101A; A102S; | K44R; V53M; K56T; I99V; | N213T |
| I114V; I134T; R150A | L30R | |||
| 589 | GT 4a | A61S; S101A; A102S; | K44R; V53M; K56T; I99T | N213T |
| I114V; I134T; R150A | ||||
| 792 | GT 4d | T95A; | D105N; D126E, A164T; | R127L; T130N |
| L168M | ||||
| 141 | unknown | A101S; T122N; S125A; | D126E; A164Q; L168M; | N62S; T131N; |
| V151A; A166E | L28M; M31L | E202D; K212R | ||
| 082 | GT 4a | A61S; S101A; A102S; | K44R; V53M; K56T; I99V | N213T |
| I114V; I134T; R150A | ||||
| 234 | GT 4d | T95A; | D105N; D126E, A164P; | R127L; |
| L168M | T130I/V |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Stefano, M.; Ismail, M.H.; Leitner, T.; Faleo, G.; Elmnan Adem, S.A.; Elamin, M.O.M.E.; Eltreifi, O.; Alwazzeh, M.J.; Fiore, J.R.; Santantonio, T.A. Genetic Subtypes and Natural Resistance Mutations in HCV Genotype 4 Infected Saudi Arabian Patients. Viruses 2021, 13, 1832. https://doi.org/10.3390/v13091832
Di Stefano M, Ismail MH, Leitner T, Faleo G, Elmnan Adem SA, Elamin MOME, Eltreifi O, Alwazzeh MJ, Fiore JR, Santantonio TA. Genetic Subtypes and Natural Resistance Mutations in HCV Genotype 4 Infected Saudi Arabian Patients. Viruses. 2021; 13(9):1832. https://doi.org/10.3390/v13091832
Chicago/Turabian StyleDi Stefano, Mariantonietta, Mona H. Ismail, Thomas Leitner, Giuseppina Faleo, Saada A. Elmnan Adem, Mohamed O. M. E. Elamin, Obeidi Eltreifi, Marwan J. Alwazzeh, Jose R. Fiore, and Teresa A. Santantonio. 2021. "Genetic Subtypes and Natural Resistance Mutations in HCV Genotype 4 Infected Saudi Arabian Patients" Viruses 13, no. 9: 1832. https://doi.org/10.3390/v13091832