
The Metabolic Response of Various Cell Lines to Microtubule-Driven Uptake of Lipid- and Polymer-Coated Layer-by-Layer Microcarriers


Abstract
:
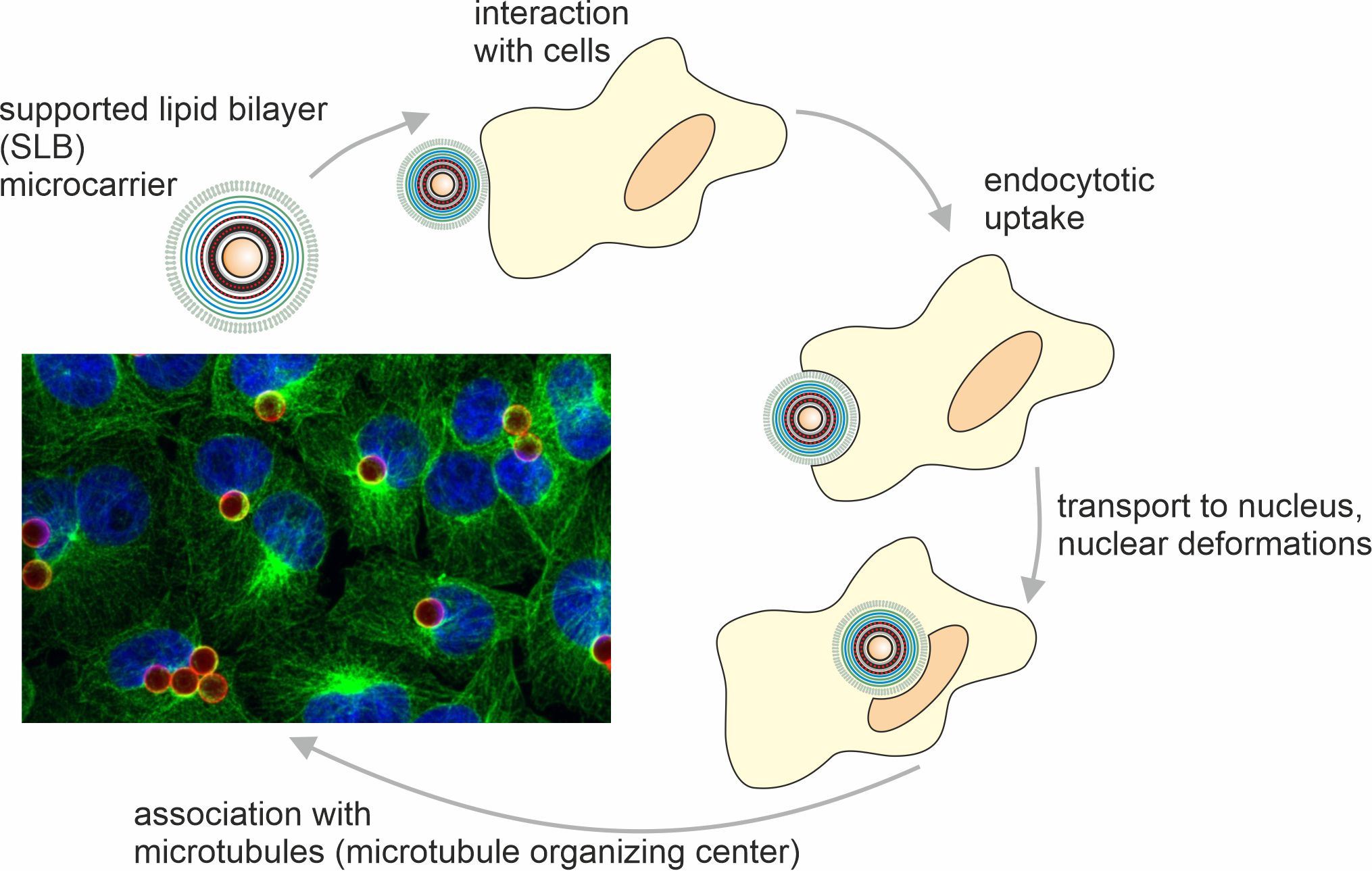{kind=link}
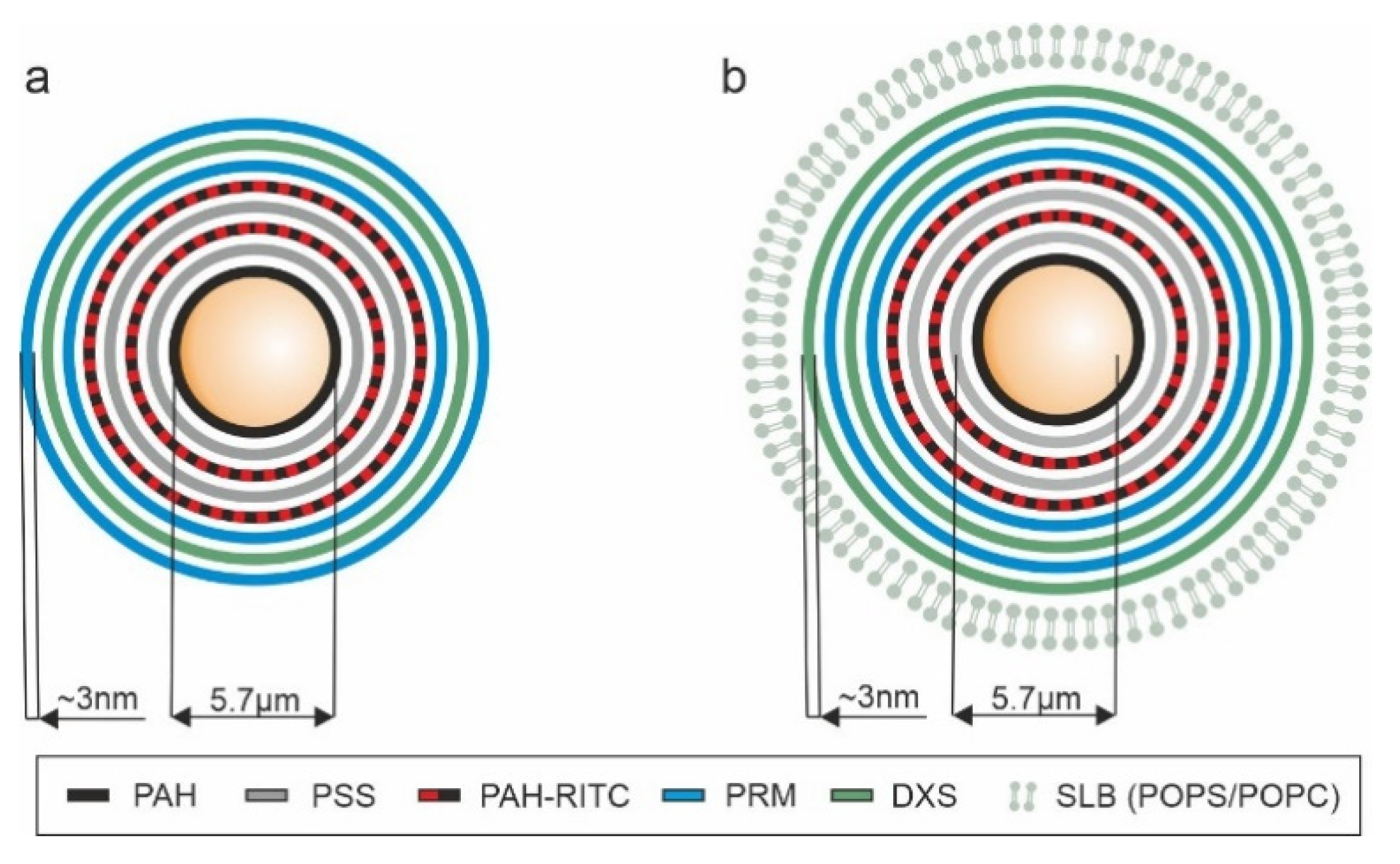{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cultivation of Permanent Cell Lines
2.3. Preparation of Human MΦ
2.4. Layer-by-Layer Coating of Microcarriers
2.5. Assembly of a Supported Lipid Bilayer (SLB)
2.6. Application of Microcarriers to Cells
2.7. Assessment of Mitochondrial Respiration and Glycolysis through Extracellular Flux Analysis
2.8. Immunofluorescence Analysis and Application of the Mito Tracker Dye
2.9. Confocal Microscopy (CLSM)
2.10. Statistical Analysis
3. Results and Discussion
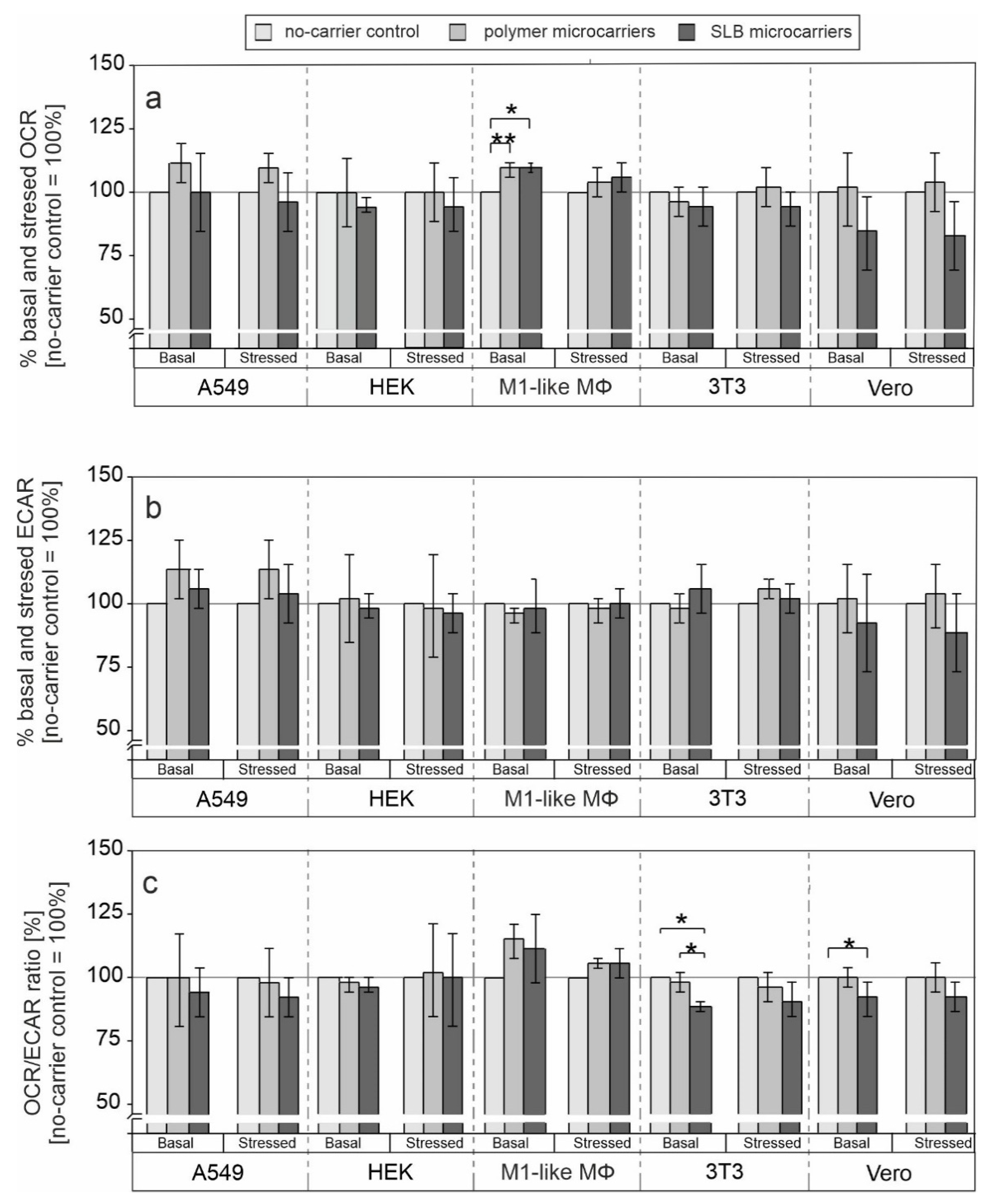
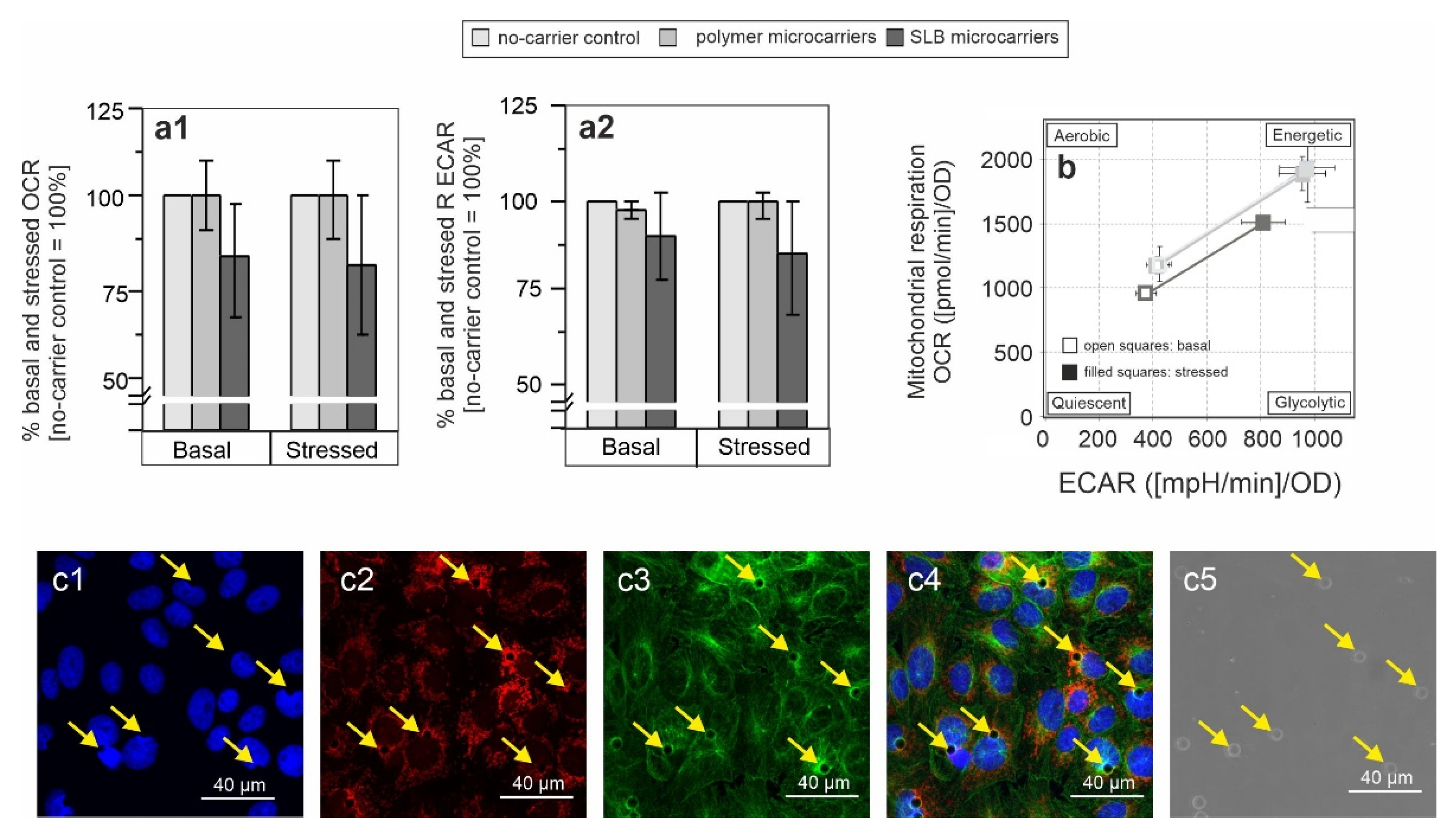
3.1. Influence of Microcarriers on Cellular Metabolic Potential
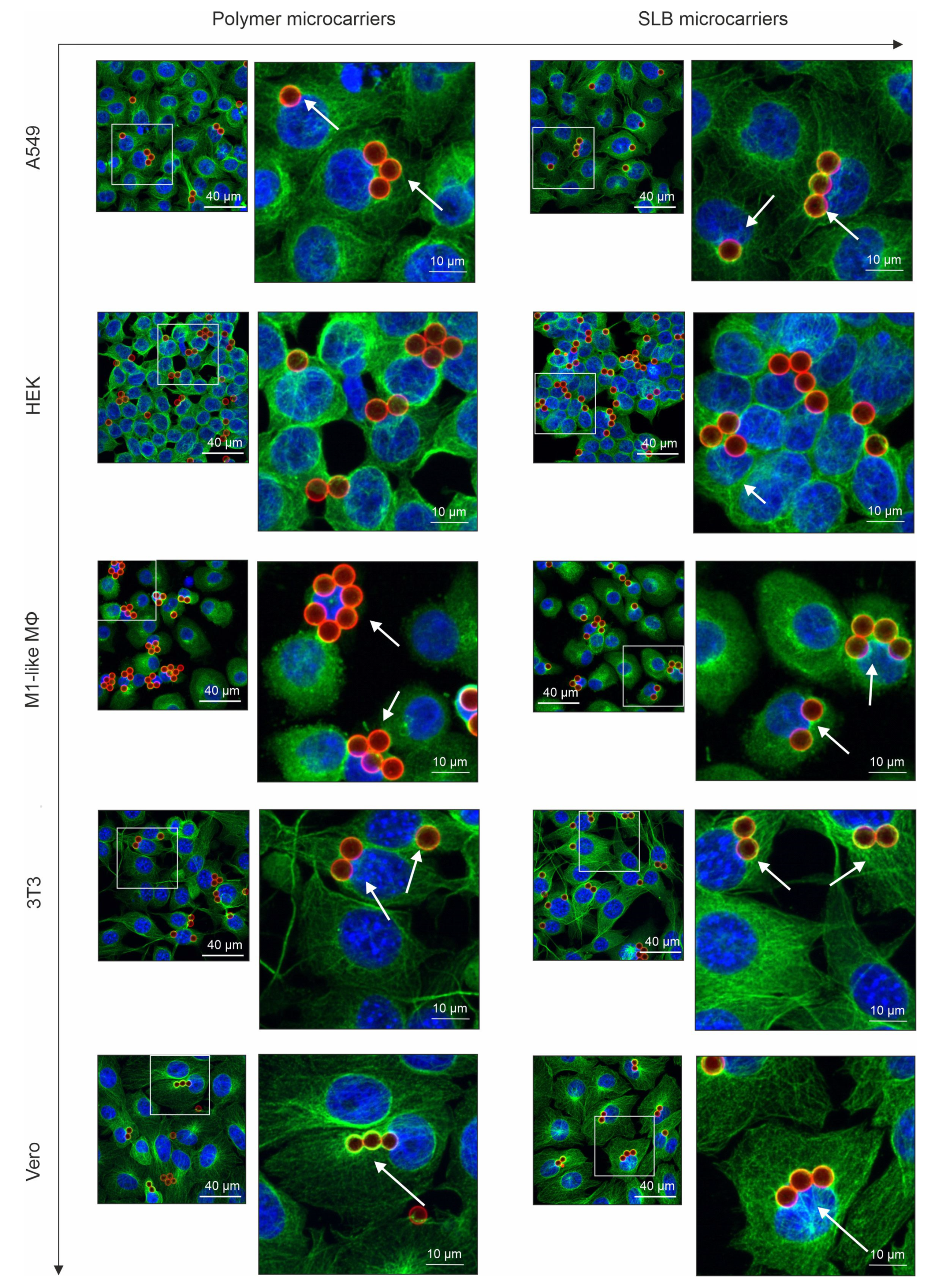
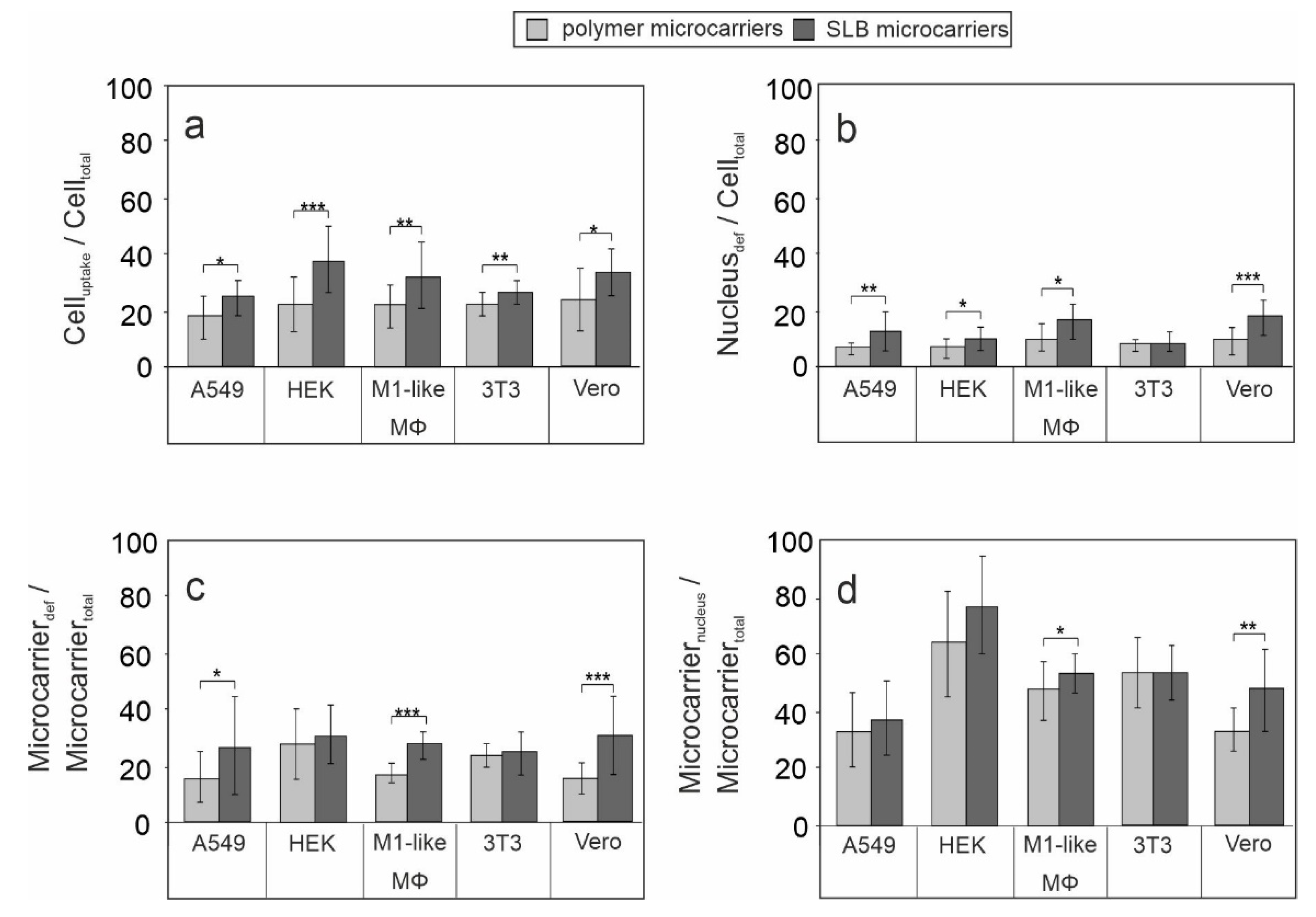
3.2. Influence of the Lipid Bilayer on Association of Microcarriers with Microtubule Filaments and Their Perinuclear Localization
3.3. Metabolic Alterations Coincide with Microcarrier Internalization and Relate to Mitochondrial Redistribution
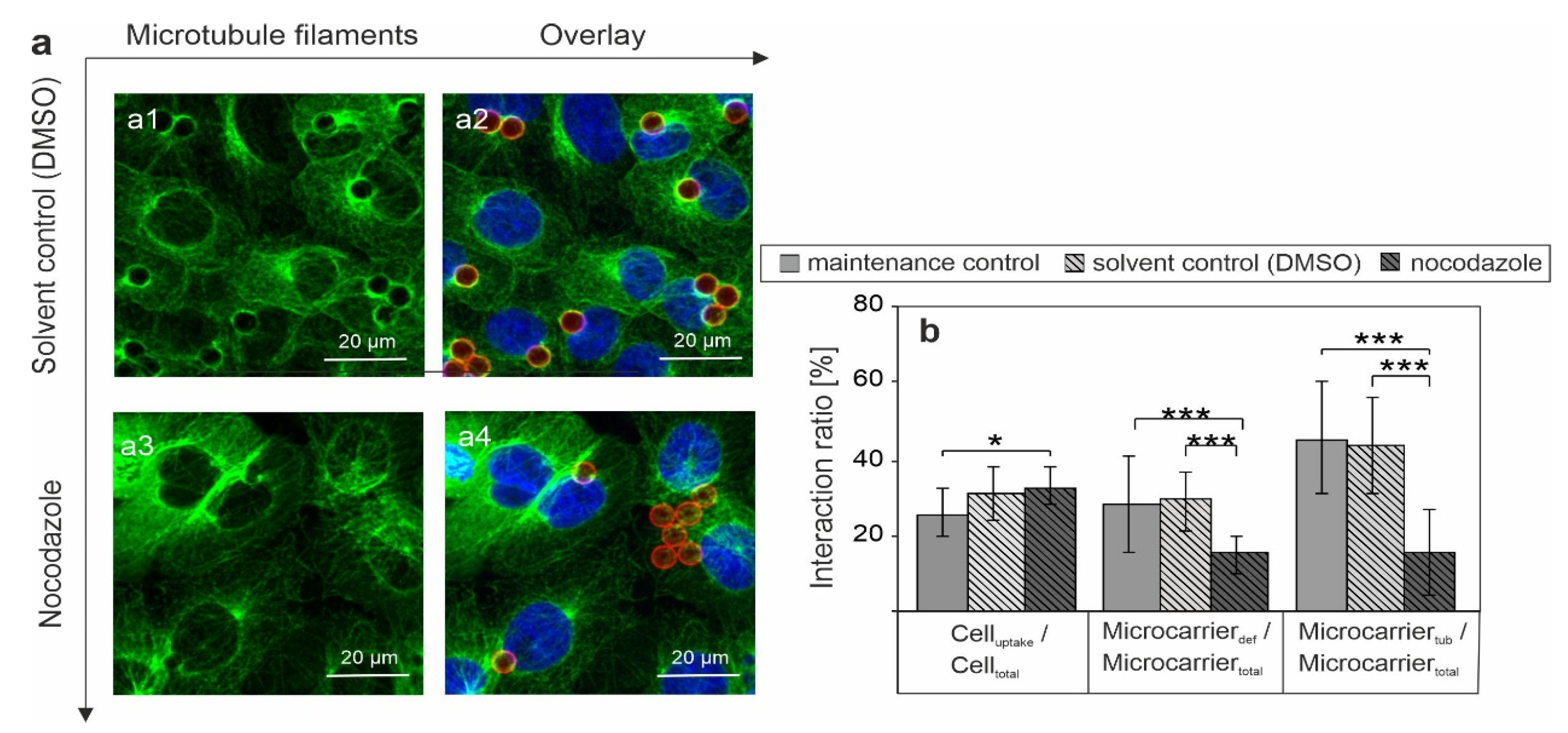
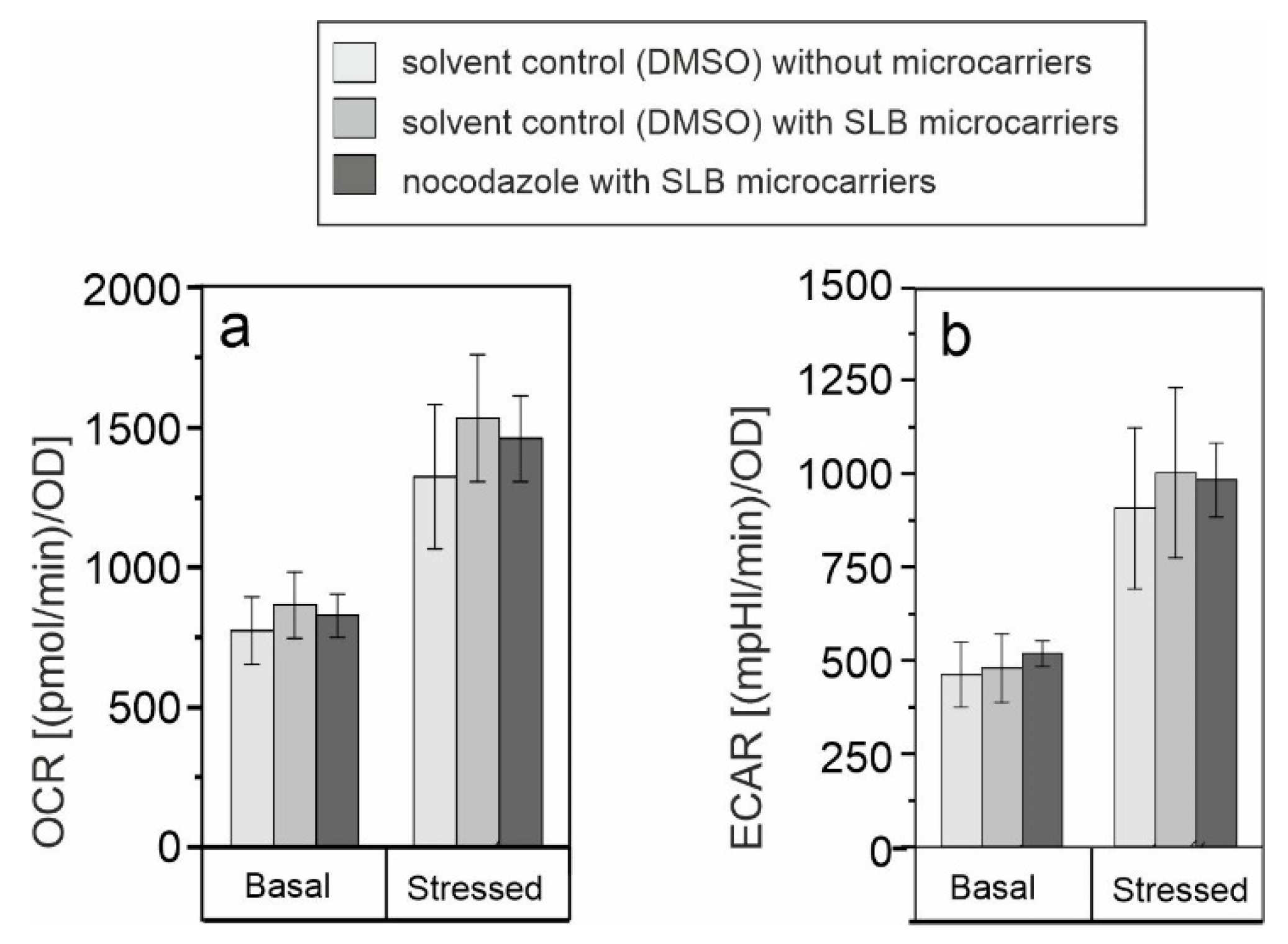
3.4. Assessment of the Association of SLB Microcarrier-Induced Metabolic Alterations with Microtubule Polymerization
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jackson, L.A.; Anderson, E.J.; Rouphael, N.G.; Roberts, P.C.; Makhene, M.; Coler, R.N.; McCullough, M.P.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; et al. An mRNA vaccine against SARS-CoV-2 — Preliminary Report. N. Engl. J. Med. 2020, 383, 1920–1931. [Google Scholar] [CrossRef]
- Wang, Z.; Schmidt, F.; Weisblum, Y.; Muecksch, F.; Barnes, C.O.; Finkin, S.; Schaefer-Babajew, D.; Cipolla, M.; Gaebler, C.; Lieberman, J.A.; et al. mRNA vaccine-elicited antibodies to SARS-CoV-2 and circulating variants. Nature 2021, 592, 616–638. [Google Scholar] [CrossRef] [PubMed]
- Gao, A.; Hu, X.-L.; Saeed, M.; Chen, B.-F.; Li, Y.-P.; Yu, H.-J. Overview of recent advances in liposomal nanoparticle-based cancer immunotherapy. Acta Pharmacol. Sin. 2019, 40, 1129–1137. [Google Scholar] [CrossRef] [Green Version]
- Billingsley, M.M.; Singh, N.; Ravikumar, P.; Zhang, R.; June, C.H.; Mitchell, M.J. Ionizable lipid nanoparticle-mediated mRNA delivery for human CAR T cell engineering. Nano Lett. 2020, 20, 1578–1589. [Google Scholar] [CrossRef]
- Jahanafrooz, Z.; Baradaran, B.; Mosafer, J.; Hashemzaei, M.; Rezaei, T.; Mokhtarzadeh, A.; Hamblin, M.R. Comparison of DNA and mRNA vaccines against cancer. Drug Discov. Today 2020, 25, 552–560. [Google Scholar] [CrossRef]
- Li, Y.; Cong, H.; Wang, S.; Yu, B.; Shen, Y. Liposomes modified with bio-substances for cancer treatment. Biomater. Sci. 2020, 8, 6442–6468. [Google Scholar] [CrossRef]
- Aldosari, B.N.; Alfagih, I.M.; Almurshedi, A.S. Lipid nanoparticles as delivery systems for RNA-based vaccines. Pharmaceutics 2021, 13, 206. [Google Scholar] [CrossRef]
- De Geest, B.G.; Vandenbroucke, R.E.; Guenther, A.M.; Sukhorukov, G.B.; Hennink, W.E.; Sanders, N.N.; Demeester, J.; De Smedt, S.C. Intracellularly degradable polyelectrolyte microcapsules. Adv. Mater. 2006, 18, 1005–1009. [Google Scholar] [CrossRef]
- De Koker, S.; De Geest, B.G.; Singh, S.K.; De Rycke, R.; Naessens, T.; Van Kooyk, Y.; Demeester, J.; De Smedt, S.C.; Grooten, J. Polyelectrolyte microcapsules as antigen delivery vehicles to dendritic cells: Uptake, processing, and cross-presentation of encapsulated antigens. Angew. Chem. Int. Ed. 2009, 48, 8485–8489. [Google Scholar] [CrossRef]
- Azarnezhad, A.; Samadian, H.; Jaymand, M.; Sobhani, M.; Ahmadi, A. Toxicological profile of lipid-based nanostructures: Are they considered as completely safe nanocarriers? Crit. Rev. Toxicol. 2020, 50, 148–176. [Google Scholar] [CrossRef]
- Nagai, N.; Ogata, F.; Otake, H.; Nakazawa, Y.; Kawasaki, N. Energy-dependent endocytosis is responsible for drug transcorneal penetration following the instillation of ophthalmic formulations containing indomethacin nanoparticles. Int. J. Nanomed. 2019, 14, 1213–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reibetanz, U.; Claus, C.; Typlt, E.; Hofmann, J.; Donath, E. Defoliation and plasmid delivery with layer-by-layer coated colloids. Macromol. Biosci. 2006, 6, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Brueckner, M.; Jankuhn, S.; Jülke, E.-M.; Reibetanz, U. Cellular interaction of a layer-by-layer based drug delivery system depending on material properties and cell types. Int. J. Nanomed. 2018, 13, 2079–2091. [Google Scholar] [CrossRef] [Green Version]
- Brueckner, M.; Scheffler, K.; Reibetanz, U. Enhanced cytoplasmic release of drug delivery systems: Chloroquine as a multilayer and template constituent of layer-by-layer microcarriers. J. Mater. Chem. B 2018, 6, 5153–5163. [Google Scholar] [CrossRef] [PubMed]
- Scheffler, K.; Bilz, N.C.; Brueckner, M.; Stanifer, M.L.; Boulant, S.; Claus, C.; Reibetanz, U. Enhanced uptake and endosomal release of LbL microcarriers functionalized with reversible fusion proteins. ACS Appl. Bio Mater. 2020, 3, 1553–1567. [Google Scholar] [CrossRef]
- Reibetanz, U.; Schönberg, M.; Rathmann, S.; Strehlow, V.; Göse, M.; Leßig, J. Inhibition of human neutrophil elastase by α1-antitrypsin functionalized colloidal microcarriers. ACS Nano 2012, 6, 6325–6336. [Google Scholar] [CrossRef]
- Reibetanz, U.; Hübner, D.; Jung, M.; Liebert, U.G.; Claus, C. Influence of growth characteristics of iPSCs on their uptake efficiency for LbL microcarriers. ACS Nano 2016, 10, 6563–6573. [Google Scholar] [CrossRef]
- Timin, A.S.; Gould, D.J.; Sukhorukov, G.B. Multi-layer microcapsules: Fresh insights and new applications. Expert Opin. Drug Deliv. 2017, 14, 583–587. [Google Scholar] [CrossRef] [Green Version]
- Tracuma, E.; Loca, D. Hyaluronic acid/polylysine composites for local drug delivery: A review. Key Eng. Mater. 2020, 850, 213–218. [Google Scholar] [CrossRef]
- Pahal, S.; Gakhar, R.; Raichur, A.M.; Varma, M.M. Polyelectrolyte multilayers for bio-applications: Recent advancements. IET Nanobiotechnol. 2017, 11, 903–908. [Google Scholar] [CrossRef]
- Sarode, A.; Annapragada, A.; Guo, J.; Mitragotri, S. Layered self-assemblies for controlled drug delivery: A translational overview. Biomaterials 2020, 242, 119929. [Google Scholar] [CrossRef]
- Skirtach, A.G.; Yashchenok, A.M.; Möhwald, H. Encapsulation, release and applications of LbL polyelectrolyte multilayer capsules. Chem. Commun. 2011, 47, 12736–12746. [Google Scholar] [CrossRef]
- Polomska, A.; Leroux, J.-C.; Brambilla, D. Layer-by-Layer coating of solid drug cores: A versatile method to improve stability, control release and tune surface properties. Macromol. Biosci. 2017, 17, 1600228. [Google Scholar] [CrossRef]
- Ghiorghita, C.-A.; Bucatariu, F.; Stela Dragan, E. Influence of cross-linking in loading/release applications of polyelectrolyte multilayer assemblies. A review. Mater. Sci. Eng. C 2019, 105, 110050. [Google Scholar] [CrossRef]
- Goese, M.; Pescador, P.; Reibetanz, U. Design of a homogeneous multifunctional supported lipid layer on LbL-coated microcarriers. Biomacromolecules 2015, 16, 757–768. [Google Scholar] [CrossRef]
- Goese, M.; Scheffler, K.; Reibetanz, U. Specific uptake of lipid-antibody-functionalized LbL microcarriers by cells. Biomacromolecules 2016, 17, 3672–3682. [Google Scholar] [CrossRef] [PubMed]
- Osada, N.; Kohara, A.; Yamaji, T.; Hirayama, N.; Kasai, F.; Sekizuka, T.; Kuroda, M.; Hanada, K. The genome landscape of the african green monkey kidney-derived vero cell line. DNA Res. 2014, 21, 673–683. [Google Scholar] [CrossRef] [Green Version]
- Schilling, E.; Weiss, R.; Grahnert, A.; Bitar, M.; Sack, U.; Hauschildt, S. Molecular mechanism of lps-induced tnf-α biosynthesis in polarized human macrophages. Mol. Immunol. 2018, 93, 206–215. [Google Scholar] [CrossRef]
- Ferrick, D.A.; Neilson, A.; Beeson, C. Advances in measuring cellular bioenergetics using extracellular flux. Drug Disov. Today 2008, 13, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Nuebel, E.; Wisidagama, D.R.R.; Setoguchi, K.; Hong, J.S.; van Horn, C.M.; Imam, S.S.; Vergnes, L.; Malone, C.S.; Koehler, C.M.; et al. Measuring energy metabolism in cultured cells, including human pluripotent stem cells and differentiated cells. Nat. Protoc. 2012, 7, 1068–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuang, Y.; Han, X.; Xu, M.; Yang, Q. Oxaloacetate induces apoptosis in HepG2 cells via inhibition of glycolysis. Cancer Med. 2018, 7, 1416–1429. [Google Scholar] [CrossRef]
- Sukhorukov, G.B.; Donath, E.; Lichtenfeld, H.; Knippel, E.; Knippel, M.; Budde, A.; Moehwald, H. Layer-by-layer self assembly of polyelectrolytes on colloidal particles. Colloids Surf. A Physicochem. Eng. Asp. 1998, 137, 253–266. [Google Scholar] [CrossRef]
- Brueckner, M.; Hollenbach-Latzko, S.; Reibetanz, U. Dual transport of active substances with a layer-by-layer-based drug delivery system to terminate inflammatory processes. Macromol. Biosci. 2020, 20, 97. [Google Scholar] [CrossRef]
- Mehta, K.; Chacko, L.A.; Chug, M.K.; Jhunjhunwala, S.; Ananthanarayanan, V. Association of mitochondria with microtubules inhibitsmitochondrial fission by precluding assembly of the fissionprotein Dnm1. J. Biol. Chem. 2019, 294, 3385–3396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, J.L.; Keith, T.J.; Knull, H.R. Glycolytic enzyme interactions with tubulin and microtubules. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1989, 999, 64–70. [Google Scholar] [CrossRef]
- Wakhloo, N.T.; Anders, S.; Badique, F.; Eichhorn, M.; Brigaud, I.; Petithory, T.; Vassaux, M.; Milan, J.-L.; Freund, J.-N.; Rühe, J.; et al. Actomyosin, vimentin and LINC complex pull on osteosarcoma nuclei to deform on micropillar topography. Biomaterials 2020, 234, 119746. [Google Scholar] [CrossRef] [PubMed]
- Doolin, M.T.; Ornstein, T.S.; Stroka, K.M. Nuclear deformation in response to mechanical confinement is cell type dependent. Cells 2019, 8, 427. [Google Scholar] [CrossRef] [Green Version]
- Pan, L.; Liu, J.; Shi, J. Cancer cell nucleus-targeting nanocomposites for advanced tumor therapeutics. Chem. Soc. Rev. 2018, 47, 6930–6947. [Google Scholar] [CrossRef]
- Rathmann, S.; Schönberg, M.; Leßig, J.; Reibetanz, U. Interaction, uptake, and processing of LbL-coated microcarriers by PMNs. Cytom. Part A 2011, 79A, 979–989. [Google Scholar] [CrossRef] [PubMed]
- Kruppa, A.J.; Buss, F. Motor proteins at the mitochondria–cytoskeleton interface. J. Cell Sci. 2021, 134, jcs226084. [Google Scholar] [CrossRef]
- Khan, S.; Raja, D.; Jaiswal, K.; Lahiri, A. Modulation of host mitochondrial dynamics during bacterial infection. Mitochondrion 2020, 53, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Galvao, J.; Davis, B.; Tilley, M.; Normando, E.; Duchen, M.R.; Cordeiro, M.F. Unexpected low-dose toxicity of the universal solvent DMSO. FASEB 2013, 28, 1317–1330. [Google Scholar] [CrossRef] [PubMed]
- de Menorval, M.-A.; Mir, L.M.; Fernandez, M.L.; Reigada, R. Effects of dimethyl sulfoxide in cholesterol-containing lipid membranes: A comparative study of experiments in silico and with cells. PLoS ONE 2012, 7, e41733. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Claus, C.; Fritz, R.; Schilling, E.; Reibetanz, U. The Metabolic Response of Various Cell Lines to Microtubule-Driven Uptake of Lipid- and Polymer-Coated Layer-by-Layer Microcarriers. Pharmaceutics 2021, 13, 1441. https://doi.org/10.3390/pharmaceutics13091441
Claus C, Fritz R, Schilling E, Reibetanz U. The Metabolic Response of Various Cell Lines to Microtubule-Driven Uptake of Lipid- and Polymer-Coated Layer-by-Layer Microcarriers. Pharmaceutics. 2021; 13(9):1441. https://doi.org/10.3390/pharmaceutics13091441
Chicago/Turabian StyleClaus, Claudia, Robert Fritz, Erik Schilling, and Uta Reibetanz. 2021. "The Metabolic Response of Various Cell Lines to Microtubule-Driven Uptake of Lipid- and Polymer-Coated Layer-by-Layer Microcarriers" Pharmaceutics 13, no. 9: 1441. https://doi.org/10.3390/pharmaceutics13091441