
Integrating Spatiotemporal Epidemiology, Eco-Phylogenetics, and Distributional Ecology to Assess West Nile Disease Risk in Horses

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Disease Data
2.2. Virus Surveillance
2.3. Climate Data
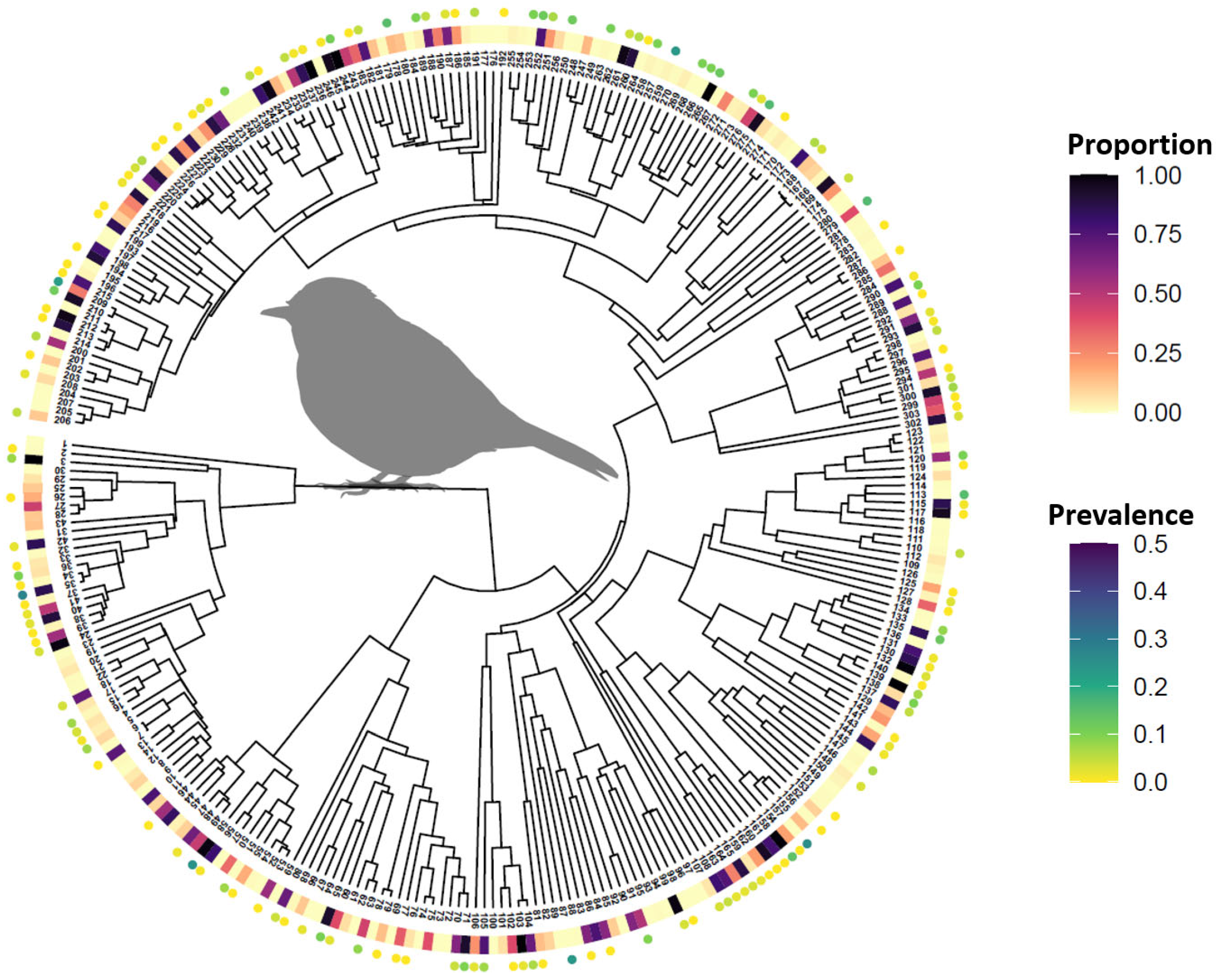
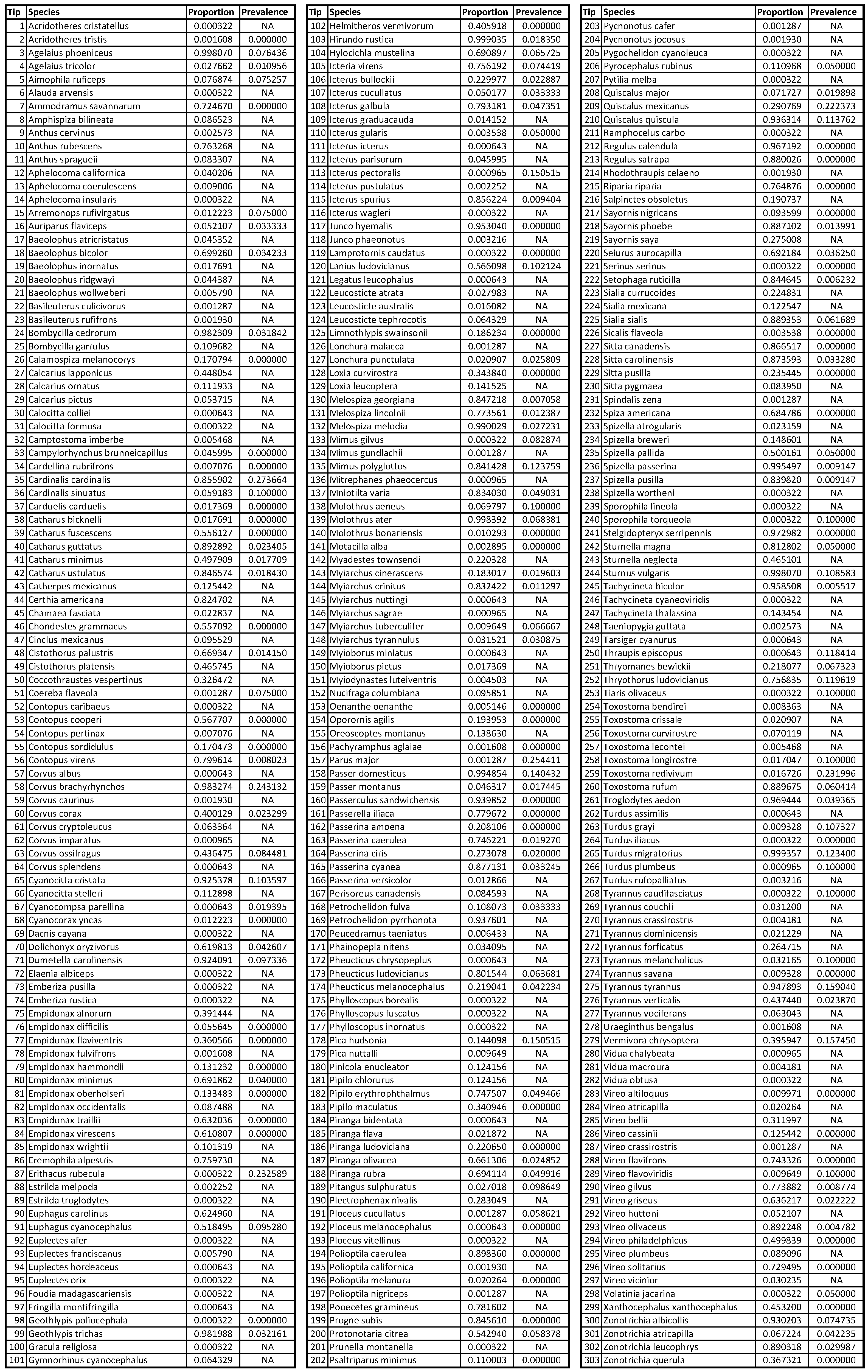
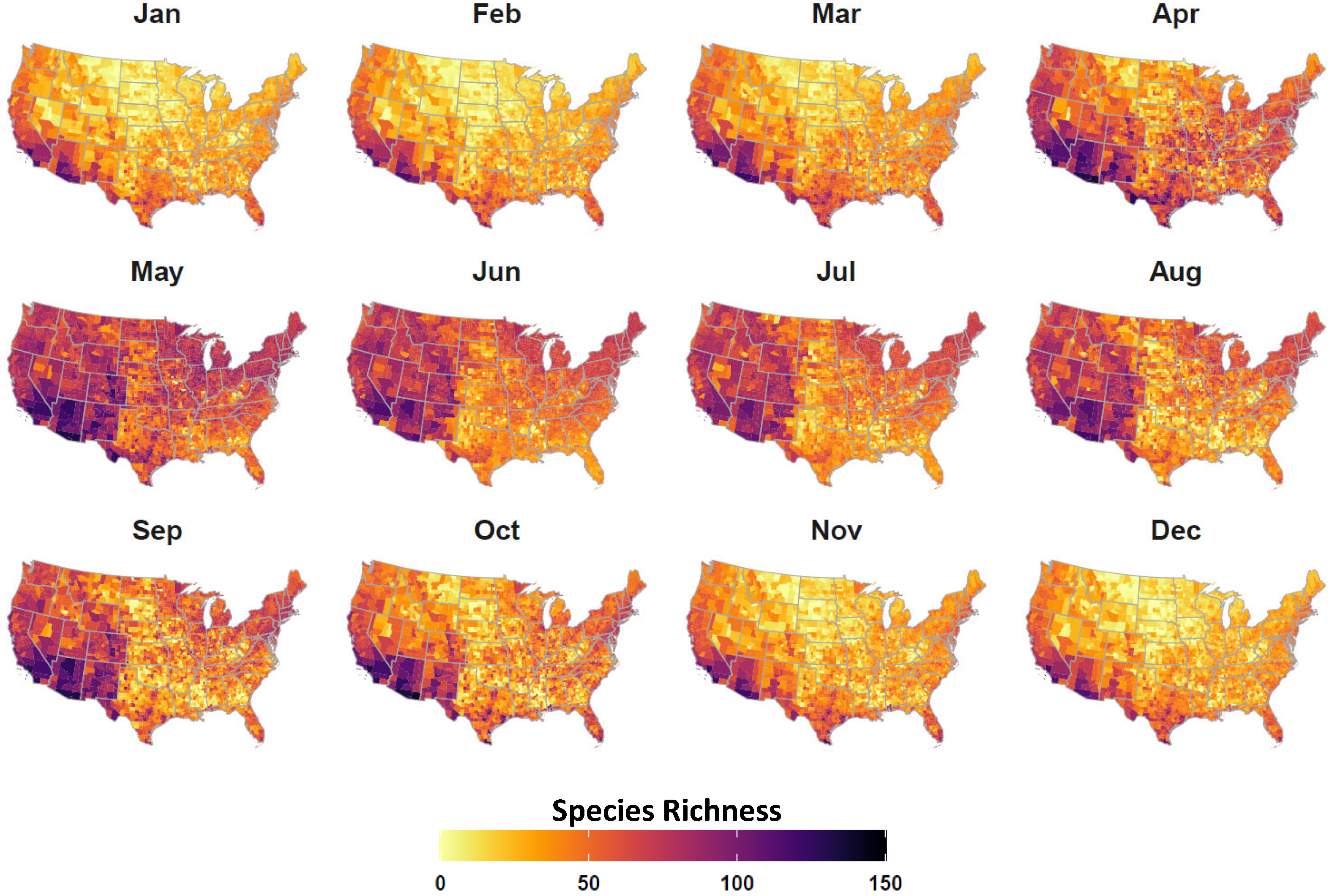
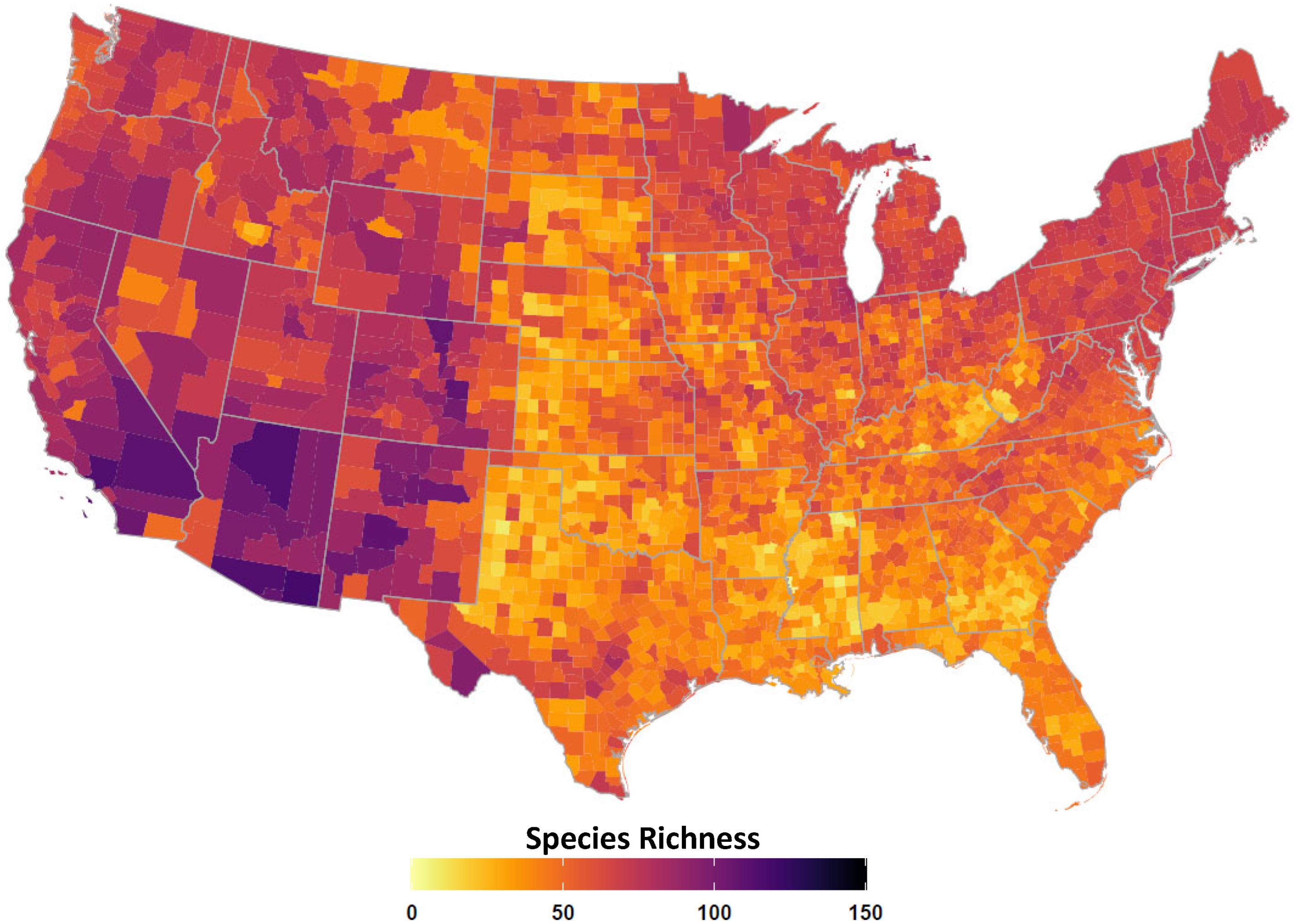
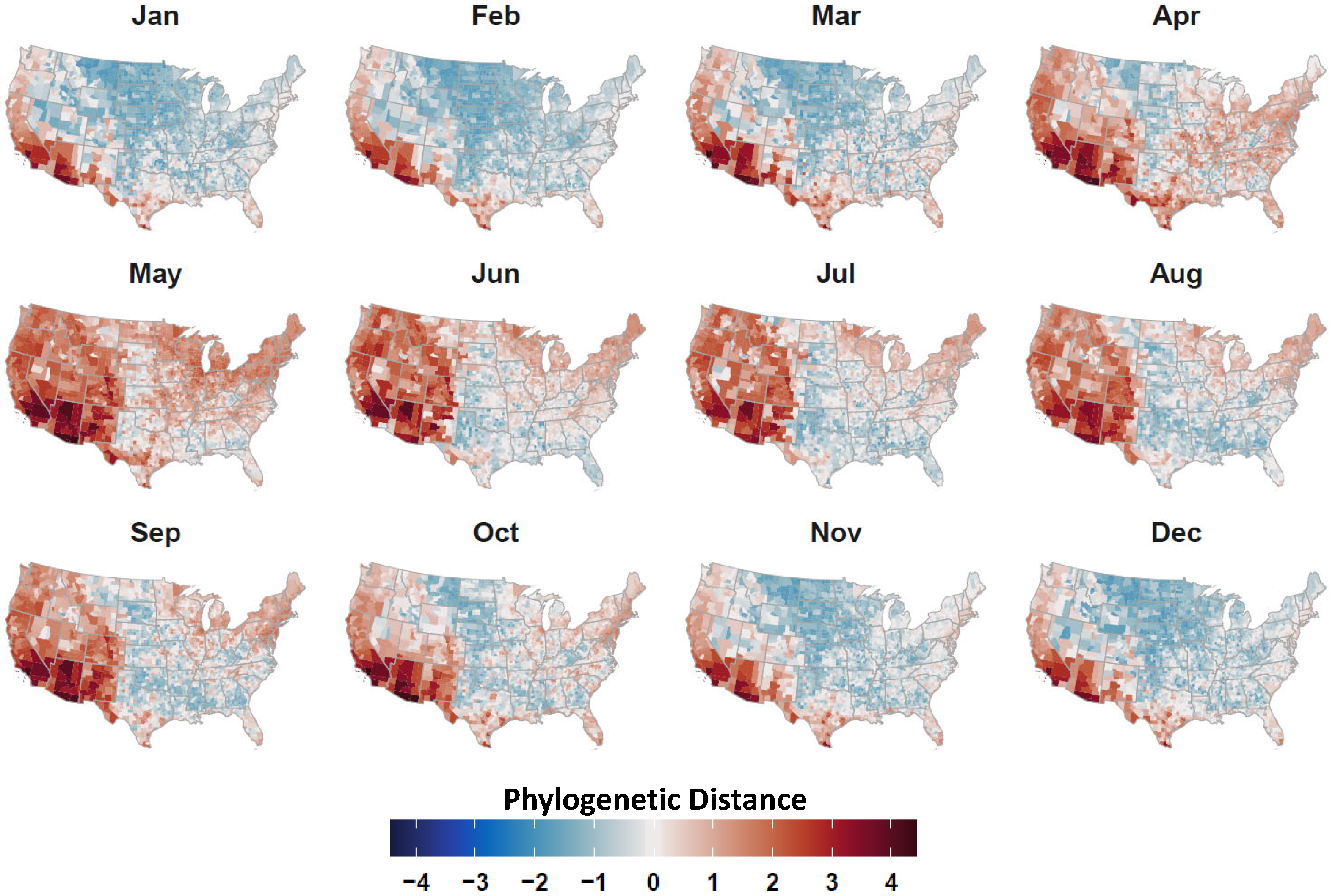
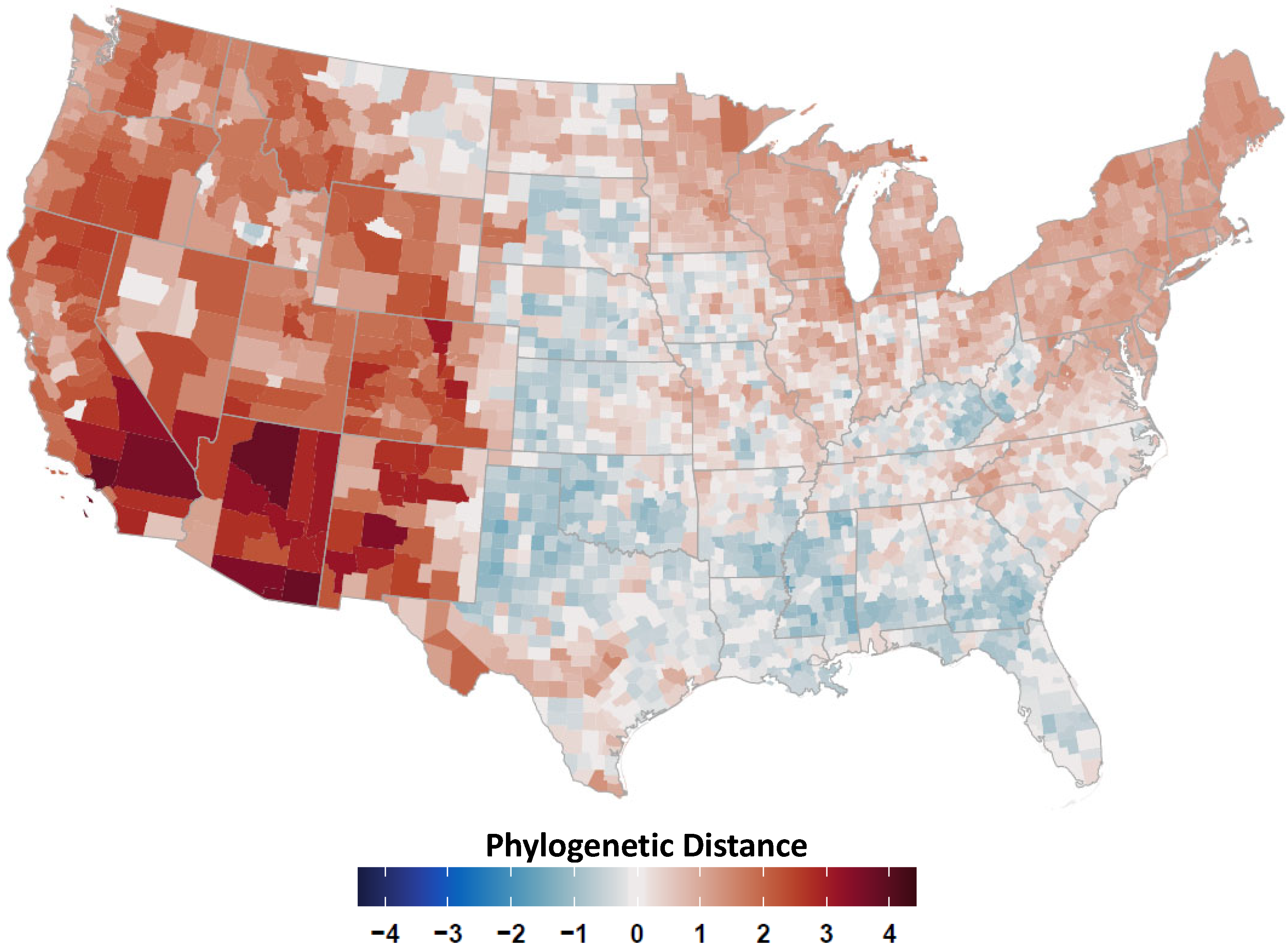
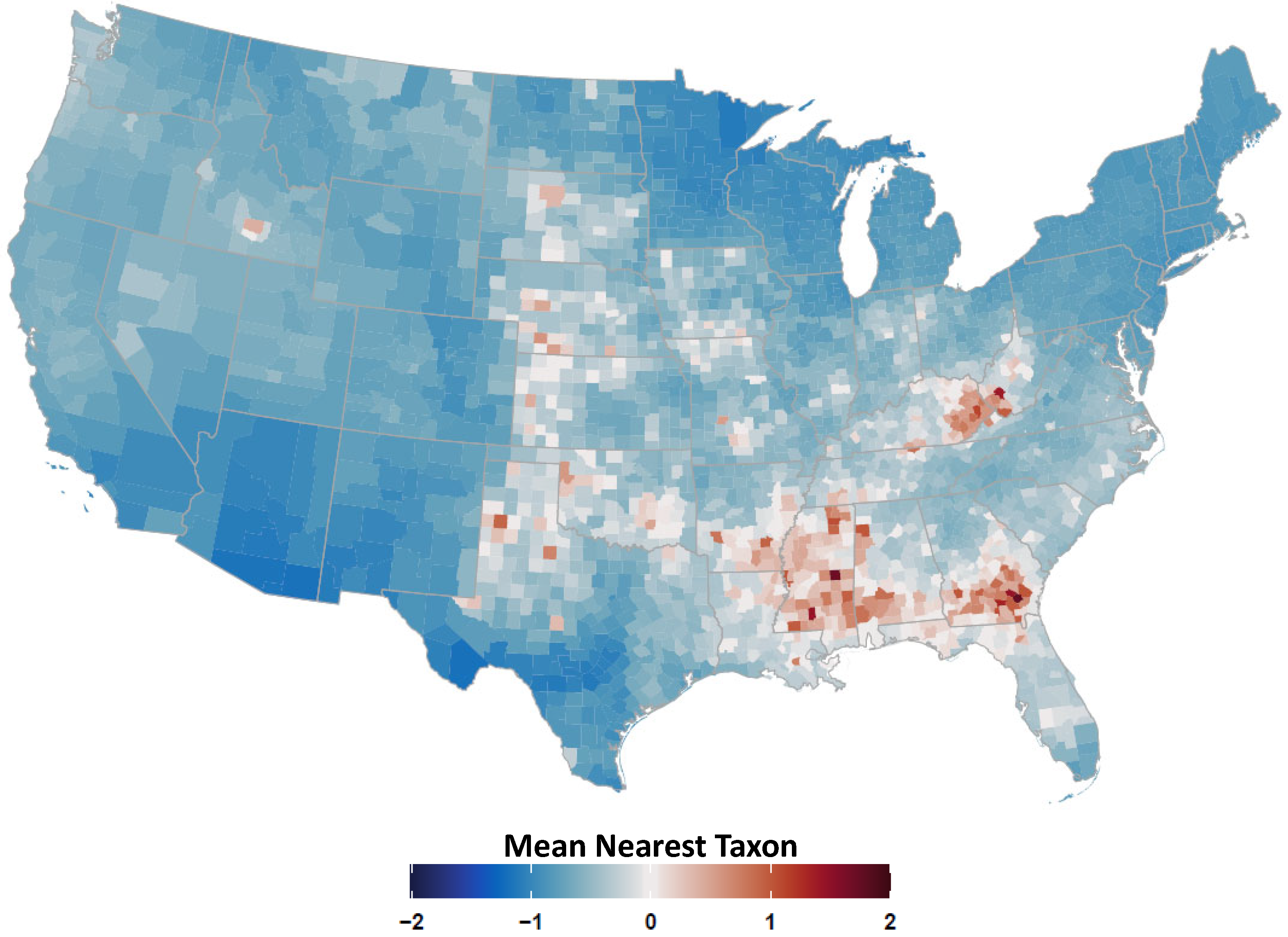
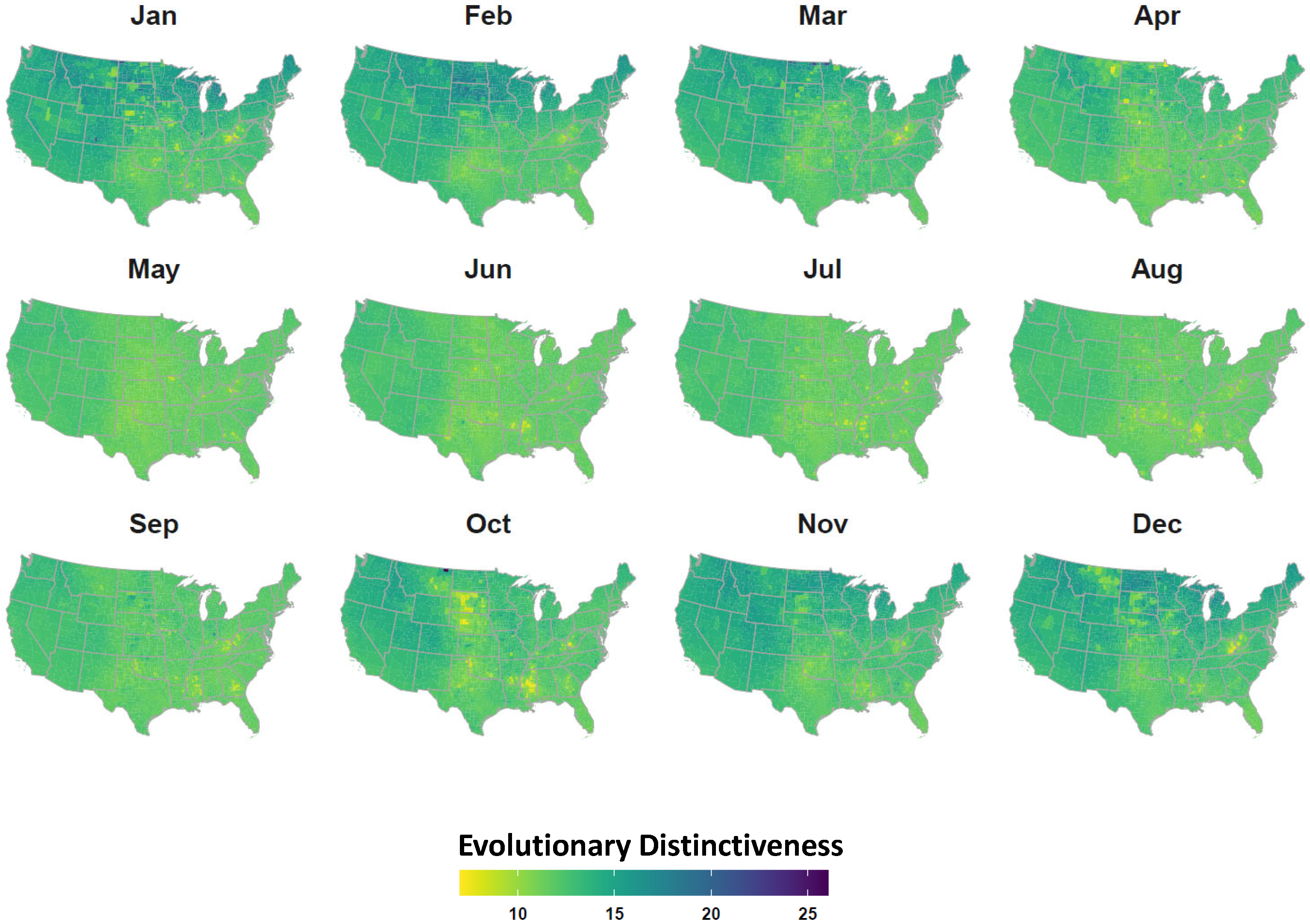
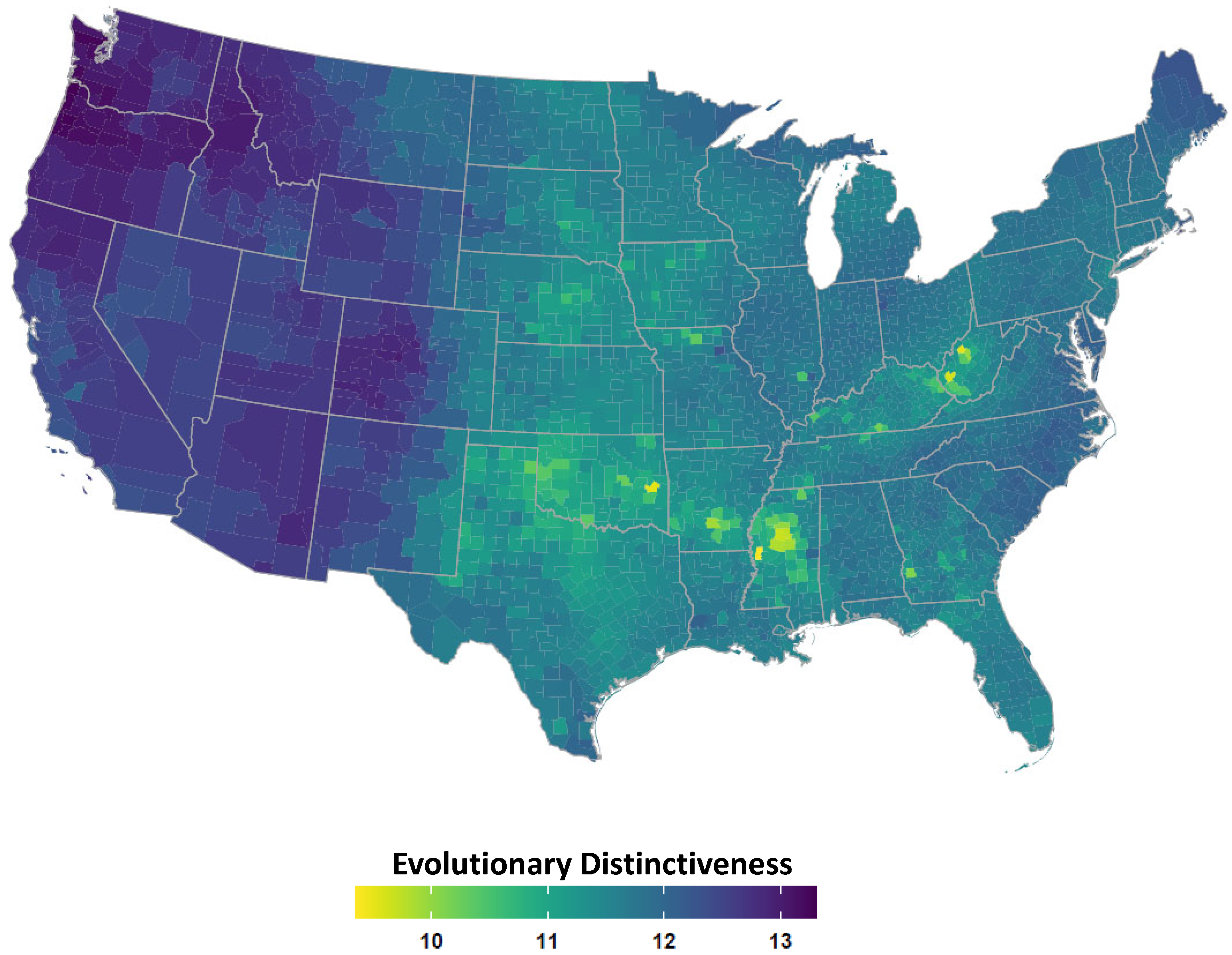
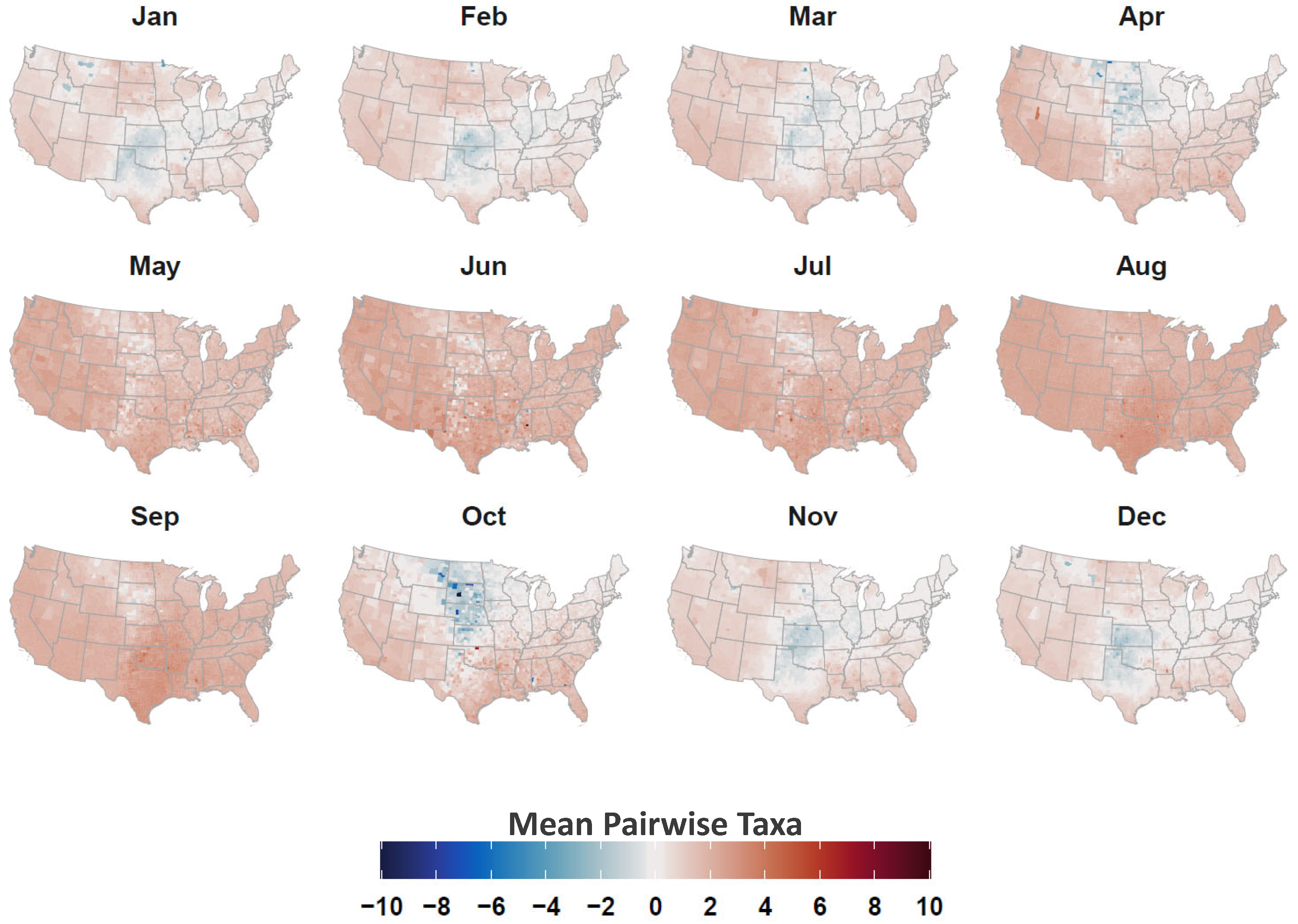
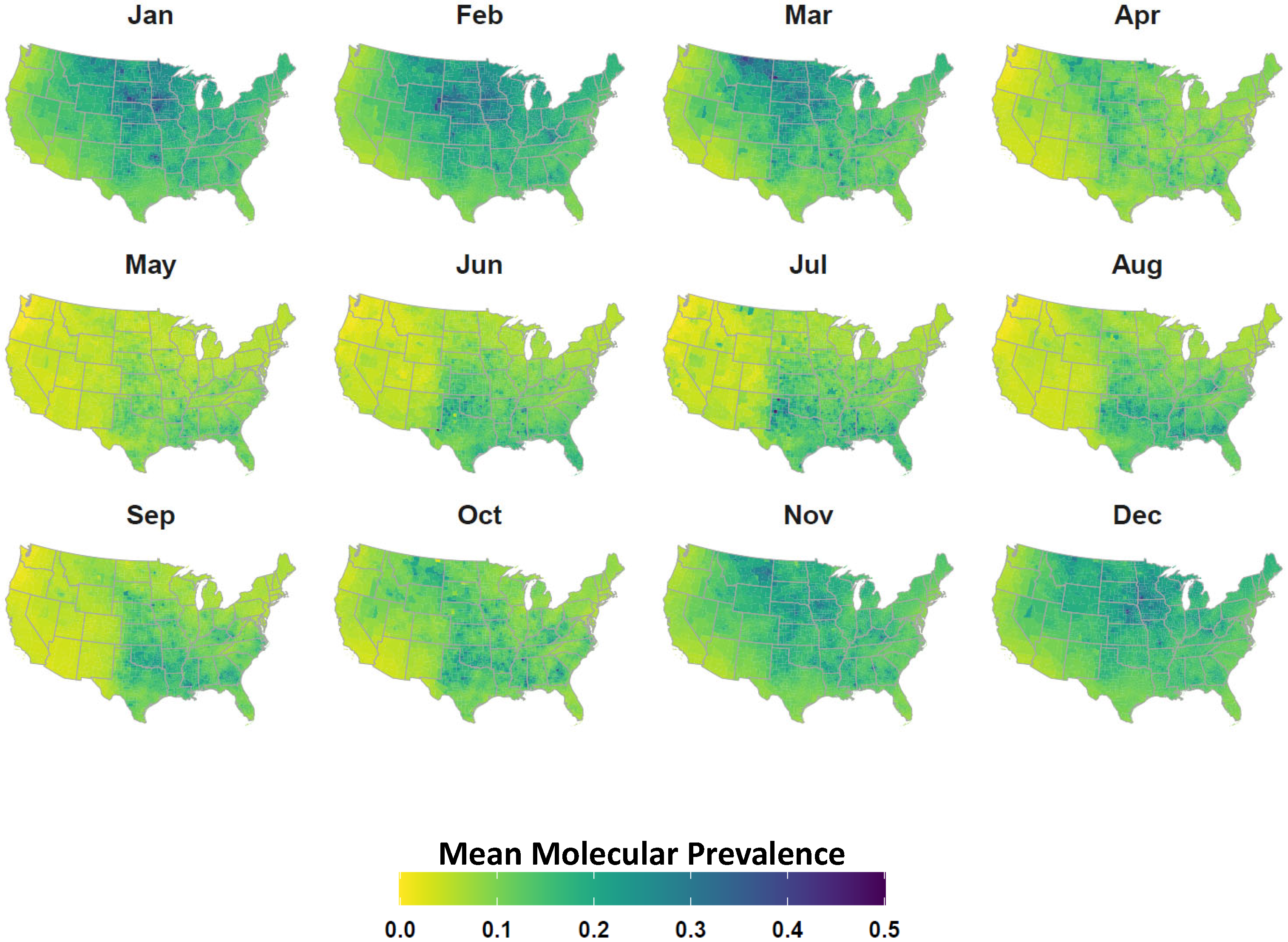
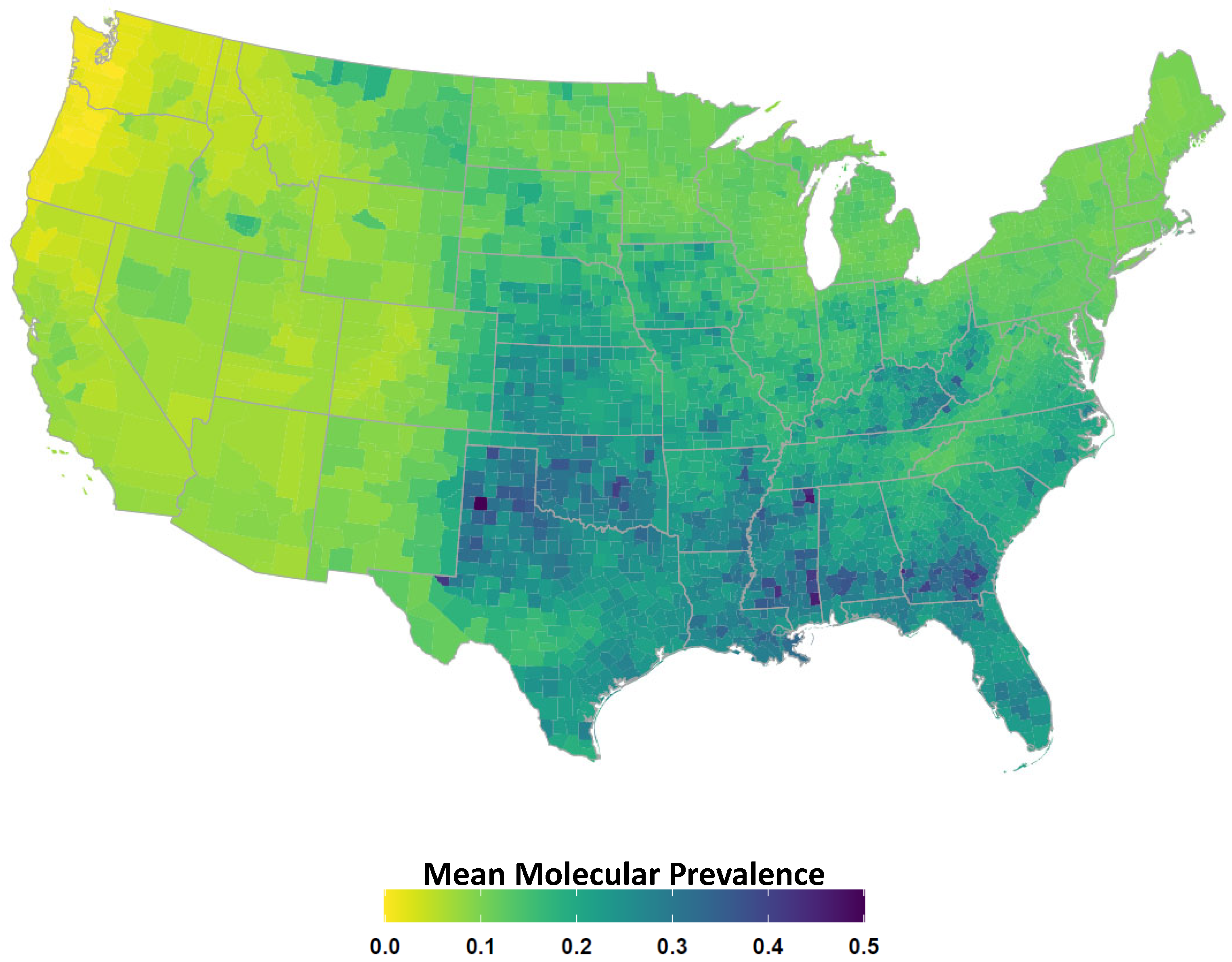
2.4. Avian Host Occurrence, Prevalence, and Phylogenetic Data
2.5. Land Cover and Human Demographic Information
2.6. Disease Model
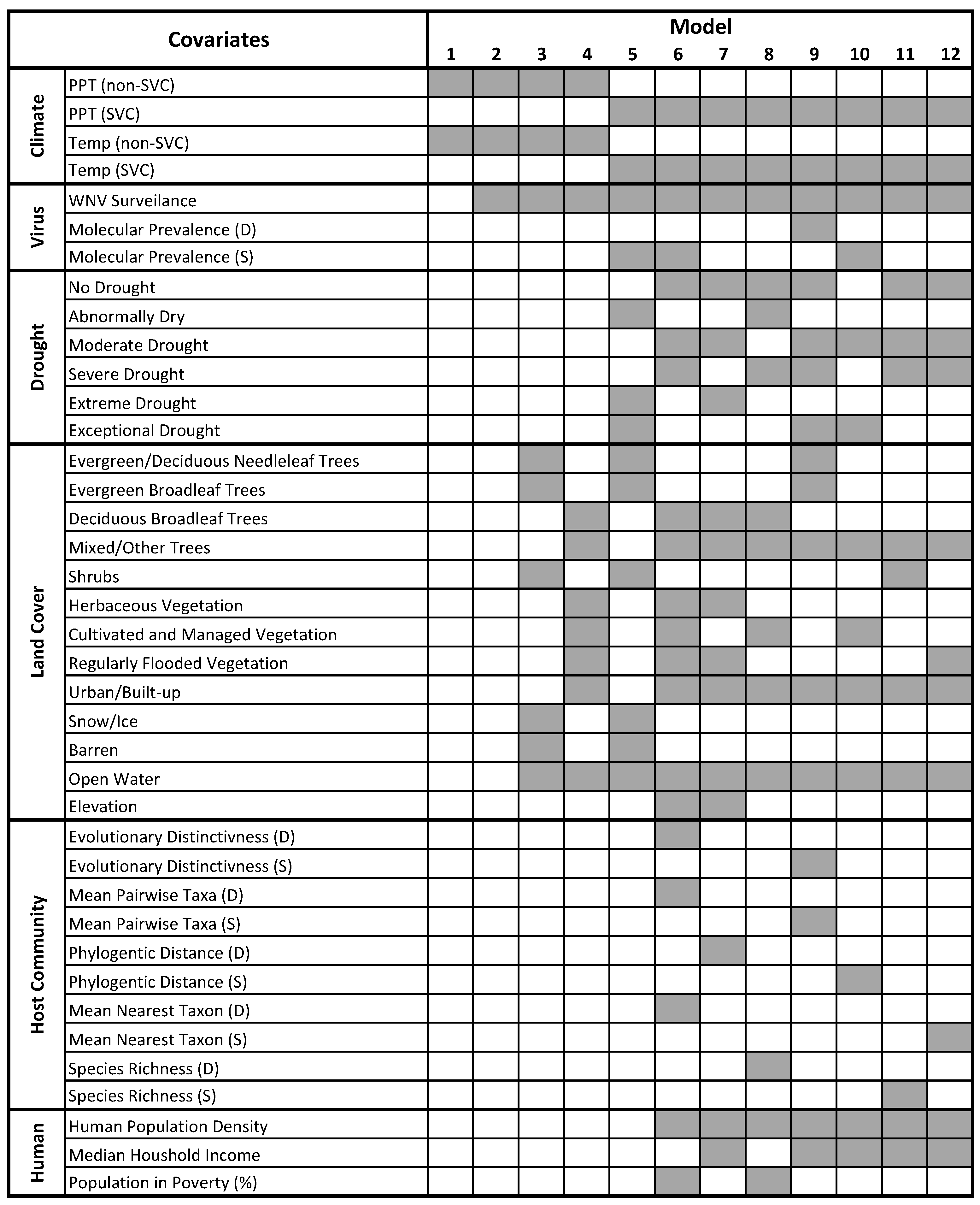
2.7. Model Selection, Consensus, and Validation
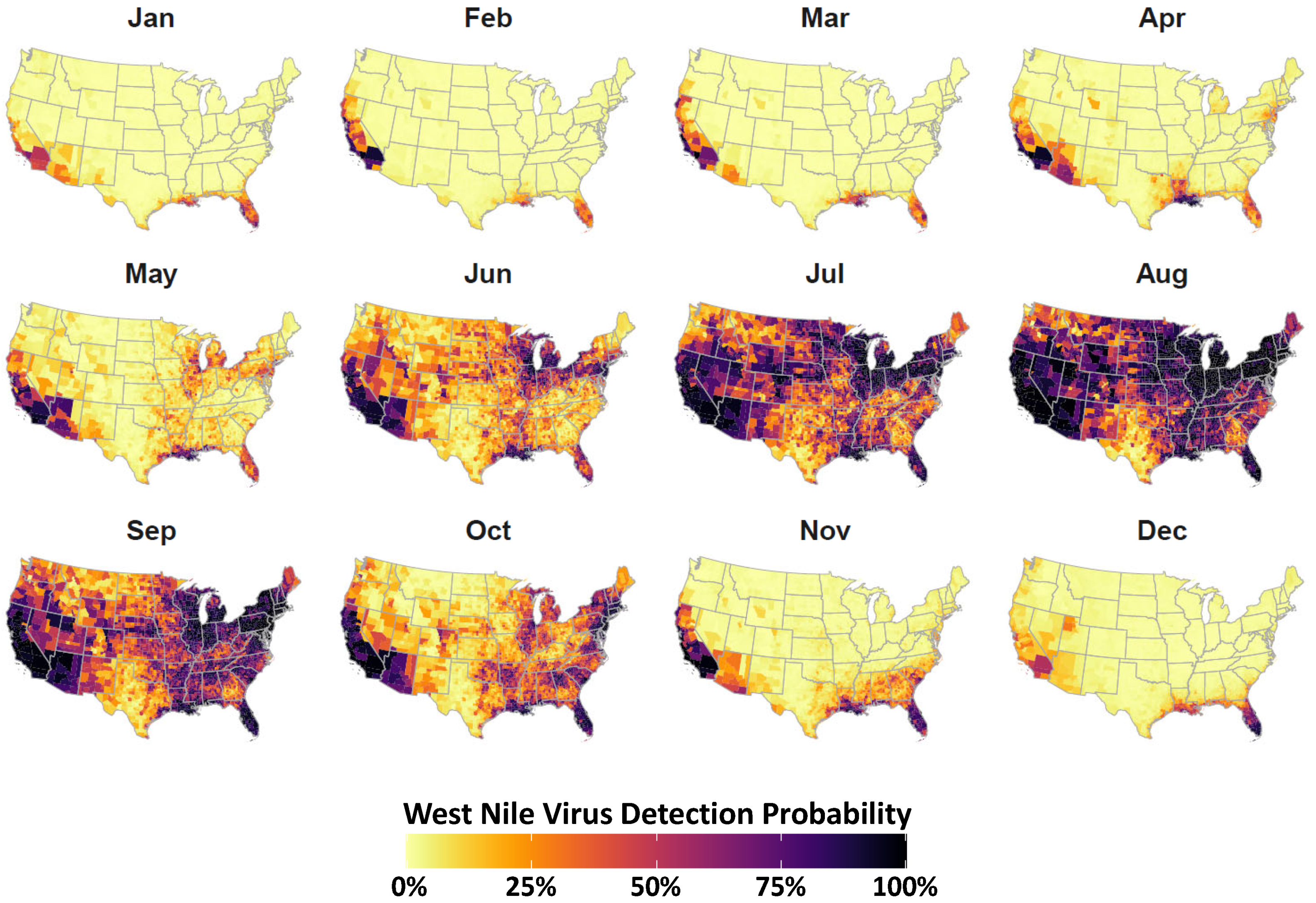
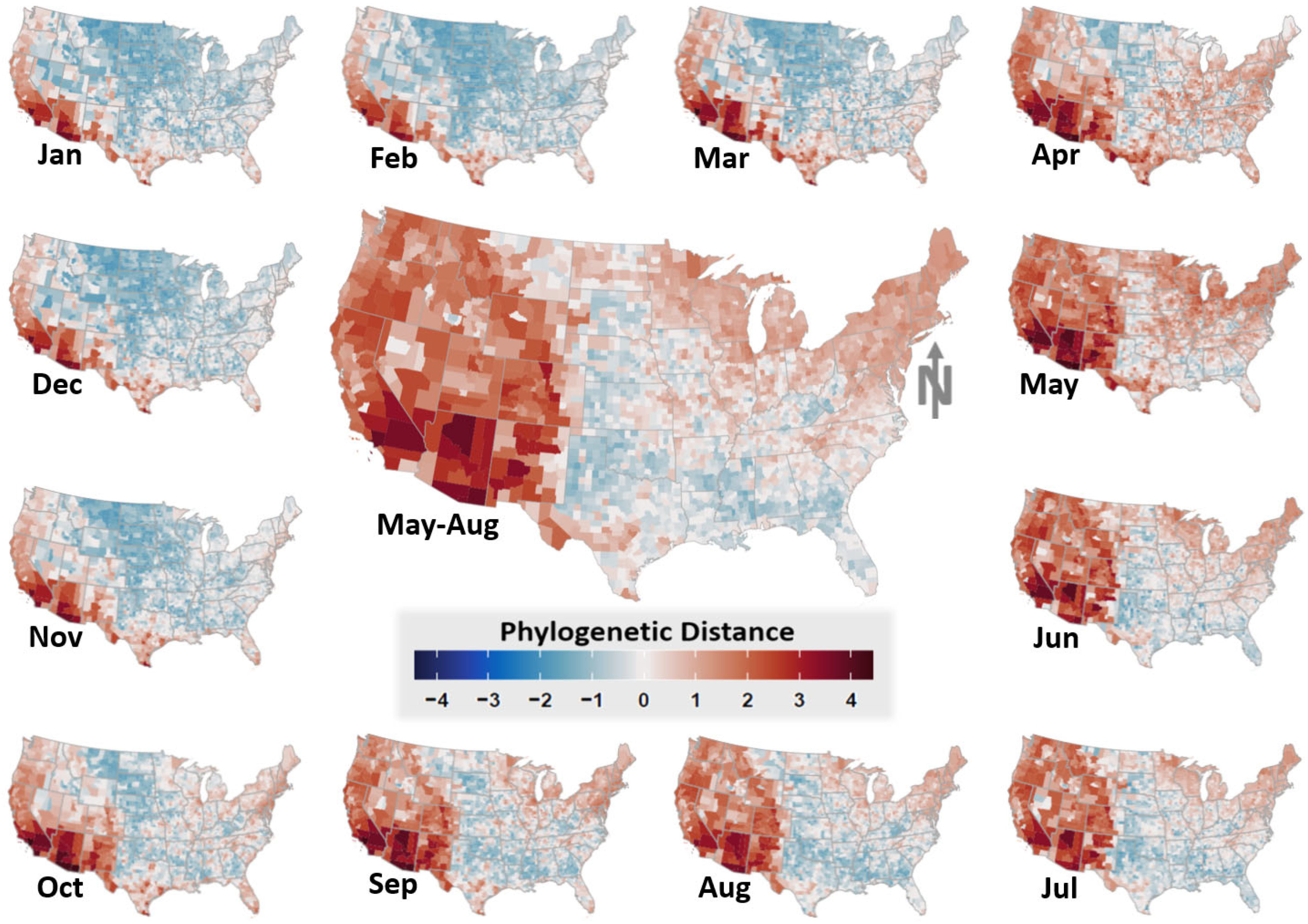
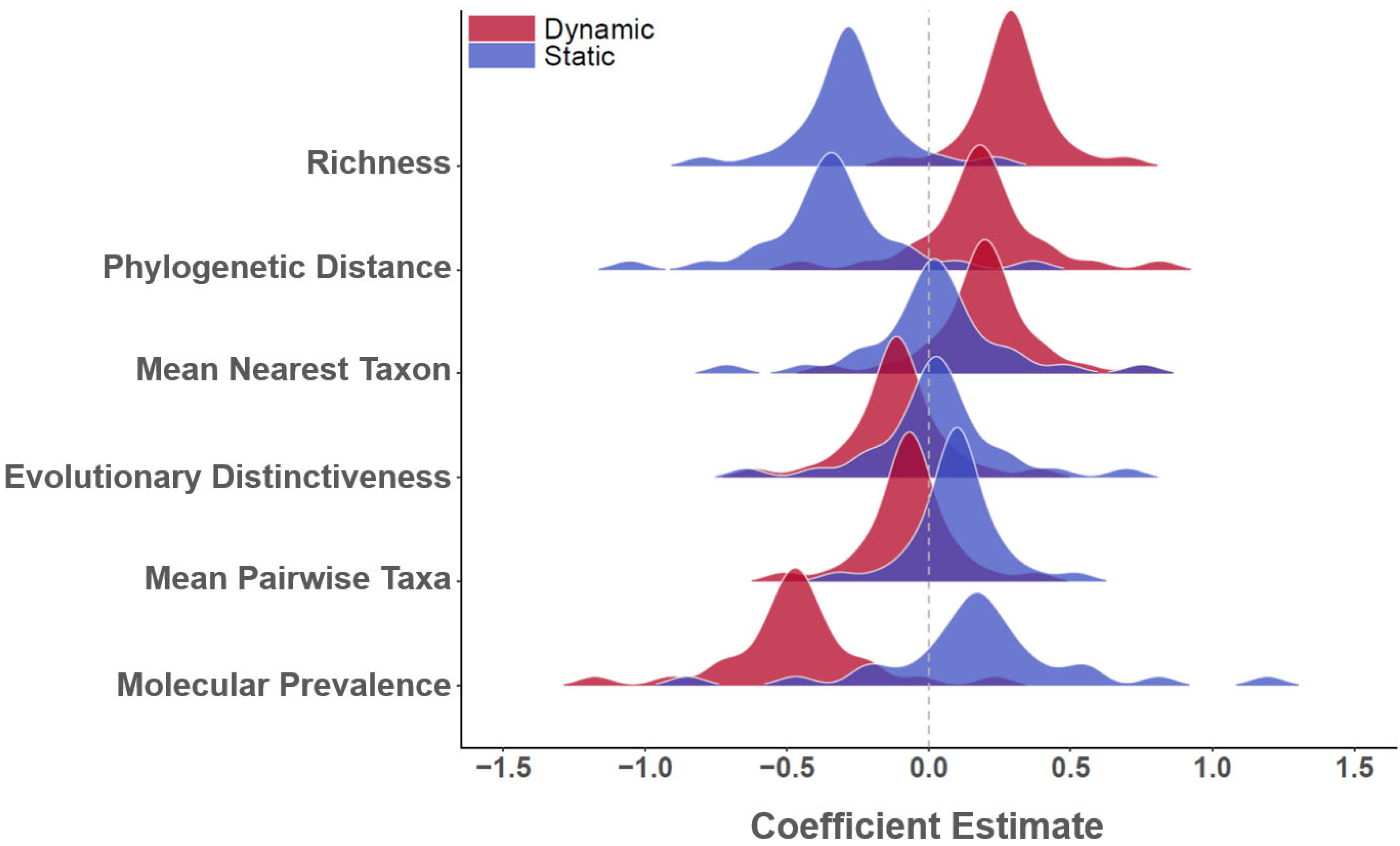
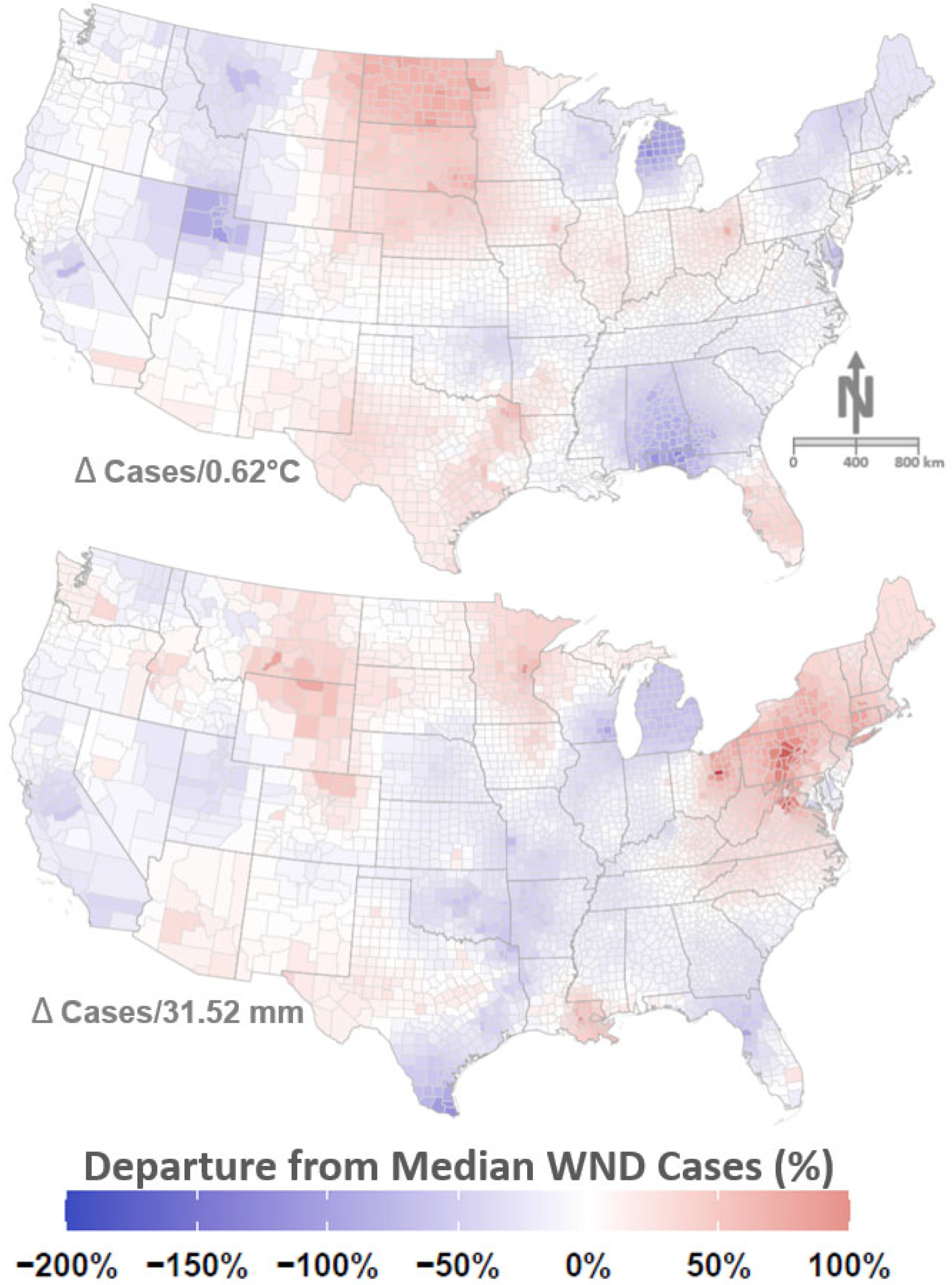
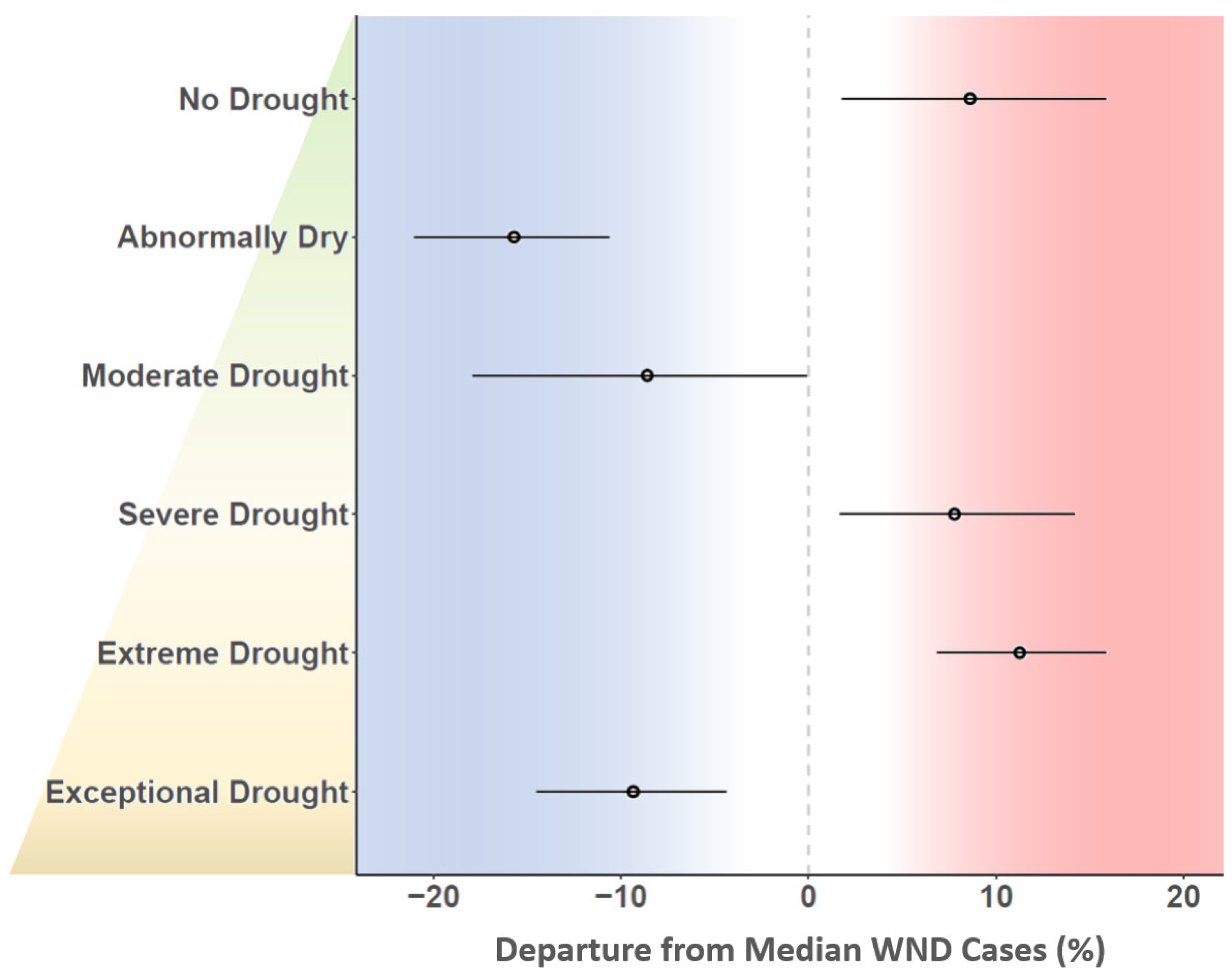
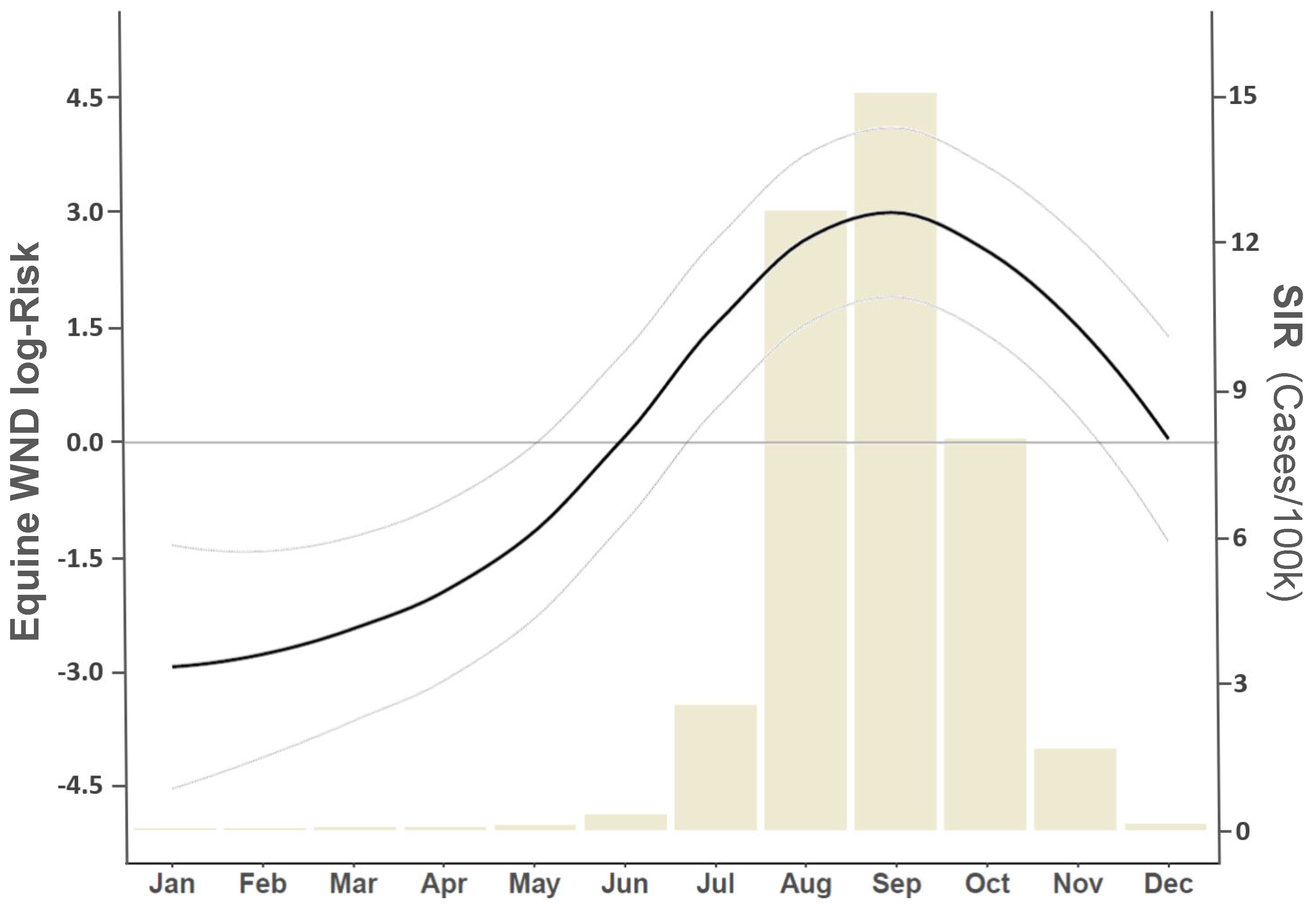
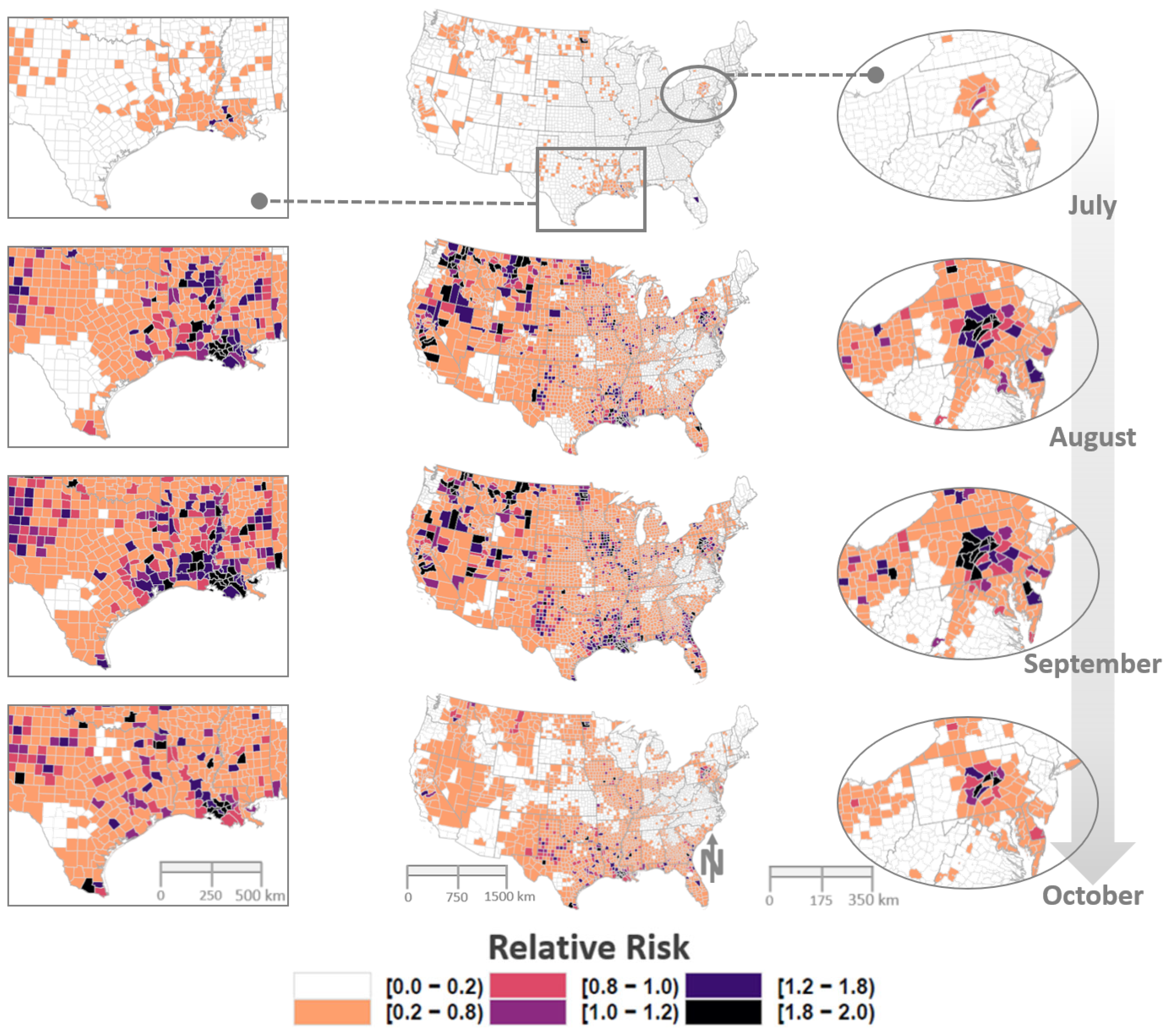
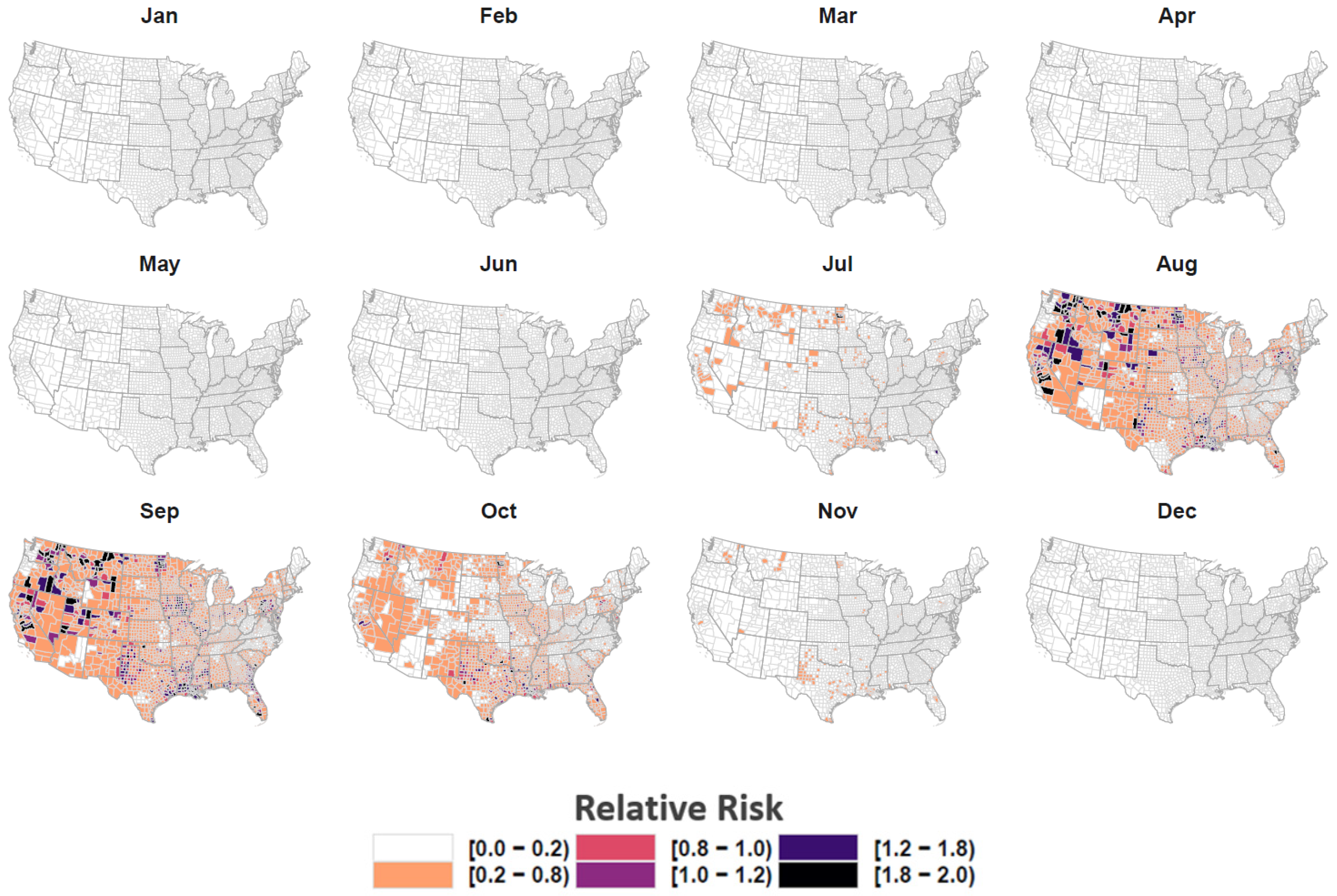
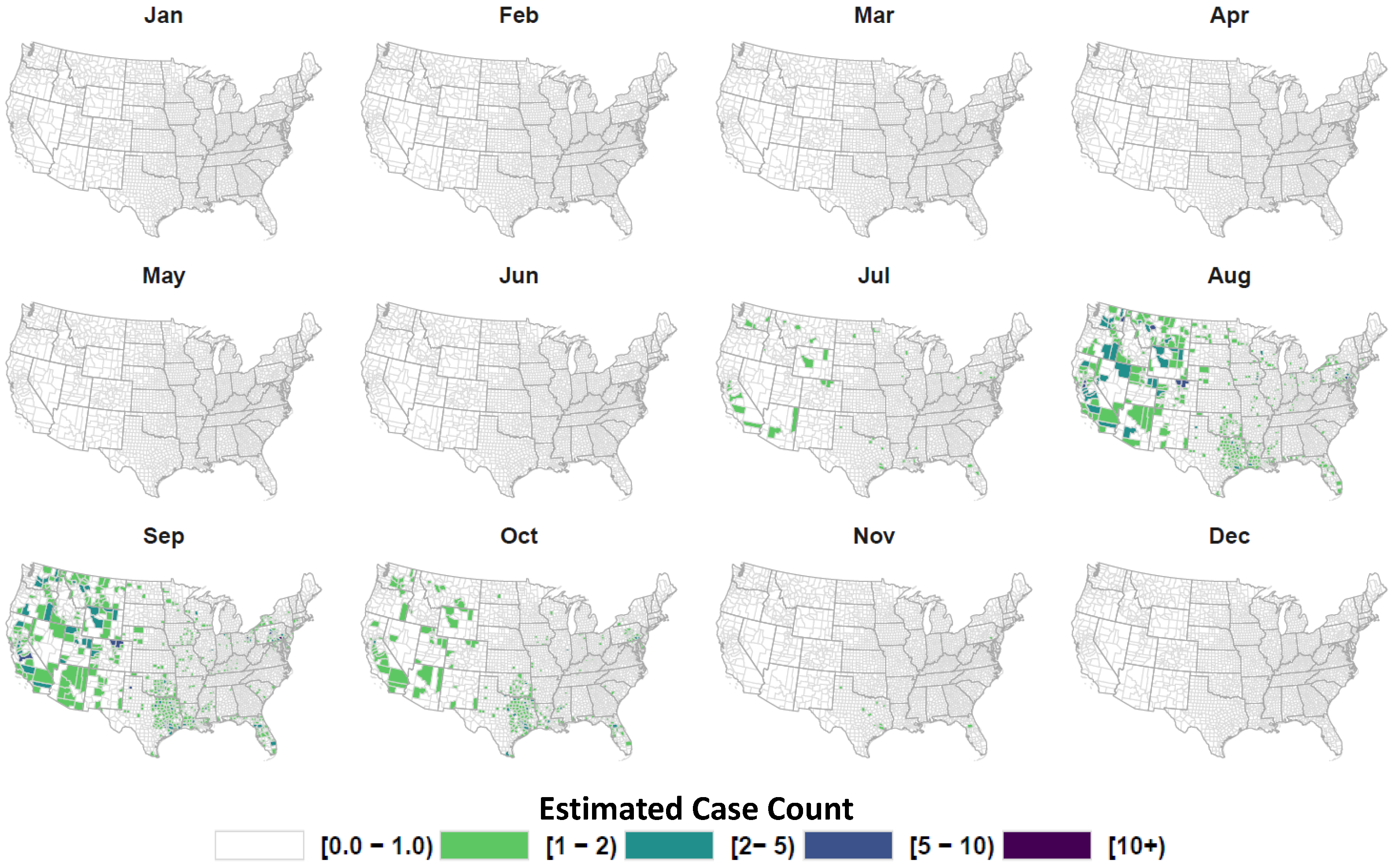
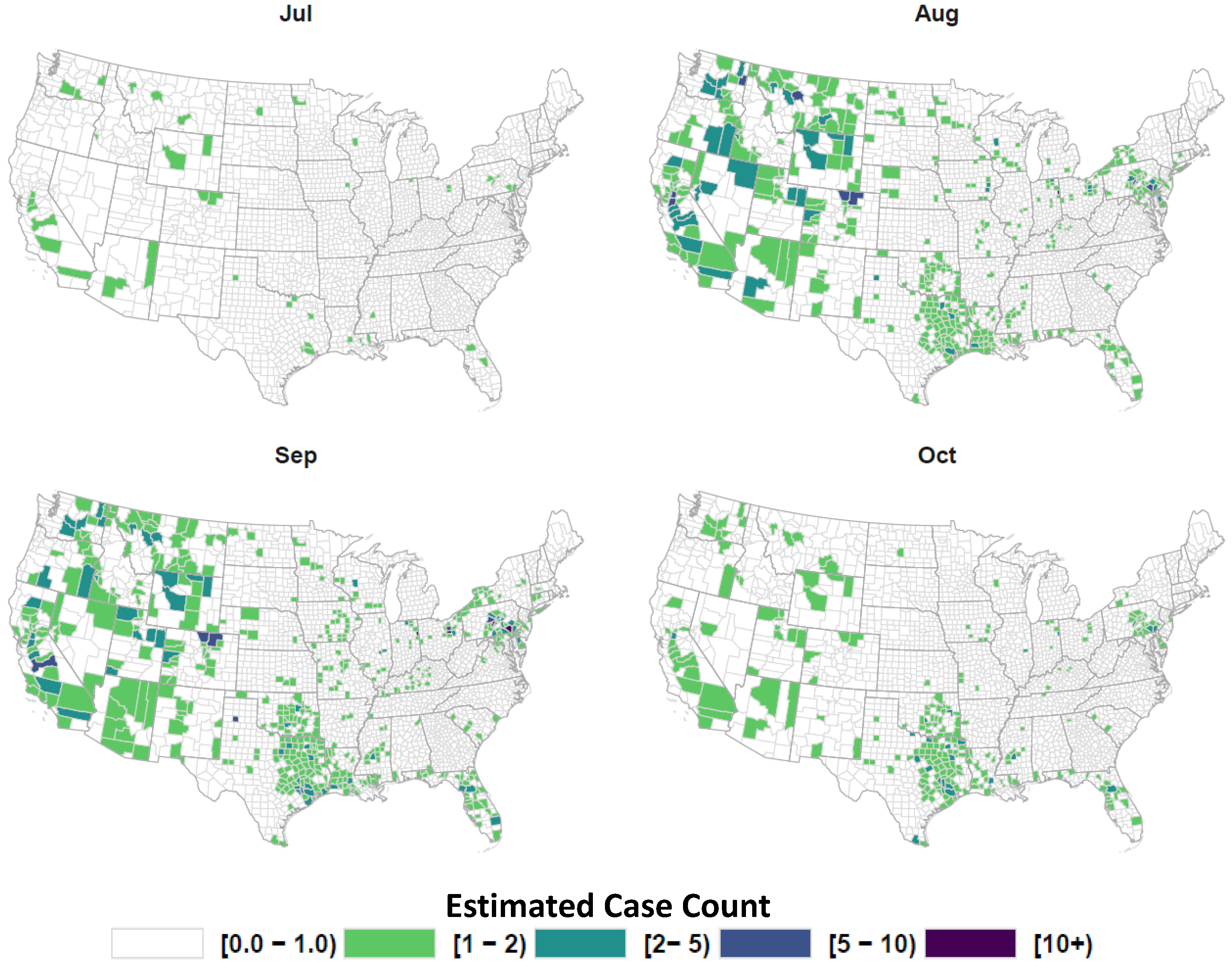
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CDC | Centers for Disease Control and Prevention |
| RR | Relative Risk |
| SIR | Standardized Incidence Rate |
| SVC | Spatially varying coefficients |
| US | United States of America |
| WNV | West Nile virus |
| WND | West Nile disease |
Appendix A. Tables and Figures


















References
- Strausbaugh, L.J.; Marfin, A.A.; Gubler, D.J. West Nile Encephalitis: An Emerging Disease in the United States. Clin. Infect. Dis. 2001, 33, 1713–1719. [Google Scholar] [CrossRef] [Green Version]
- Rochlin, I.; Faraji, A.; Healy, K.; Andreadis, T.G. West Nile Virus Mosquito Vectors in North America. J. Med. Entomol. 2019, 56, 1475–1490. [Google Scholar] [CrossRef] [PubMed]
- Byas, A.D.; Ebel, G.D. Comparative Pathology of West Nile Virus in Humans and Non-Human Animals. Pathogens 2020, 9, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Hurk, A.F.; Ritchie, S.A.; Mackenzie, J.S. Ecology and geographical expansion of Japanese encephalitis virus. Annu. Rev. Entomol. 2009, 54, 17–35. [Google Scholar] [CrossRef] [Green Version]
- Schuh, A.J.; Ward, M.J.; Leigh Brown, A.J.; Barrett, A.D.T. Dynamics of the emergence and establishment of a newly dominant genotype of Japanese encephalitis virus throughout Asia. J. Virol. 2014, 88, 4522–4532. [Google Scholar] [CrossRef] [Green Version]
- Clé, M.; Beck, C.; Salinas, S.; Lecollinet, S.; Gutierrez, S.; Van de Perre, P.; Baldet, T.; Foulongne, V.; Simonin, Y. Usutu virus: A new threat? Epidemiol. Infect. 2019, 147, e232. [Google Scholar] [CrossRef] [Green Version]
- Mackenzie, J.S.; Lindsay, M.D.A.; Smith, D.W.; Imrie, A. The ecology and epidemiology of Ross River and Murray Valley encephalitis viruses in Western Australia: Examples of One Health in Action. Trans. R. Soc. Trop. Med. Hyg. 2017, 111, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Curren, E.J.; Lehman, J.; Kolsin, J.; Walker, W.L.; Martin, S.W.; Staples, J.E.; Hills, S.L.; Gould, C.V.; Rabe, I.B.; Fischer, M.; et al. West Nile Virus and Other Nationally Notifiable Arboviral Diseases—United States, 2017. MMWR. Morb. Mortal. Wkly. Rep. 2018, 67, 1137–1142. [Google Scholar] [CrossRef] [PubMed]
- Humphreys, J.M.; Young, K.I.; Cohnstaedt, L.W.; Hanley, K.A.; Peters, D.P.C. Vector Surveillance, Host Species Richness, and Demographic Factors as West Nile Disease Risk Indicators. Viruses 2021, 13, 934. [Google Scholar] [CrossRef]
- Bosco-Lauth, A.M.; Bowen, R.A. West Nile Virus: Veterinary Health and Vaccine Development. J. Med. Entomol. 2019, 56, 1463–1466. [Google Scholar] [CrossRef]
- Saiz, J.C. Animal and Human Vaccines against West Nile Virus. Pathogens 2020, 9, 1073. [Google Scholar] [CrossRef]
- Mongoh, N.; Hearne, R.; Dyer, N.; Khaitsa, M. The economic impact of West Nile virus infection in horses in the North Dakota equine industry in 2002. Trop. Anim. Health Prod. 2008, 40, 69–76. [Google Scholar] [CrossRef]
- Lonker, N.S.; Fechner, K.; Abd El Wahed, A. Horses as a Crucial Part of One Health. Vet. Sci. 2020, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Ronca, S.E.; Ruff, J.C.; Murray, K.O. A 20-year historical review of West Nile virus since its initial emergence in North America: Has West Nile virus become a neglected tropical disease? PLoS Negl. Trop. Dis. 2021, 15, e0009190. [Google Scholar] [CrossRef] [PubMed]
- APHIS. USDA, Animal and Plant Health Inspection Service (APHIS), West Nile Virus. 2019. Available online: https://www.aphis.usda.gov/aphis/ourfocus/animalhealth/animal-disease-information/equine/wnv/west-nile-virus (accessed on 2 July 2021).
- Cantile, C.; Piero, F.D.; Guardo, G.D.; Arispici, M. Pathologic and Immunohistochemical Findings in Naturally Occurring West Nile Virus Infection in Horses. Vet. Pathol. 2001, 38, 414–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostlund, E.; Crom, R.; Pedersen, D.; Johnson, D.; Williams, W.; Schmitt, B. Equine West Nile encephalitis, United States. Emerg. Infect. Dis. 2001, 7, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Minke, J.; Siger, L.; Cupillard, L.; Powers, B.; Bakonyi, T.; Boyum, S.; Nowotny, N.; Bowen, R. Protection provided by a recombinant ALVAC®-WNV vaccine expressing the prM/E genes of a lineage 1 strain of WNV against a virulent challenge with a lineage 2 strain. Vaccine 2011, 29, 4608–4612. [Google Scholar] [CrossRef] [PubMed]
- Angenvoort, J.; Brault, A.; Bowen, R.; Groschup, M. West Nile viral infection of equids. Vet. Microbiol. 2013, 167, 168–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schvartz, G.; Farnoushi, Y.; Berkowitz, A.; Edery, N.; Hahn, S.; Steinman, A.; Lublin, A.; Erster, O. Molecular characterization of the re-emerging West Nile virus in avian species and equids in Israel, 2018, and pathological description of the disease. Parasites Vectors 2020, 13, 528. [Google Scholar] [CrossRef] [PubMed]
- Barros, S.C.; Ramos, F.; Fagulha, T.; Duarte, M.; Henriques, A.M.; Waap, H.; Luís, T.; Costa, T.; Amador, R.; Quintans, S.; et al. West Nile virus in horses during the summer and autumn seasons of 2015 and 2016, Portugal. Vet. Microbiol. 2017, 212, 75–79. [Google Scholar] [CrossRef]
- Bertram, F.M.; Thompson, P.N.; Venter, M. Epidemiology and Clinical Presentation of West Nile Virus Infection in Horses in South Africa, 2016–2017. Pathogens 2021, 10, 20. [Google Scholar] [CrossRef] [PubMed]
- Smithburn, K.C.; Hughes, T.P.; Burke, A.W.; Paul, J.H. A Neurotropic Virus Isolated from the Blood of a Native of Uganda. Am. J. Trop. Med. 1940, s1-20, 471–492. [Google Scholar] [CrossRef]
- Castillo-Olovares, J.; Wood, J. West Nile virus infection of horses. Vet. Res. 2004, 35, 467–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, J.R.; Mansoury, H.K.E. Natural and Experimental Infection of Egyptian Equines with West Nile Virus. Ann. Trop. Med. Parasitol. 1963, 57, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Blitvich, B.J.; Bowen, R.A.; Marlenee, N.L.; Hall, R.A.; Bunning, M.L.; Beaty, B.J. Epitope-Blocking Enzyme-Linked Immunosorbent Assays for Detection of West Nile Virus Antibodies in Domestic Mammals. J. Clin. Microbiol. 2003, 41, 2676–2679. [Google Scholar] [CrossRef] [Green Version]
- Martín-Acebes, M.A.; Saiz, J.C. West Nile virus: A re-emerging pathogen revisited. World J. Virol. 2012, 1, 51–70. [Google Scholar] [CrossRef]
- Silva, A.S.G.; Matos, A.C.D.; da Cunha, M.A.C.R.; Rehfeld, I.S.; Galinari, G.C.F.; Marcelino, S.A.C.; Saraiva, L.H.G.; Martins, N.R.d.S.; Maranhão, R.d.P.A.; Lobato, Z.I.P.; et al. West Nile virus associated with equid encephalitis in Brazil, 2018. Transbound. Emerg. Dis. 2019, 66, 445–453. [Google Scholar] [CrossRef] [Green Version]
- Pierson, T.C.; Diamond, M.S. The continued threat of emerging flaviviruses. Nat. Microbiol. 2020, 5, 796–812. [Google Scholar] [CrossRef]
- Cantile, C.; Di Guardo, G.; Eleni, C.; Arispici, M. Clinical and neuropathological features of West Nile virus equine encephalomyelitis in Italy. Equine Vet. J. 2000, 32, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Murgue, B.; Murri, S.; Zientara, S.; Durand, B.; Durand, J.; Zeller, H. West Nile outbreak in horses in southern France, 2000: The return after 35 years. Emerg. Infect. Dis. 2001, 7, 692–696. [Google Scholar] [CrossRef]
- Trock, S.; Meade, B.; Glaser, A.; Ostlund, E.; Lanciotti, R.; Cropp, B.; Kulasekera, V.; Kramer, L.; Komar, N. West Nile virus outbreak among horses in New York State, 1999 and 2000. Emerg. Infect. Dis. 2001, 7, 745–747. [Google Scholar] [CrossRef]
- Petersen, L.; Roehrig, J. West Nile virus: A reemerging global pathogen. Emerg. Infect. Dis. 2001, 7, 611–614. [Google Scholar] [CrossRef] [PubMed]
- Komar, N.; Clark, G.G. West Nile virus activity in Latin America and the Caribbean. Rev. Panam. Salud Publica = Pan Am. J. Public Health 2006, 19, 112–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, L.D.; Ciota, A.T.; Marm Kilpatrick, A. Introduction, Spread, and Establishment of West Nile Virus in the Americas. J. Med. Entomol. 2019, 56, 1448–1455. [Google Scholar] [CrossRef]
- Bunning, M.L.; Bowen, R.A.; Cropp, B.; Sullivan, K.; Davis, B.; Komar, N.; Godset, M.; Baker, D.; Hettler, D.; Holmes, D.; et al. Experimental Infection of Horses with West Nile Virus and Their Potential to Infect Mosquitoes and Serve as Amplifying Hosts. Ann. N. Y. Acad. Sci. 2001, 951, 338–339. [Google Scholar] [CrossRef]
- Peterson, A.T. Biogeography of diseases: A framework for analysis. Die Naturwissenschaften 2008, 95, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Escobar, L.E.; Craft, M.E. Advances and Limitations of Disease Biogeography Using Ecological Niche Modeling. Front. Microbiol. 2016, 7, 1174. [Google Scholar] [CrossRef] [Green Version]
- Lawson, A.B. Bayesian Disease Mapping: Hierarchical Modeling in Spatial Epidemiology, 3rd ed.; Chapman and Hall/CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Peterson, A.T. Predicting species’ geographic distributions based on ecological niche modeling. Condor 2001, 103, 599–605. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J. Species distribution modeling and ecological niche modeling: Getting the Concepts Right. Nat. Conserv. 2012, 10, 102–107. [Google Scholar] [CrossRef]
- Stenkamp-Strahm, C.; Patyk, K.; McCool-Eye, M.J.; Fox, A.; Humphreys, J.; James, A.; South, D.; Magzamen, S. Using geospatial methods to measure the risk of environmental persistence of avian influenza virus in South Carolina. Spat. Spatio-Temporal Epidemiol. 2020, 34, 100342. [Google Scholar] [CrossRef]
- Humphreys, J.M.; Ramey, A.M.; Douglas, D.C.; Mullinax, J.M.; Soos, C.; Link, P.; Walther, P.; Prosser, D.J. Waterfowl occurrence and residence time as indicators of H5 and H7 avian influenza in North American Poultry. Sci. Rep. 2020, 10, 2592. [Google Scholar] [CrossRef] [PubMed]
- Humphreys, J.M.; Douglas, D.C.; Ramey, A.M.; Mullinax, J.M.; Soos, C.; Link, P.; Walther, P.; Prosser, D.J. The spatial-temporal relationship of blue-winged teal to domestic poultry: Movement state modeling of a highly mobile avian influenza host. J. Appl. Ecol. 2021. [Google Scholar] [CrossRef]
- Swetnam, D.; Widen, S.G.; Wood, T.G.; Reyna, M.; Wilkerson, L.; Debboun, M.; Symonds, D.A.; Mead, D.G.; Beaty, B.J.; Guzman, H.; et al. Terrestrial Bird Migration and West Nile Virus Circulation, United States. Emerg. Infect. Dis. 2018, 24, 2184–2194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sales, L.P.; Hayward, M.W.; Loyola, R. What do you mean by “niche”? Modern ecological theories are not coherent on rhetoric about the niche concept. Acta Oecologica 2021, 110, 103701. [Google Scholar] [CrossRef]
- Soberón, J.; Peterson, A.T. Interpretation of Models of Fundamental Ecological Niches and Species’ Distributional Areas. Biodivers. Infor. 2005, 2, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Komar, N. West Nile viral encephalitis. Rev. Sci. Tech. (Int. Off. Epizoot.) 2000, 19, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Rappole, J.H.; Derrickson, S.R.; Hubálek, Z. Migratory birds and spread of West Nile virus in the Western Hemisphere. Emerg. Infect. Dis. 2000, 6, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Salas, I.; Contreras-Cordero, J.F.; Blitvich, B.J.; González-Rojas, J.I.; Cavazos-Alvarez, A.; Marlenee, N.L.; Elizondo-Quiroga, A.; Loroño-Pino, M.A.; Gubler, D.J.; Cropp, B.C.; et al. Serologic evidence of West Nile Virus infection in birds, Tamaulipas State, México. Vector Borne Zoonotic Dis. 2003, 3, 209–213. [Google Scholar] [CrossRef] [Green Version]
- Komar, N.; Langevin, S.; Hinten, S.; Nemeth, N.; Edwards, E.; Hettler, D.; Davis, B.; Bowen, R.; Bunning, M. Experimental infection of North American birds with the New York 1999 strain of West Nile virus. Emerg. Infect. Dis. 2003, 9, 311–322. [Google Scholar] [CrossRef]
- Shelite, T.R.; Rogers, C.M.; Litzner, B.R.; Johnson, R.R.; Schneegurt, M.A. West Nile Virus Antibodies in Permanent Resident and Overwintering Migrant Birds in South-Central Kansas. Vector-Borne Zoonotic Dis. 2008, 8, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Durand, B.; Tran, A.; Balança, G.; Chevalier, V. Geographic variations of the bird-borne structural risk of West Nile virus circulation in Europe. PLoS ONE 2017, 12, e0185962. [Google Scholar] [CrossRef] [Green Version]
- Tolsá, M.J.; García-Peña, G.E.; Rico-Chávez, O.; Roche, B.; Suzán, G. Macroecology of birds potentially susceptible to West Nile virus. Proc. R. Soc. B Biol. Sci. 2018, 285, 20182178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzán, G.; García-Peña, G.E.; Castro-Arellano, I.; Rico, O.; Rubio, A.V.; Tolsá, M.J.; Roche, B.; Hosseini, P.R.; Rizzoli, A.; Murray, K.A.; et al. Metacommunity and phylogenetic structure determine wildlife and zoonotic infectious disease patterns in time and space. Ecol. Evol. 2015, 5, 865–873. [Google Scholar] [CrossRef]
- Huang, Z.Y.X.; de Boer, W.F.; van Langevelde, F.; Olson, V.; Blackburn, T.M.; Prins, H.H.T. Species’ Life-History Traits Explain Interspecific Variation in Reservoir Competence: A Possible Mechanism Underlying the Dilution Effect. PLoS ONE 2013, 8, e54341. [Google Scholar] [CrossRef] [Green Version]
- Webb, C.O.; Ackerly, D.D.; McPeek, M.A.; Donoghue, M.J. Phylogenies and Community Ecology. Annu. Rev. Ecol. Syst. 2002, 33, 475–505. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, G.S.; Parker, I.M. The Evolutionary Ecology of Plant Disease: A Phylogenetic Perspective. Annu. Rev. Phytopathol. 2016, 54, 549–578. [Google Scholar] [CrossRef]
- Fountain-Jones, N.M.; Pearse, W.D.; Escobar, L.E.; Alba-Casals, A.; Carver, S.; Davies, T.J.; Kraberger, S.; Papeş, M.; Vandegrift, K.; Worsley-Tonks, K.; et al. Towards an eco-phylogenetic framework for infectious disease ecology. Biol. Rev. 2018, 93, 950–970. [Google Scholar] [CrossRef] [Green Version]
- Swaddle, J.P.; Calos, S.E. Increased avian diversity is associated with lower incidence of human West Nile infection: Observation of the dilution effect. PLoS ONE 2008, 3, e2488. [Google Scholar] [CrossRef] [Green Version]
- Loss, S.R.; Hamer, G.L.; Walker, E.D.; Ruiz, M.O.; Goldberg, T.L.; Kitron, U.D.; Brawn, J.D. Avian host community structure and prevalence of West Nile virus in Chicago, Illinois. Oecologia 2009, 159, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Kain, M.P.; Bolker, B.M. Predicting West Nile virus transmission in North American bird communities using phylogenetic mixed effects models and eBird citizen science data. Parasites Vectors 2019, 12, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Levine, R.S.; Hedeen, D.L.; Hedeen, M.W.; Hamer, G.L.; Mead, D.G.; Kitron, U.D. Avian species diversity and transmission of West Nile virus in Atlanta, Georgia. Parasites Vectors 2017, 10, 62. [Google Scholar] [CrossRef] [Green Version]
- Luis, A.D.; Kuenzi, A.J.; Mills, J.N. Species diversity concurrently dilutes and amplifies transmission in a zoonotic host–pathogen system through competing mechanisms. Proc. Natl. Acad. Sci. USA 2018, 115, 7979–7984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centers for Disease Control and Prevention (CDC). ArboNET. 2019. Available online: https://wwwn.cdc.gov/arbonet (accessed on 21 August 2019).
- Breslow, N.; Leroux, B.; Platt, R. Approximate hierarchical modelling of discrete data in epidemiology. Stat. Methods Med. Res. 1998, 7, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Dean, C.B.; Ugarte, M.D.; Militino, A.F. Detecting interaction between random region and fixed age effects in disease mapping. Biometrics 2001, 57, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Simpson, D.; Illian, J.; Lindgren, F.; Sørbye, S.H.; Rue, H. Going off grid: Computationally efficient inference for log-Gaussian Cox processes. Biometrika 2016, 103, 49–70. [Google Scholar] [CrossRef] [Green Version]
- Riebler, A.; Sørbye, S.H.; Simpson, D.; Rue, H.; Lawson, A.B.; Lee, D.; MacNab, Y. An intuitive Bayesian spatial model for disease mapping that accounts for scaling. Stat. Methods Med. Res. 2016, 25, 1145–1165. [Google Scholar] [CrossRef] [Green Version]
- Bivand, R.; Piras, G. Comparing Implementations of Estimation Methods for Spatial Econometrics. J. Stat. Softw. 2015, 63. [Google Scholar] [CrossRef] [Green Version]
- PRISM. Total Annual Precipitation and Mean Maximum Temperature Climate Data. 2020. Available online: http://prism.oregonstate.edu (accessed on 12 July 2020).
- Hart, E.M.; Bell, K. prism: Download Data from the Oregon Prism Project; R Package Version 0.0.6; 2015. [Google Scholar] [CrossRef]
- Sullivan, B.L.; Aycrigg, J.L.; Barry, J.H.; Bonney, R.E.; Bruns, N.; Cooper, C.B.; Damoulas, T.; Dhondt, A.A.; Dietterich, T.; Farnsworth, A.; et al. The eBird enterprise: An integrated approach to development and application of citizen science. Biol. Conserv. 2014, 169, 31–40. [Google Scholar] [CrossRef]
- Jetz, W.; Thomas, G.H.; Joy, J.B.; Hartmann, K.; Mooers, A.O. The global diversity of birds in space and time. Nature 2012, 491, 444–448. [Google Scholar] [CrossRef]
- Strimas-Mackey, M.; Miller, E.; Hochachka, W. auk: EBird Data Extraction and Processing with AWK; R Package Version 0.3.0; 2018. [Google Scholar]
- Hartley, D.M.; Barker, C.M.; Le Menach, A.; Niu, T.; Gaff, H.D.; Reisen, W.K. Effects of Temperature on Emergence and Seasonality of West Nile Virus in California. Am. J. Trop. Med. Hyg. 2012, 86, 884. [Google Scholar] [CrossRef] [Green Version]
- Ukawuba, I.; Shaman, J. Association of spring-summer hydrology and meteorology with human West Nile virus infection in West Texas, USA, 2002–2016. Parasites Vectors 2018, 11, 224. [Google Scholar] [CrossRef]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A Software Platform for Bayesian Evolutionary Analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef] [Green Version]
- Kembel, S.; Cowan, P.; Helmus, M.; Cornwell, W.; Morlon, H.; Ackerly, D.; Blomberg, S.; Webb, C. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [Green Version]
- Cadotte, M.W.; Davies, T.J. Phylogenies in Ecology: A Guide to Concepts and Methods; Princeton University Press: Princeton, NJ, USA, 2016. [Google Scholar]
- Redding, D.W.; Mooers, A.Ø. Incorporating Evolutionary Measures into Conservation Prioritization. Conserv. Biol. 2006, 20, 1670–1678. [Google Scholar] [CrossRef]
- Tuanmu, M.N.; Jetz, W. A global 1-km consensus land-cover product for biodiversity and ecosystem modelling. Glob. Ecol. Biogeogr. 2014, 23, 1031–1045. [Google Scholar] [CrossRef]
- Robinson, N.; Regetz, J.; Guralnick, R.P. EarthEnv-DEM90: A nearly-global, void-free, multi-scale smoothed, 90m digital elevation model from fused ASTER and SRTM data. ISPRS J. Photogramm. Remote Sens. 2014, 87, 57–67. [Google Scholar] [CrossRef]
- Census Bureau. Small Area Income and Poverty Estimates (SAIPE) Program. 2018. Available online: https://www.census.gov/programs-surveys/saipe.html (accessed on 15 July 2019).
- Recht, H. censusapi: Retrieve Data from the Census APIs; R Package Version 0.7.1; 2020. [Google Scholar]
- Belsley, D.A.; Kuh, E.; Welsch, R.E. Identifying Influential Data and Sources of Collinearity; John Wiley & Sons: Hoboken, NJ, USA, 1980. [Google Scholar]
- Hendricks, J.; Pelzer, B. Collinearity involving ordered and unordered categorical variables. In RC33 Conference in Amsterdam; 2004; pp. 1–17. Available online: https://www.researchgate.net/publication/235994590_Collinearity_involving_ordered_and_unordered_categorical_variables (accessed on 2 July 2021).
- Elliott, P.; Wakefield, J.; Best, N.; Briggs, D. Spatial Epidemiology: Methods and Applications; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Rue, H.; Martino, S.; Chopin, N. Approximate Bayesian inference for latent Gaussian models by using integrated nested Laplace approximations. J. R. Stat. Soc. Ser. B Stat. Methodol. 2009, 71, 319–392. [Google Scholar] [CrossRef]
- Martins, T.G.; Simpson, D.; Lindgren, F.; Rue, H. Bayesian computing with INLA: New features. Comput. Stat. Data Anal. 2013, 67, 68–83. [Google Scholar] [CrossRef] [Green Version]
- Lindgren, F.; Rue, H. Bayesian Spatial Modelling with R-INLA. J. Stat. Softw. 2015, 63, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Simpson, D.; Rue, H.; Riebler, A.; Martins, T.G.; Sørbye, S.H. Penalising Model Component Complexity: A Principled, Practical Approach to Constructing Priors. Stat. Sci. 2017, 32, 1–28. [Google Scholar] [CrossRef]
- Hoeting, J.A.; Madigan, D.; Raftery, A.E.; Volinsky, C.T. Bayesian Model Averaging: A Tutorial. Stat. Sci. 1999, 14, 382–401. [Google Scholar]
- Gómez-Rubio, V.; Bivand, R.S.; Rue, H. Bayesian Model Averaging with the Integrated Nested Laplace Approximation. Econometrics 2020, 8, 23. [Google Scholar] [CrossRef]
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. InterJournal Complex Syst. 2006, 1695. [Google Scholar]
- Harris, R. Visualizing Multicollinearity Using Igraph. 2020. Available online: https://rpubs.com/profrichharris/visual-collinearity (accessed on 2 July 2021).
- Brier, G.W. Verification of forecasts expressed in terms of probability. Mon. Weather Rev. 1950, 78, 1–3. [Google Scholar] [CrossRef]
- Gneiting, T.; Raftery, A.E. Strictly Proper Scoring Rules, Prediction, and Estimation. J. Am. Stat. Assoc. 2007, 102, 359–378. [Google Scholar] [CrossRef]
- Richardson, W.J. Timing and Amount of Bird Migration in Relation to Weather: A Review. Oikos 1978, 30, 224–272. [Google Scholar] [CrossRef]
- Humphreys, J.M.; Murrow, J.L.; Sullivan, J.D.; Prosser, D.J. Seasonal occurrence and abundance of dabbling ducks across the continental United States: Joint spatio-temporal modelling for the Genus Anas. Divers. Distrib. 2019, 25, 1497–1508. [Google Scholar] [CrossRef] [Green Version]
- Nasci, R.; Savage, H.; White, D.; Miller, J.; Cropp, B.; Godsey, M.; Kerst, A.; Bennett, P.; Gottfried, K.; Lanciotti, R. West Nile virus in overwintering Culex mosquitoes, New York City, 2000. Emerg. Infect. Dis. 2001, 7, 742–744. [Google Scholar] [CrossRef]
- Davis, J.K.; Gebrehiwot, T.; Worku, M.; Awoke, W.; Mihretie, A.; Nekorchuk, D.; Wimberly, M.C. A genetic algorithm for identifying spatially-varying environmental drivers in a malaria time series model. Environ. Model. Softw. Environ. Data News 2019, 119, 275–284. [Google Scholar] [CrossRef]
- Rollinson, C.R.; Finley, A.O.; Alexander, M.R.; Banerjee, S.; Dixon Hamil, K.A.; Koenig, L.E.; Locke, D.H.; Peterson, M.; Tingley, M.W.; Wheeler, K.; et al. Working across space and time: Nonstationarity in ecological research and application. Front. Ecol. Environ. 2021, 19, 66–72. [Google Scholar] [CrossRef]
- Olson, S.H.; Gangnon, R.; Elguero, E.; Durieux, L.; Guégan, J.F.; Foley, J.A.; Patz, J.A. Links between climate, malaria, and wetlands in the Amazon Basin. Emerg. Infect. Dis. 2009, 15, 659–662. [Google Scholar] [CrossRef] [PubMed]
- Landesman, W.J.; Allan, B.F.; Langerhans, R.B.; Knight, T.M.; Chase, J.M. Inter-Annual Associations Between Precipitation and Human Incidence of West Nile Virus in the United States. Vector-Borne Zoonotic Dis. 2007, 7, 337–343. [Google Scholar] [CrossRef]
- Ruiz, M.O.; Chaves, L.F.; Hamer, G.L.; Sun, T.; Brown, W.M.; Walker, E.D.; Haramis, L.; Goldberg, T.L.; Kitron, U.D. Local impact of temperature and precipitation on West Nile virus infection in Culex species mosquitoes in northeast Illinois, USA. Parasites Vectors 2010, 3, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paull, S.H.; Kilpatrick, A.M.; Horton, D.E.; Diffenbaugh, N.S.; Ashfaq, M.; Rastogi, D.; Kramer, L.D. Drought and immunity determine the intensity of west nile virus epidemics and climate change impacts. Proc. R. Soc. B Biol. Sci. 2017, 284, 20162078. [Google Scholar] [CrossRef]
- Hahn, M.B.; Monaghan, A.J.; Hayden, M.H.; Eisen, R.J.; Delorey, M.J.; Lindsey, N.P.; Nasci, R.S.; Fischer, M. Meteorological conditions associated with increased incidence of west nile virus disease in the United States, 2004–2012. Am. J. Trop. Med. Hyg. 2015, 92, 1013. [Google Scholar] [CrossRef]
- Karl, T.R.; Koss, W.J. Regional and National Monthly, Seasonal, and Annual Temperature Weighted by Area, 1895–1983 (Historical Climatology Series 4-3); Technical report; National Climatic Data Center: Asheville, NC, USA, 1984. [Google Scholar]
- Schöning, J.M.; Cerny, N.; Prohaska, S.; Wittenbrink, M.M.; Smith, N.H.; Bloemberg, G.; Pewsner, M.; Schiller, I.; Origgi, F.C.; Ryser-Degiorgis, M.P. Surveillance of Bovine Tuberculosis and Risk Estimation of a Future Reservoir Formation in Wildlife in Switzerland and Liechtenstein. PLoS ONE 2013, 8, e54253. [Google Scholar] [CrossRef] [Green Version]
- Gallana, M.; Ryser-Degiorgis, M.P.; Wahli, T.; Segner, H. Climate change and infectious diseases of wildlife: Altered interactions between pathogens, vectors and hosts. Curr. Zool. 2013, 59, 427–437. [Google Scholar] [CrossRef]
- Shaman, J.; Day, J.F.; Stieglitz, M. Drought-induced amplification of Saint Louis encephalitis virus, Florida. Emerg. Infect. Dis. 2002, 8, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Shaman, J.; Day, J.F.; Stieglitz, M. Drought-Induced Amplification and Epidemic Transmission of West Nile Virus in Southern Florida. J. Med. Entomol. 2005, 42, 134–141. [Google Scholar] [CrossRef]
- Johnson, B.J.; Sukhdeo, M. Drought-Induced Amplification of Local and Regional West Nile Virus Infection Rates in New Jersey. J. Med. Entomol. 2013, 50, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Gaunt, M.W.; Sall, A.A.; Lamballerie, X.D.; Falconar, A.K.I.; Dzhivanian, T.I.; Gould, E.A. Phylogenetic relationships of flaviviruses correlate with their epidemiology, disease association and biogeography. J. Gen. Virol. 2001, 82, 1867–1876. [Google Scholar] [CrossRef] [PubMed]
- Davies, T.J.; Pedersen, A.B. Phylogeny and geography predict pathogen community similarity in wild primates and humans. Proc. R. Soc. B Biol. Sci. 2008, 275, 1695–1701. [Google Scholar] [CrossRef] [Green Version]
- Streicker, D.G.; Turmelle, A.S.; Vonhof, M.J.; Kuzmin, I.V.; McCracken, G.F.; Rupprecht, C.E. Host Phylogeny Constrains Cross-Species Emergence and Establishment of Rabies Virus in Bats. Science 2010, 329, 676–679. [Google Scholar] [CrossRef] [Green Version]
- Parker, I.M.; Saunders, M.; Bontrager, M.; Weitz, A.P.; Hendricks, R.; Magarey, R.; Suiter, K.; Gilbert, G.S. Phylogenetic structure and host abundance drive disease pressure in communities. Nature 2015, 520, 542–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiens, J.J.; Graham, C.H. Niche Conservatism: Integrating Evolution, Ecology, and Conservation Biology. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 519–539. [Google Scholar] [CrossRef] [Green Version]
- Dawson, W.R.; Marsh, R.L.; Yacoe, M.E. Metabolic adjustments of small passerine birds for migration and cold. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 1983, 245, R755–R767. [Google Scholar] [CrossRef] [PubMed]
- Tucker, C.M.; Cadotte, M.W.; Carvalho, S.B.; Jonathan Davies, T.; Ferrier, S.; Fritz, S.A.; Grenyer, R.; Helmus, M.R.; Jin, L.S.; Mooers, A.O.; et al. A guide to phylogenetic metrics for conservation, community ecology and macroecology. Biol. Rev. 2017, 92, 698–715. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
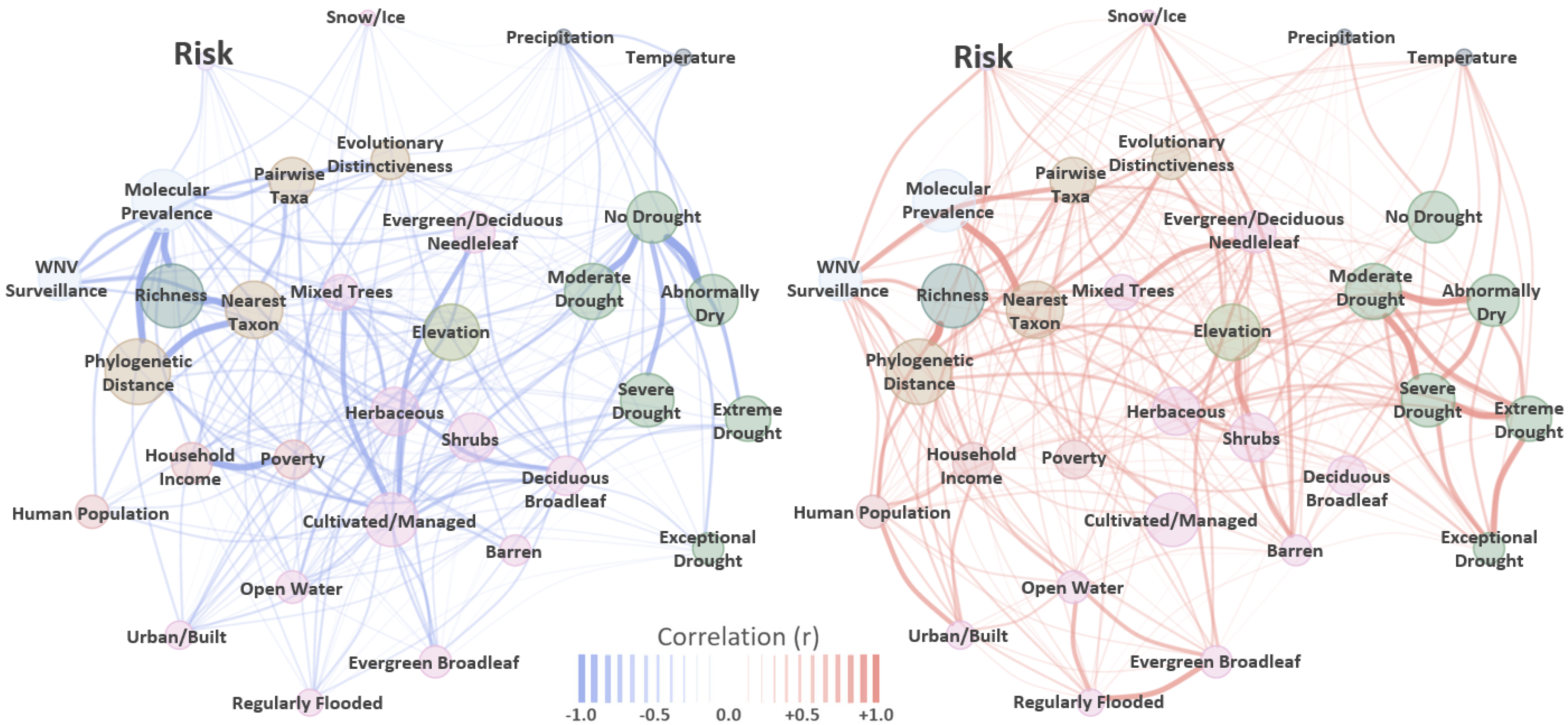
| Covariate | Mean | SD | 2.5 Q | 97.5 Q |
|---|---|---|---|---|
| WNV Surveillance | 0.14 | 0.03 | 0.09 | 0.20 |
| Human Population Density | −0.03 | 0.03 | −0.09 | 0.02 |
| Median Household Income | −0.12 | 0.04 | −0.19 | −0.05 |
| Population in Poverty (%) | −0.02 | 0.03 | −0.05 | 0.01 |
| Evergreen/Deciduous Needleleaf Trees | −0.09 | 0.05 | −0.20 | 0.01 |
| Evergreen Broadleaf Trees | −0.01 | 0.05 | −0.10 | 0.09 |
| Deciduous Broadleaf Trees | −0.16 | 0.10 | −0.35 | 0.04 |
| Mixed/Other Trees | −0.20 | 0.09 | −0.38 | −0.02 |
| Shrubs | 0.05 | 0.06 | −0.07 | 0.16 |
| Herbaceous Vegetation | 0.02 | 0.08 | −0.14 | 0.18 |
| Cultivated and Managed Vegetation | 0.02 | 0.10 | −0.19 | 0.22 |
| Regularly Flooded Vegetation | 0.02 | 0.05 | −0.08 | 0.11 |
| Urban/Built-Up | −0.12 | 0.05 | −0.21 | −0.03 |
| Snow/Ice | −0.02 | 0.02 | −0.07 | 0.02 |
| Barren | 0.07 | 0.03 | 0.01 | 0.13 |
| Open Water | −0.16 | 0.05 | −0.25 | −0.06 |
| Elevation | −0.11 | 0.08 | −0.26 | 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Humphreys, J.M.; Pelzel-McCluskey, A.M.; Cohnstaedt, L.W.; McGregor, B.L.; Hanley, K.A.; Hudson, A.R.; Young, K.I.; Peck, D.; Rodriguez, L.L.; Peters, D.P.C. Integrating Spatiotemporal Epidemiology, Eco-Phylogenetics, and Distributional Ecology to Assess West Nile Disease Risk in Horses. Viruses 2021, 13, 1811. https://doi.org/10.3390/v13091811
Humphreys JM, Pelzel-McCluskey AM, Cohnstaedt LW, McGregor BL, Hanley KA, Hudson AR, Young KI, Peck D, Rodriguez LL, Peters DPC. Integrating Spatiotemporal Epidemiology, Eco-Phylogenetics, and Distributional Ecology to Assess West Nile Disease Risk in Horses. Viruses. 2021; 13(9):1811. https://doi.org/10.3390/v13091811
Chicago/Turabian StyleHumphreys, John M., Angela M. Pelzel-McCluskey, Lee W. Cohnstaedt, Bethany L. McGregor, Kathryn A. Hanley, Amy R. Hudson, Katherine I. Young, Dannele Peck, Luis L. Rodriguez, and Debra P. C. Peters. 2021. "Integrating Spatiotemporal Epidemiology, Eco-Phylogenetics, and Distributional Ecology to Assess West Nile Disease Risk in Horses" Viruses 13, no. 9: 1811. https://doi.org/10.3390/v13091811