
Beneficial Soil Microbes Negatively Affect Spider Mites and Aphids in Pepper


Laboratory of Agricultural Entomology & Zoology, Department of Agricultural Development, Democritus University of Thrace, Pantazidou 193, 68200 Orestiada, Greece
*
Author to whom correspondence should be addressed.
Agronomy 2021, 11(9), 1831; https://doi.org/10.3390/agronomy11091831
Submission received: 16 August 2021
/
Revised: 7 September 2021
/
Accepted: 9 September 2021
/
Published: 13 September 2021
(This article belongs to the Special Issue Beneficial Soil Microorganisms for Improved Plant Performance and Resilience in Sustainable Agricultural Systems)
Abstract
:Beneficial soil microbes have long been recognized for their ability to improve plant growth, to antagonize pathogens and to prime plants against biotic stressors. Nevertheless, their ability to enhance plant resistance against arthropod pests remains largely unexplored, especially in crop plants such as pepper. Herein, we assessed the effects of several fungal and bacterial species/strains applied in the soil on the performance of key pests of pepper plants. Specifically, we recorded the impact of pepper inoculation with commercial strains of beneficial bacteria (Bacillus amyloliquefaciens and Pseudomonas spp.) as well as fungi (Trichoderma spp. and Cordyceps fumosorosea) on the population growth of the green peach aphid, Myzus persicae, and the two-spotted spider mite, Tetranychus urticae. Furthermore, we recorded the effects of microbial inoculation on plant growth parameters, such as stem and root weight. We found that both pests can be negatively affected by microbial inoculation: spider mites laid up to 40% fewer eggs, and the number of aphids were up to 50% less on pepper-inoculated plants, depending on the microbe. We also recorded a variation among the tested microbes in their impact on herbivore performance, but no significant effects were found on plant biomass. Our results add to the growing literature that beneficial soil microbes may be capable of exerting biocontrol capabilities against aboveground herbivorous pests possibly, among other means, via the elicitation of plant defense responses.
Keywords:
aphids; biological control; endophytes; Myzus persicae; pepper; Tetranychus urticae; soil drench1. Introduction
Herbivorous arthropods, such as insects and mites, are key pests in several crops, causing considerable yield losses worldwide [1,2]. Chemical control is included as one of the most effective tools against pests, although it may contribute to considerable environmental problems. Consistent with the general trend of replacing pesticides with effective environmentally friendly alternatives [3], biological control methods are promising in collectively synergizing efficient pest control. Biological control, i.e., the use of living organisms to control harmful ones, includes the natural enemies of pests (predators and parasitoids) and beneficial microorganisms. In the latter group, several biocontrol agents are promising as they can affect herbivores directly as entomopathogens or indirectly via altering plant quality, producing metabolites, changing herbivores and natural enemies’ behavior or inducing essential plant defenses [3,4,5,6,7,8,9,10].
Currently, a number of beneficial microbes, such as fungi and bacteria, are commercially available as biofungicides against pathogenic microbes that cause plant diseases. The main genera in these fungi include different strains of Trichoderma spp., while Bacillus spp. and Pseudomonas spp. strains are included in bacteria. With regard to beneficial microbes that are applied against insect pests, these are rather limited to entomopathogenic bacteria and fungi, such as Bacillus thuringiensis, Beauveria bassiana and Metarhizium spp. Mechanisms involved in the fungicidal activity of soil microbes include the production of toxic metabolites, direct competition and plant defense induction [3,11,12].
Nevertheless, in nature, plants are associated with a high diversity of beneficial microbes. Selected root colonizing microbes, in particular, are also known for their beneficial impact on plant growth and nutrition, with some of them being biostimulants or pathogen competitors [13,14,15]. Improved plant growth and quality by plant growth-promoting fungi and rhizobacteria have been shown to positively affect herbivore performance, whereas Induced Systemic Resistance (ISR) by soil microbes can negatively impact herbivores [5,15,16]. For example, selected soil microbes alter the composition and emission rate of volatile blends emitted by plants, hence their attractiveness to natural enemies of herbivores [6,14]. Nevertheless, to date, we only have limited data on the effects of beneficial soil microbes on the herbivorous pests of economically important crops, and even less is known about the regulatory mechanisms underlying these interactions [4].
The ability of beneficial soil microbes to enhance plant resistance to herbivorous arthropods is an important attribute that has gained the attention of the scientific community [3,4,17]. Thus far, there is evidence that certain soil microbes can affect aboveground herbivores via the plant and several involved mechanisms, such as the elicitation of plant defenses, among others [4,5,6]. However, most studies focus on model plants (Arabidopsis) or specific crop plants (tomato) [4]. Other crops have been largely neglected, although studying the plant-mediated effects of beneficial microbes and the ways that they can shape the above- and belowground interactions of plants with their herbivorous enemies is especially needed for plants of economic importance [4]. In the context of Integrated Pest Management (IPM), these studies are crucial, considering, for example, that beneficial soil microbes could act as not only biofungicides but also biostimulants against herbivorous pests.
In our attempt to assess the effects of beneficial soil microbes on herbivorous pests and to understand the mechanisms involved, we performed a screening of several commercial microbial strains against key pests of pepper. Pepper is an important vegetable crop, with an annual production of 22.5 million tonnes in Europe [18]. It is attacked by several arthropod pests, among which are the two-spotted spider mite Tetranychus urticae Koch, a polyphagous herbivore attacking more than 1000 plant species [19], and the green peach aphid Myzus persicae Sulzer, a cosmopolitan polyphagous aphid species responsible for economic losses due to its phytophagy and ability to vector plant viruses [20].
Hence, in this study, we present our results after assessing five bacterial and four fungal strains for their plant-mediated effects against T. urticae and the green peach aphid M. persicae. We applied the microbes in the soil at a standard dose that falls within the range of field-recommended doses and then recorded performance indices, such as the number of aphids or spider mites after two weeks. We hypothesized that beneficial microbes would negatively affect the two herbivores and that variation would be recorded among the different strains.
2. Materials and Methods
2.1. Plants
Pepper (Capsicum annuum L., cv. P13, a commonly used cultivar in Greece) plants were used in all of the experiments as well as in herbivore rearing. Plants were grown from seeds that were surface-sterilized with a bleach solution (30% commercial bleach) for 10 min, washed with water and left to dry before sowing. The seeds were sown in sterilized peat (Klasmann-TS2) that was autoclaved for 1 h at 115 °C. Young seedlings were transplanted in plastic pots (~0.35 L) filled with sterilized peat. All of the plants were maintained in climate chambers (25 ± 2 °C, 16:8 LD, 60–70% RH) and watered every other day. Once a week, they were fertilized with an N–P–K fertilizer (20-20-20). The plants used in the experiments were 4–5 weeks old.
2.2. Herbivores
Spider mites (Tetranychus urticae) sampled from infested pepper plants were used to establish a lab colony. The mites were reared on intact pepper plants at 25 ± 2 °C, 16:8 LD, 60–70% RH. Young female mites (2–4 days old) were used in the experiments. These were obtained by infesting pepper plants with a high number (approx. 300) of female mites that were allowed to lay eggs for 48 h at 25 ± 2 °C, 16:8 LD. The next day, the mites were removed and the plants were maintained at the same conditions until adult mites emerged (after approximately 16 days).
Green peach aphids (Myzus persicae) were sampled from pepper plants grown in the greenhouse and were used to establish a lab colony. The aphid colony was maintained on intact pepper plants that were placed in insect cages at 25 ± 2 °C, 16:8 LD, 60–70% RH. To obtain the young (two-day-old) experimental females, plants were infested with approximately 200 aphids per plant for 48 h. Adult aphids were removed, and approximately 10 days after, the young females were used in the experiments.
2.3. Plant Inoculation with Microbes
We assessed the effects of nine microbes, five bacterial and four fungal commercial strains against spider mites and aphids. Since most of the tested microbes were used for foliar spray application (Table 1), for each microbe, the label-recommended dose expressed in kg/ha was used to estimate the equivalent application rate as if each microbe was directly applied on the soil surface. All of the products were dissolved in water and applied as water drench in sterilized peat in pots where young (approximately 10 days from seed sowing) plants had been transplanted. After one week, a second drench was performed, and two weeks after, the plants were infested with spider mites or aphids. During the experiments, the plants were maintained at 25 ± 1 °C, 16:8 LD, 60–70% RH and watered every other day. The two groups of experiments were performed separately due to space restrictions. From the microbes tested, all of the bacteria and fungi were currently commercially available as biofungicides except Cordyceps fumosorosea Apopka 97, which is an entomopathogenic fungus used to control whiteflies (Table 1).
2.4. Herbivore Performance
The herbivore performance on pepper plants that were treated with the microbes was assessed by infesting them with 45 female spider mites per plant on three leaves (15 females per leaf) or 10 aphids per plant for 14 days. Subsequently, the numbers of eggs, and live spider mites or aphids per plant were recorded. The two experiments were performed in parallel but in different chambers, both including a separate control. Per experiment (different herbivores), we used six plants per treatment (5–6 treatments per experiment).
2.5. Plant Growth Parameters
To assess the impact of microbe inoculation on the growth of peppers, another set of experimental plants was prepared as above and infested with the standard number of spider mites or aphids as in the performance experiments above. On day 14 of herbivore feeding (i.e., when the plants were 5–6 weeks old), all of the plants were harvested and weighed on a microbalance. Before weighing, the roots were gently rinsed in water and dried on tissue paper. The two experiments were performed in parallel but in different chambers, both including a separate control. Per experiment (different herbivore), we used six plants per treatment (5–6 treatments per experiment).
2.6. Statistical Analysis
To evaluate the effect of microbial inoculation on the number of spider mite eggs, spider mite individuals, number of aphids per plant and plant biomass, t-tests were used for all pairwise comparisons among the different treatments and the respective controls. Prior to statistical analysis, the normality and homogeneity of the variances were checked with Shapiro–Wilk’s and Levene’s tests, respectively. All of the statistics were performed using SPSS [21].
3. Results
3.1. Spider Mite Performance
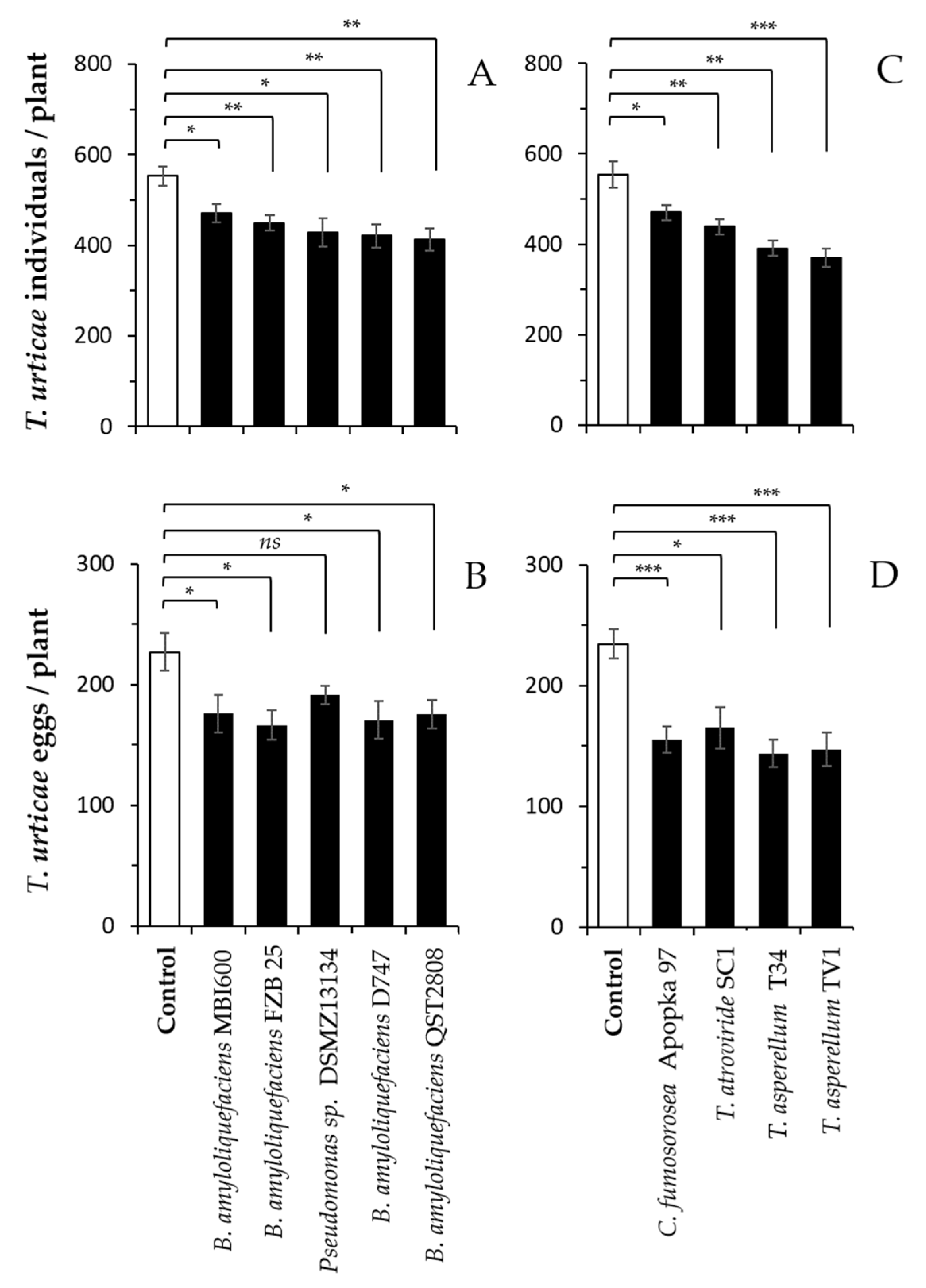
All of the tested microbes were shown to significantly affect spider mite performance. The number of individuals and eggs recorded on the leaves of inoculated plants 14 days following infestation was significantly less than those on the control (un-colonized) plants (Figure 1). The experiments with fungi and bacteria were not performed in parallel; hence, the results are presented separately per microbe group.
All of the bacterial strains significantly reduced the number of spider mites (Figure 1A: 2.75 ≤ t ≤ 4.33, df = 10, 0.0015 ≤ p ≤ 0.02) as well as the number of spider mite eggs per plant (Figure 1B: 2.06 ≤ t ≤ 3.11, df = 10, 0.01 < p ≤ 0.04) except for Pseudomonas sp. DSMZ 13134, which resulted in a similar number of eggs as the control (t = 2.065, df = 10, p = 0.066). The inoculation of the plants with B. amyloliquefaciens QST2808 resulted in the lowest number of spider mites, and that with B. amyloliquefaciens FZB25, B. amyloliquefaciens D747 and B. amyloliquefaciens QST 2808 resulted in the lowest number of spider mite eggs compared with the control.
Similarly, all of the fungal strains significantly reduced the number of spider mites (Figure 1C: 2.53 ≤ t ≤ 5.29, df = 10, 0.0003 ≤ p ≤ 0.029) as well as the number of spider mite eggs per plant (Figure 1D: 3.24 ≤ t ≤ 5.44, df = 10, 0.0003 ≤ p ≤ 0.008), with T. asperellum T34 and T. asperellum TV1 resulting in the lowest number of spider mites and eggs. Although a direct comparison among the two microbe groups (fungi vs. bacteria) or the microbes within each group cannot be made, T. asperellum T34 and T. asperellum TV1 from the fungal group are shown to result in the lowest number of spider mite eggs per plant (approximately 145 eggs/plant) compared with all of the treatments (Figure 1).
3.2. Aphid Performance
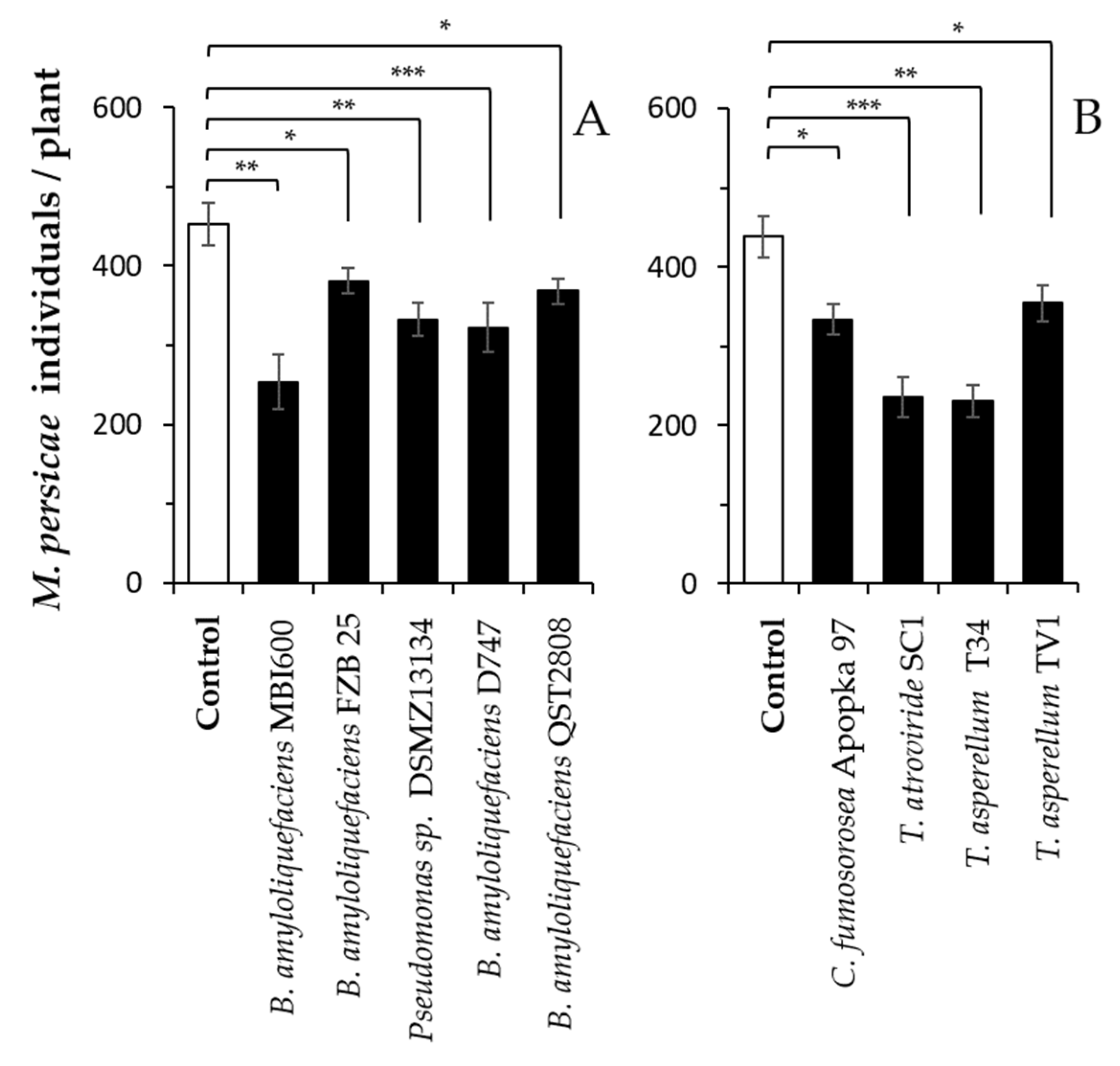
Aphid performance was significantly affected by plant inoculation with the tested microbes. The number of aphids per plant 14 days following infestation was significantly less than those on the control (un-colonized) plants (Figure 2). As with the spider mites, the experiments with aphids were also not performed in parallel for fungi and bacteria
All of the bacteria strains significantly reduced the number of aphids (Figure 2A: 2.31 ≤ t ≤ 4.59, df = 10, 0.001 ≤ p ≤ 0.041), with B. amyloliquefaciens MBI600 resulting in the lowest number (approx. 253 aphids/plant) compared with the control.
Similarly, all of the fungal strains significantly reduced the number of aphids (Figure 2B: 2.42 ≤ t ≤ 6.24, df = 10, 0.0001 ≤ p ≤ 0.035), with T. asperellum T34 and T. asperellum SC1, resulting in the lowest numbers compared with the control. From the two microbe groups (fungi vs. bacteria), B. amyloliquefaciens MBI600, T. asperellum T34 and T. asperellum SC1 resulted in a similar number of aphids per plant (approx. 230 aphids/plant) (Figure 2).
3.3. Plant Growth
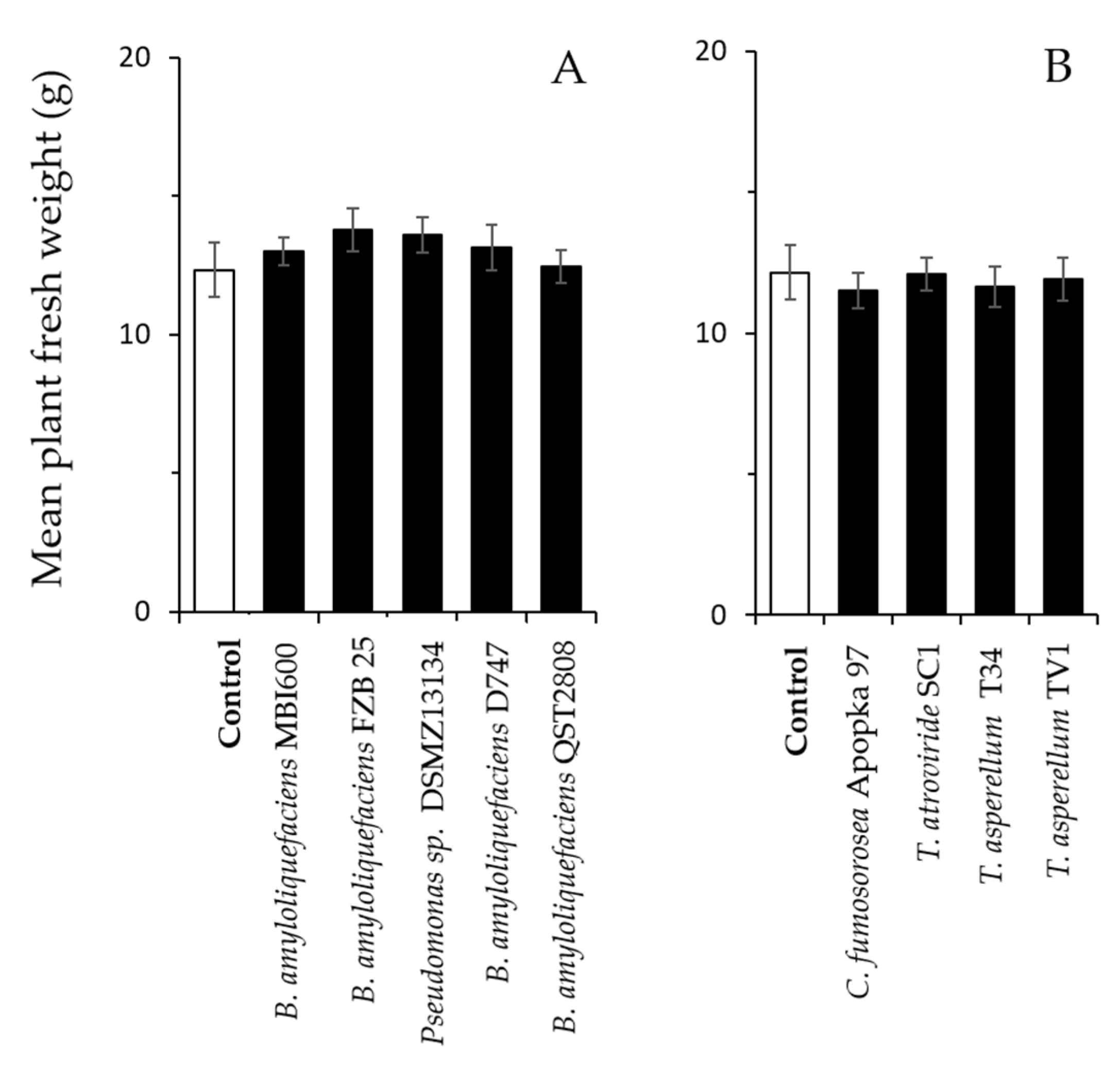
We tested whether pepper inoculation with the microbes affect the plant growth parameters of spider mite- or aphid-infested plants. We found that plant inoculation with microbes had no significant effect on the fresh weight of inoculated plants that were infested with 45 spider mite females for 14 days compared with the control (non-inoculated) plants (Figure 3A; bacteria: −1.16 ≤ t ≤ −1.04, df = 10, 0.273 ≤ p ≤ 0.919; Figure 3B; fungi: 0.05 ≤ t ≤ 0.55, df = 10, 0.594 ≤ p ≤ 0.962).
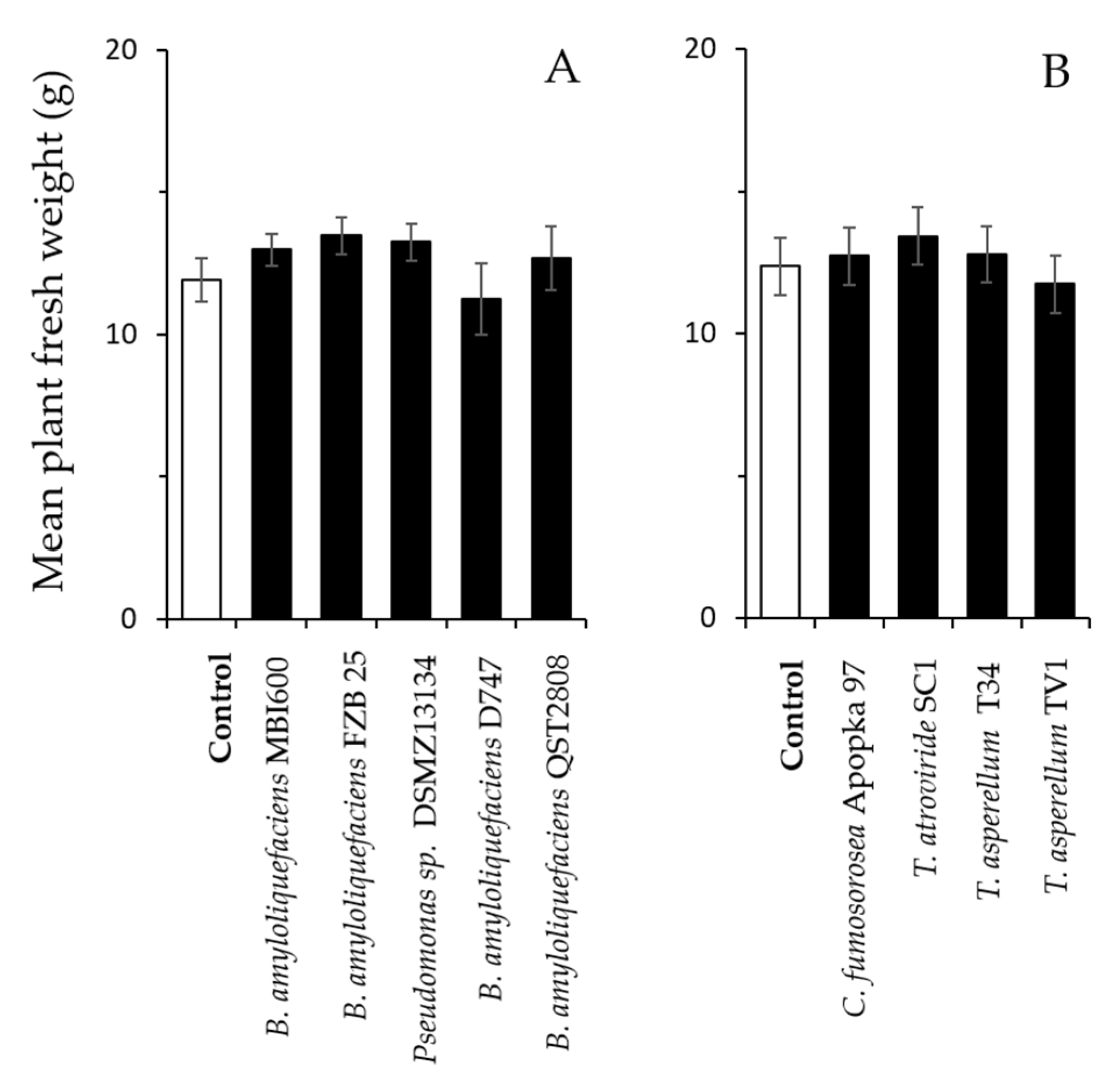
Similarly, no significant effect was recorded in the fresh weight of plants that were inoculated with the tested microbes and infested with 10 aphids for 14 days compared with the control plants (Figure 4A; bacteria: − 1.56 ≤ t ≤ 0.46, df = 10, 0.148 ≤ p ≤ 0.651; Figure 4B; fungi: − 0.71 ≤ t ≤ 0.489, df = 10, 0.496 ≤ p ≤ 0.837).
4. Discussion
In the present study, we recorded the effects of different beneficial soil microbes on the performance of spider mites and aphids in pepper. We found that both herbivores are negatively affected, with spider mites laying a lower number of eggs on microbe-treated plants except for the plants treated with Pseudomonas sp. DSMZ 13134. We also observed a lower number of spider mites as well as aphid individuals on microbe-inoculated compared with non-inoculated plants. Finally, plant biomass was not affected in the presence of the microbes in herbivore-infested plants compared with the control (non-inoculated) plants. We argue that there are indications for the existence of plant defense induction capabilities in the tested microbes, with some variation recorded within each microbe group (fungi and bacteria).
The plant-mediated effects of beneficial soil microbes other than entomopathogenic bacteria and fungi have been scarcely addressed so far. The current literature on the topic mainly includes studies using entomopathogens, such as Beauveria spp. and Metarhizium spp. strains, as a soil drench or treating seeds or roots to assess their impact on T. urticae performance [22,23,24,25,26]. Most of these studies have been performed in tomato, bean and strawberry plants, whereas remarkably, no study has been conducted on the impact of endophytic entomopathogens in pepper against spider mites. Furthermore, to the best of our knowledge, no studies have been performed to assess the plant-mediated effects of entomopathogenic bacteria or of non-entomopathogenic microbes (e.g., plant growth-promoting rhizobacteria) against spider mites. Instead, Pseudomonas spp. and Bacillus spp. isolates have mainly been assessed for their acaricidal effects in mortality tests or after being sprayed on the plants [27,28]. Remarkably, all of the above studies have identified several strains of entomopathogens with plant protection capabilities when applied in the soil. For example, Canassa et al. [22] evaluated 25 entomopathogenic fungal isolates of three genera (Metarhizium, Beauveria and Cordyceps) that almost all negatively affected spider mite performance in strawberry plants, and tomato seed treatments by isolates of B. bassiana and M. robertsii but not M. brunneum resulted in a reduction in spider mite population growth [29], as did the seed inoculation of beans with isolates of B. bassiana, Cordyceps fumosorosea (Isaria fumosorosea) and Akanthomyces lecanii (Lecanicillium lecanii) [26] as well as with M. robertsii [30]. Nevertheless, the symbioses of soil microbes with other plants, such as tomato or pepper, and their interactions with aboveground spider mites remain unexplored. An exception is the recent report of a beneficial soil endophytic fungus, Fusarium solani strain K, that was shown to negatively affect spider mite performance in tomatoes via the elicitation of plant defense responses [31].
Similarly, studies performed on the effects of beneficial microbes against the green peach aphid Myzus persicae follow the general trend of assessing entomopathogenic fungi (Beauveria spp., Metarhizium spp.), and only a few have dealt with plant growth-promoting rhizobacteria (Bacillus spp. and Pseudomonas spp.) [32,33,34,35]. In the studies assessing plant-mediated effects, microbial application in the soil was performed either as a seed coating or as a soil drench. Different crop plants have been studied in this regard, such as sweet pepper, bell pepper, tobacco and Arabidopsis thaliana [32,35,36,37], with the microbial application in soil resulting in negative effects on aphid performance in accordance with the results of the present study. Beneficial fungi, such as Trichoderma spp. and the entomopathogen C. fumosorosea, have not been studied for their plant-mediated efficacy against M. persicae in pepper so far.
The mechanisms involved in the recorded effects of beneficial microbes applied in the soil on herbivore performance remain to be elucidated so that we can better exploit their potential as biological control agents. Several mechanisms can be involved, such as the production of secondary metabolites, antibiotic effects, feeding deterrents and plant defense induction [3,5,6,7,8,9,10,22]. With regard to C. fumosorosea in particular, we also cannot exclude the possibility that negative effects on herbivore performance are related to the entomopathogenic activity of the fungus colonizing the plant [3,38]. Spider mites and aphids were negatively affected on microbe-inoculated plants, suggesting the absence of nutritional benefits or that defense induction outcompetes the putative benefits of improved nutrition to the herbivores. In addition, we recorded a difference in the number of live spider mites on plants and non-inoculated control plants, suggesting that recorded differences cannot be attributed to only plant responses affecting spider mite reproduction. We only recorded the number of live individuals or eggs on the plants, so we cannot draw safe conclusions about the potent mechanisms involved. Further studies are required to elucidate whether our results could be explained by one or more of the above mechanisms.
In addition, we should note that, despite all of the microbes negatively affecting the two herbivores, we found a variation in the efficacy of each microbe depending on the species tested. Aphids are phloem-feeders and spider mites are mesophyll cell content feeders, and the ability of specific microbes to move within the plants, i.e., from roots to leaves, may vary, although most possibly in a genus-specific manner [22]. In addition, we did not record any effect on the plant biomass 14 days after infestation with the herbivores, which may be explained by a potent trade-off between plant growth and defense induction [6,16,39,40,41] in the inoculated plants compared with the non-inoculated herbivore-infested plants. Finally, as most of the microbes tested herein are biofungicides, we cannot exclude mechanisms underlying their efficacy against fungi to be efficient against arthropod pests. Hence, understanding the molecular and chemical mechanisms involved in these interactions will cast light on the ways that beneficial soil microbes affect aboveground herbivores.
With regard to the applied aspects of our findings, we show herein that beneficial soil microbes currently applied as biofungicides can also exert plant protection capabilities against aboveground arthropod pests. Therefore, our results contribute to the growing literature, showing that beneficial soil microbes can shape plant–herbivore interactions to the benefit of the plants. Moreover, we reveal herein the ability of C. fumosorosea, an entomopathogen largely unexplored as an endophyte, and less as a plant growth-promoter, to suppress herbivore populations via the plant. The efficacy of entomopathogens may be affected by biotic and abiotic factors when sprayed on the plants under field conditions; hence, applying them in the soil may improve their consistency in impacting pests. Nevertheless, long-term experiments coupled with field trials are still required to assess the population dynamics of the two herbivores on microbe-inoculated plants under field-realistic conditions as well as to assess the impact of the tested soil microbes on plant productivity. Although we did not assess the plants’ colonization with the microbes on the day of our recordings (35 days after the first microbe soil drench), our results indicate persistence in the plant-mediated efficacy of the tested microbes against pests in pepper. In practice, persistence in microbial colonization in plants could be prolonged by repeating microbial applications. Finally, the effects on the natural enemies of the herbivores should also be investigated to be able to fully exploit the benefits of both groups of biological control agents (for a trade-off, see [42]). This is particularly important considering that beneficial soil microbes also impact indirect plant defenses via altering the composition and emission rate of volatiles emitted by inoculated plants in response to herbivory [6,8,31].
5. Conclusions
Beneficial microbes applied as a soil drench can exert plant-mediated effects and can negatively affect the performance of spider mites and aphids in pepper. The mechanisms involved as well as their effects on natural enemies need to be studied in order identify the potential of each microbe against aboveground arthropod pests.
Author Contributions
Conceptualization, M.L.P. and G.D.B.; methodology, M.L.P. and G.D.B.; validation, M.L.P. and K.S.; formal analysis, G.D.B.; investigation, M.L.P., K.S. and I.K.; resources, G.D.B.; writing—original draft preparation, M.L.P. and G.D.B.; writing—review and editing, M.L.P. and G.D.B.; visualization, G.D.B.; supervision, G.D.B. and M.L.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was co-financed by Greece and the European Union (European Social Fund- ESF) through the Operational Programme “Human Resources Development, Education and Lifelong Learning 2014–2020” in the context of the project “Beneficial soil microbes to combat major agricultural crop pests” (MIS 5049563).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated and analyzed during the current study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef]
- Pappas, M.L.; Baptista, P.; Broufas, G.D.; Dalakouras, A.; Djobbi, W.; Flors, V.; Guerfali, M.M.; Khayi, S.; Mentag, R.; Pastor, V.; et al. Biological and Molecular Control Tools in Plant Defense. In Plant Defence: Biological Control; Mérillon, J.-M., Ramawat, K.G., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 3–43. [Google Scholar]
- Gruden, K.; Lidoy, J.; Petek, M.; Podpečan, V.; Flors, V.; Papadopoulou, K.K.; Pappas, M.L.; Martinez-Medina, A.; Bejarano, E.; Biere, A.; et al. Ménage à Trois: Unraveling the Mechanisms Regulating Plant–Microbe–Arthropod Interactions. Trends Plant Sci. 2020, 25, 1215–1226. [Google Scholar] [CrossRef] [PubMed]
- Hartley, S.E.; Gange, A.C. Impacts of plant symbiotic fungi on insect herbivores: Mutualism in a multitrophic context. Annu. Rev. Entomol. 2009, 54, 323–342. [Google Scholar] [CrossRef]
- Pineda, A.; Zheng, S.J.; van Loon, J.J.A.; Pieterse, C.M.J.; Dicke, M. Helping plants to deal with insects: The role of beneficial soil-borne microbes. Trends Plant Sci. 2010, 15, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J. Trichoderma as biocontrol agent against pests: New uses for a mycoparasite. Biol. Control. 2021, 159, 104634. [Google Scholar] [CrossRef]
- Rasmann, S.; Bennett, A.; Biere, A.; Karley, A.; Guerrieri, E. Root symbionts: Powerful drivers of plant above- and belowground indirect defenses. Insect Sci. 2017, 24, 947–960. [Google Scholar] [CrossRef]
- Vega, F.E. Insect pathology and fungal endophytes. J. Invertebr. Pathol. 2008, 98, 277–279. [Google Scholar] [CrossRef]
- Vega, F.E. The use of fungal entomopathogens as endophytes in biological control: A review. Mycologia 2018, 110, 4–30. [Google Scholar] [CrossRef]
- Sindhu, S.S.; Sehrawat, A.; Sharma, R.; Khandelwal, A. Biological Control of Insect Pests for Sustainable Agriculture. In Advances in Soil Microbiology: Recent Trends and Future Prospects: Volume 2: Soil-Microbe-Plant Interaction; Adhya, T.K., Mishra, B.B., Annapurna, K., Verma, D.K., Kumar, U., Eds.; Springer: Singapore, 2017; pp. 189–218. [Google Scholar]
- Singh, A.; Bhardwaj, R.; Singh, I.K. Biocontrol Agents: Potential of Biopesticides for Integrated Pest Management. In Biofertilizers for Sustainable Agriculture and Environment; Giri, B., Prasad, R., Wu, Q.-S., Varma, A., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 413–433. [Google Scholar]
- Finkel, O.M.; Castrillo, G.; Herrera Paredes, S.; Salas González, I.; Dangl, J.L. Understanding and exploiting plant beneficial microbes. Curr. Opin. Plant Biol. 2017, 38, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Pineda, A.; Soler, R.; Weldegergis, B.T.; Shimwela, M.M.; Van Loon, J.J.A.; Dicke, M. Non-pathogenic rhizobacteria interfere with the attraction of parasitoids to aphid-induced plant volatiles via jasmonic acid signalling. Plant Cell Environ. 2013, 36, 393–404. [Google Scholar] [CrossRef]
- Smith, S.E.; Smith, F.A. Roles of arbuscular mycorrhizas in plant nutrition and growth: New paradigms from cellular to ecosystem scales. Annu. Rev. Plant Biol. 2011, 62, 227–250. [Google Scholar] [CrossRef] [Green Version]
- Shikano, I.; Rosa, C.; Tan, C.W.; Felton, G.W. Tritrophic Interactions: Microbe-Mediated Plant Effects on Insect Herbivores. Annu. Rev. Phytopathol. 2017, 55, 313–331. [Google Scholar] [CrossRef]
- Pozo, M.J.; Zabalgogeazcoa, I.; Vazquez de Aldana, B.R.; Martinez-Medina, A. Untapping the potential of plant mycobiomes for applications in agriculture. Curr. Opin. Plant Biol. 2021, 60, 102034. [Google Scholar] [CrossRef]
- Food and Agriculture Organization (FAO). FAOSTAT Statistical Database of the United Nation Food and Agriculture Organization (FAO) Statistical Division. Crops and Livestock Products. 2019. Available online: http://www.fao.org/faostat/en/#data/QCL (accessed on 13 August 2021).
- Helle, W.; Sabelis, M.W. Spider Mites Their Biology, Natural Enemies and Control; Elsevier Science Publishers: Amsterdam, The Netherlands, 1985. [Google Scholar]
- Blackman, R.L.; Eastop, V.F. Aphids on the World’s Crops: An Identification and Information Guide; John Wiley: Chichester, UK, 1984. [Google Scholar]
- IBM Corp. Released 2020. IBM SPSS Statistics for Windows, SPSS Version 27.0; IBM Corp: Armonk, NY, USA, 2020. [Google Scholar]
- Canassa, F.; D’Alessandro, C.P.; Sousa, S.B.; Demétrio, C.G.B.; Meyling, N.V.; Klingen, I.; Delalibera, I., Jr. Fungal isolate and crop cultivar influence the beneficial effects of root inoculation with entomopathogenic fungi in strawberry. Pest Manag. Sci. 2020, 76, 1472–1482. [Google Scholar] [CrossRef] [PubMed]
- Canassa, F.; Esteca, F.C.N.; Moral, R.A.; Meyling, N.V.; Klingen, I.; Delalibera, I. Root inoculation of strawberry with the entomopathogenic fungi Metarhizium robertsii and Beauveria bassiana reduces incidence of the twospotted spider mite and selected insect pests and plant diseases in the field. J. Pest Sci. 2020, 93, 261–274. [Google Scholar] [CrossRef]
- Castro, T.; Eilenberg, J.; Delalibera, I. Exploring virulence of new and less studied species of Metarhizium spp. from Brazil for two-spotted spider mite control. Exp. Appl. Acarol. 2018, 74, 139–146. [Google Scholar] [CrossRef]
- Elhakim, E.; Mohamed, O.; Elazouni, I. Virulence and proteolytic activity of entomopathogenic fungi against the two-spotted spider mite, Tetranychus urticae Koch (Acari: Tetranychidae). Egypt. J. Biol. Pest Control. 2020, 30, 30. [Google Scholar] [CrossRef] [Green Version]
- Dash, C.K.; Bamisile, B.S.; Keppanan, R.; Qasim, M.; Lin, Y.; Islam, S.U.; Hussain, M.; Wang, L. Endophytic entomopathogenic fungi enhance the growth of Phaseolus vulgaris L. (Fabaceae) and negatively affect the development and reproduction of Tetranychus urticae Koch (Acari: Tetranychidae). Microb. Pathog. 2018, 125, 385–392. [Google Scholar] [CrossRef]
- Aksoy, H.M.; Ozman-Sullivan, S.K.; Ocal, H.; Celik, N.; Sullivan, G.T. The effects of Pseudomonas putida biotype B on Tetranychus urticae (Acari: Tetranychidae). Exp. Appl. Acarol. 2008, 46, 223. [Google Scholar] [CrossRef]
- Qessaoui, R.; Bouharroud, R.; Amarraque, A.; Ajerrar, A.; El Hassan, M.; Chebli, B.; Dadi, M.; Elaini, R.; El Filali, F.; Walters, A.S. Ecological applications of Pseudomonas as a biopesticide to control two-spotted mite Tetranychus urticae: Chitinase and HCN production. J. Plant Prot. Res. 2017, 57, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Rasool, S.; Cárdenas, P.D.; Pattison, D.I.; Jensen, B.; Meyling, N.V. Isolate-specific effect of entomopathogenic endophytic fungi on population growth of two-spotted spider mite (Tetranychus urticae Koch) and levels of steroidal glycoalkaloids in tomato. J. Chem. Ecol. 2021, 47, 476–488. [Google Scholar] [CrossRef]
- Canassa, F.; Tall, S.; Moral, R.A.; Lara, I.A.R.D.; Delalibera, I.; Meyling, N.V. Effects of bean seed treatment by the entomopathogenic fungi Metarhizium robertsii and Beauveria bassiana on plant growth, spider mite populations and behavior of predatory mites. Biol. Control. 2019, 132, 199–208. [Google Scholar] [CrossRef]
- Pappas, M.L.; Liapoura, M.; Papantoniou, D.; Avramidou, M.; Kavroulakis, N.; Weinhold, A.; Broufas, G.D.; Papadopoulou, K.K. The beneficial endophytic fungus Fusarium solani strain K alters tomato responses against spider mites to the benefit of the plant. Front. Plant Sci. 2018, 9, 1603. [Google Scholar] [CrossRef] [Green Version]
- Herman, M.A.B.; Nault, B.A.; Smart, C.D. Effects of plant growth-promoting rhizobacteria on bell pepper production and green peach aphid infestations in New York. Crop. Prot. 2008, 27, 996–1002. [Google Scholar] [CrossRef]
- Jaber, L.R.; Araj, S.-E. Interactions among endophytic fungal entomopathogens (Ascomycota: Hypocreales), the green peach aphid Myzus persicae Sulzer (Homoptera: Aphididae), and the aphid endoparasitoid Aphidius colemani Viereck (Hymenoptera: Braconidae). Biol. Control. 2018, 116, 53–61. [Google Scholar] [CrossRef]
- Naeem, M.; Aslam, Z.; Khaliq, A.; Ahmed, J.N.; Nawaz, A.; Hussain, M. Plant growth promoting rhizobacteria reduce aphid population and enhance the productivity of bread wheat. Braz. J. Microbiol. 2018, 49, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Zhao, X.; Huang, S.; Deng, J.; Li, X.; Luo, Z.; Zhang, Y. Pest management via endophytic colonization of tobacco seedlings by the insect fungal pathogen Beauveria bassiana. Pest Manag. Sci. 2021, 77, 2007–2018. [Google Scholar] [CrossRef]
- Boutard-Hunt, C.; Smart, C.D.; Thaler, J.; Nault, B.A. Impact of plant growth-promoting rhizobacteria and natural enemies on Myzus persicae (Hemiptera: Aphididae) infestations in pepper. J. Econ. Entomol. 2009, 102, 2183–2191. [Google Scholar] [CrossRef]
- Brock, A.K.; Berger, B.; Schreiner, M.; Ruppel, S.; Mewis, I. Plant growth-promoting bacteria Kosakonia radicincitans mediate anti-herbivore defense in Arabidopsis thaliana. Planta 2018, 248, 1383–1392. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, G. The entomopathogenic fungi Isaria farinosa (formerly Paecilomyces farinosus) and the Isaria fumosorosea species complex (formerly Paecilomyces fumosoroseus): Biology, ecology and use in biological control. Biocontrol Sci. Technol. 2008, 18, 865–901. [Google Scholar] [CrossRef]
- Kempel, A.; Schmidt, A.K.; Brandl, R.; Schädler, M. Support from the underground: Induced plant resistance depends on arbuscular mycorrhizal fungi. Funct. Ecol. 2010, 24, 293–300. [Google Scholar] [CrossRef]
- Gehring, C.; Bennett, A. Mycorrhizal fungal-plant-insect interactions: The importance of a community approach. Environ. Entomol. 2009, 38, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Pozo, M.J.; Azcón-Aguilar, C. Unraveling mycorrhiza-induced resistance. Curr. Opin. Plant Biol. 2007, 10, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Blubaugh, C.K.; Carpenter-Boggs, L.; Reganold, J.P.; Schaeffer, R.N.; Snyder, W.E. Bacteria and competing herbivores weaken top–down and bottom–up aphid suppression. Front. Plant Sci. 2018, 9, 1239. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Plant-mediated effects of beneficial microbes on two-spotted spider mite performance in pepper. Plants were inoculated at transplantation and one week later with the different bacterial (A,B) and fungal (C,D) microbes. Shown are the means (±SE) of T. urticae individuals (A,C) as well as eggs (B,D) per plant 14 days following infestation with 45 adult females per plant (n = 6 per treatment). In each panel, asterisks indicate significant differences between each treatment and the respective control (t-test; * p < 0.05, ** p < 0.01, *** p < 0.001); “ns” indicates nonsignificant differences: p > 0.05).
Figure 1.
Plant-mediated effects of beneficial microbes on two-spotted spider mite performance in pepper. Plants were inoculated at transplantation and one week later with the different bacterial (A,B) and fungal (C,D) microbes. Shown are the means (±SE) of T. urticae individuals (A,C) as well as eggs (B,D) per plant 14 days following infestation with 45 adult females per plant (n = 6 per treatment). In each panel, asterisks indicate significant differences between each treatment and the respective control (t-test; * p < 0.05, ** p < 0.01, *** p < 0.001); “ns” indicates nonsignificant differences: p > 0.05).

Figure 2.
Plant-mediated effects of beneficial microbes on green peach aphid performance in peppers. Plants were inoculated at transplantation and one week later with the different bacterial (A) and fungal (B) microbes. Shown are the means (±SE) of M. persicae individuals 14 days following infestation with 10 adult females per plant (n = 6 per treatment). In each panel, asterisks indicate significant differences between each treatment and the respective control (t-test; * p < 0.05, ** p < 0.01, *** p < 0.001).
Figure 2.
Plant-mediated effects of beneficial microbes on green peach aphid performance in peppers. Plants were inoculated at transplantation and one week later with the different bacterial (A) and fungal (B) microbes. Shown are the means (±SE) of M. persicae individuals 14 days following infestation with 10 adult females per plant (n = 6 per treatment). In each panel, asterisks indicate significant differences between each treatment and the respective control (t-test; * p < 0.05, ** p < 0.01, *** p < 0.001).

Figure 3.
Effects of microbial colonization on plant growth of two-spotted spider mite-infested pepper. Plants were inoculated at transplantation and one week later with the different bacterial (A) and fungal (B) microbes. Shown are means (±SE) of fresh weight per plant (n = 6 per treatment) 14 days following infestation with 45 adult T. urticae females per plant (t-test; p > 0.05).
Figure 3.
Effects of microbial colonization on plant growth of two-spotted spider mite-infested pepper. Plants were inoculated at transplantation and one week later with the different bacterial (A) and fungal (B) microbes. Shown are means (±SE) of fresh weight per plant (n = 6 per treatment) 14 days following infestation with 45 adult T. urticae females per plant (t-test; p > 0.05).

Figure 4.
Effects of microbial colonization on plant growth of green peach aphid-infested peppers. Plants were inoculated at transplantation and one week later with the different bacterial (A) and fungal (B) microbes. Shown are means (±SE) of fresh weight per plant (n = 6 per treatment) 14 days following infestation with 10 M. persicae females per plant (t-test; p > 0.05).
Figure 4.
Effects of microbial colonization on plant growth of green peach aphid-infested peppers. Plants were inoculated at transplantation and one week later with the different bacterial (A) and fungal (B) microbes. Shown are means (±SE) of fresh weight per plant (n = 6 per treatment) 14 days following infestation with 10 M. persicae females per plant (t-test; p > 0.05).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Strains of beneficial microbes that were tested for their plant-mediated effects against spider mites and aphids in pepper.
Table 1.
Strains of beneficial microbes that were tested for their plant-mediated effects against spider mites and aphids in pepper.
| Strains | Product | Dose (mg/pot) | Commercial Use (Pest/Application Method) |
|---|---|---|---|
| Fungi | |||
| Trichoderma asperellum T34 | Asperello® T34 BiocontrolTM, Biobest Group NV 1 × 1012 cfu/g 1 | 3.50 | Fusarium sp., Pythium sp., Rhizoctonia sp., Phytophthora sp. (soil application) |
| Trichoderma atroviride SC1 | Vintec®, Bi-PA NV/SA 1 × 1013 cfu/g | 0.09 | Botrytis cinerea (foliar application) |
| Trichoderma asperellum TV-1 | Xedavir, Intrachem Hellas 1 × 1010 cfu/g | 0.32 | Pythium spp., Rhizoctonia solani, Verticillium spp. (soil irrigation) |
| Cordyceps (Isaria) fumosorosea Apopka 97 | PreFeRal®, Biobest Group NV 2 × 109 cfu/g | 0.64 | Trialeurodes vaporariorum, Bemisia tabaci (foliar application) |
| Bacteria | |||
| Bacillus amyloliquefaciens subsp. plantarum D747 | Amylo-X®, K&N Efthymiadis 2 × 1011 cfu/g | 1.60 | Leveillula taurica, Botrytis cinerea, Sclerotinia sp. (foliar application) |
| B. amyloliquefaciens MBI600 | Serifel®, BASF Hellas >5.5 × 1010 cfu/g | 0.32 | Botrytis cinerea (foliar application) |
| B. amyloliquefaciens FZB24 | Taegro®, Syngenta Hellas 1 × 1010 cfu/g | 0.24 | Oidium neolycopersici, Leveillula taurica, Alternaria solani (foliar application) |
| B. amyloliquefaciens QST 2808 | Sonata®, Bayer Hellas >1 × 109 cfu/g | 6.40 | Leveillula taurica, Oidium neolycopersici (foliar application) |
| Pseudomonas sp. DSMZ 13134 | Proradix®, Anthesis 6.6 × 1010 cfu/g | 0.08 | Rhizoctonia solani, Fusarium oxysporum (soil irrigation) |
1 cfu/g: colony-forming units (cfu)/gram of dry weight.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pappas, M.L.; Samaras, K.; Koufakis, I.; Broufas, G.D. Beneficial Soil Microbes Negatively Affect Spider Mites and Aphids in Pepper. Agronomy 2021, 11, 1831. https://doi.org/10.3390/agronomy11091831
AMA Style
Pappas ML, Samaras K, Koufakis I, Broufas GD. Beneficial Soil Microbes Negatively Affect Spider Mites and Aphids in Pepper. Agronomy. 2021; 11(9):1831. https://doi.org/10.3390/agronomy11091831
Chicago/Turabian StylePappas, Maria L., Konstantinos Samaras, Ioannis Koufakis, and George D. Broufas. 2021. "Beneficial Soil Microbes Negatively Affect Spider Mites and Aphids in Pepper" Agronomy 11, no. 9: 1831. https://doi.org/10.3390/agronomy11091831
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.