
Role of Phytochemicals in Cancer Chemoprevention: Insights



Laser Research Centre, Faculty of Health Sciences, University of Johannesburg, Doornfontein, Johannesburg 2028, South Africa
*
Author to whom correspondence should be addressed.
Antioxidants 2021, 10(9), 1455; https://doi.org/10.3390/antiox10091455
Submission received: 16 August 2021
/
Revised: 2 September 2021
/
Accepted: 3 September 2021
/
Published: 14 September 2021
(This article belongs to the Special Issue Antioxidant and Chemopreventive Activity of Natural Compounds)
Abstract
:Cancer is a condition where the body cells multiply in an uncontrollable manner. Chemoprevention of cancer is a broad term that describes the involvement of external agents to slow down or suppress cancer growth. Synthetic and natural compounds are found useful in cancer chemoprevention. The occurrence of global cancer type varies, depending on many factors such as environmental, lifestyle, genetic etc. Cancer is often preventable in developed countries with advanced treatment modalities, whereas it is a painful death sentence in developing and low-income countries due to the lack of modern therapies and awareness. One best practice to identify cancer control measures is to study the origin and risk factors associated with common types. Based on these factors and the health status of patients, stage, and severity of cancer, type of treatment is decided. Even though there are well-established therapies, cancer still stands as one of the major causes of death and a public health burden globally. Research shows that most cancers can be prevented, treated, or the incidence can be delayed. Phytochemicals from various medicinal plants were reported to reduce various risk factors associated with different types of cancer through their chemopreventive role. This review highlights the role of bioactive compounds or natural products from plants in the chemoprevention of cancer. There are many plant based dietary factors involved in the chemoprevention process. The review discusses the process of carcinogenesis and chemoprevention using plants and phytocompounds, with special reference to five major chemopreventive phytocompounds. The article also summarizes the important chemopreventive mechanisms and signaling molecules involved in the process. Since the role of antioxidants in chemoprevention is inevitable, an insight into plant-based antioxidant compounds that fight against this dreadful disease at various stages of carcinogenesis and disease progression is discussed. This will fill the research gap in search of chemopreventive natural compounds and encourage scientists in clinical trials of anticancer agents from plants.
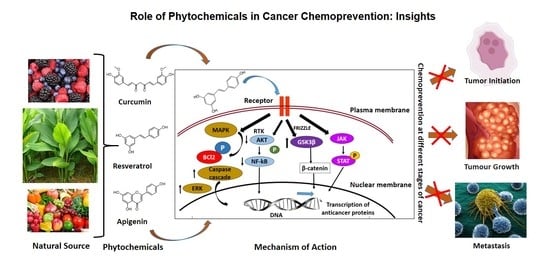
1. Introduction
Cancer is a condition where cells anywhere in the body behave abnormally and multiply in an uncontrollable manner. There are over 200 different types of cancer. The symptoms and signs of cancer vary based on the size, location, stage, and cancer types. The cancer type and staging can be determined by biopsy outcomes. The treatment protocols vary based on the type and severity of the tumor. Surgery, chemotherapy, and radiation are the widely used conventional cancer treatment modalities. There are many complementary and alternative treatment modalities for cancer including herbal medicine. Chemoprevention is an approach with great potential in controlling the incidence of cancer. Chemoprevention works in different ways to stop, delay, and control cancer incidence and progression [1]. Many products from natural and synthetic agents were employed to prevent carcinogenesis or metastasis of cancer [2]. Chemoprevention research has noticeably increased with advanced insight into carcinogenesis and the detection of potent molecular targets to hinder the process of carcinogenesis. Worldwide, every year around 11 million people are diagnosed and out of which 6.7 million people die due to cancer. Breast, lung, and colorectal cancers are the most common cancers diagnosed globally and the mortality rate of lung, stomach, and liver cancers are the highest among all cancers [3]. The drawbacks of modern cancer treatments, disease severity, increased mortality rate, and loss of patient quality of life upon treatment show the need for a new and alternative herbal approach in disease control and prevention.
Chemoprevention is one of the growing areas of anticancer research with the focus on various interventions such as nutritional factors, biological and pharmacological. There are three different approaches involved in cancer chemoprevention: primary, secondary, and tertiary chemoprevention. First one involves interventions intended to prevent healthy individuals from showing high-risk features, such as genetic predisposition to prevent the development of certain cancer. Second type is mainly developed for treating premalignant tumors to prevent cancer progression (example: colon polyps, skin actinic keratosis, cervical dysplasia, lung metaplasia). Third approach is to support patients with cancer history to prevent further cancer relapse or the development of new primary tumors [3]. Nutrients, and other dietary factors are importantly linked with the risk of developing various cancer types. Manson in 2003 reported that around 35% of global cancer mortality rates are directly associated with dietary factors [4]. Recently, dietary polyphenolic compounds were reported to possess anticancer benefits and other related pharmacological activities and received a great deal of attention for their health benefits. Polyphenolic compounds are found in majority of fruits and vegetables. Based on the structural complexity, chemical nature and structure of the polyphenols, they are categorized into 10 classes with over 8000 compounds [5]. Among the pool of dietary polyphenols, flavonoids and phenolic acids are the most common and account for about 60 and 30%, while phenolic acids, flavonoids, stilbenes, and lignans are the most widely occurring plant polyphenols. This review discusses the concept of chemoprevention with reference to the plant-based chemopreventive agents, their mechanism of action, related signaling molecules, and the role of antioxidant compounds.
2. Cancer Chemoprevention: Rapidly Growing Field
The characteristic feature of cancer cells is their ability to grow faster than normal cells. Most of the chemotherapy agents are designed in such a way that they target these fast-growing cells and block, kill, or slow down their growth. Nevertheless, these chemotherapy drugs adversely affect or kill normal healthy cells too. Due to this reason, the patient experiences severe side effects, and the therapy efficacy will be reduced or limited. Carcinogenesis progresses through various molecular mechanisms and signalling molecules. The chemoprevention of cancer fosters the utilization of natural and artificial mediators to disrupt carcinogenesis by stopping or defeating the precise molecular signaling mechanisms [6]. These mediators are mainly classified into two: blocking and suppressing agents. Chemoprevention using medicinal plant-derived compounds is an advanced cancer research field that concentrates on prevention through nutritional mediations. Development of a chemopreventive agent from a basic biological observation to clinically efficient anticancer compounds or agent is challenging. The chemoprevention process must always be safe and lengthy, and the compounds used must be acceptable for long-term administration to healthy and elderly patients, as well as those with comorbidities.
There are several epidemiological studies that reported that consistent fruit and vegetable intake significantly reduces the possibility of the incidence of many cancers [7]. Bioactive compounds from medicinal plants or from herbal diets, dietary phytochemicals lately showed their significant role in cancer chemoprevention [6], leading to the development of a new and alternative method of cancer prevention and therapy [8]. Several phytochemicals underwent clinical trials to investigate their possible cancer chemopreventive effects [9]. Wattenberg in 1985 classified chemopreventive agents into inhibitors of carcinogen formation, blocking agents, and suppressing agents. The first type prevents the carcinogen formation from the precursors, and the second type prevents carcinogen-induced mutations by blocking specific metabolic carcinogen activation pathways, and it also enhances the detoxification process by trapping specific reactive oxygen species (ROS). Finally, the third type prevents the tumor progression by stopping cell proliferation and differentiation, and by inducing apoptosis, autophagy, necrosis, etc. Most of the bioactive compounds of plant origin block the cancer development process by any of these pathways. Chemopreventive compounds were also shown to enhance the effectiveness of conventional chemotherapeutic drugs. The important aim of cancer chemoprevention is to reduce the early-stage cancer risk and to prevent the secondary spreading of tumors [10,11].
Carcinogenesis is a multistep process involving multiple genes and several genetic alterations that are expected in a normal healthy cell to develop into a tumor cell. The process of carcinogenesis, or cancer cell formation, is often divided into three steps such as initiation, promotion, and progression. The number of hereditary changes engaged in these different steps is not yet fully verified. The first step entails the initiation of an irreversibly distorted cell and is often associated with a mutation and different initiation pathways. During the second stage, the initiated mutated cells expand and form a visible mass of cells, which are probably a nonmalignant lesion. The promotion stage certainly involves epigenetic factors that affect the multiplication of the initiated cells. The detailed mechanism involved in the second stage of carcinogenesis is not well-understood. The final product of promotion is usually nonmalignant or benign cells, or sometimes preneoplastic cells. These benign cells undergo a couple of extra genetic alterations during the progression stage into neoplastic cells. The final stage in carcinogenesis, i.e., progression involves in the formation of malignant tumors from nonmalignant benign tumors, are different from other earlier two steps. Stem cells play a vital role in the initiation of carcinogenesis through various factors such as physical, chemical, or biological, including viruses. Such initiated cells will then be exposed to a promoting factor to stimulate the full neoplastic cell formation, and the sequential steps are important in the malignant transformation of preneoplastic cells [12,13]. The carcinogenesis process in a multicellular animal is the consequence of various chemical, physical, biological, or genomic changes in the cells. Though carcinogenesis is mostly driven by mutation, several other factors are also involved in this development. Chemoprevention is a pharmacological intervention approach to arrest or reverse the process of carcinogenesis. Chemoprevention influences and stops the carcinogenesis process in each stage as shown in Figure 1.
3. Role of Plant Polyphenols in Chemoprevention of Cancer
Many plant-derived products are used for the treatment and management of various diseases. Several purified plant products are available in the market for use, and such products are either entire plants or parts of the plant available in powder, capsule, or liquid forms. These plant products can be consumed as food, tablets, prepared in tea, used as gels to apply over the skin, and sometimes can be used in bathing water. Even though plant products were used for centuries in anticancer and cancer chemopreventive roles, investigation of active compounds from plants drove interest in the last few decades. Nevertheless, appropriate prudence must be held during the use of such herbal products in cancer patients because some of the herbal products may interfere with the conventional treatments patients undergo, or the dosages of the plant products may cause adverse effects on the patients’ health [14,15,16,17].
One of the main drawbacks of anticancer chemotherapies is the resistance developed during the continued therapies. Recent research is focused on this area to find out the resistance genes and drug resistance processes. There are some reports on the use of herbal products in combination with conventional anticancer drugs, and the outcome showed these herbal combinations resensitize the chemotherapy resistance established after the continued use of the anticancer chemotherapeutic drugs. Therefore, these combination therapies using herbal product anticancer drug conjugates improve the healing efficacy and result in the desired treatment outcomes. The herbal combination chemotherapies also showed significantly reduced undesirable side effects of synthetic drug-based chemotherapies [18,19]. The dietetic phytochemicals are widely used in the chemoprevention process, and this idea of cancer prevention is achieving increased research focus due to the diverse chemical components in plants, complex structure of chemicals, characteristic biological effects, cost-effectiveness, easy accessibility, and reduced or less toxic side effects. Phytocompounds can alter various molecular signaling pathways and impart protective role in cancer chemoprevention. The dietary chemopreventive phytochemicals not only played an important role in cancer prevention, but they were also employed as a primary competitor as a natural lead compound in the development of potent cancer chemotherapeutic drugs. Polyphenolic compounds among the phytochemicals have a significant role in cancer therapies and prevention due to their prominent role in interfering carcinogenesis at the initiation, promotion, progression stages, etc. Phytochemicals lead to the alteration of proteins in different signal transduction pathways and integrate with distinct molecular signals to exert the definitive chemopreventive and/or chemotherapeutic role [20].
Environmental factors including diet are one of the major causes of the development of cancer. Nutrients, and other dietary elements are strongly correlated to the risk of developing various cancer types. Research investigations showed that one-third of cancer mortality rates are associated with diet and nutrition [4]. Phenolic acids, flavonoid compounds, stilbenes, lignans, etc., are the commonly appearing polyphenolic compounds in plants; among them, flavonoids and phenolic acids account for about 90% of dietetic polyphenols. Based on the complexity of molecular and chemical structure, there are around 4,000 different types of flavonoids reported. The dietary sources of some of the important chemopreventive polyphenolic compounds are shown in Figure 2. Various clinical studies showed the positive correlation between the intake of polyphenolic compounds or flavonoids as food or drinks rich in polyphenols and reduced cancer incidence, lowered the risk of developing different cancer types, and decreased cancer relapse [21,22,23,24,25,26,27]. Most of the potent chemopreventive polyphenols disrupt or reverse carcinogenesis through initiation, promotion, and progression steps via the network of intracellular signaling molecules [28]. Therefore, the anticancer and chemopreventive role of nutritive and dietary polyphenols are based on the combination role as cytoprotective and cytotoxic efficacies. They protect the normal healthy cells, while they kill or are toxic to the premalignant and neoplastic or malignant cells.
Phenolic compounds of plants alter carcinogenesis through various pathways and their efficacies will vary based on the type of tissue and dosages used. Though there are some clinical studies reported on the role of polyphenols, many of the anticarcinogenesis roles of nutritive polyphenolic compounds are studied in in vitro and in vivo models. Studies showed the safe doses of green tea extract are daily up to 4.2 g or 1.0 g thrice daily intake to the patients with advanced stage of cancers, and 3 g per day for patients with advanced lung cancers. In these studies, the toxicity observed was directly linked to the caffeine (neurological and gastrointestinal effects) doses [29,30]. Another study reported the anticancer activity of green tea based on the expression of molecular biomarkers. The decreased levels of PSA and urinary 8-hydroxydeoxyguanosine markers were noted in liver and prostate cancer patients with a 6 g/m2 intake of green tea daily [31,32]. Some of the common medicinal plants used in the chemoprevention of cancer, their therapeutic role, and cancer types treated or prevented are listed in Table 1.
4. Natural Chemopreventive Agents in Clinical Setting
Many natural compounds were widely examined for their possible use in the prevention of cancers over the years. The increasing volume of in vitro and in vivo data on the cancer chemopreventive and chemotherapeutic outcomes of plant-derived compounds urged scientists to do clinical trials focusing on the pharmacokinetics, efficiency, and safety of the phytocompounds.
4.1. Curcumin
Curcumin is a potential natural compound that originated from Curcuma longa and is widely used for cancer chemoprevention. Apoptosis induction, inhibition of molecular signals, free radical scavenging, and inhibition of inflammatory responses on tumor microenvironment are the various molecular mechanisms by which curcumin exerts chemoprevention of cancer. Curcumin exhibits ideal chemopreventive features, such as low toxic effects, cost-effectiveness, and easy accessibility. In different continents, especially in Asia, the powder form of rhizome of the Curcuma plant was used widely as a spice and as a coloring and flavoring agent in many foods, and it also showed anti-inflammatory effects. Diferuloylmethane (1,7-bis- (4- hydroxy-3-methoxyphenyl)-1,6-heptadine-3,5-dione) or curcumin is the chief pigment of turmeric, with strong antioxidant and anti-inflammatory activities [60]. Turmeric contains many bioactive compounds of which curcuminoids such as curcumin, tetrahydrocurcumin, bisdemethoxycurcumin, and demethoxycurcumin are common, and the levels vary based on the species type, farming, and rhizome treating conditions. Tetrahydrocurcumin is the key naturally occurring curcuminoids compound, and it is of great interest in anticancer research due to its increased solubility in water. They are chemically stable and showed maximum bioavailability with strong antioxidant activity. Anticancer activities of tetrahydrocurcumin are linked to many molecular pathways such as they modulate oxidative stress created in cells, reduce inflammation and proliferation, and induce immunity and cell death [61].
The major molecular targets of curcumin that help in cancer cell death are cyclooxygenase-2 (COX-2), nuclear factor kappa B (NF-kB), tumor necrosis factor-alpha (TNF-a), and cyclin D1. The molecular targets exert substantial anti-inflammatory and anti-tumorigenic properties in different clinical and preclinical studies [62,63]. Many in vitro studies also showed different mechanisms of curcumin in the antiproliferation of cancer cells. Cyclin D1 and CDK-4 expressions were significantly reduced in breast and skin cancers after curcumin treatments [64]. Curcumin also downregulates the expression of angiogenic genes including VEGF, angiopoietin and MMP-9, and MMP-3; this effect was mediated by AP-1 inhibitory action [65,66] a promising in cancer therapeutic and chemopreventive approach. The chemopreventive action of curcumin is altogether mediated by several biochemical and molecular pathways that regulate apoptotic cell death and the role of different types of transcription factors and enzymes.
4.2. Resveratrol
Grapes, berries, and many other plant products exhibit strong polyphenolic compounds are rich in resveratrol (3, 5, 4′-trihydroxy-trans-stilbene). Not only in cancer research but many other clinical research data also showed that resveratrol has significant effects with many age-related diseases, such as diabetes, neurodegenerative disorders, arthritis, and coronary and pulmonary illnesses. Reports suggest that resveratrol modifies all the different steps in carcinogenesis, such as initiation, promotion, and progression. They alter various signaling mechanisms to reduce the cancer cell multiplication and progression, inducing programmed cell death and reducing inflammatory responses and angiogenesis, and also stopping the metastatic spread of tumors. They modulate multiple signaling pathways of the cell cycle, inflammation, and apoptosis [67,68,69,70]. One of the promising roles of resveratrol is how it significantly decreases the toxic side effects associated with therapies and enhances the cancer therapeutic effects. It can also be employed in the treatment of some autoimmune diseases [71,72,73,74].
Side effects of chemotherapies were always a serious complication in the application of chemotherapy drugs in clinical practice. Resveratrol is a safe and potent natural compound that showed a multifunctional role which has a protective role against chemotherapy side effects with strong chemopreventive and chemotherapy properties [75,76]. Resveratrol significantly reduced the accumulation of arsenic and decrease the arsenic-induced toxicities in renal cells. They also inhibit the arsenic trioxide-induced oxidative stress, and decline the arsenic accumulation in hepatic cells and increase antioxidant enzyme effects to prevent arsenic-induced toxicity in liver [77]. The promising antioxidant role of resveratrol also helps to protect acetaminophen-induced liver toxicity, as well as cisplatin-induced gastrointestinal tract disorders by improving the tubular dilation and cell vacuolization of kidney tubes [78,79]. The topical application of resveratrol inhibits UVB-mediated skin edema and reduces hydrogen peroxide production and leukocyte infiltration in mice. This study led found that the long-term application of resveratrol before and after the treatment significantly reduced the tumor incidence or delayed the start of oncogenesis, while the short-term application led to the inhibition of cellular proliferation [80,81]. Some studies reported that regulatory cell cycle signaling molecules and apoptotic inhibitory proteins expression after resveratrol treatments showed inhibition of photocarcinogenesis [82,83].
4.3. Apigenin
A flavonoid rich in fruits and vegetables with chemopreventive action is apigenin (4′, 5, 7, -trihydroxyflavone). Antiangiogenic properties of apigenin are linked to the regulation of signaling pathways, induction of apoptosis, prevention of cancer cell transformation, and cell cycle arrest [84,85,86,87]. Apigenin has an inhibitory effect on tobacco smoke-related cancers and HPV infections. They substantially reduced HPV-altered human prostate cells by cell cycle alteration and apoptotic induction [88], and this effect was due to the inhibiting action of apigenin on NNK, a tobacco-specific carcinogen through focal adhesion kinase (FAK) and extracellular regulated protein kinase (ERK) signals [89]. Furthermore, they specifically inhibit HPV-positive HeLa and SiHa cells while reduced toxic effects are observed in HPV-negative C33A and HaCaT cells [90].
There are many in vivo studies that reported the chemopreventive role of apigenin. The dosages, different modes of administration, and frequencies of treatments, etc., were studied on various animal models. The in vivo results showed the inhibition of phosphoinositide 3-kinase (PI3K)/Akt/Forkhead box O-signaling pathway [91], and apigenin reduced Her2/neu protein expression in breast cancer mice models [92]. In a xenograft tumor model, the intake of apigenin also reduced the serum-immunoglobulin F-I concentration, and stimulated apoptotic cell death and cell cycle arrest at different phases [93]. The interruption of apigenin in the NF-kB pathway is responsible for prostate cancer inhibition [94]. A clinical study was conducted to investigate the absorption of apigenin systemically after the intake of parsley rich diet, the results of which showed improved biological antioxidant levels, such as SOD and GR, whereas the GPx, CAT activities seemed to be reduced in blood cells [95]. Apigenin also showed other bioactivities, such as reduction in plasma level LDLs, platelet aggregation, and cell proliferation [28,95,96].
4.4. Epigallocatechin Gallate
Green tea is rich in a strong antioxidant chemopreventive compound epigallocatechin gallate (EGCG). The chemical structure of EGCG consists of three attached heterocyclic rings, and electron delocalization led to the free radical scavenging [97]. Tea catechins including EGCG have redox properties and react with reactive oxygen species. The metal chelating abilities of EGCG helps in the prevention of ROS production and catechin oxidation process [98,99]. The chemical structure of EGCG imparts antioxidant properties and the air oxidation process with ions and leads to the generation of unstable dimers of catechin. Though EGCG has many health benefits, it showed low bioavailability due to increased instability, low distribution in the digestive tract, and active efflux properties [100,101,102,103]. Due to these drawbacks, the effect and application of EGCG in human trials is reduced. Recently, many clinical trials were conducted on controlled pharmacokinetic factors to choose highly suitable dosages and administration modes for proper EGCG intervention [103].
Several in vitro studies conducted to investigate the different mechanisms of EGCG chemoprevention. EGCG shown prominent regulatory effects on various signalling pathways such as JAK/STAT, MAPK, PI3K/AKT, Wnt, Notch, NF-κB and AP-1 [104,105,106,107,108,109,110,111]. The bioactive compounds of green tea also established tumour suppressor activities, including p53, p21, p16 and Rb [112,113,114,115]; these activities play a vital role in the prevention of cancer [116,117]. Another important property of EGCG is its regulatory role on several receptors in host; for example, EGCG control 67-KDa laminin receptor activity, formerly recognized as extracellular matrix as part of the translational assembly and as surface receptors [118,119]. Furthermore, they control the androgen receptor activity in prostate tumors [120] and the estrogen receptor action in the mammary tumors [121].
4.5. Genistein
Legumes produce an important isoflavone known as genistein (4′, 5, 7-Trihydroxyisoflavone), commonly called phytoestrogens with a similar structure to human estrogen. They are ketone or nonketone polyhydroxy polyphenolic compounds of leguminous plants [122]. Genistein exhibit a wide range of biological properties including cancer chemoprevention. Epidemiological studies showed that in Asian countries, the incidence of certain types of cancers such as breast and prostate cancers is lower compared to that of western countries due to diets rich in soy and soy products [123]. The report says the genistein was first isolated from Genista tinctoria, Fabaceae family, in 1899, but the most abundant amounts of genistein are sourced from legumes [124].
Biochemical properties of genistein mediate many health benefits including antitumor. Genistein phytoestrogen competes with 17β-estradiol in estrogen binding tests in vitro cell cultures. The report says genistein blocks the proliferation and multiplication of estrogen and androgen receptor-positive and negative mammary and prostate tumor cells in vitro [125]. Genistein blocks PTK signaling mechanism via protein-tyrosine kinase (PTK) inhibition, which indirectly suppresses the proliferation of cancer cells [126]. Other important molecular cancer targets of genistein are topoisomerase I and II [127], 5α-reductase [128], and protein histidine kinase [129]. Inhibition of these targets also imparts antiproliferative and apoptotic induction of cancer cells. Genistein showed strong antioxidant potential and protects the cells from ROS action via free radical scavenging, and it also inhibits the stress response-related genes expression and reduces their role in carcinogenesis [130]. Genistein is a strong inhibitor of cell survival pathways, such as NF-κB and Akt [131], and these inhibitory effects play a major role in the apoptosis induction in genistein-treated cells.
Some of the most well-known plant-derived chemopreventive agents, their chemical structure and chemopreventive mechanisms are listed in Table 2.
5. Phytochemicals Induced Chemopreventive Mechanisms
5.1. Antiangiogenesis and Metastasis
The physiological process by which new blood vessels are formed from the old ones is called angiogenesis. This process is a vital step in the growth, invasion, and spread of tumors. There are many phytochemicals including phenolics that act as angiogenesis inhibitors to block tumor cell growth and multiplication. The preliminary mechanisms of antiangiogenic properties of phenolic compounds are well understood in many in vitro and in vivo studies. The antiangiogenic properties of ellagic acid, EGCG, genistein, and anthocyanin-rich berry extracts are via suppression of vascular endothelial growth factor, vascular endothelial growth factor receptor-2, platelet derived growth factor, platelet-derived growth factor receptor, hypoxia-inducible factor 1a, and matrix metalloproteases. The blocking of epidermal growth factor receptor, vascular endothelial growth factor receptor, and platelet-derived growth factor receptor phosphorylation also impart in antiangiogenesis [136,137,138,139].
Many phenolics show a variance of effects on antiangiogenic factors of tumor and healthy normal cells. Green tea phenolic EGCG can chelate ferrous ions and prevent hypoxia-inducible factor-1a-induced cancer cell growth. In vitro studies showed a decrease in hypoxia-inducible factor-1-mediated transcription and hypoxia-inducible factor-1a protein level under normal oxygen levels after treatment with EGCG in prostate tumor cells [140]. Kaempferol, quercetin, myricetin, and galanin suppress the vascular endothelial growth factor-mediated human umbilical vein endothelial cells (HUVEC) tubular formation and block the U937 cell adhesion to HUVECs to prevent the angiogenesis process [141]. Flavonoids, such as naringin, rutin, apigenin, genistein, and kaempferol, decreased the vascular endothelial growth factor from human mammary cancer cells [142]. Metastasis is the process of spreading cancer cells to distal organs through lymph nodes. This process interchange with the degradation of extracellular matrix, proteolysis, cell adhesion, cell migration, angiogenesis, and invasion [143]. Many dietary polyphenolic compounds possess anti-invasive and antimetastatic properties by interfering with the tumor cell adhesion and migration via various mechanisms. However, their exact molecular mechanism and signal transduction pathways are yet to be discovered [4].
5.2. Apoptosis and Cell Cycle Arrest
Apoptosis is the most common programmed cell death and acts as the therapeutic target for many cancers. Several dietary chemopreventive compounds (resveratrol, quercetin, EGCG, curcumin, apigenin, chrysin, silymarin, and ellagic acid) showed the inhibition of carcinogenesis via apoptosis induction [4,28]. Cancer cells are more sensitive to these compounds than healthy normal cells [144,145,146]. EGCG mediated apoptosis induction in sarcoma cells was by the cell cycle arrest at G2/M phase, suppression of Bcl-2 and myc, expression of p53 and Bax, whereas the expression of other important targets of apoptosis such as p21, p27, Bcl-xL, mdm2, and cyclin D1 remains unchanged in sarcoma cells [147]. However, the overexpression of p21, p53 and Bax were observed along with the activation of caspases-3 and -9 and PARP cleavage to achieve apoptosis in prostate cancer cells. EGCG primarily activates and promotes the cell cycle arrest and apoptosis through p53-mediated signal transduction along with the effect of p21 and Bax [148]. Another study showed that a black tea phenolic called theaflavin stimulated the fragmentation of DNA, caspases-3 and -8, Bax expression, and Bcl-2 downregulation to achieve apoptosis [149]. Remarkably, theaflavin also stimulated apoptosis by p53 expression, altering the Bax/Bcl-2 ratio, increased release of cytochrome c from mitochondria, and activates the caspases-9 and -3 expressions in prostate cancer cells [150]. An important class of flavonoid compound, anthocyanins, showed an ability to decrease cell proliferation in colon cancer cells in a dose-dependent manner. Anthocyanins-mediated apoptotic induction in colon cancer cells was due to the DNA fragmentation and imbalance of Bax and Bcl-2 expressions. However, some bioflavonoid compounds (rutin, epicatechin, chlorogenic acid, or p-hydroxybenzoic acid) showed no growth inhibitory effect in cells [145]. Delphinidin is an anthocyanidin plant pigment that inhibited vascular endothelial growth factor-mediated cell movement and proliferation through the cell cycle arrest at G0/G1 phase. During this process, the expression of p21 and p27 increased whereas the levels of cyclin D1, cyclin A significantly reduced [151]. Additionally, the delphinidin-mediated antiproliferative effect was also caused by the early activation of extracellular signal-regulated protein kinase1/2, upregulation of caveolin- 1, and downregulation of Ras [152].
6. Signaling Molecules Involved in Cancer Chemoprevention
There are several factors leading to the progression of cancer, including complex interactions of signaling molecules [153]. With advancement in science and research technologies, these pathways are being unveiled to track the conditions leading to cancer progression. Molecular oncology has identified key mechanism of actions with a precise understanding of gene expression, suppression, mutations, etc., [154,155]. Several medicine and drugs performed wonders in cancer therapy. However, cancer incidence and progression can be prevented by modifying lifestyle and dietary changes [28]. These dietary components as chemopreventive agents attained much interest in resolving many medical conditions.
6.1. Phytochemicals Modulating Signaling Molecules
Naturally occurring chemopreventive agents, especially from plants, inspired the scientific community to conduct many experiments to investigate the underlying mechanisms of action against cancer prevention [156]. Plant-based compounds or phytochemicals interfere with cellular mechanisms including action of antioxidant enzymes, reducing oxidative stress, cell cycle arrest, apoptosis, necrosis, autophagy, and inhibit expression of genes suppressing cancer progression and lead to activation of oncogenes, modulating signaling pathways and inhibiting angiogenesis and metastasis. All these and many other pathways have targeted molecules that activate chemoprevention of cancer [157,158].
These target molecules include kinases, receptors, caspases, tumor-suppressor proteins, transcriptional factors, miRNAs, and cyclins. Due to internal or external factors, oncogenes get activated inducing carcinogenesis. However, cells initiate their self-defense mechanisms through apoptosis, cell cycle arrest, autophagy etc., by altering cell signaling pathways. Phytochemicals can accelerate these defense mechanisms on cellular matrix, cytoplasm and nucleus through multiple molecules controlling these pathways which decide cell fate [71,159,160].
6.2. Major Signaling Pathways
MAPK Pathways. Phytochemicals can target the extracellular signal regulated kinase (ERK) and mitogen-activated protein kinase (MAPK) pathway, which essentially regulates cellular growth and survival. These natural compounds from plants potentially control cancer progression through various mechanisms [160,161]. The major plant compounds that were reported to induce apoptosis through MAPK and ERK pathways include ursolic acid, kaempferol, resveratrol, gingerol, sulforaphane, genistein, and isothiocyanates [161,162,163].
Akt Signaling Pathways. In cancer control and progression, Akt/PI3 signaling pathway plays a pivotal role. Levels of epidermal growth factor (EGF) regulates series of molecular mechanisms including activation of NF-κB and phosphorylation of Akt leading to resistance to apoptosis and uncontrolled cell proliferation, while downstream, it leads to the regulation of caspases, Bcl-2, glycogen synthase kinase 3-beta (GSK3β), and mammalian target of rapamycin (mTOR) [164]. Alkaloids and phenolics significantly contributed to controlling the expression of these factors. Resveratrol, luteolin, luteolin, apigenin, flavone, sulforaphane, and curcumin were reported to induce anticancer activities through cell cycle arrest and apoptosis, inhibit Akt/PI3K signaling, proapoptosis, activation of FOXO3a (forkhead box O3), antiproliferation, and anti-invasion [165,166,167,168,169,170,171].
JAK/STAT Signaling Pathways. Activation of Janus kinases (JAKs) subsequently phosphorylates signal transducer and activator of transcriptions (STATs), and translocates to the nucleus, where it controls the transcription of p53, Bcl-2, cyclin D, and interlukin-6 (IL-6) involved in cell death, proliferation, and apoptosis [172]. Compounds isolated from plants significantly induce cell death in various cancer forms by inhibiting activity of JAK/STAT signaling and activating apoptotic cascades [173].
Wnt/β-Catenin Signaling Pathways. Common cancers, including that of breast, lung, colon, blood, ovary, skin, and brain, are frequently associated with abnormal signaling of Wnt/β-catenin pathway [174]. Activation of this pathway is initiated with the binding of Wnt-protein to frizzled family transmembrane receptors and accumulation of β-catenin in the nucleus leading to the activation of transcriptional factors regulating cell proliferation, survival, and migration [175,176]. Phytochemicals, such as curcumin, resveratrol and epigallocatechin-3-gallate (EGCG), inhibit the translocation and accumulation of β-catenin in the nucleus by activation of glycogen synthase kinase 3 (GSK3) [174,177,178].
Tumor Suppressor p53. Similar to other signaling pathways, p53 accounts for anticancer activities in cell. This tumor suppressor protein facilitates activation of apoptotic cascades [179]. Several phytochemicals, including resveratrol, Quercetin, EGCG, and piceatannol, were reported to upregulate p53, downregulate Akt, and induce cellular apoptosis and elevated cell cycle rest [179,180,181]. Another significant contribution is with p53, MAPK, and JNK pathway crosslink [182].
Curcumin and ursolic acid reportedly promote autophagy, wherein the cells shut down irregular cell proliferation and progression. In cancer cells, autophagy is induced through downregulation Akt/mTOR pathways [183]. Dietary phytochemicals also regulates of antitumor functions in tissues by regulating inflammation, angiogenesis, invasion, and metastasis [184]. Phytochemicals, hence through these signaling pathways, suppress cancer development (Figure 3).
7. Role of Antioxidants in Chemoprevention of Cancer
Many factors promote cancer development, and one among them are free radicals. These free radicals fundamentally belong to reactive nitrogen species (RNS) and reactive oxygen species (ROS). Singlet oxygen, superoxide radicals, hydrogen peroxide, and nitric oxide (NO) are known to impart potential impact on cellular mechanisms [185]. These free radicals cause damage to lipid bilayer disrupting the cellular membrane, also tampering with amino acids and DNA, thereby activating a series of enzymatic and nonenzymatic reactions inside the cell, leading to irregular gene expressions [186]. In cells, some of the ROS produced are neutralized by endogenous antioxidants in the body. However, imbalance between pro and antioxidants results in accumulation of free radicals promoting malignancy and metastasis.
The oxidative damage of DNA induces gene mutations and chromosomal aberrations, leading to failure in cell cycle arrest in G1 and diminishing DNA repair capacity. This will promote replication errors, inactivation of tumor suppressor gene, activation of oncogenes, and malignancy. The depletion of normal cell population is more evident with free radical induced resistance in cells due to incorrect DNA replication and chromosomal rearrangements leading to carcinogenesis. Studies reported activation of C-Raf-1 and K-RAS oncogenes by hydroxyl radicals. This activation initiates through N-terminal deletion of genes and GC base pair mutations [187].
To counteract these free radicals and promote cell death in tumor tissues, antioxidants are being studied widely as they neutralize free radicals that enhance cancer progression. Endogenous and exogenous antioxidants are the major forms of classified based on their source. This includes enzymes, phenolics, carotenoids, minerals, and vitamins [188]. Despite the availability of synthetic antioxidants, antioxidants from natural sources are abundant with immense health benefits. Dietary antioxidants are among the most explored ones as they can be included in daily diet which can mitigate free radical induced oxidative stress and disease [189]. Plant-derived antioxidants showed explicit potential in preventing diseases, and hence, researchers’ interest towards identifying novel antioxidants from plants are increasing.
Cancer and its subtypes are a challenging concern for most of the medical experts and researchers all over the world. However, plant-derived compounds, especially those with antioxidant properties to counteract free radicals and oxidative stress, are promising solutions in cancer chemoprevention. Moreover, these phytochemicals are proved to possess none-to-minimal toxicity towards normal cells, making them preferred agents in chemoprevention.
8. Possible Actions of Antioxidant Phytochemicals
Several phytochemicals induce antimigratory, anti-invasive, and antiproliferative effects on cancer cells. Free radicals induced carcinogenesis and activation of nuclear factor erythroid 2-related factor 2 (Nrf2), resulting in the accumulation of ROS and increased oxidative stress. These results in translocation of Nrf2 into nucleus once dissociated from Kelch-like ECH-associated protein 1 (KEAP1). Meanwhile, KEAP1 undergoes proteasomal degradation. In the nucleus, Nrf2 binds with Musculoaponeurotic Fibrosarcoma (MAF), initiating transcription of genes activating superoxide dismutase (SOD), glutathione-S transferases (GSTs), quinone oxidoreductase 1 (NQO1), heme oxygenase-1 (HMOX-1), suppressing free radical-induced stress, as well as DNA and protein damage, and downregulating several other signaling pathways responsible for carcinogenesis [190]. In the following paragraphs, we discuss a few important antioxidant phytochemicals with proven chemopreventive roles against cancer.
Curcumin, one of the major alkaloids found in plants of cucurbitaceae family, is known to reduce ROS-induced tumorigenesis and simultaneously protect normal tissues from ROS-mediated DNA damage [191]. Curcumin showed the ability to modulate several signaling pathways by reducing free radical load in cancer cells. Extracellular signal regulated protein kinase (ERK1/2) is one such signaling molecule involved in reverting ROS production, tumor invasion, and migration [192]. Nuclear factor kappa B (NFkB) is also activated in cancer cells in response to ERK1/2 pathway [193]. Endogenous enzymes such as catalase (CAT), superoxide dismutase (SOD), and heme oxygenase-1 are important in fighting free radical imbalance in the cells. Phenolic compounds were potentially proven to be an agent in activating these enzymes inhibiting the production of ROS in several organs and reducing metastases by altering levels of matrix metalloproteinase (MMPs), Vascular endothelial growth factor (VEGF), and Protein kinase (PKC) levels [194]. Garattini et al. studied the effective role of all trans retinoic acids and other retinoids in the prevention of breast cancer. This was attained by modulating several growth factor pathways such as epidermal growth factors including (EGF), insulin growth factor (IGF), mitogen activated protein kinase (MAPKs) and Akt signaling pathways [195]. Angiogenesis, being one of the major promoters of cancer proliferation, became a target of cancer prevention. Retinoids prevent this process of angiogenesis in cancer populations [196]. These and other classes of phytochemicals are known to eliminate cancer stem cells responsible for cancer relapse [197], inducing apoptosis and modulating gene and immune functions [198]. The preventive role of tocopherols were studied in many cell lines inducing antiproliferative, apoptotic, cell cycle arrest, and proapoptotic activity in pancreas, colorectum, lung, breast, prostate, liver, bladder, stomach, and glioblastoma [199,200,201]. Shin-Kang et al., [202] studied the mechanism behind these properties and concluded that tocols act on several signaling cascades, including inhibition of ERK, Akt, cyclins, NF-κB, P13K/Akt signaling, TGF-β, and EGF-2, upregulation of caspases 8, 6, and 9, and suppression of cell proliferation [199].
One of the major classes of compounds found in plants with known wide range of properties are phenolic compounds (PCs). They were shown to kill cancer cell lines in many ways. Similar to phytochemical actions discussed above, phenolic compounds also exert their preventive role though cell cycle arrest, apoptosis, and most importantly, scavenging free radicals responsible for carcinogenesis and its progression [203,204,205]. Phytochemicals involved in cancer chemoprevention were always a topic of controversy due to their pro-oxidant action. However, compounds from plants are sought after to effectively prevent cancer. The role of such natural compounds are shown in Table 3.
9. Role of Natural Products in CYP450 Inhibition
Several natural compounds or phytochemicals were proven to exert inhibitory activity towards different isoforms of P450 protein family as potential anticancer and chemopreventive therapeutic agents. Phytochemicals, such as polycyclic aromatic hydrocarbons (PAH), naphthoquinones and anthraquinones, stilbenoids, flavonoids, coumarins, alkaloids etc., inhibit the P450 enzyme activity. Mechanistically, these phytochemicals are classified as reversible (competitive or noncompetitive), quasi-irreversible, and irreversible inhibitors of P450 enzyme. Phytochemicals, such as polycyclic aromatic hydrocarbons, flavones, and coumarins, exhibit very high P450 inhibitory activity. Nonsubstituted PAHs, 5-hydroxy-2-methyl-NQ, 2-methyl-NQ, 2-hydroxy-NQ, and 1,4-NQ, 2,4,3′,5′-Tetramethoxystilbene, protoberberine alkaloids, cannabinoids etc., inhibit P450 enzymes in a competitive manner. The well-known anti-inflammatory and anticancer agent berberine acts as a noncompetitive inhibitor of P450 CYP1 family enzymes. Linderane and thujopsene terpenoids show an irreversible inhibition of CYP2 enzymes, while methylenedioxyphenyl lignans act as an irreversible inhibitor of CYP34A [214,215,216]. Resveratrol exhibits various P450 inhibitory mechanisms such as it inhibits human P450 CYP1A1 activity in a mixed-type inhibition (competitive–noncompetitive), while also acting as a noncompetitive inhibitor against P450 CYP1B1 and a mechanism-based inhibitor of cytochrome P450 CYP3A4 [214]. A unique inhibition type known as mechanism-based inactivation (MBI) is also observed in certain natural compounds. In MBI the original substrate of the enzyme undergoes additional metabolic conversion, which chemically activates the substrate to irreversibly binds and inactivates the enzyme. The natural mechanism-based inhibitors of P450 enzymes include anthraquinone derivatives (4-amino-1-chloro-3-methylanthraquinone), stilbenoids (rhapontigenin), flavones (7-hydroxyflavone, 3-flavon propargyl ether), coumarins (coriandrin), alkaloids (furafylline), etc. However, application of MBIs as anticancer agents is not recommended, as irreversible and permanent inhibition of P450 enzyme causes disturbed pharmacokinetics and pharmacodynamics of drugs metabolized by P450. This causes a significant increase in drug concentration in blood circulation, leading to toxicity [214,215]. Other than direct inhibition, resveratrol also acts as a competitive antagonist of arylhydrocarbon receptor (AhR) ligands such as dioxin by blocking the activation of dioxin-inducible genes and inhibits cytochrome P450 1A1 expression [217,218]. In an in vivo study, resveratrol was shown to abrogate the benzo[a]pyrene-induced P450 1A1 expression and its subsequent effects on B[a]P-DNA adducts formation and apoptosis induction [219].
10. Conclusions
Chemoprevention of cancer using plant-based compounds became a preferred approach in cancer management. Exploration of new chemopreventive phytocompounds became a central goal of anticancer research, and it also helps with finding new therapeutic targets. Specific dietary compounds explained in this review may constitute potent cancer chemopreventive agents, and the consumption of food that involves such bioactive compounds was shown to have protective and therapeutic effects on various types of cancers. The polyphenolic compounds from plants have an immunomodulatory role that identifies and destroys cancer cells by antiangiogenic effects. Chemopreventive drugs enhance chemo and radiotherapy efficacies via multiple signal transduction pathways. Since oxidative stress plays a vital role in the pathogenesis of many cancers, the antioxidant effect of dietary phenolic compounds might act as a promising strategy to prevent cancer. Dietary antioxidant phytochemicals are abundant in plants representing different classes of compounds, which exert various mechanisms of actions on tumors. Hence, chemoprevention through diets rich in plant-based antioxidants show great potential in reducing the risk factors associated with cancer progression.
Author Contributions
Conceptualization, B.P.G.; writing—original draft preparation, B.P.G., R.C. and H.A.; writing—review and editing, B.P.G., R.C. and H.A. funding acquisition, H.A. All authors have read and agreed to the published version of the manuscript.
Funding
South African Research Chairs Initiative of the Department of Science and Technology and National Research Foundation of South Africa (Grant No 98337).
Acknowledgments
This work is based on the research supported by the South African Research Chairs Initiative of the Department of Science and Technology and National Research Foundation of South Africa (Grant No 98337). The authors sincerely thank the University of Johannesburg, the National Laser Centre (CSIR-NLC), and the National Research Foundation (NRF) of South Africa for their financial support.
Conflicts of Interest
The authors declare that there is no conflict of interest.
References
- De Flora, S.; Ferguson, L.R. Overview of Mechanisms of Cancer Chemopreventive Agents. Mutat. Res. 2005, 591, 8–15. [Google Scholar] [CrossRef]
- Sporn, M.B. Approaches to Prevention of Epithelial Cancer during the Preneoplastic Period. Cancer Res. 1976, 36, 2699–2702. [Google Scholar]
- Bonovas, S.; Tsantes, A.; Drosos, T.; Sitaras, N.M. Cancer Chemoprevention: A Summary of the Current Evidence. Anticancer Res. 2008, 28, 1857–1866. [Google Scholar]
- Manson, M.M. Cancer Prevention—the Potential for Diet to Modulate Molecular Signalling. Trends Mol. Med. 2003, 9, 11–18. [Google Scholar] [CrossRef]
- Ramos, S. Effects of Dietary Flavonoids on Apoptotic Pathways Related to Cancer Chemoprevention. J. Nutr. Biochem. 2007, 18, 427–442. [Google Scholar] [CrossRef] [Green Version]
- Landis-Piwowar, K.R.; Iyer, N.R. Cancer Chemoprevention: Current State of the Art. Cancer Growth Metastasis 2014, 7, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Vallinas, M.; González-Castejón, M.; Rodríguez-Casado, A.; Ramírez de Molina, A. Dietary Phytochemicals in Cancer Prevention and Therapy: A Complementary Approach with Promising Perspectives. Nutr. Rev. 2013, 71, 585–599. [Google Scholar] [CrossRef]
- Wattenberg, L.W. Chemoprevention of Cancer. Cancer Res. 1985, 45, 1–8. [Google Scholar] [CrossRef]
- Hosseini, A.; Ghorbani, A. Cancer Therapy with Phytochemicals: Evidence from Clinical Studies. Avicenna J. Phytomed. 2015, 5, 84–97. [Google Scholar] [PubMed]
- Sharma, G.; Tyagi, A.K.; Singh, R.P.; Chan, D.C.F.; Agarwal, R. Synergistic Anti-Cancer Effects of Grape Seed Extract and Conventional Cytotoxic Agent Doxorubicin against Human Breast Carcinoma Cells. Breast Cancer Res. Treat. 2004, 85, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lev-Ari, S.; Strier, L.; Kazanov, D.; Madar-Shapiro, L.; Dvory-Sobol, H.; Pinchuk, I.; Marian, B.; Lichtenberg, D.; Arber, N. Celecoxib and Curcumin Synergistically Inhibit the Growth of Colorectal Cancer Cells. Clin. Cancer. Res. 2005, 11, 6738–6744. [Google Scholar] [CrossRef] [Green Version]
- Kabirai, A.; Chahar, A.; Chahar, N.; Gupta, J. Chemical Carcinogenesis: A Brief Review on Mechanism & Metabolism. J. Oral Med. Oral Surg. Oral Pathol. Oral Radiol. 2020, 6, 120–124. [Google Scholar] [CrossRef]
- Malarkey, D.E.; Hoenerhoff, M.; Maronpot, R.R. Chapter 5-Carcinogenesis: Mechanisms and Manifestations. In Haschek and Rousseaux’s Handbook of Toxicologic Pathology, 3rd ed.; Haschek, W.M., Rousseaux, C.G., Wallig, M.A., Eds.; Academic Press: Boston, MA, USA, 2013; pp. 107–146. [Google Scholar] [CrossRef]
- Saha, S.K.; Sikdar, S.; Mukherjee, A.; Bhadra, K.; Boujedaini, N.; Khuda-Bukhsh, A.R. Ethanolic Extract of the Goldenseal, Hydrastis Canadensis, Has Demonstrable Chemopreventive Effects on HeLa Cells in Vitro: Drug-DNA Interaction with Calf Thymus DNA as Target. Environ. Toxicol. Pharmacol. 2013, 36, 202–214. [Google Scholar] [CrossRef]
- Bishayee, K.; Paul, A.; Ghosh, S.; Sikdar, S.; Mukherjee, A.; Biswas, R.; Boujedaini, N.; Khuda-Bukhsh, A.R. Condurango-Glycoside-A Fraction of Gonolobus Condurango Induces DNA Damage Associated Senescence and Apoptosis via ROS-Dependent P53 Signalling Pathway in HeLa Cells. Mol. Cell. Biochem. 2013, 382, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Bishayee, K.; Khuda-Bukhsh, A.R. Graveoline Isolated from Ethanolic Extract of Ruta Graveolens Triggers Apoptosis and Autophagy in Skin Melanoma Cells: A Novel Apoptosis-Independent Autophagic Signaling Pathway. Phytother. Res. 2014, 28, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Sikdar, S.; Mukherjee, A.; Ghosh, S.; Khuda-Bukhsh, A.R. Condurango Glycoside-Rich Components Stimulate DNA Damage-Induced Cell Cycle Arrest and ROS-Mediated Caspase-3 Dependent Apoptosis through Inhibition of Cell-Proliferation in Lung Cancer, in Vitro and in Vivo. Environ. Toxicol. Pharmacol. 2014, 37, 300–314. [Google Scholar] [CrossRef]
- Wang, Z.J.; Chen, X.; Huang, Y.; Lam, W.K.; Lam, W.K.C.; Chow, M.S.S. Overcoming Chemotherapy Resistance with Herbal Medicines: Past, Present and Future Perspectives. Phytochem. Rev. 2014, 13, 323–337. [Google Scholar] [CrossRef]
- Mondal, J.; Bishayee, K.; Panigrahi, A.K.; Khuda-Bukhsh, A.R. Low Doses of Ethanolic Extract of Boldo (Peumus Boldus) Can Ameliorate Toxicity Generated by Cisplatin in Normal Liver Cells of Mice in Vivo and in WRL-68 Cells in Vitro, but Not in Cancer Cells in Vivo or in Vitro. J. Integr. Med. 2014, 12, 425–438. [Google Scholar] [CrossRef]
- Ramos, S. Cancer Chemoprevention and Chemotherapy: Dietary Polyphenols and Signalling Pathways. Mol. Nutr. Food Res. 2008, 52, 507–526. [Google Scholar] [CrossRef]
- Arts, I.C.W.; Jacobs, D.R.; Gross, M.; Harnack, L.J.; Folsom, A.R. Dietary Catechins and Cancer Incidence among Postmenopausal Women: The Iowa Women’s Health Study (United States). Cancer Causes Control. 2002, 13, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Key, T.J.; Sharp, G.B.; Appleby, P.N.; Beral, V.; Goodman, M.T.; Soda, M.; Mabuchi, K. Soya Foods and Breast Cancer Risk: A Prospective Study in Hiroshima and Nagasaki, Japan. Br. J. Cancer 1999, 81, 1248–1256. [Google Scholar] [CrossRef] [Green Version]
- Cutler, G.J.; Nettleton, J.A.; Ross, J.A.; Harnack, L.J.; Jacobs, D.R.; Scrafford, C.G.; Barraj, L.M.; Mink, P.J.; Robien, K. Dietary Flavonoid Intake and Risk of Cancer in Postmenopausal Women: The Iowa Women’s Health Study. Int. J. Cancer 2008, 123, 664–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knekt, P.; Järvinen, R.; Seppänen, R.; Hellövaara, M.; Teppo, L.; Pukkala, E.; Aromaa, A. Dietary Flavonoids and the Risk of Lung Cancer and Other Malignant Neoplasms. Am. J. Epidemiol. 1997, 146, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Su, L.J.; Arab, L. Tea Consumption and the Reduced Risk of Colon Cancer—Results from a National Prospective Cohort Study. Public Health Nutr. 2002, 5, 419–425. [Google Scholar] [CrossRef]
- Le Marchand, L.; Murphy, S.P.; Hankin, J.H.; Wilkens, L.R.; Kolonel, L.N. Intake of Flavonoids and Lung Cancer. J. Natl. Cancer Inst. 2000, 92, 154–160. [Google Scholar] [CrossRef] [Green Version]
- Nakachi, K.; Suemasu, K.; Suga, K.; Takeo, T.; Imai, K.; Higashi, Y. Influence of Drinking Green Tea on Breast Cancer Malignancy among Japanese Patients. Jpn. J. Cancer Res. 1998, 89, 254–261. [Google Scholar] [CrossRef]
- Surh, Y.-J. Cancer Chemoprevention with Dietary Phytochemicals. Nat. Rev. Cancer 2003, 3, 768–780. [Google Scholar] [CrossRef]
- Pisters, K.M.; Newman, R.A.; Coldman, B.; Shin, D.M.; Khuri, F.R.; Hong, W.K.; Glisson, B.S.; Lee, J.S. Phase I Trial of Oral Green Tea Extract in Adult Patients with Solid Tumors. J. Clin. Oncol. 2001, 19, 1830–1838. [Google Scholar] [CrossRef]
- Laurie, S.A.; Miller, V.A.; Grant, S.C.; Kris, M.G.; Ng, K.K. Phase I Study of Green Tea Extract in Patients with Advanced Lung Cancer. Cancer Chemother. Pharmacol. 2005, 55, 33–38. [Google Scholar] [CrossRef]
- Jatoi, A.; Ellison, N.; Burch, P.A.; Sloan, J.A.; Dakhil, S.R.; Novotny, P.; Tan, W.; Fitch, T.R.; Rowland, K.M.; Young, C.Y.F.; et al. A Phase II Trial of Green Tea in the Treatment of Patients with Androgen Independent Metastatic Prostate Carcinoma. Cancer 2003, 97, 1442–1446. [Google Scholar] [CrossRef]
- Luo, H.; Tang, L.; Tang, M.; Billam, M.; Huang, T.; Yu, J.; Wei, Z.; Liang, Y.; Wang, K.; Zhang, Z.-Q.; et al. Phase IIa Chemoprevention Trial of Green Tea Polyphenols in High-Risk Individuals of Liver Cancer: Modulation of Urinary Excretion of Green Tea Polyphenols and 8-Hydroxydeoxyguanosine. Carcinogenesis 2006, 27, 262–268. [Google Scholar] [CrossRef] [Green Version]
- Khan, K. Role of Emblica Officinalis in Medicine—A Review. Bot. Res. Int. 2009, 2, 218–228. [Google Scholar]
- Tattelman, E. Health Effects of Garlic. Am. Fam. Physician 2005, 72, 103–106. [Google Scholar]
- Shukla, Y.; Kalra, N. Cancer Chemoprevention with Garlic and Its Constituents. Cancer Lett. 2007, 247, 167–181. [Google Scholar] [CrossRef]
- Hatcher, H.; Planalp, R.; Cho, J.; Torti, F.M.; Torti, S.V. Curcumin: From Ancient Medicine to Current Clinical Trials. Cell. Mol. Life Sci. 2008, 65, 1631–1652. [Google Scholar] [CrossRef]
- Jurenka, J.S. Anti-Inflammatory Properties of Curcumin, a Major Constituent of Curcuma Longa: A Review of Preclinical and Clinical Research. Altern. Med. Rev. 2009, 14, 141–153. [Google Scholar]
- Akula, A.; Butchi Raju, A.; Prakash, G.; Challa, S. Anti-Cancer Activity of Curcuma Longa Linn. (Turmeric). J. Pharm. Res. 2011, 4, 1274–1276. [Google Scholar]
- Pettit, G.R.; Tan, R.; Ichihara, Y.; Williams, M.D.; Doubek, D.L.; Tackett, L.P.; Schmidt, J.M.; Cerny, R.L.; Boyd, M.R.; Hooper, J.N.A. Antineoplastic Agents, 325. Isolation and Structure of the Human Cancer Cell Growth Inhibitory Cyclic Octapeptides Phakellistatin 10 and 11 from Phakellia Sp. J. Nat. Prod. 1995, 58, 961–965. [Google Scholar] [CrossRef]
- Jagetia, G.C.; Nayak, V.; Vidyasagar, M.S. Evaluation of the Antineoplastic Activity of Guduchi (Tinospora Cordifolia) in Cultured HeLa Cells. Cancer Lett. 1998, 127, 71–82. [Google Scholar] [CrossRef]
- Jada, S.R.; Subur, G.S.; Matthews, C.; Hamzah, A.S.; Lajis, N.H.; Saad, M.S.; Stevens, M.F.G.; Stanslas, J. Semisynthesis and in Vitro Anticancer Activities of Andrographolide Analogues. Phytochemistry 2007, 68, 904–912. [Google Scholar] [CrossRef]
- Sousa, O.; Vieira, G.; Pinho, J.; Yamamoto, C.; Alves, M. Antinociceptive and Anti-Inflammatory Activities of the Ethanol Extract of Annona Muricata L. Leaves in Animal Models. Int. J. Mol. Sci. 2010, 11, 2067–2078. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.-T.; Yang, R.-C.; Lee, P.-N.; Yang, S.-H.; Liao, S.-K.; Chen, T.-Y.; Pang, J.-H.S. Anti-Tumor and Anti-Angiogenic Effects of Phyllanthus Urinaria in Mice Bearing Lewis Lung Carcinoma. Int. Immunopharmacol. 2006, 6, 870–879. [Google Scholar] [CrossRef] [PubMed]
- Wall, M.E.; Wani, M.C. Antineoplastic Agents from Plants. Annu. Rev. Pharmacol. Toxicol. 1977, 17, 117–132. [Google Scholar] [CrossRef] [PubMed]
- Malik, F.; Kumar, A.; Bhushan, S.; Khan, S.; Bhatia, A.; Suri, K.A.; Qazi, G.N.; Singh, J. Reactive Oxygen Species Generation and Mitochondrial Dysfunction in the Apoptotic Cell Death of Human Myeloid Leukemia HL-60 Cells by a Dietary Compound Withaferin A with Concomitant Protection by N-Acetyl Cysteine. Apoptosis 2007, 12, 2115–2133. [Google Scholar] [CrossRef]
- Shashi, B.; Jaswant, S.; Madhusudana, R.J.; Kumar, S.A.; Nabi, Q.G. A Novel Lignan Composition from Cedrus Deodara Induces Apoptosis and Early Nitric Oxide Generation in Human Leukemia Molt-4 and HL-60 Cells. Nitric Oxide 2006, 14, 72–88. [Google Scholar] [CrossRef]
- Jagetia, G.C.; Rao, S.K. Evaluation of the Antineoplastic Activity of Guduchi (Tinospora Cordifolia) in Ehrlich Ascites Carcinoma Bearing Mice. Biol. Pharm. Bull. 2006, 29, 460–466. [Google Scholar] [CrossRef] [Green Version]
- George, V.C.; Kumar, D.R.N.; Rajkumar, V.; Suresh, P.K.; Kumar, R.A. Quantitative Assessment of the Relative Antineoplastic Potential of the N-Butanolic Leaf Extract of Annona Muricata Linn. in Normal and Immortalized Human Cell Lines. Asian Pac. J. Cancer Prev. 2012, 13, 699–704. [Google Scholar] [CrossRef]
- Liu, W.; Li, S.-Y.; Huang, X.-E.; Cui, J.-J.; Zhao, T.; Zhang, H. Inhibition of Tumor Growth in Vitro by a Combination of Extracts from Rosa Roxburghii Tratt and Fagopyrum Cymosum. Asian Pac. J. Cancer Prev. 2012, 13, 2409–2414. [Google Scholar] [CrossRef] [Green Version]
- Hematulin, A.; Ingkaninan, K.; Limpeanchob, N.; Sagan, D. Ethanolic Extract from Derris Scandens Benth Mediates Radiosensitzation via Two Distinct Modes of Cell Death in Human Colon Cancer HT-29 Cells. Asian Pac. J. Cancer Prev. 2014, 15, 1871–1877. [Google Scholar] [CrossRef] [Green Version]
- Alabsi, A.M.; Ali, R.; Ali, A.M.; Harun, H.; Al-Dubai, S.A.R.; Ganasegeran, K.; Alshagga, M.A.; Salem, S.D.; Abu Kasim, N.H.B. Induction of Caspase-9, Biochemical Assessment and Morphological Changes Caused by Apoptosis in Cancer Cells Treated with Goniothalamin Extracted from Goniothalamus Macrophyllus. Asian Pac. J. Cancer Prev. 2013, 14, 6273–6280. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Xu, C.-Y.; Cai, S.-Z.; Zhou, Y.; Li, J.; Jiang, R.; Wang, Y.-P. Senescence Effects of Angelica Sinensis Polysaccharides on Human Acute Myelogenous Leukemia Stem and Progenitor Cells. Asian Pac. J. Cancer Prev. 2014, 14, 6549–6556. [Google Scholar] [CrossRef]
- Guo, W.-Q.; Li, L.-Z.; He, Z.-Y.; Zhang, Q.; Liu, J.; Hu, C.-Y.; Qin, F.-J.; Wang, T.-Y. Anti-Proliferative Effects of Atractylis Lancea (Thunb.) DC. via down-Regulation of the c-Myc/HTERT/Telomerase Pathway in Hep-G2 Cells. Asian Pac. J. Cancer Prev. 2013, 14, 6363–6367. [Google Scholar] [CrossRef] [Green Version]
- Cho, W.C.S.; Leung, K.N. In Vitro and in Vivo Anti-Tumor Effects of Astragalus Membranaceus. Cancer Lett. 2007, 252, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Valcic, S.; Timmermann, B.N.; Alberts, D.S.; Wächter, G.A.; Krutzsch, M.; Wymer, J.; Guillén, J.M. Inhibitory Effect of Six Green Tea Catechins and Caffeine on the Growth of Four Selected Human Tumor Cell Lines. Anticancer Drugs 1996, 7, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.; Parihar, L.; Parihar, P. Review on Cancer and Anticancerous Properties of Some Medicinal Plants. J. Med. Plants Res. 2011, 5, 1818–1835. [Google Scholar]
- Bharali, R.; Azad, M.R.H.; Tabassum, J. Chemopreventive Action of Boerhaavia Diffusa on DMBA-Induced Skin Carcinogenesis in Mice. Indian J. Physiol. Pharmacol. 2003, 47, 459–464. [Google Scholar] [PubMed]
- Patel, S.; Gheewala, N.; Suthar, A.; Shah, A.; Patel, S. In-Vitro Cytotoxicity Activity of Solanum Nigrum Extract against Hela Cell Line and Vero Cell Line. Int. J. Pharm. Pharm. Sci. 2008, 1, 38–46. [Google Scholar]
- Chung, M.J.; Chung, C.-K.; Jeong, Y.; Ham, S.-S. Anticancer Activity of Subfractions Containing Pure Compounds of Chaga Mushroom (Inonotus Obliquus) Extract in Human Cancer Cells and in Balbc/c Mice Bearing Sarcoma-180 Cells. Nutr. Res. Pract. 2010, 4, 177–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, B.S.; Rao, C.V. Chemoprevention of Cancer by Curcumin. In Cancer Chemoprevention: Promising Cancer Chemopreventive Agents; Kelloff, G.J., Hawk, E.T., Sigman, C.C., Eds.; Cancer Drug Discovery and Development; Humana Press: Totowa, NJ, USA, 2004; pp. 169–175. [Google Scholar] [CrossRef]
- Lai, C.-S.; Ho, C.-T.; Pan, M.-H. The Cancer Chemopreventive and Therapeutic Potential of Tetrahydrocurcumin. Biomolecules 2020, 10, 831. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Kumar, A.; Bharti, A.C. Anticancer Potential of Curcumin: Preclinical and Clinical Studies. Anticancer Res. 2003, 23, 363–398. [Google Scholar]
- Prakobwong, S.; Khoontawad, J.; Yongvanit, P.; Pairojkul, C.; Hiraku, Y.; Sithithaworn, P.; Pinlaor, P.; Aggarwal, B.B.; Pinlaor, S. Curcumin Decreases Cholangiocarcinogenesis in Hamsters by Suppressing Inflammation-Mediated Molecular Events Related to Multistep Carcinogenesis. Int. J. Cancer. 2011, 129, 88–100. [Google Scholar] [CrossRef]
- Mukhopadhyay, A.; Banerjee, S.; Stafford, L.J.; Xia, C.; Liu, M.; Aggarwal, B.B. Curcumin-Induced Suppression of Cell Proliferation Correlates with down-Regulation of Cyclin D1 Expression and CDK4-Mediated Retinoblastoma Protein Phosphorylation. Oncogene 2002, 21, 8852–8861. [Google Scholar] [CrossRef] [Green Version]
- Gururaj, A.E.; Belakavadi, M.; Venkatesh, D.A.; Marmé, D.; Salimath, B.P. Molecular Mechanisms of Anti-Angiogenic Effect of Curcumin. Biochem. Biophys. Res. Commun. 2002, 297, 934–942. [Google Scholar] [CrossRef]
- Hahm, E.-R.; Gho, Y.S.; Park, S.; Park, C.; Kim, K.-W.; Yang, C.-H. Synthetic Curcumin Analogs Inhibit Activator Protein-1 Transcription and Tumor-Induced Angiogenesis. Biochem. Biophys. Res. Commun. 2004, 321, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Baur, J.A.; Sinclair, D.A. Therapeutic Potential of Resveratrol: The In vivo Evidence. Nat. Rev. Drug Discov. 2006, 5, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Saiko, P.; Szakmary, A.; Jaeger, W.; Szekeres, T. Resveratrol and Its Analogs: Defense against Cancer, Coronary Disease and Neurodegenerative Maladies or Just a Fad? Mutat. Res. 2008, 658, 68–94. [Google Scholar] [CrossRef] [PubMed]
- Harikumar, K.B.; Aggarwal, B.B. Resveratrol: A Multitargeted Agent for Age-Associated Chronic Diseases. Cell Cycle 2008, 7, 1020–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishayee, A. Cancer Prevention and Treatment with Resveratrol: From Rodent Studies to Clinical Trials. Cancer Prev. Res. 2009, 2, 409–418. [Google Scholar] [CrossRef] [Green Version]
- Kotecha, R.; Takami, A.; Espinoza, J.L. Dietary Phytochemicals and Cancer Chemoprevention: A Review of the Clinical Evidence. Oncotarget 2016, 7, 52517–52529. [Google Scholar] [CrossRef] [Green Version]
- Berman, A.Y.; Motechin, R.A.; Wiesenfeld, M.Y.; Holz, M.K. The Therapeutic Potential of Resveratrol: A Review of Clinical Trials. NPJ. Precis. Oncol. 2017, 1, 35. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, A.L.d.B.; Monteiro, V.V.S.; Navegantes-Lima, K.C.; Reis, J.F.; Gomes, R.d.S.; Rodrigues, D.V.S.; Gaspar, S.L.d.F.; Monteiro, M.C. Resveratrol Role in Autoimmune Disease-A Mini-Review. Nutrients 2017, 9, 1306. [Google Scholar] [CrossRef] [Green Version]
- Ko, J.-H.; Sethi, G.; Um, J.-Y.; Shanmugam, M.K.; Arfuso, F.; Kumar, A.P.; Bishayee, A.; Ahn, K.S. The Role of Resveratrol in Cancer Therapy. Int. J. Mol. Sci. 2017, 18, 2589. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Xue, J.; Li, Y.; Zhang, W.; Ma, D.; Liu, L.; Zhang, Z. Resveratrol Protects against Arsenic Trioxide-Induced Nephrotoxicity by Facilitating Arsenic Metabolism and Decreasing Oxidative Stress. Arch. Toxicol. 2013, 87, 1025–1035. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, F.H.; Li, Y.; Wang, Z.; Kong, D. Cellular Signaling Perturbation by Natural Products. Cell. Signal. 2009, 21, 1541–1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Xue, J.; Ge, M.; Yu, M.; Liu, L.; Zhang, Z. Resveratrol Attenuates Hepatotoxicity of Rats Exposed to Arsenic Trioxide. Food Chem. Toxicol. 2013, 51, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, G.; Singh, A.K.; Kumar, A.; Prakash, O.; Singh, M.P. Resveratrol Modulates Pyrogallol-Induced Changes in Hepatic Toxicity Markers, Xenobiotic Metabolizing Enzymes and Oxidative Stress. Eur. J. Pharmacol. 2008, 596, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Afaq, F.; Adhami, V.M.; Ahmad, N. Prevention of Short-Term Ultraviolet B Radiation-Mediated Damages by Resveratrol in SKH-1 Hairless Mice. Toxicol. Appl. Pharmacol. 2003, 186, 28–37. [Google Scholar] [CrossRef]
- Aziz, M.H.; Afaq, F.; Ahmad, N. Prevention of Ultraviolet-B Radiation Damage by Resveratrol in Mouse Skin Is Mediated via Modulation in Survivin. Photochem. Photobiol. 2005, 81, 25–31. [Google Scholar] [CrossRef]
- Aziz, M.H.; Reagan-Shaw, S.; Wu, J.; Longley, B.J.; Ahmad, N. Chemoprevention of Skin Cancer by Grape Constituent Resveratrol: Relevance to Human Disease? FASEB J. 2005, 19, 1193–1195. [Google Scholar] [CrossRef] [Green Version]
- Adhami, V.M.; Afaq, F.; Ahmad, N. Suppression of Ultraviolet B Exposure-Mediated Activation of NF-KappaB in Normal Human Keratinocytes by Resveratrol. Neoplasia 2003, 5, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Reagan-Shaw, S.; Afaq, F.; Aziz, M.H.; Ahmad, N. Modulations of Critical Cell Cycle Regulatory Events during Chemoprevention of Ultraviolet B-Mediated Responses by Resveratrol in SKH-1 Hairless Mouse Skin. Oncogene 2004, 23, 5151–5160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Mao, Y.; Chen, H.; Lin, Y.; Hu, Z.; Wu, J.; Xu, X.; Xu, X.; Qin, J.; Xie, L. Apigenin Promotes Apoptosis, Inhibits Invasion and Induces Cell Cycle Arrest of T24 Human Bladder Cancer Cells. Cancer Cell Int. 2013, 13, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Ning, C.; Wang, Y.; Ma, T.; Huang, H.; Ge, Y.; Liu, J.; Jiang, Y. Natural Plant Flavonoid Apigenin Directly Disrupts Hsp90/Cdc37 Complex and Inhibits Pancreatic Cancer Cell Growth and Migration. J. Funct. Foods 2015, 18, 10–21. [Google Scholar] [CrossRef]
- Salmani, J.M.M.; Zhang, X.-P.; Jacob, J.A.; Chen, B.-A. Apigenin’s Anticancer Properties and Molecular Mechanisms of Action: Recent Advances and Future Prospectives. Chin. J. Nat. Med. 2017, 15, 321–329. [Google Scholar] [CrossRef]
- Yang, J.; Pi, C.; Wang, G. Inhibition of PI3K/Akt/MTOR Pathway by Apigenin Induces Apoptosis and Autophagy in Hepatocellular Carcinoma Cells. Biomed. Pharmacother. 2018, 103, 699–707. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Afaq, F.; Mukhtar, H. Selective Growth-Inhibitory, Cell-Cycle Deregulatory and Apoptotic Response of Apigenin in Normal versus Human Prostate Carcinoma Cells. Biochem. Biophys. Res. Commun. 2001, 287, 914–920. [Google Scholar] [CrossRef]
- Pham, H.; Chen, M.; Takahashi, H.; King, J.; Reber, H.A.; Hines, O.J.; Pandol, S.; Eibl, G. Apigenin Inhibits NNK-Induced Focal Adhesion Kinase Activation in Pancreatic Cancer Cells. Pancreas 2012, 41, 1306–1315. [Google Scholar] [CrossRef] [Green Version]
- Souza, R.P.; Bonfim-Mendonça, P.d.S.; Gimenes, F.; Ratti, B.A.; Kaplum, V.; Bruschi, M.L.; Nakamura, C.V.; Silva, S.O.; Maria-Engler, S.S.; Consolaro, M.E.L. Oxidative Stress Triggered by Apigenin Induces Apoptosis in a Comprehensive Panel of Human Cervical Cancer-Derived Cell Lines. Oxid. Med. Cell. Longev. 2017, 2017, 1512745. [Google Scholar] [CrossRef]
- Shukla, S.; Bhaskaran, N.; Babcook, M.A.; Fu, P.; Maclennan, G.T.; Gupta, S. Apigenin Inhibits Prostate Cancer Progression in TRAMP Mice via Targeting PI3K/Akt/FoxO Pathway. Carcinogenesis 2014, 35, 452–460. [Google Scholar] [CrossRef] [Green Version]
- Mafuvadze, B.; Liang, Y.; Besch-Williford, C.; Zhang, X.; Hyder, S.M. Apigenin Induces Apoptosis and Blocks Growth of Medroxyprogesterone Acetate-Dependent BT-474 Xenograft Tumors. Horm. Cancer 2012, 3, 160–171. [Google Scholar] [CrossRef] [Green Version]
- Shukla, S.; Fu, P.; Gupta, S. Apigenin Induces Apoptosis by Targeting Inhibitor of Apoptosis Proteins and Ku70-Bax Interaction in Prostate Cancer. Apoptosis 2014, 19, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Kanwal, R.; Shankar, E.; Datt, M.; Chance, M.R.; Fu, P.; MacLennan, G.T.; Gupta, S. Apigenin Blocks IKKα Activation and Suppresses Prostate Cancer Progression. Oncotarget 2015, 6, 31216–31232. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.E.; Young, J.F.; Daneshvar, B.; Lauridsen, S.T.; Knuthsen, P.; Sandström, B.; Dragsted, L.O. Effect of Parsley (Petroselinum Crispum) Intake on Urinary Apigenin Excretion, Blood Antioxidant Enzymes and Biomarkers for Oxidative Stress in Human Subjects. Br. J. Nutr. 1999, 81, 447–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiery-Vuillemin, A.; Nguyen, T.; Pivot, X.; Spano, J.P.; Dufresnne, A.; Soria, J.C. Molecularly Targeted Agents: Their Promise as Cancer Chemopreventive Interventions. Eur. J. Cancer 2005, 41, 2003–2015. [Google Scholar] [CrossRef] [PubMed]
- Bimonte, S.; Cascella, M.; Schiavone, V.; Mehrabi-Kermani, F.; Cuomo, A. The Roles of Epigallocatechin-3-Gallate in the Treatment of Neuropathic Pain: An Update on Preclinical in vivo Studies and Future Perspectives. Drug Des. Devel. Ther. 2017, 11, 2737–2742. [Google Scholar] [CrossRef] [Green Version]
- Gupte, A.; Mumper, R.J. Elevated Copper and Oxidative Stress in Cancer Cells as a Target for Cancer Treatment. Cancer Treat. Rev. 2009, 35, 32–46. [Google Scholar] [CrossRef]
- Khan, H.Y.; Zubair, H.; Ullah, M.F.; Ahmad, A.; Hadi, S.M. Oral Administration of Copper to Rats Leads to Increased Lymphocyte Cellular DNA Degradation by Dietary Polyphenols: Implications for a Cancer Preventive Mechanism. Biometals 2011, 24, 1169–1178. [Google Scholar] [CrossRef] [PubMed]
- Auger, C.; Mullen, W.; Hara, Y.; Crozier, A. Bioavailability of Polyphenon E Flavan-3-Ols in Humans with an Ileostomy. J. Nutr. 2008, 138, 1535S–1542S. [Google Scholar] [CrossRef] [Green Version]
- Stalmach, A.; Troufflard, S.; Serafini, M.; Crozier, A. Absorption, Metabolism and Excretion of Choladi Green Tea Flavan-3-Ols by Humans. Mol. Nutr. Food Res. 2009, 53, S44–S53. [Google Scholar] [CrossRef]
- Roowi, S.; Stalmach, A.; Mullen, W.; Lean, M.E.J.; Edwards, C.A.; Crozier, A. Green Tea Flavan-3-Ols: Colonic Degradation and Urinary Excretion of Catabolites by Humans. J. Agric. Food Chem. 2010, 58, 1296–1304. [Google Scholar] [CrossRef]
- Mereles, D.; Hunstein, W. Epigallocatechin-3-Gallate (EGCG) for Clinical Trials: More Pitfalls than Promises? Int. J. Mol. Sci. 2011, 12, 5592–5603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tedeschi, E.; Suzuki, H.; Menegazzi, M. Antiinflammatory Action of EGCG, the Main Component of Green Tea, through STAT-1 Inhibition. Ann. N. Y. Acad. Sci. 2002, 973, 435–437. [Google Scholar] [CrossRef] [PubMed]
- Sen, T.; Dutta, A.; Chatterjee, A. Epigallocatechin-3-Gallate (EGCG) Downregulates Gelatinase-B (MMP-9) by Involvement of FAK/ERK/NFkappaB and AP-1 in the Human Breast Cancer Cell Line MDA-MB-231. Anticancer Drugs 2010, 21, 632–644. [Google Scholar] [CrossRef] [PubMed]
- Masuda, M.; Wakasaki, T.; Toh, S.; Shimizu, M.; Adachi, S. Chemoprevention of Head and Neck Cancer by Green Tea Extract: EGCG-The Role of EGFR Signaling and “Lipid Raft”. J. Oncol. 2011, 2011, 540148. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, H.; Tighiouart, M.; Lee, J.E.; Shin, H.J.; Khuri, F.R.; Yang, C.S.; Chen, Z.G.; Shin, D.M. Synergistic Inhibition of Head and Neck Tumor Growth by Green Tea (−)-Epigallocatechin-3-Gallate and EGFR Tyrosine Kinase Inhibitor. Int. J. Cancer 2008, 123, 1005–1014. [Google Scholar] [CrossRef] [Green Version]
- Pahlke, G.; Ngiewih, Y.; Kern, M.; Jakobs, S.; Marko, D.; Eisenbrand, G. Impact of Quercetin and EGCG on Key Elements of the Wnt Pathway in Human Colon Carcinoma Cells. J. Agric. Food Chem. 2006, 54, 7075–7082. [Google Scholar] [CrossRef]
- Hossain, M.M.; Banik, N.L.; Ray, S.K. Survivin Knockdown Increased Anti-Cancer Effects of (−)-Epigallocatechin-3-Gallate in Human Malignant Neuroblastoma SK-N-BE2 and SH-SY5Y Cells. Exp. Cell Res. 2012, 318, 1597–1610. [Google Scholar] [CrossRef] [Green Version]
- Vézina, A.; Chokor, R.; Annabi, B. EGCG Targeting Efficacy of NF-ΚB Downstream Gene Products Is Dictated by the Monocytic/Macrophagic Differentiation Status of Promyelocytic Leukemia Cells. Cancer Immunol. Immunother. 2012, 61, 2321–2331. [Google Scholar] [CrossRef]
- Kim, H.S.; Kim, M.H.; Jeong, M.; Hwang, Y.S.; Lim, S.H.; Shin, B.A.; Ahn, B.W.; Jung, Y.D. EGCG Blocks Tumor Promoter-Induced MMP-9 Expression via Suppression of MAPK and AP-1 Activation in Human Gastric AGS Cells. Anticancer Res. 2004, 24, 747–753. [Google Scholar]
- Kang, S.U.; Lee, B.-S.; Lee, S.-H.; Baek, S.J.; Shin, Y.S.; Kim, C.-H. Expression of NSAID-Activated Gene-1 by EGCG in Head and Neck Cancer: Involvement of ATM-Dependent P53 Expression. J. Nutr. Biochem. 2013, 24, 986–999. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, N.; Agarwal, C.; Agarwal, R. Differential Responses of Skin Cancer-Chemopreventive Agents Silibinin, Quercetin, and Epigallocatechin 3-Gallate on Mitogenic Signaling and Cell Cycle Regulators in Human Epidermoid Carcinoma A431 Cells. Nutr. Cancer 2001, 39, 292–299. [Google Scholar] [CrossRef]
- Nandakumar, V.; Vaid, M.; Katiyar, S.K. (−)-Epigallocatechin-3-Gallate Reactivates Silenced Tumor Suppressor Genes, Cip1/P21 and P16INK4a, by Reducing DNA Methylation and Increasing Histones Acetylation in Human Skin Cancer Cells. Carcinogenesis 2011, 32, 537–544. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, N.; Adhami, V.M.; Gupta, S.; Cheng, P.; Mukhtar, H. Role of the Retinoblastoma (PRb)-E2F/DP Pathway in Cancer Chemopreventive Effects of Green Tea Polyphenol Epigallocatechin-3-Gallate. Arch. Biochem. Biophys. 2002, 398, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Erker, L.; Schubert, R.; Yakushiji, H.; Barlow, C.; Larson, D.; Mitchell, J.B.; Wynshaw-Boris, A. Cancer Chemoprevention by the Antioxidant Tempol Acts Partially via the P53 Tumor Suppressor. Hum. Mol. Genet. 2005, 14, 1699–1708. [Google Scholar] [CrossRef] [PubMed]
- Papadimitrakopoulou, V.A.; Izzo, J.; Mao, L.; Keck, J.; Hamilton, D.; Shin, D.M.; El-Naggar, A.; den Hollander, P.; Liu, D.; Hittelman, W.N.; et al. Cyclin D1 and P16 Alterations in Advanced Premalignant Lesions of the Upper Aerodigestive Tract: Role in Response to Chemoprevention and Cancer Development. Clin. Cancer Res. 2001, 7, 3127–3134. [Google Scholar] [PubMed]
- Tsukamoto, S.; Yamashita, S.; Kim, Y.H.; Kumazoe, M.; Huang, Y.; Yamada, K.; Tachibana, H. Oxygen Partial Pressure Modulates 67-KDa Laminin Receptor Expression, Leading to Altered Activity of the Green Tea Polyphenol, EGCG. FEBS Lett. 2012, 586, 3441–3447. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, Y.; Sumida, M.; Sugihara, K.; Tsukamoto, S.; Yamada, K.; Tachibana, H. Green Tea Polyphenol EGCG Sensing Motif on the 67-KDa Laminin Receptor. PLoS ONE 2012, 7, e37942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.-H.; Kwak, J.; Choi, H.-K.; Choi, K.-C.; Kim, S.; Lee, J.; Jun, W.; Park, H.-J.; Yoon, H.-G. EGCG Suppresses Prostate Cancer Cell Growth Modulating Acetylation of Androgen Receptor by Anti-Histone Acetyltransferase Activity. Int. J. Mol. Med. 2012, 30, 69–74. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Yuan, Y.-Y.; Meeran, S.M.; Tollefsbol, T.O. Synergistic Epigenetic Reactivation of Estrogen Receptor-α (ERα) by Combined Green Tea Polyphenol and Histone Deacetylase Inhibitor in ERα-Negative Breast Cancer Cells. Mol. Cancer 2010, 9, 274. [Google Scholar] [CrossRef] [Green Version]
- Sirtori, C.R.; Arnoldi, A.; Johnson, S.K. Phytoestrogens: End of a Tale? Ann. Med. 2005, 37, 423–438. [Google Scholar] [CrossRef]
- Adjakly, M.; Ngollo, M.; Boiteux, J.-P.; Bignon, Y.-J.; Guy, L.; Bernard-Gallon, D. Genistein and Daidzein: Different Molecular Effects on Prostate Cancer. Anticancer Res. 2013, 33, 39–44. [Google Scholar]
- Dixon, R.A.; Ferreira, D. Genistein. Phytochemistry 2002, 60, 205–211. [Google Scholar] [CrossRef]
- Peterson, G.; Barnes, S. Genistein Inhibition of the Growth of Human Breast Cancer Cells: Independence from Estrogen Receptors and the Multi-Drug Resistance Gene. Biochem. Biophys. Res. Commun. 1991, 179, 661–667. [Google Scholar] [CrossRef]
- Ullrich, A.; Schlessinger, J. Signal Transduction by Receptors with Tyrosine Kinase Activity. Cell 1990, 61, 203–212. [Google Scholar] [CrossRef]
- Okura, A.; Arakawa, H.; Oka, H.; Yoshinari, T.; Monden, Y. Effect of Genistein on Topoisomerase Activity and on the Growth of [Val 12]Ha-Ras-Transformed NIH 3T3 Cells. Biochem. Biophys. Res. Commun. 1988, 157, 183–189. [Google Scholar] [CrossRef]
- Evans, B.A.; Griffiths, K.; Morton, M.S. Inhibition of 5 Alpha-Reductase in Genital Skin Fibroblasts and Prostate Tissue by Dietary Lignans and Isoflavonoids. J. Endocrinol. 1995, 147, 295–302. [Google Scholar] [CrossRef]
- Huang, J.; Nasr, M.; Kim, Y.; Matthews, H.R. Genistein Inhibits Protein Histidine Kinase. J. Biol. Chem. 1992, 267, 15511–15515. [Google Scholar] [CrossRef]
- Zhou, Y.; Lee, A.S. Mechanism for the Suppression of the Mammalian Stress Response by Genistein, an Anticancer Phytoestrogen from Soy. J. Natl. Cancer Inst. 1998, 90, 381–388. [Google Scholar] [CrossRef] [Green Version]
- Brunet, A.; Bonni, A.; Zigmond, M.J.; Lin, M.Z.; Juo, P.; Hu, L.S.; Anderson, M.J.; Arden, K.C.; Blenis, J.; Greenberg, M.E. Akt Promotes Cell Survival by Phosphorylating and Inhibiting a Forkhead Transcription Factor. Cell 1999, 96, 857–868. [Google Scholar] [CrossRef] [Green Version]
- Park, W.; Amin, A.R.M.R.; Chen, Z.G.; Shin, D.M. New Perspectives of Curcumin in Cancer Prevention. Cancer Prev. Res. 2013, 6, 387–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, B.; Chung, H.Y.; Kim, N.D. Role of Apigenin in Cancer Prevention via the Induction of Apoptosis and Autophagy. J. Cancer Prev. 2016, 21, 216–226. [Google Scholar] [CrossRef] [Green Version]
- Schramm, L. Going Green: The Role of the Green Tea Component EGCG in Chemoprevention. J. Carcinog. Mutagen. 2013, 4, 1000142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavese, J.M.; Farmer, R.L.; Bergan, R.C. Inhibition of Cancer Cell Invasion and Metastasis by Genistein. Cancer Metastasis Rev. 2010, 29, 465–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annabi, B.; Lachambre, M.-P.; Bousquet-Gagnon, N.; Pagé, M.; Gingras, D.; Béliveau, R. Green Tea Polyphenol (−)-Epigallocatechin 3-Gallate Inhibits MMP-2 Secretion and MT1-MMP-Driven Migration in Glioblastoma Cells. Biochim. Biophys. Acta (BBA)–Mol. Cell Res. 2002, 1542, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Bagchi, D.; Sen, C.K.; Bagchi, M.; Atalay, M. Anti-Angiogenic, Antioxidant, and Anti-Carcinogenic Properties of a Novel Anthocyanin-Rich Berry Extract Formula. Biochemistry 2004, 69, 75–80. [Google Scholar] [CrossRef]
- Yamakawa, S.; Asai, T.; Uchida, T.; Matsukawa, M.; Akizawa, T.; Oku, N. (−)-Epigallocatechin Gallate Inhibits Membrane-Type 1 Matrix Metalloproteinase, MT1-MMP, and Tumor Angiogenesis. Cancer Lett. 2004, 210, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Labrecque, L.; Lamy, S.; Chapus, A.; Mihoubi, S.; Durocher, Y.; Cass, B.; Bojanowski, M.W.; Gingras, D.; Béliveau, R. Combined Inhibition of PDGF and VEGF Receptors by Ellagic Acid, a Dietary-Derived Phenolic Compound. Carcinogenesis 2005, 26, 821–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, R.; Kim, M.H. Epigallocatechin Gallate Inhibits HIF-1alpha Degradation in Prostate Cancer Cells. Biochem. Biophys. Res. Commun. 2005, 334, 543–548. [Google Scholar] [CrossRef]
- Kim, J.-D.; Liu, L.; Guo, W.; Meydani, M. Chemical Structure of Flavonols in Relation to Modulation of Angiogenesis and Immune-Endothelial Cell Adhesion. J. Nutr. Biochem. 2006, 17, 165–176. [Google Scholar] [CrossRef]
- Schindler, R.; Mentlein, R. Flavonoids and Vitamin E Reduce the Release of the Angiogenic Peptide Vascular Endothelial Growth Factor from Human Tumor Cells. J. Nutr. 2006, 136, 1477–1482. [Google Scholar] [CrossRef] [Green Version]
- Woodhouse, E.C.; Chuaqui, R.F.; Liotta, L.A. General Mechanisms of Metastasis. Cancer 1997, 80, 1529–1537. [Google Scholar] [CrossRef]
- Babich, H.; Pinsky, S.M.; Muskin, E.T.; Zuckerbraun, H.L. In vitro Cytotoxicity of a Theaflavin Mixture from Black Tea to Malignant, Immortalized, and Normal Cells from the Human Oral Cavity. Toxicol. In Vitro 2006, 20, 677–688. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.K.; Koponen, J.M.; Mykkänen, H.M.; Törrönen, A.R. Berry Phenolic Extracts Modulate the Expression of P21(WAF1) and Bax but Not Bcl-2 in HT-29 Colon Cancer Cells. J. Agric. Food Chem. 2007, 55, 1156–1163. [Google Scholar] [CrossRef]
- Feng, R.; Ni, H.-M.; Wang, S.Y.; Tourkova, I.L.; Shurin, M.R.; Harada, H.; Yin, X.-M. Cyanidin-3-Rutinoside, a Natural Polyphenol Antioxidant, Selectively Kills Leukemic Cells by Induction of Oxidative Stress. J. Biol. Chem. 2007, 282, 13468–13476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manna, S.; Banerjee, S.; Mukherjee, S.; Das, S.; Panda, C.K. Epigallocatechin Gallate Induced Apoptosis in Sarcoma180 Cells In vivo: Mediated by P53 Pathway and Inhibition in U1B, U4-U6 UsnRNAs Expression. Apoptosis 2006, 11, 2267–2276. [Google Scholar] [CrossRef]
- Hastak, K.; Agarwal, M.K.; Mukhtar, H.; Agarwal, M.L. Ablation of Either P21 or Bax Prevents P53-Dependent Apoptosis Induced by Green Tea Polyphenol Epigallocatechin-3-Gallate. FASEB J. 2005, 19, 789–791. [Google Scholar] [CrossRef]
- Kundu, T.; Dey, S.; Roy, M.; Siddiqi, M.; Bhattacharya, R.K. Induction of Apoptosis in Human Leukemia Cells by Black Tea and Its Polyphenol Theaflavin. Cancer Lett. 2005, 230, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Kalra, N.; Seth, K.; Prasad, S.; Singh, M.; Pant, A.B.; Shukla, Y. Theaflavins Induced Apoptosis of LNCaP Cells Is Mediated through Induction of P53, down-Regulation of NF-Kappa B and Mitogen-Activated Protein Kinases Pathways. Life Sci. 2007, 80, 2137–2146. [Google Scholar] [CrossRef]
- Favot, L.; Martin, S.; Keravis, T.; Andriantsitohaina, R.; Lugnier, C. Involvement of Cyclin-Dependent Pathway in the Inhibitory Effect of Delphinidin on Angiogenesis. Cardiovasc. Res. 2003, 59, 479–487. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.; Favot, L.; Matz, R.; Lugnier, C.; Andriantsitohaina, R. Delphinidin Inhibits Endothelial Cell Proliferation and Cell Cycle Progression through a Transient Activation of ERK-1/-2. Biochem. Pharmacol. 2003, 65, 669–675. [Google Scholar] [CrossRef]
- Pérez-Ruiz, E.; Melero, I.; Kopecka, J.; Sarmento-Ribeiro, A.B.; García-Aranda, M.; De Las Rivas, J. Cancer Immunotherapy Resistance Based on Immune Checkpoints Inhibitors: Targets, Biomarkers, and Remedies. Drug Resist. Updates 2020, 53, 100718. [Google Scholar] [CrossRef]
- Farooqi, A.A.; de la Roche, M.; Djamgoz, M.B.A.; Siddik, Z.H. Overview of the Oncogenic Signaling Pathways in Colorectal Cancer: Mechanistic Insights. Semin. Cancer Biol. 2019, 58, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Gasparri, M.L.; Bardhi, E.; Ruscito, I.; Papadia, A.; Farooqi, A.A.; Marchetti, C.; Bogani, G.; Ceccacci, I.; Mueller, M.D.; Benedetti Panici, P. PI3K/AKT/MTOR Pathway in Ovarian Cancer Treatment: Are We on the Right Track? Geburtshilfe Frauenheilkd 2017, 77, 1095–1103. [Google Scholar] [CrossRef] [Green Version]
- Patterson, S.L.; Colbert Maresso, K.; Hawk, E. Cancer Chemoprevention: Successes and Failures. Clin. Chem. 2013, 59, 94–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, R.; Kong, A.-N.T. Activation of MAP Kinases, Apoptosis and Nutrigenomics of Gene Expression Elicited by Dietary Cancer-Prevention Compounds. Nutrition 2004, 20, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Kong, A.-N.T. Dietary Cancer-Chemopreventive Compounds: From Signaling and Gene Expression to Pharmacological Effects. Trends Pharmacol. Sci. 2005, 26, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Michl, C.; Vivarelli, F.; Weigl, J.; Nicola, G.R.D.; Canistro, D.; Paolini, M.; Iori, R.; Rascle, A. The Chemopreventive Phytochemical Moringin Isolated from Moringa Oleifera Seeds Inhibits JAK/STAT Signaling. PLoS ONE 2016, 11, e0157430. [Google Scholar] [CrossRef]
- Cohen, S.M.; Arnold, L.L. Chemical Carcinogenesis. Toxicol. Sci. 2011, 120, S76–S92. [Google Scholar] [CrossRef] [PubMed]
- Burotto, M.; Chiou, V.L.; Lee, J.-M.; Kohn, E.C. The MAPK Pathway across Different Malignancies: A New Perspective. Cancer 2014, 120, 3446–3456. [Google Scholar] [CrossRef] [Green Version]
- Adachi, S.; Shimizu, M.; Shirakami, Y.; Yamauchi, J.; Natsume, H.; Matsushima-Nishiwaki, R.; To, S.; Weinstein, I.B.; Moriwaki, H.; Kozawa, O. (−)-Epigallocatechin Gallate Downregulates EGF Receptor via Phosphorylation at Ser1046/1047 by P38 MAPK in Colon Cancer Cells. Carcinogenesis 2009, 30, 1544–1552. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.-H.; Jan, R.-L.; Kuo, C.-H.; Chu, Y.-T.; Wang, W.-L.; Lee, M.-S.; Chen, H.-N.; Hung, C.-H. Natural Flavone Kaempferol Suppresses Chemokines Expression in Human Monocyte THP-1 Cells through MAPK Pathways. J. Food Sci. 2010, 75, H254–H259. [Google Scholar] [CrossRef] [PubMed]
- Karar, J.; Maity, A. PI3K/AKT/MTOR Pathway in Angiogenesis. Front. Mol. Neurosci. 2011, 4, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.M.; Jin, K.-S.; Lee, Y.-W.; Song, Y.S. Luteolin and Chicoric Acid Synergistically Inhibited Inflammatory Responses via Inactivation of PI3K-Akt Pathway and Impairment of NF-ΚB Translocation in LPS Stimulated RAW 264.7 Cells. Eur. J. Pharmacol. 2011, 660, 454–459. [Google Scholar] [CrossRef] [PubMed]
- Zanotto-Filho, A.; Braganhol, E.; Edelweiss, M.I.; Behr, G.A.; Zanin, R.; Schröder, R.; Simões-Pires, A.; Battastini, A.M.O.; Moreira, J.C.F. The Curry Spice Curcumin Selectively Inhibits Cancer Cells Growth In vitro and in Preclinical Model of Glioblastoma. J. Nutr. Biochem. 2012, 23, 591–601. [Google Scholar] [CrossRef]
- Sun, Z.-J.; Chen, G.; Hu, X.; Zhang, W.; Liu, Y.; Zhu, L.-X.; Zhou, Q.; Zhao, Y.-F. Activation of PI3K/Akt/IKK-Alpha/NF-KappaB Signaling Pathway Is Required for the Apoptosis-Evasion in Human Salivary Adenoid Cystic Carcinoma: Its Inhibition by Quercetin. Apoptosis 2010, 15, 850–863. [Google Scholar] [CrossRef]
- Prasad, R.; Vaid, M.; Katiyar, S.K. Grape Proanthocyanidin Inhibit Pancreatic Cancer Cell Growth In vitro and In vivo through Induction of Apoptosis and by Targeting the PI3K/Akt Pathway. PLoS ONE 2012, 7, e43064. [Google Scholar] [CrossRef]
- Shin, D.Y.; Kim, G.-Y.; Lee, J.H.; Choi, B.T.; Yoo, Y.H.; Choi, Y.H. Apoptosis Induction of Human Prostate Carcinoma DU145 Cells by Diallyl Disulfide via Modulation of JNK and PI3K/AKT Signaling Pathways. Int. J. Mol. Sci. 2012, 13, 14158–14171. [Google Scholar] [CrossRef]
- Pawlik, A.; Wiczk, A.; Kaczyńska, A.; Antosiewicz, J.; Herman-Antosiewicz, A. Sulforaphane Inhibits Growth of Phenotypically Different Breast Cancer Cells. Eur. J. Nutr. 2013, 52, 1949–1958. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-H.; Chang, C.-Y.; Lee, K.-R.; Lin, H.-J.; Chen, T.-H.; Wan, L. Flavones Inhibit Breast Cancer Proliferation through the Akt/FOXO3a Signaling Pathway. BMC Cancer 2015, 15, 958. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Lee, H.; Herrmann, A.; Buettner, R.; Jove, R. Revisiting STAT3 Signalling in Cancer: New and Unexpected Biological Functions. Nat. Rev. Cancer 2014, 14, 736–746. [Google Scholar] [CrossRef]
- Kim, D.-H.; Park, K.-W.; Chae, I.G.; Kundu, J.; Kim, E.-H.; Kundu, J.K.; Chun, K.-S. Carnosic Acid Inhibits STAT3 Signaling and Induces Apoptosis through Generation of ROS in Human Colon Cancer HCT116 Cells. Mol. Carcinog. 2016, 55, 1096–1110. [Google Scholar] [CrossRef]
- Wang, D.; Wise, M.L.; Li, F.; Dey, M. Phytochemicals Attenuating Aberrant Activation of β-Catenin in Cancer Cells. PLoS ONE 2012, 7, e50508. [Google Scholar] [CrossRef] [Green Version]
- Taipale, J.; Beachy, P.A. The Hedgehog and Wnt Signalling Pathways in Cancer. Nature 2001, 411, 349–354. [Google Scholar] [CrossRef]
- Driehuis, E.; Clevers, H. WNT Signalling Events near the Cell Membrane and Their Pharmacological Targeting for the Treatment of Cancer. Br. J. Pharmacol. 2017, 174, 4547–4563. [Google Scholar] [CrossRef] [Green Version]
- Tsai, J.-H.; Hsu, L.-S.; Lin, C.-L.; Hong, H.-M.; Pan, M.-H.; Way, T.-D.; Chen, W.-J. 3,5,4′-Trimethoxystilbene, a Natural Methoxylated Analog of Resveratrol, Inhibits Breast Cancer Cell Invasiveness by Downregulation of PI3K/Akt and Wnt/β-Catenin Signaling Cascades and Reversal of Epithelial-Mesenchymal Transition. Toxicol. Appl. Pharmacol. 2013, 272, 746–756. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Gwak, J.; Park, S.; Yang, C.S. Green Tea Polyphenol EGCG Suppresses Wnt/β-Catenin Signaling by Promoting GSK-3β- and PP2A-Independent β-Catenin Phosphorylation/Degradation. Biofactors 2014, 40, 586–595. [Google Scholar] [CrossRef]
- Miyashita, T.; Reed, J.C. Tumor Suppressor P53 Is a Direct Transcriptional Activator of the Human Bax Gene. Cell 1995, 80, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.-H.; Lee, J.-G.; Yang, Y.-I.; Kim, J.-H.; Ahn, J.-H.; Baek, N.-I.; Lee, K.-T.; Choi, J.-H. Eupatilin, a Dietary Flavonoid, Induces G2/M Cell Cycle Arrest in Human Endometrial Cancer Cells. Food Chem. Toxicol. 2011, 49, 1737–1744. [Google Scholar] [CrossRef]
- Wang, X.; Bai, H.; Zhang, X.; Liu, J.; Cao, P.; Liao, N.; Zhang, W.; Wang, Z.; Hai, C. Inhibitory Effect of Oleanolic Acid on Hepatocellular Carcinoma via ERK-P53-Mediated Cell Cycle Arrest and Mitochondrial-Dependent Apoptosis. Carcinogenesis 2013, 34, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Cordero-Herrera, I.; Martín, M.A.; Bravo, L.; Goya, L.; Ramos, S. Epicatechin Gallate Induces Cell Death via P53 Activation and Stimulation of P38 and JNK in Human Colon Cancer SW480 Cells. Nutr. Cancer 2013, 65, 718–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sui, Y.; Yao, H.; Li, S.; Jin, L.; Shi, P.; Li, Z.; Wang, G.; Lin, S.; Wu, Y.; Li, Y.; et al. Delicaflavone Induces Autophagic Cell Death in Lung Cancer via Akt/MTOR/P70S6K Signaling Pathway. J. Mol. Med. 2017, 95, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Wung, B.S.; Hsu, M.C.; Wu, C.C.; Hsieh, C.W. Resveratrol Suppresses IL-6-Induced ICAM-1 Gene Expression in Endothelial Cells: Effects on the Inhibition of STAT3 Phosphorylation. Life Sci. 2005, 78, 389–397. [Google Scholar] [CrossRef]
- Rahimi-Madiseh, M.; Malekpour-Tehrani, A.; Bahmani, M.; Rafieian-Kopaei, M. The Research and Development on the Antioxidants in Prevention of Diabetic Complications. Asian Pac. J. Trop. Med. 2016, 9, 825–831. [Google Scholar] [CrossRef] [Green Version]
- Akanji, M.A.; Fatinukun, H.D.; Rotimi, D.E.; Afolabi, B.L.; Adeyemi, O.S. The Two Sides of Dietary Antioxidants in Cancer Therapy; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef]
- Dizdaroglu, M.; Jaruga, P.; Birincioglu, M.; Rodriguez, H. Free Radical-Induced Damage to DNA: Mechanisms and Measurement. Free Radic. Biol. Med. 2002, 32, 1102–1115. [Google Scholar] [CrossRef]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. CPB 2017, 44, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Yeung, A.W.K.; Tzvetkov, N.T.; El-Tawil, O.S.; Bungǎu, S.G.; Abdel-Daim, M.M.; Atanasov, A.G. Antioxidants: Scientific Literature Landscape Analysis. Oxid. Med. Cell. Longev. 2019, 2019, 8278454. [Google Scholar] [CrossRef] [PubMed]
- Chikara, S.; Nagaprashantha, L.D.; Singhal, J.; Horne, D.; Awasthi, S.; Singhal, S.S. Oxidative Stress and Dietary Phytochemicals: Role in Cancer Chemoprevention and Treatment. Cancer Lett. 2018, 413, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Wilken, R.; Veena, M.S.; Wang, M.B.; Srivatsan, E.S. Curcumin: A Review of Anti-Cancer Properties and Therapeutic Activity in Head and Neck Squamous Cell Carcinoma. Mol. Cancer 2011, 10, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.-S.; Wu, J.-R.; Hu, C.-T. Signal Cross Talks for Sustained MAPK Activation and Cell Migration: The Potential Role of Reactive Oxygen Species. Cancer Metastasis Rev. 2008, 27, 303–314. [Google Scholar] [CrossRef]
- Dhawan, P.; Richmond, A. A Novel NF-Kappa B-Inducing Kinase-MAPK Signaling Pathway up-Regulates NF-Kappa B Activity in Melanoma Cells. J. Biol. Chem. 2002, 277, 7920–7928. [Google Scholar] [CrossRef] [Green Version]
- Das, L.; Vinayak, M. Long Term Effect of Curcumin in Regulation of Glycolytic Pathway and Angiogenesis via Modulation of Stress Activated Genes in Prevention of Cancer. PLoS ONE 2014, 9, e99583. [Google Scholar] [CrossRef] [Green Version]
- Garattini, E.; Bolis, M.; Garattini, S.K.; Fratelli, M.; Centritto, F.; Paroni, G.; Gianni’, M.; Zanetti, A.; Pagani, A.; Fisher, J.N.; et al. Retinoids and Breast Cancer: From Basic Studies to the Clinic and Back Again. Cancer Treat. Rev. 2014, 40, 739–749. [Google Scholar] [CrossRef]
- Doldo, E.; Costanza, G.; Agostinelli, S.; Tarquini, C.; Ferlosio, A.; Arcuri, G.; Passeri, D.; Scioli, M.G.; Orlandi, A. Vitamin A, Cancer Treatment and Prevention: The New Role of Cellular Retinol Binding Proteins. BioMed Res. Int. 2015, 2015, 624627. [Google Scholar] [CrossRef] [Green Version]
- Werner, B.; Gallagher, R.E.; Paietta, E.M.; Litzow, M.R.; Tallman, M.S.; Wiernik, P.H.; Slack, J.L.; Willman, C.L.; Sun, Z.; Traulsen, A.; et al. Dynamics of Leukemia Stem-like Cell Extinction in Acute Promyelocytic Leukemia. Cancer Res. 2014, 74, 5386–5396. [Google Scholar] [CrossRef] [Green Version]
- van Breemen, R.B.; Pajkovic, N. Multitargeted Therapy of Cancer by Lycopene. Cancer Lett. 2008, 269, 339–351. [Google Scholar] [CrossRef] [Green Version]
- Montagnani Marelli, M.; Marzagalli, M.; Fontana, F.; Raimondi, M.; Moretti, R.M.; Limonta, P. Anticancer Properties of Tocotrienols: A Review of Cellular Mechanisms and Molecular Targets. J. Cell. Physiol. 2019, 234, 1147–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constantinou, C.; Charalambous, C.; Kanakis, D. Vitamin E and Cancer: An Update on the Emerging Role of γ and δ Tocotrienols. Eur. J. Nutr. 2020, 59, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Stevens-Barrón, J.; Astiazarán-Garcia, H.; Robles-Zepeda, R.E. Efectividad y principales mecanismos anticancerígenos de tocotrienoles en líneas celulares malignas. Rev. Mex. Cienc. Farm. 2017, 13, 16–27. [Google Scholar]
- Shin-Kang, S.; Ramsauer, V.P.; Lightner, J.; Chakraborty, K.; Stone, W.; Campbell, S.; Reddy, S.A.G.; Krishnan, K. Tocotrienols Inhibit AKT and ERK Activation and Suppress Pancreatic Cancer Cell Proliferation by Suppressing the ErbB2 Pathway. Free Radic. Biol. Med. 2011, 51, 1164–1174. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnekenburger, M.; Dicato, M.; Diederich, M. Plant-Derived Epigenetic Modulators for Cancer Treatment and Prevention. Biotechnol. Adv. 2014, 32, 1123–1132. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.N.; Almoyad, M.; Huq, F. Polyphenols in Colorectal Cancer: Current State of Knowledge Including Clinical Trials and Molecular Mechanism of Action. BioMed Res. Int. 2018, 2018, 4154185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.-L.; Liu, Y.-Y.; Ma, Y.-G.; Xue, Y.-X.; Liu, D.-G.; Ren, Y.; Liu, X.-B.; Li, Y.; Li, Z. Curcumin Blocks Small Cell Lung Cancer Cells Migration, Invasion, Angiogenesis, Cell Cycle and Neoplasia through Janus Kinase-STAT3 Signalling Pathway. PLoS ONE 2012, 7, e37960. [Google Scholar] [CrossRef] [PubMed]
- Bimonte, S.; Barbieri, A.; Palma, G.; Luciano, A.; Rea, D.; Arra, C. Curcumin Inhibits Tumor Growth and Angiogenesis in an Orthotopic Mouse Model of Human Pancreatic Cancer. BioMed Res. Int. 2013, 2013, 810423. [Google Scholar] [CrossRef]
- Murakami, C.; Hirakawa, Y.; Inui, H.; Nakano, Y.; Yoshida, H. Effect of Tea Catechins on Cellular Lipid Peroxidation and Cytotoxicity in HepG2 Cells. Biosci. Biotechnol. Biochem. 2002, 66, 1559–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.-H.; Li, Y.-Q.; Yang, X.-Y. Inhibition of Epigallocatechin Gallate on Orthotopic Colon Cancer by Upregulating the Nrf2-UGT1A Signal Pathway in Nude Mice. PHA 2007, 80, 269–278. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Mohammed, H.M.; Khadrawy, S.M.; Galaly, S.R. Hesperidin Protects against Chemically Induced Hepatocarcinogenesis via Modulation of Nrf2/ARE/HO-1, PPARγ and TGF-Β1/Smad3 Signaling, and Amelioration of Oxidative Stress and Inflammation. Chem. Biol. Interact. 2017, 277, 146–158. [Google Scholar] [CrossRef]
- Granado-Serrano, A.B.; Martín, M.A.; Bravo, L.; Goya, L.; Ramos, S. Quercetin Modulates Nrf2 and Glutathione-Related Defenses in HepG2 Cells: Involvement of P38. Chem. Biol. Interact. 2012, 195, 154–164. [Google Scholar] [CrossRef] [Green Version]
- Ryu, M.J.; Chung, H.S. [10]-Gingerol Induces Mitochondrial Apoptosis through Activation of MAPK Pathway in HCT116 Human Colon Cancer Cells. In Vitro Cell. Dev. Biol. Anim. 2015, 51, 92–101. [Google Scholar] [CrossRef]
- Lewinska, A.; Adamczyk-Grochala, J.; Kwasniewicz, E.; Deregowska, A.; Wnuk, M. Ursolic Acid-Mediated Changes in Glycolytic Pathway Promote Cytotoxic Autophagy and Apoptosis in Phenotypically Different Breast Cancer Cells. Apoptosis 2017, 22, 800–815. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Sridhar, J.; Foroozesh, M. Cytochrome P450 family 1 inhibitors and structure-activity relationships. Molecules 2013, 18, 14470–14495. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Rao, J.; Li, W.; Wang, K.; Qiu, F. Mechanism-based inactivation of cytochrome P450 enzymes by natural products based on metabolic activation. Drug Metab. Rev. 2020, 52, 501–530. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Gao, Q.; Zhang, T.; Rao, J.; Ding, L.; Qiu, F. Inhibition of CYP2C9 by natural products: Insight into the potential risk of herb-drug interactions. Drug Metab. Rev. 2020, 52, 235–257. [Google Scholar] [CrossRef]
- Ciolino, H.P.; Daschner, P.J.; Yeh, G.C. Resveratrol inhibits transcription of CYP1A1 in vitro by preventing activation of the aryl hydrocarbon receptor. Cancer Res. 1998, 58, 5707–5712. [Google Scholar] [PubMed]
- Casper, R.F.; Quesne, M.; Rogers, I.M.; Shirota, T.; Jolivet, A.; Milgrom, E.; Savouret, J.F. Resveratrol has antagonist activity on the aryl hydrocarbon receptor: Implications for prevention of dioxin toxicity. Mol. Pharmacol. 1999, 56, 784–790. [Google Scholar] [PubMed]
- Revel, A.; Raanani, H.; Younglai, E.; Xu, J.; Rogers, I.; Han, R.; Savouret, J.F.; Casper, R.F. Resveratrol, a natural aryl hydrocarbon receptor antagonist, protects lung from DNA damage and apoptosis caused by benzo[a]pyrene. J. Appl. Toxicol. 2003, 23, 255–261. [Google Scholar] [CrossRef]
Figure 1.
Stages of carcinogenesis and chemoprevention.

Figure 2.
Common examples of chemopreventive dietary phytochemicals.

Figure 3.
Interaction of phytochemicals with signaling molecules in cancer chemoprevention. Figure represents action of phytochemicals on MAPK, Akt, Wnt, and JAK/STAT pathways inducing cancer cell death through activation of several internal signaling molecules.
Figure 3.
Interaction of phytochemicals with signaling molecules in cancer chemoprevention. Figure represents action of phytochemicals on MAPK, Akt, Wnt, and JAK/STAT pathways inducing cancer cell death through activation of several internal signaling molecules.


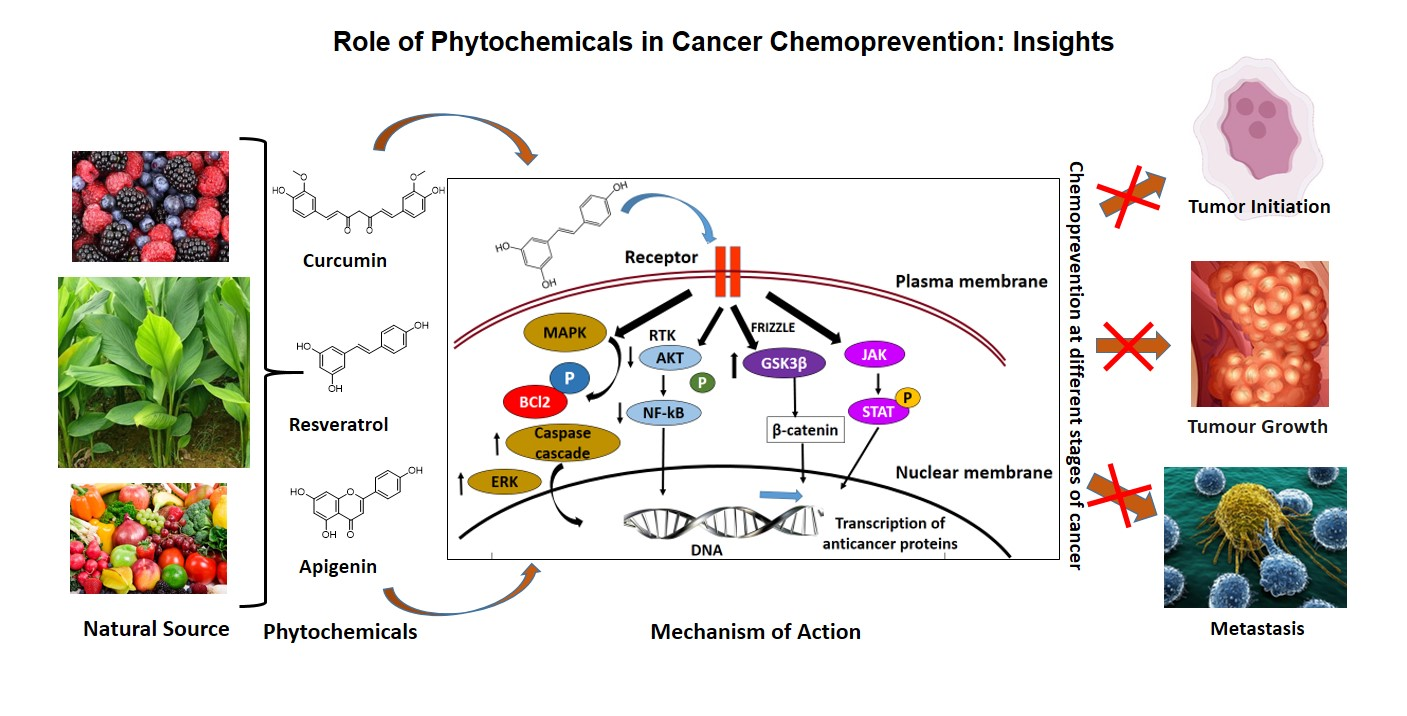{kind=link}
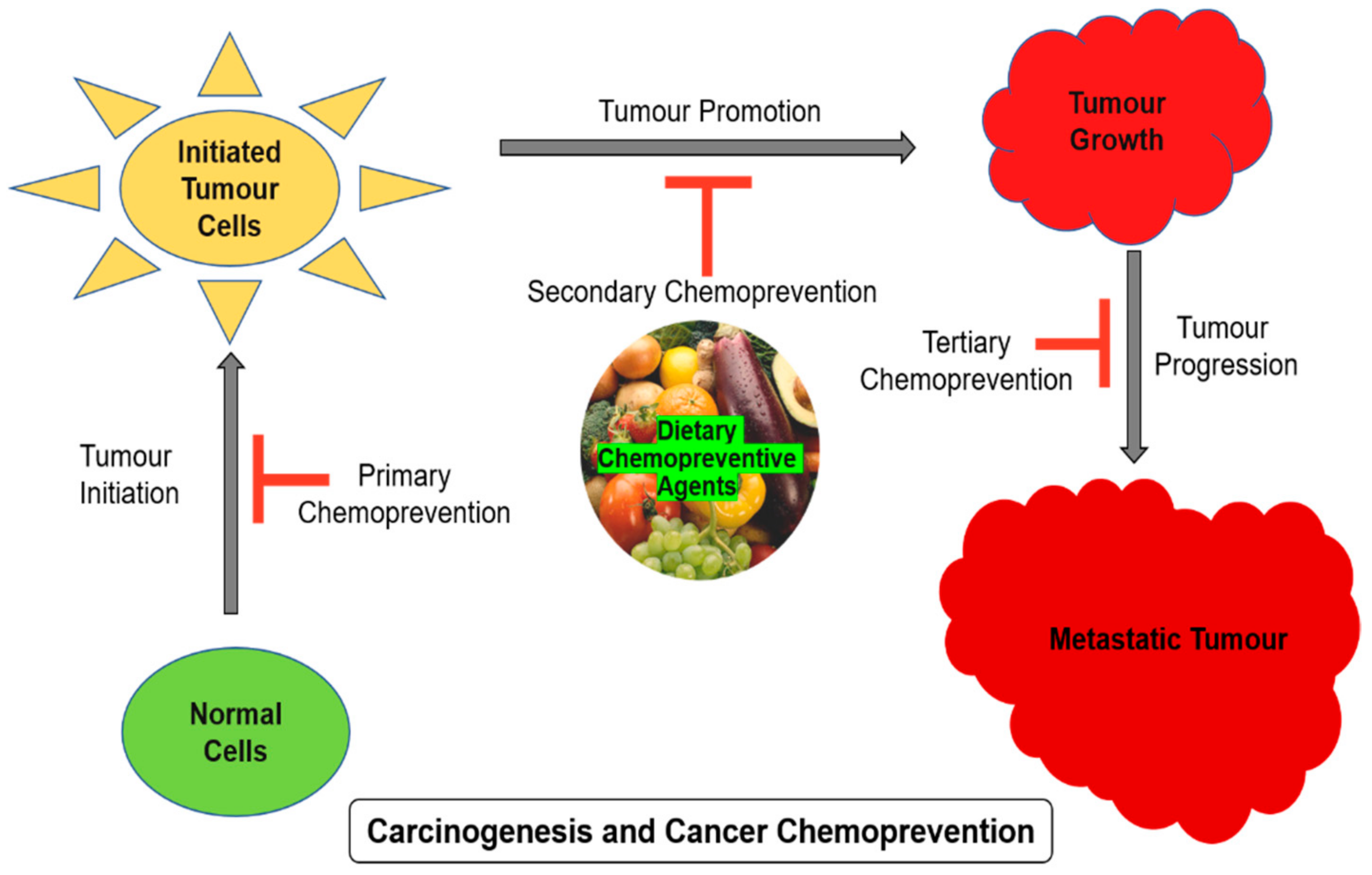{kind=link}
{kind=link}
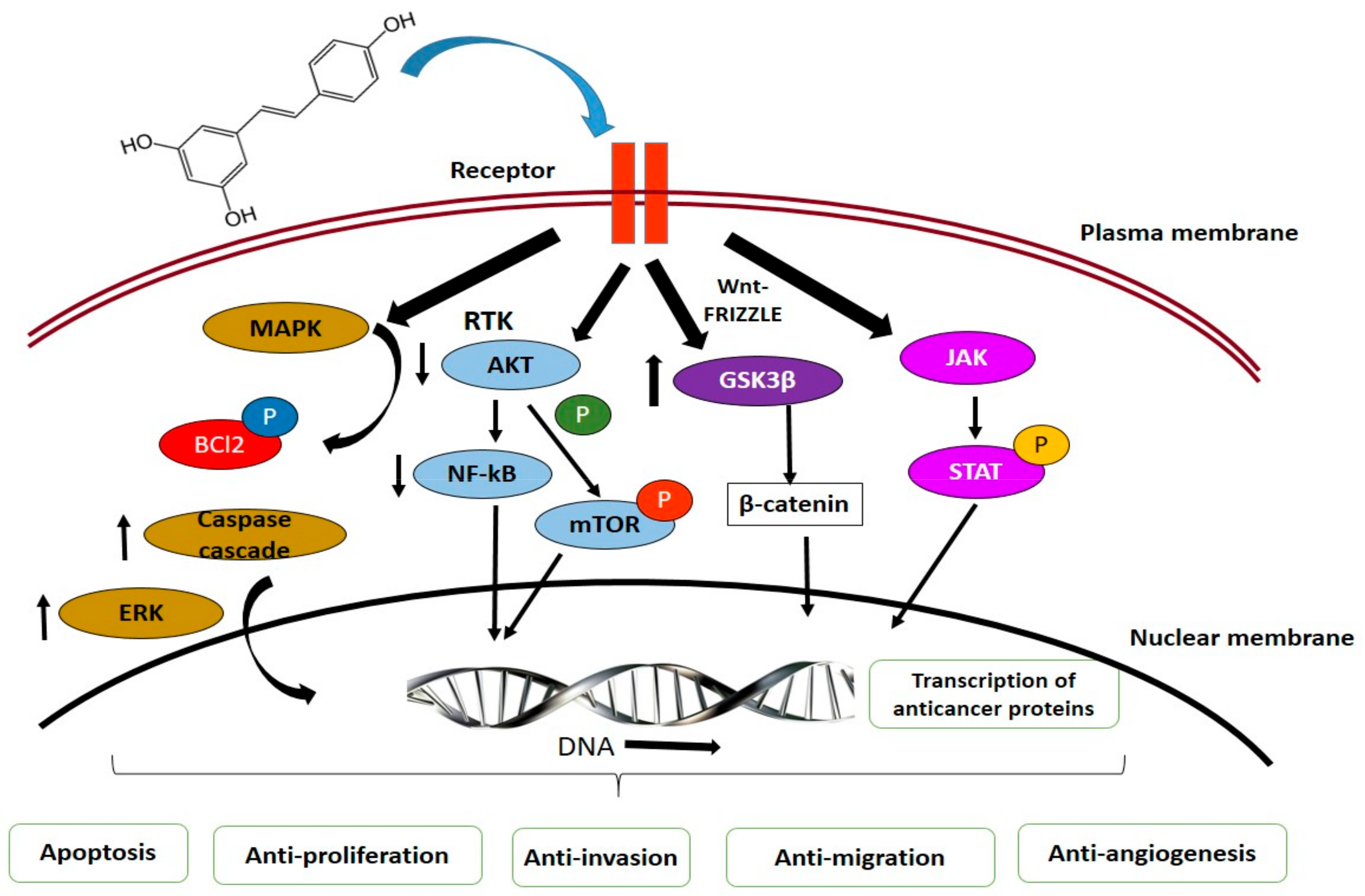{kind=link}
Table 1.
Medicinal plants used in chemoprevention of cancer and their therapeutic role.
| Common Name | Botanical Name | Therapeutic Role | Type of Cancer | References |
|---|---|---|---|---|
| Indian Gooseberry | Phyllanthus emblica | Immunomodulatory, cytoprotective | Breast cancer | [33] |
| Garlic | Allium sativum | Anticancer, immuno-stimulant | Oesophageal, colorectal cancer | [34,35] |
| Turmeric | Curcuma longa | Anti-inflammatory, anticancer; chemo-resistance | Breast cancer | [36,37,38] |
| Common mayapple | Podophyllum peltatum | Anticancer | Testicular, lung cancer | [39] |
| Heartleaf moonseed plant | Tinospora cordifolia | Immunomodulatory, antioxidant, anticancer | Cervical cancer | [40] |
| King of bitters | Andrographis paniculata | Immune stimulator | Leukaemia, Colon, breast cancer | [41] |
| Atemoya | Annona atemoya | Anticancer | Lung, Colon, breast cancer | [42] |
| Stone breaker | Phyllanthus amarus | Cell cycle arrest, DNA repair, anti-angiogenic | Lung | [43] |
| Amruta | Mappia foetida | Antineoplastic | Leukaemia, lymphoma, cervical | [44] |
| Winter cherry | Withania somnifera | Anti-inflammatory, antitumor, antioxidant, immuno-modulatory | Leukaemia | [45] |
| Himalayan cedar | Cedrus deodara | Apoptosis induction | Leukaemia | [46] |
| Heart leaved moonseed | Tinospora cordifolia | Cytotoxic | Cervical | [47] |
| Soursop | Annona muricata | Cytotoxic | Breast | [48] |
| Chestnut rose | Rosa roxburghii | Immunomodulatory, antiaging | Oesophageal, gastric, pulmonary | [49] |
| Jewel Vine | Derris scandens | Radiosensitizer | Colon | [50] |
| Penawar Hitam | Goniothalamus macrophyllus | Apoptosis induction | Cervical | [51] |
| Dong quai | Angelica sinensis | Cytotoxic | Leukaemia | [52] |
| Cang Zhu | Atractylis lancea | Apoptotic, cell cycle arrest | Liver | [53] |
| Mongolian milkvetch | Astragalus membranaceus | Immunomodulatory | Myeloid Leukaemia | [54] |
| Tea plant | Camellia sinensis | Antioxidant, antitumor, antibacterial, antimutagenic | Breast, lung, colon, skin | [55] |
| Fire-flame bush | Woodfordia fruticosa | Cytotoxic | Lung, colon, Liver, Neuroblastoma | [56] |
| Red spiderling | Boerhaavia diffusa | Cytotoxic, anticarcinogenic | Cervical | [57] |
| Umbrella cheese tree | Glochidion zeylanicum | Cytotoxic | Prostate, liver, colon | [56] |
| Black nightshade | Solanum nigrum | Antimicrobial, antioxidant, cytotoxic, antiulcerogenic, hepatoprotective | Cervical | [58] |
| Chaga | Inonotus obliquus | Anticancer | Lung, breast, cervical, stomach | [59] |
Table 2.
Chemopreventive agents and mechanisms.
| Chemopreventive Agents | Chemical Structure | Chemopreventive Mechanism | References |
|---|---|---|---|
| Curcumin |  | Inhibits lipid peroxidation, free radical generation, lipo- and cyclooxygenase, protein kinase C | [132] |
| Resveratrol |  | Alter signalling mechanism of cell division and proliferation, cell death, inflammation, angiogenesis, metastasis | [70] |
| Apigenin |  | Inhibit angiogenesis, malignant transformation, cell invasion, metastasis, induce apoptosis, regulate cell cycle | [133] |
| Epigallocatechin gallate |  | Regulate signalling pathways- PI3K/AKT, JAK/STAT, Notch, Wnt, AP-1, NFκB. tumor suppressor activities- p53; p21, p16 and Rb | [134] |
| Genistein |  | Inhibit cell growth, protein-tyrosine kinase, NF-κB, Akt signalling, PTK, regulate cell cycle regulation, induce apoptosis | [135] |
Table 3.
Gene regulatory mechanisms of phytochemicals.
| Phytochemical | Action | Gene Regulation | References |
|---|---|---|---|
| Curcumin | Antiproliferation, antimigration, anti-invasion, cytotoxicity Reduces tumor invasiveness | ↓ ERK1/2 and NFkB STAT | [206,207] |
| Epigallocatechin-3-gallate | Antiproliferation, antioxidant defence capacity Reduces exogenous oxidative stress | ↓ NFkB ↑ GSH ↑ Nrf2, UGT1A, UGT1A8, and UGT1A10 | [208,209] |
| Resveratrol | Inhibits ROS-induced proliferation and Migration Protects against (4-OHE2)- induced migration and transformation | ↓ pERK, pAKT, and pNFkB ERK, NFkB and p38, MAPK/NFkB signaling | [210] |
| Hesperedin | Reducing oxidative stress | ↓ NFkB and COX-2 | [167] |
| Quercetin | Accelerates endogenous antioxidant enzymes, Apoptosis | GST and GPx | [211,212] |
| Gingerol | Apoptosis | ↑ MAPK | [213] |
| Ursolic acid | Antiproliferation, proapoptosis, proautophagy | ↓ Akt, ↓ MAPK | [171] |
| Apigenin, flavone, eupatilin | Antiproliferation, proapoptosis | ↓ Akt | [180] |
‘↓’ indicate down-regulation and ‘↑’ indicate up-regulation of genes.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
George, B.P.; Chandran, R.; Abrahamse, H. Role of Phytochemicals in Cancer Chemoprevention: Insights. Antioxidants 2021, 10, 1455. https://doi.org/10.3390/antiox10091455
AMA Style
George BP, Chandran R, Abrahamse H. Role of Phytochemicals in Cancer Chemoprevention: Insights. Antioxidants. 2021; 10(9):1455. https://doi.org/10.3390/antiox10091455
Chicago/Turabian StyleGeorge, Blassan P., Rahul Chandran, and Heidi Abrahamse. 2021. "Role of Phytochemicals in Cancer Chemoprevention: Insights" Antioxidants 10, no. 9: 1455. https://doi.org/10.3390/antiox10091455
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.