
Traditional Free-Ranging Livestock Farming as a Management Strategy for Biological and Cultural Landscape Diversity: A Case from the Southern Apennines

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
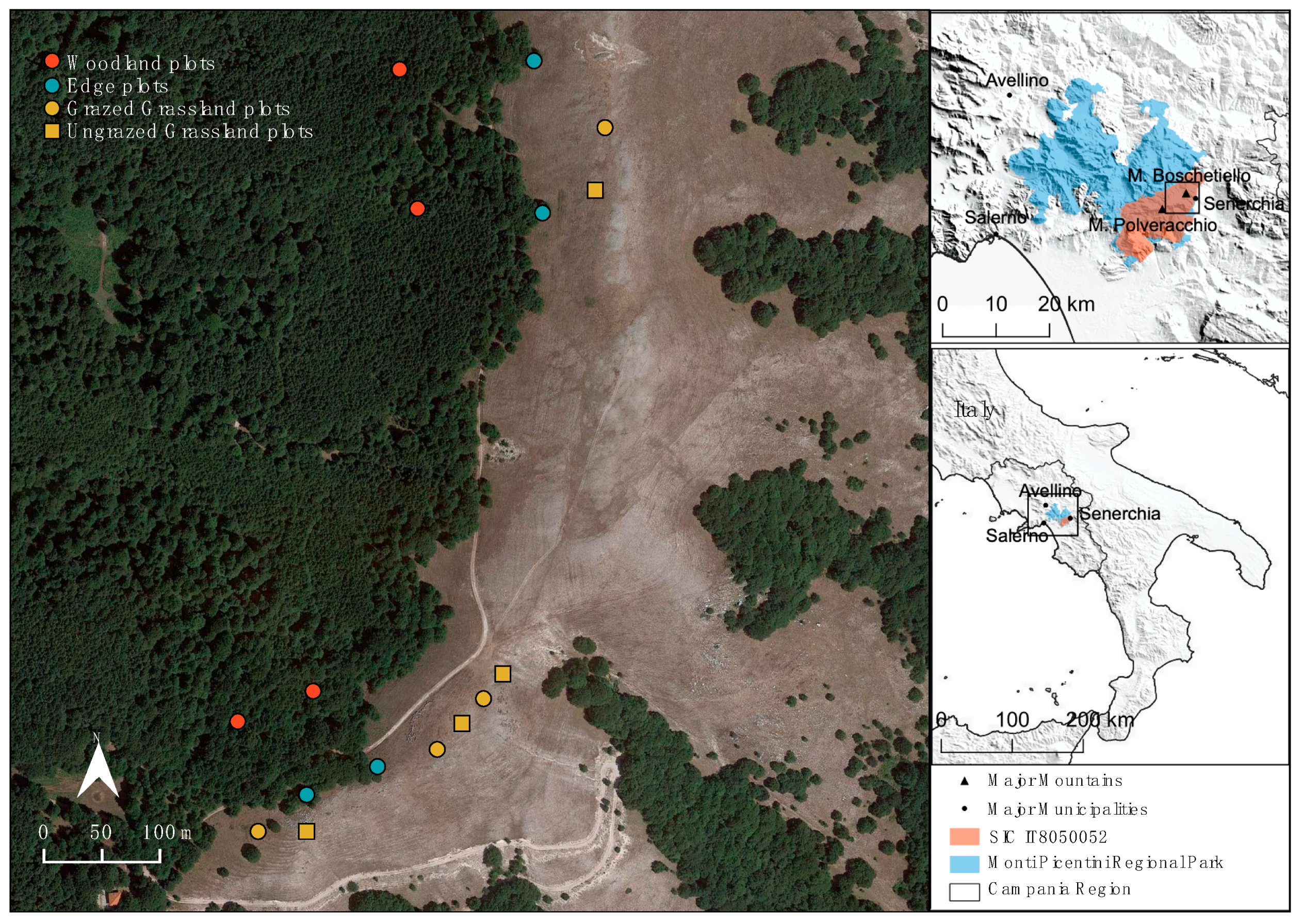
2.1. Study Area
2.2. Experimental Design
2.3. Arthropod Sampling
2.4. Floristic Characterization
2.5. Data Analysis
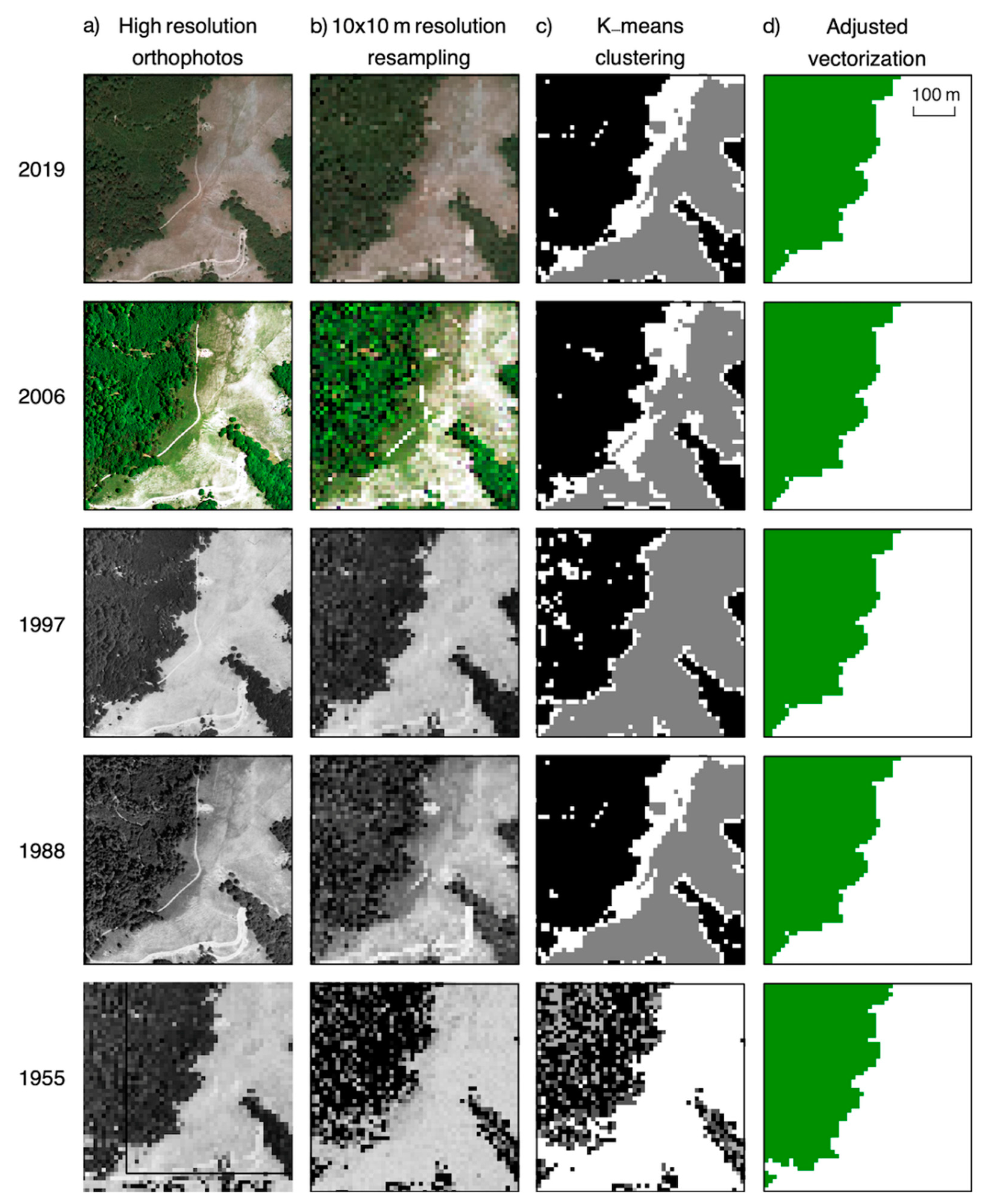
2.6. Forest Margin Expansion
2.7. Interviews
3. Results
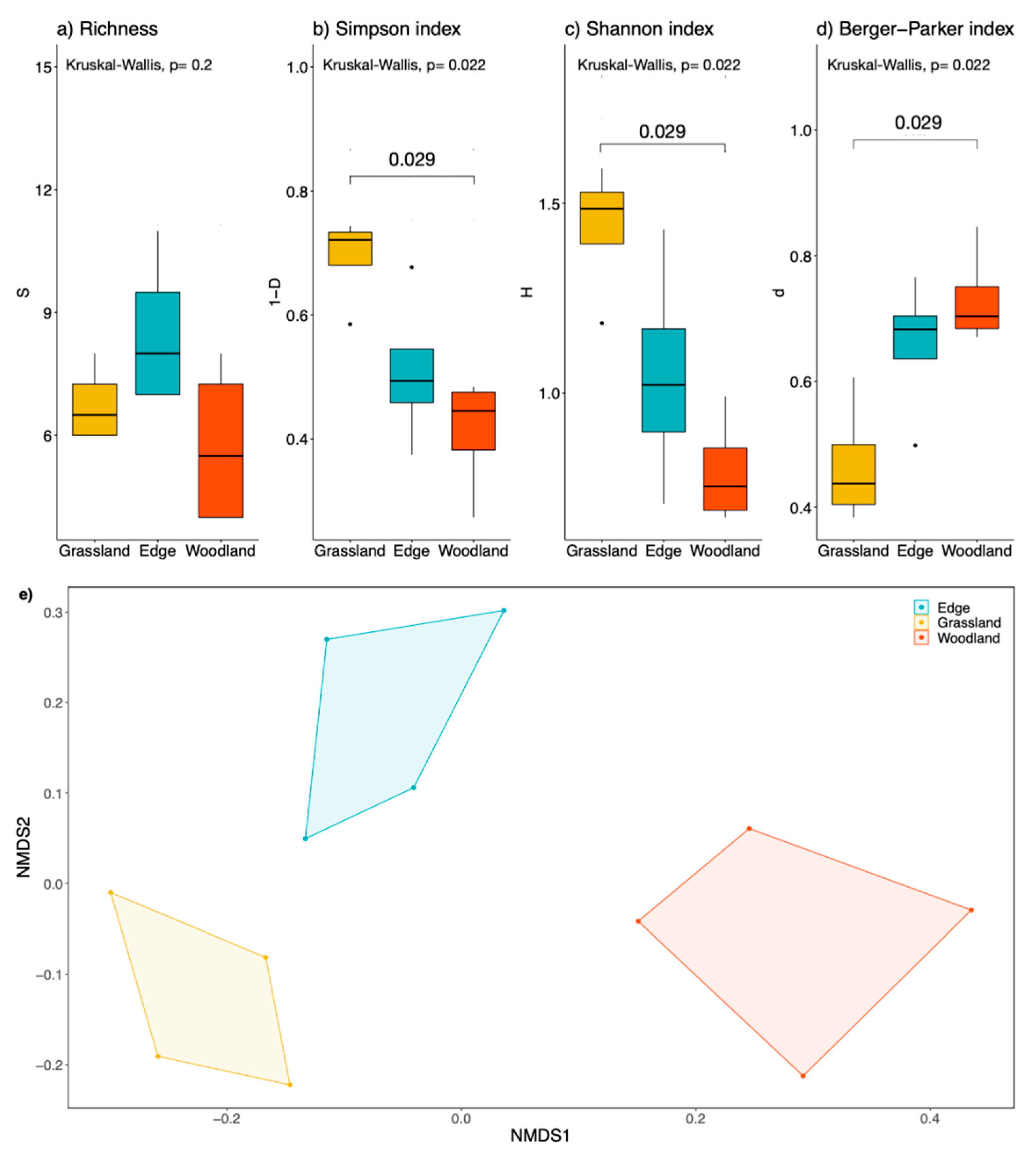
3.1. Animal Diversity in the Pastures
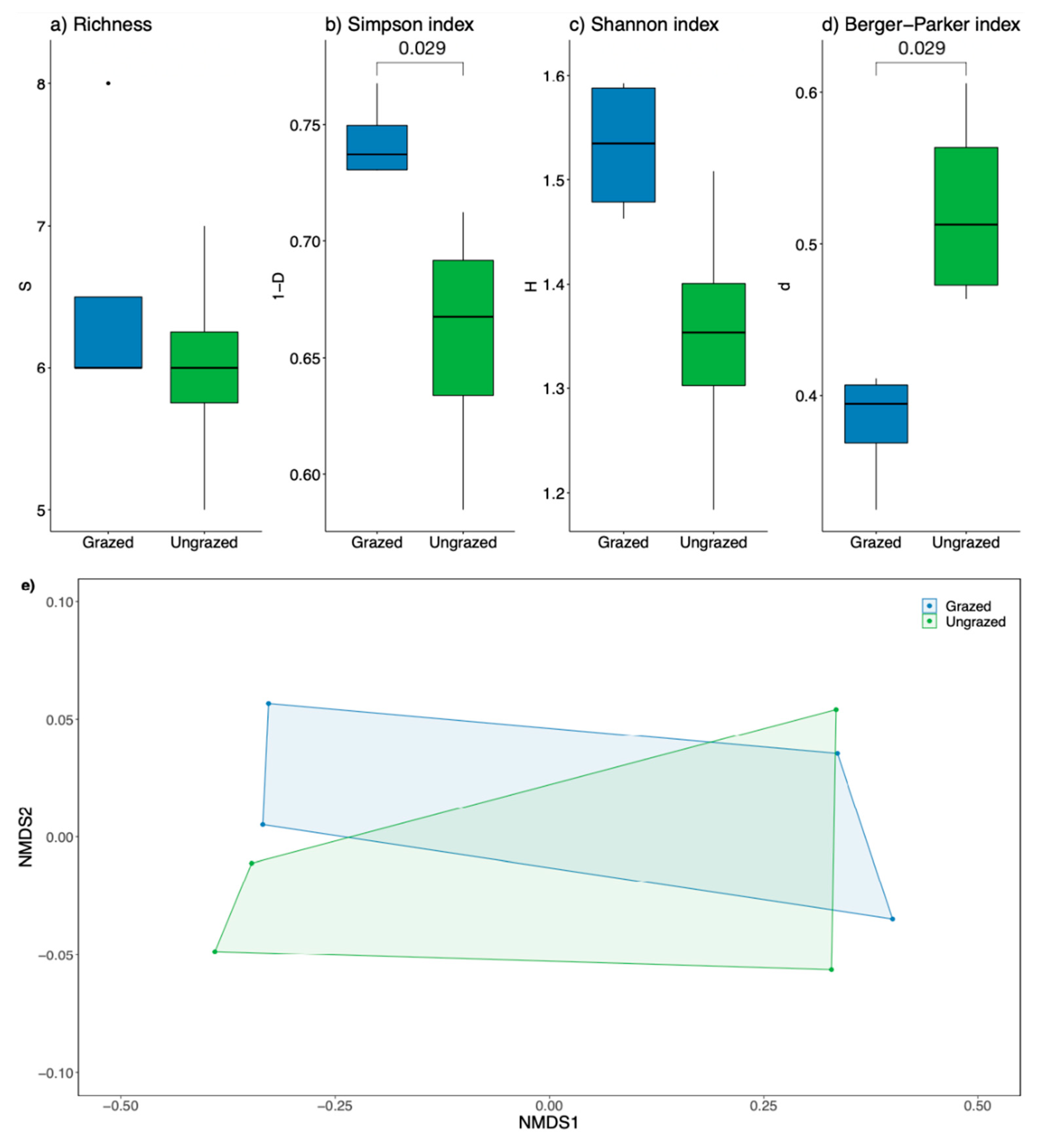
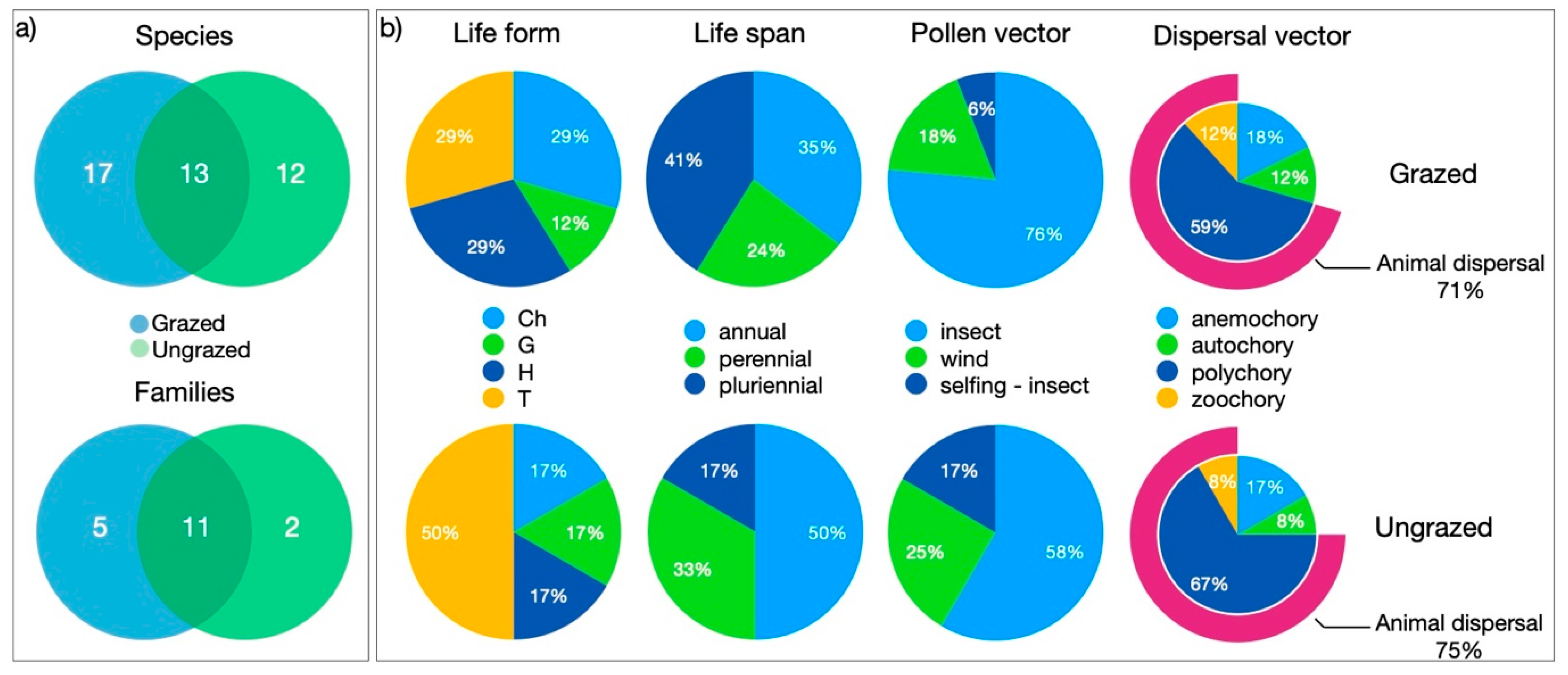
3.2. Floristic Diversity
3.3. Forest Expansion
3.4. Interviews
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barnosky, A.D.; Matzke, N.; Tomiya, S.; Wogan, G.O.; Swartz, B.; Quental, T.B.; Marshall, C.; McGuire, J.L.; Lindsey, E.L.; Maguire, K.C.; et al. Has the earth’s sixth mass extinction already arrived? Nature 2011, 471, 51–57. [Google Scholar] [CrossRef]
- Millennium Ecosystem Assessment. Ecosystems and Human Well-Being: Synthesis; Island Press: Washington, DC, USA, 2005; ISBN 978-1-59726-040-4. [Google Scholar]
- Bravo, D.N.; Araújo, M.B.; Lasanta, T.; Moreno, J.I.L. Climate change in mediterranean mountains during the 21st century. AMBIO A J. Hum. Environ. 2008, 37, 280–285. [Google Scholar] [CrossRef]
- Foley, J.A. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [Green Version]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity: Biodiversity and climate change. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [Green Version]
- Queiroz, C.; Beilin, R.; Folke, C.; Lindborg, R. Farmland abandonment: Threat or opportunity for biodiversity conservation? A global review. Front. Ecol. Environ. 2014, 12, 288–296. [Google Scholar] [CrossRef]
- Debussche, M.; Lepart, J.; Dervieux, A. Mediterranean landscape changes: Evidence from old postcards. Glob. Ecol. Biogeogr. 1999, 8, 3–15. [Google Scholar] [CrossRef]
- The Mediterranean Region: Biological Diversity in Space and Time, 2nd ed.; Blondel, J. (Ed.) Oxford University Press: New York, NY, USA, 2010; ISBN 978-0-19-955799-8. [Google Scholar]
- Agnoletti, M. Rural landscape, nature conservation and culture: Some notes on research trends and management approaches from a (southern) European perspective. Landsc. Urban Plan. 2014, 126, 66–73. [Google Scholar] [CrossRef]
- Di Rita, F.; Magri, D. Holocene drought, deforestation and evergreen vegetation development in the central Mediterranean: A 5500 year record from Lago Alimini Piccolo, Apulia, Southeast Italy. Holocene 2009, 19, 295–306. [Google Scholar] [CrossRef]
- Woodbridge, J.; Roberts, N.; Fyfe, R. Pan-Mediterranean holocene vegetation and land-cover dynamics from synthesized pollen data. J. Biogeogr. 2018, 45, 2159–2174. [Google Scholar] [CrossRef] [Green Version]
- UNESCO World Heritage Cultural Landscapes: A Handbook for Conservation and Management. In World Heritage Papers; UNESCO World Heritage Centre: Paris, France, 2009; Volume 26.
- Guidelines for Protected Area Management Categories; IUCN/UICN: Cambridge, UK, 1994; ISBN 978-2-8317-0201-8.
- Naveh, Z. Interactions of landscapes and cultures. Landsc. Urban Plan. 1995, 32, 43–54. [Google Scholar] [CrossRef]
- The Protected Landscape Approach: Linking Nature, Culture, and Community; Brown, J.; Mitchell, N.; Beresford, M. (Eds.) IUCN—The World Conservation Union: Gland, Switzerland, 2005; ISBN 978-2-8317-0797-6. [Google Scholar]
- Taylor, K.; Lennon, J. Cultural landscapes: A bridge between culture and nature? Int. J. Herit. Stud. 2011, 17, 537–554. [Google Scholar] [CrossRef]
- Mittermeier, R.A.; Turner, W.R.; Larsen, F.W.; Brooks, T.M.; Gascon, C. Global biodiversity conservation: The critical Role of hotspots. In Biodiversity Hotspots; Zachos, F.E., Habel, J.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 3–22. ISBN 978-3-642-20991-8. [Google Scholar]
- Vacchiano, G.; Garbarino, M.; Lingua, E.; Motta, R. Forest dynamics and disturbance regimes in the Italian Apennines. For. Ecol. Manag. 2017, 388, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Malandra, F.; Vitali, A.; Urbinati, C.; Garbarino, M. 70 Years of land use/land cover changes in the Apennines (Italy): A Meta-Analysis. Forests 2018, 9, 551. [Google Scholar] [CrossRef] [Green Version]
- Barker, G. Agriculture, pastoralism, and Mediterranean landscapes in prehistory. In The Archaeology of Mediterranean Prehistory; Blake, E., Knapp, A.B., Eds.; Blackwell Pub.: Malden, MA, USA, 2005; pp. 46–76. ISBN 978-0-631-23268-1. [Google Scholar]
- Brown, A.G.; Hatton, J.; Selby, K.A.; Leng, M.J.; Christie, N. Multi-proxy study of holocene environmental change and human activity in the central Apennine mountains, Italy. J. Quat. Sci. 2013, 28, 71–82. [Google Scholar] [CrossRef]
- Schreiber, H. 14th Session of the intergovernmental committee for the safeguarding of the intangible cultural heritage. Santander Art Cult. Law Rev. 2019, 5, 293–295. [Google Scholar]
- Bakudila, A.; Fassio, F.; Sallustio, L.; Marchetti, M.; Munafò, M.; Ritano, N. I Comuni e le Comunità Appenninici: Evoluzione del Territorio. Available online: http://www.slowfood.it/stati-generali-delle-comunita-dellappennino/ (accessed on 2 January 2021).
- Hanke, W.; Böhner, J.; Dreber, N.; Jürgens, N.; Schmiedel, U.; Wesuls, D.; Dengler, J. The impact of livestock grazing on plant diversity: An analysis across dryland ecosystems and scales in Southern Africa. Ecol. Appl. 2014, 24, 1188–1203. [Google Scholar] [CrossRef] [Green Version]
- Komac, B.; Domènech, M.; Fanlo, R. Effects of grazing on plant species diversity and pasture quality in subalpine grasslands in the Eastern Pyrenees (Andorra): Implications for conservation. J. Nat. Conserv. 2014, 22, 247–255. [Google Scholar] [CrossRef]
- Vera, F.W.M. Grazing Ecology and Forest History; Cabi: Wallingford, UK, 2000. [Google Scholar]
- Rotherham, I.D. Trees, Forested Landscapes and Grazing Animals: A European Perspective on Woodlands and Grazed Treescapes; Routledge: London, UK, 2013; ISBN 0-415-62611-0. [Google Scholar]
- Bruun, H.H.; Fritzbøger, B. The past impact of livestock husbandry on dispersal of plant seeds in the landscape of Denmark. AMBIO A J. Hum. Environ. 2002, 31, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Wolański, P.; Bobiec, A.; Ortyl, B.; Makuch-Pietraś, I.; Czarnota, P.; Ziobro, J.; Korol, M.; Havryliuk, S.; Paderewski, J.; Kirby, K. The importance of livestock grazing at Woodland-Grassland interface in the conservation of rich oakwood plant communities in temperate Europe. Biodivers Conserv. 2021, 30, 741–760. [Google Scholar] [CrossRef]
- Sorino, R.; Santarcangelo, V.; Tarasco, E.; Corriero, G. Struttura di comunità degli ortotteri e integrità dei sistemi agro-pastorali del sic-zps murgia alta (Puglia, Italia). Entomologica 2008, 41, 219–231. [Google Scholar]
- Villanueva-López, G.; Lara-Pérez, L.A.; Oros-Ortega, I.; Ramirez-Barajas, P.J.; Casanova-Lugo, F.; Ramos-Reyes, R.; Aryal, D.R. Diversity of soil macro-arthropods correlates to the richness of plant species in traditional agroforestry systems in the humid tropics of Mexico. Agric. Ecosyst. Environ. 2019, 286, 106658. [Google Scholar] [CrossRef]
- Ancillotto, L.; Festa, F.; De Benedetta, F.; Cosentino, F.; Pejic, B.; Russo, D. Free-ranging livestock and a diverse landscape structure increase bat foraging in mountainous landscapes. Agrofor. Syst. 2021, 95, 407–418. [Google Scholar] [CrossRef]
- Buglione, M.; Petrelli, S.; de Filippo, G.; Troiano, C.; Rivieccio, E.; Notomista, T.; Maselli, V.; di Martino, L.; Carafa, M.; Gregorio, R.; et al. Contribution to the ecology of the Italian hare (Lepus Corsicanus). Sci. Rep. 2020, 10, 13071. [Google Scholar] [CrossRef]
- Buglione, M.; Troisi, S.R.; Petrelli, S.; van Vugt, M.; Notomista, T.; Troiano, C.; Bellomo, A.; Maselli, V.; Gregorio, R.; Fulgione, D. The first report on the ecology and distribution of the wolf population in Cilento, Vallo Di Diano and Alburni National Park. Biol. Bull. 2020, 47, 640–654. [Google Scholar] [CrossRef]
- Pinilla, V.; Ayuda, M.-I.; Sáez, L.-A. Rural depopulation and the migration turnaround in Mediterranean western Europe: A case study of Aragon. J. Rural. Community Dev. 2008, 3, 1–22. [Google Scholar]
- Rippa, D.; Maselli, V.; Soppelsa, O.; Fulgione, D. The Impact of Agro-pastoral Abandonment on the Rock Partridge Alectoris Graeca in the Apennines. Ibis 2011, 153, 721–734. [Google Scholar] [CrossRef]
- Métailié, J.-P.; Paegelow, M. Land abandonment and the spreading of the forest in the eastern French Pyrenees in the nineteenth to twentieth centuries. In Recent Dynamics of the Mediterranean Vegetation and Landscape; Wiley: Chichester, UK, 2004; pp. 219–236. [Google Scholar]
- Sokos, C.K.; Mamolos, A.P.; Kalburtji, K.L.; Birtsas, P.K. Farming and wildlife in Mediterranean agroecosystems. J. Nat. Conserv. 2013, 21, 81–92. [Google Scholar] [CrossRef]
- De Sillo, R.; De Sanctis, M.; Bruno, F.; Attorre, F. Vegetation and landscape of the Simbruini mountains (Central Appenines). Plant Sociol. 2012, 3–64. [Google Scholar] [CrossRef]
- Munafò, M.; Assennato, F.; Congedo, L.; Luti, T.; Marinosci, I.; Monti, G.; Riitano, N.; Sallustio, L.; Strollo, A.; Tomboliniand, I.; et al. Il Consumo di Suolo in Italia 2015; Ispra: Rome, Italy, 2015. [Google Scholar]
- Kozak, J.; Ziółkowska, E.; Vogt, P.; Dobosz, M.; Kaim, D.; Kolecka, N.; Ostafin, K. Forest-cover increase does not trigger forest-fragmentation decrease: Case study from the Polish carpathians. Sustainability 2018, 10, 1472. [Google Scholar] [CrossRef] [Green Version]
- Malandra, F.; Vitali, A.; Urbinati, C.; Weisberg, P.J.; Garbarino, M. Patterns and drivers of forest landscape change in the Apennines range, Italy. Reg. Environ. Chang. 2019, 19, 1973–1985. [Google Scholar] [CrossRef]
- Guilherme, J.L.; Miguel Pereira, H. Adaptation of bird communities to farmland abandonment in a mountain landscape. PLoS ONE 2013, 8, e73619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regos, A.; Domínguez, J.; Gil-Tena, A.; Brotons, L.; Ninyerola, M.; Pons, X. Rural abandoned landscapes and bird assemblages: Winners and losers in the rewilding of a Marginal mountain area (NW Spain). Reg. Environ. Chang. 2016, 16, 199–211. [Google Scholar] [CrossRef]
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef] [Green Version]
- Durant, J.M.; Hjermann, D.Ø.; Ottersen, G.; Stenseth, N.C. Climate and the match or mismatch between predator requirements and resource availability. Clim. Res. 2007, 33, 271–283. [Google Scholar] [CrossRef] [Green Version]
- Halada, L.; Evans, D.; Romão, C.; Petersen, J.-E. Which habitats of European importance depend on agricultural practices? Biodivers Conserv. 2011, 20, 2365–2378. [Google Scholar] [CrossRef]
- Salvati, L.; Tombolini, I.; Perini, L.; Ferrara, A. Landscape changes and environmental quality: The evolution of land vulnerability and potential resilience to degradation in Italy. Reg. Environ. Change 2013, 13, 1223–1233. [Google Scholar] [CrossRef]
- Assini, S.; Filipponi, F.; Zucca, F. Land cover changes in an abandoned agricultural land in the Northern Apennine (Italy) between 1954 and 2008: Spatio-temporal dynamics. Plant Biosyst.—Int. J. Deal. All Asp. Plant Biol. 2015, 149, 807–817. [Google Scholar] [CrossRef]
- Lasanta, T.; González-Hidalgo, J.C.; Vicente-Serrano, S.M.; Sferi, E. Using landscape ecology to evaluate an alternative management scenario in abandoned Mediterranean mountain areas. Landsc. Urban Plan. 2006, 78, 101–114. [Google Scholar] [CrossRef]
- Maselli, V.; Rippa, D.; Russo, G.; Ligrone, R.; Soppelsa, O.; D’Aniello, B.; Fulgione, D. Wild boars’ social structure in the Mediterranean habitat. Ital. J. Zool. 2014, 81, 610–617. [Google Scholar] [CrossRef] [Green Version]
- Maselli, V.; Rippa, D.; Deluca, A.; Larson, G.; Wilkens, B.; Linderholm, A.; Masseti, M.; Fulgione, D. Southern Italian wild boar population, hotspot of genetic diversity. Hystrix Ital. J. Mammal. 2016, 27. [Google Scholar] [CrossRef]
- Acevedo, P.; Farfán, M.Á.; Márquez, A.L.; Delibes-Mateos, M.; Real, R.; Vargas, J.M. Past, present and future of wild ungulates in relation to changes in land use. Landsc. Ecol 2011, 26, 19–31. [Google Scholar] [CrossRef]
- Massei, G.; Genov, P. The environmental impact of wild boar. Galemys 2004, 16, 135–145. [Google Scholar]
- Barrios-Garcia, M.N.; Ballari, S.A. Impact of wild boar (Sus Scrofa) in its introduced and native range: A review. Biol. Invasions 2012, 14, 2283–2300. [Google Scholar] [CrossRef]
- Scandurra, A.; Magliozzi, L.; Fulgione, D.; Aria, M.; D’Aniello, B. Lepidoptera papilionoidea communities as a sentinel of biodiversity threat: The case of wild boar rooting in a mediterranean habitat. J. Insect Conserv. 2016, 20, 353–362. [Google Scholar] [CrossRef]
- Conti, G.; Fagarazzi, L. Avanzamento del bosco in ecosistemi montani: “sogno degli ambientalisti o incubo per la società”? Cause, aspetti ed impatti di una delle principali trasformazioni ambientali, territoriali e paesistiche del XX secolo in Italia. PLANUM: Eur. Online J. Plan. 2004, 10, 1–20. [Google Scholar]
- Bakudila, A. LA PASTORIZIA NELLE AREE APPENNINICHE: ANALISI Dell’Ultimo DECENNIO. Available online: https://www.slowfood.it/wp-content/uploads/2018/07/LA-PASTORIZIA-NELLE-AREE-APPENNINICHE-1.pdf (accessed on 3 January 2021).
- Antrop, M. The concept of traditional landscapes as a base for landscape evaluation and planning. the example of flanders region. Landsc. Urban Plan. 1997, 38, 105–117. [Google Scholar] [CrossRef]
- Bender, O.; Boehmer, H.J.; Jens, D.; Schumacher, K.P. Using GIS to analyse long-term cultural landscape change in Southern Germany. Landsc. Urban Plan. 2005, 70, 111–125. [Google Scholar] [CrossRef]
- Hussain, R.I.; Walcher, R.; Eder, R.; Allex, B.; Wallner, P.; Hutter, H.-P.; Bauer, N.; Arnberger, A.; Zaller, J.G.; Frank, T. Management of mountainous meadows associated with biodiversity attributes, perceived health benefits and cultural ecosystem services. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Díaz, S.; Demissew, S.; Carabias, J.; Joly, C.; Lonsdale, M.; Ash, N.; Larigauderie, A.; Adhikari, J.R.; Arico, S.; Báldi, A.; et al. The IPBES conceptual framework—connecting nature and people. Curr. Opin. Environ. Sustain. 2015, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Nazarea, V.D. Local knowledge and memory in biodiversity conservation. Annu. Rev. Anthropol. 2006, 35, 317–335. [Google Scholar] [CrossRef]
- Sobala, M. Landscape effects of conflicts in space management. a historical approach based on the Silesian and Żywiec Beskids (West Carpathians, Poland). Environ. Socio-Econ. Stud. 2016, 4, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Van Meerbeek, K.; Muys, B.; Schowanek, S.D.; Svenning, J.-C. Reconciling conflicting paradigms of biodiversity conservation: Human intervention and rewilding. BioScience 2019, 69, 997–1007. [Google Scholar] [CrossRef]
- Gonçalves, P.; Alcobia, S.; Simões, L.; Santos-Reis, M. Effects of management options on mammal richness in a Mediterranean Agro-Silvo-Pastoral system. Agrofor. Syst. 2012, 85, 383–395. [Google Scholar] [CrossRef]
- Feurdean, A.; Ruprecht, E.; Molnár, Z.; Hutchinson, S.M.; Hickler, T. Biodiversity-rich european grasslands: Ancient, forgotten ecosystems. Biol. Conserv. 2018, 228, 224–232. [Google Scholar] [CrossRef]
- Biaggini, M.; Consorti, R.; Dapporto, L.; Dellacasa, M.; Paggetti, E.; Corti, C. The taxonomic level order as a possible tool for rapid assessment of arthropod diversity in agricultural landscapes. Agric. Ecosyst. Environ. 2007, 122, 183–191. [Google Scholar] [CrossRef]
- Raunkiaer, C. The life forms of plants and statistical plant geography; being the collected papers of C. Raunkiaer. In The Life Forms of Plants and Statistical Plant Geography; Being the Collected Papers of C. Raunkiaer; Clarendon Press: Oxford, UK, 1934. [Google Scholar]
- Klotz, S.; Kühn, I.; Durka, W. BIOLFLOR: Eine Datenbank Mit Biologisch-Ökologischen Merkmalen zur Flora von Deutschland; Schriftenreihe für Vegetationskunde; Bundesamt für Naturschutz: Bonn, Germany, 2002; ISBN 978-3-7843-3508-7. [Google Scholar]
- Fitter, A.H.; Peat, H.J. The ecological flora database. J. Ecol. 1994, 82, 415. [Google Scholar] [CrossRef]
- Pignatti, S.; Menegoni, P.; Pietrosanti, S. Biondicazione attraverso le piante Vascolari. Valori Di Indicazione Second. Ellenberg (Zeigerwerte) Per Le Specie Della Flora D’italia. Braun-Blanquetia 2005, 39, 97. [Google Scholar]
- Kleyer, M.; Bekker, R.M.; Knevel, I.C.; Bakker, J.P.; Thompson, K.; Sonnenschein, M.; Poschlod, P.; van Groenendael, J.M.; Klimeš, L.; Klimešová, J.; et al. The LEDA traitbase: A database of life-history traits of the Northwest European flora. J. Ecol. 2008, 96, 1266–1274. [Google Scholar] [CrossRef]
- Tavşanoğlu, Ç.; Pausas, J.G. A functional trait database for mediterranean basin plants. Sci. Data 2018, 5, 180135. [Google Scholar] [CrossRef] [Green Version]
- Bocci, G. TR8: An r package for easily retrieving plant species traits. Methods Ecol. Evol. 2015, 6, 347–350. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Champaign, IL, USA, 1949; Volume 96. [Google Scholar]
- Magurran, A.E. Measuring Biological Diversity; Wiley-Blackwell: Hoboken, NJ, USA, 2013; ISBN 978-1-118-68792-5. [Google Scholar]
- Berger, W.H.; Parker, F.L. Diversity of planktonic foraminifera in Deep-Sea sediments. Science 1970, 168, 1345–1347. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; ISBN 3-900051-07-0. [Google Scholar]
- Savage, A.M.; Youngsteadt, E.; Ernst, A.F.; Powers, S.A.; Dunn, R.R.; Frank, S.D. Homogenizing an urban habitat mosaic: Arthropod diversity declines in New York City Parks after super storm Sandy. Ecol. Appl. 2018, 28, 225–236. [Google Scholar] [CrossRef] [PubMed]
- MacQueen, J. Some methods for classification and analysis of multivariate observations. In Proceedings of the Fifth Berkeley Symposium on Mathematical Statistics and Probability, Berkeley, CA, USA, 21 June–18 July 1965; Volume 1, pp. 281–297. [Google Scholar]
- Batterton, K.A.; Hale, K.N. The likert scale what it is and how to use it. Phalanx 2017, 50, 32–39. [Google Scholar]
- Kark, S. Effects of ecotones on biodiversity. In Encyclopedia of Biodiversity; Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- San Miguel-Ayanz, A.; García-Calvo, R.P.; García-Olalla, M. Wild ungulates vs extensive livestock. Looking back to face the future. Options Meditérranneenes 2010, 92, 27–34. [Google Scholar]
- Svenning, J.-C. Rewilding should be central to global restoration efforts. One Earth 2020, 3, 657–660. [Google Scholar] [CrossRef]
- Prishchepov, A.V.; Radeloff, V.C.; Baumann, M.; Kuemmerle, T.; Müller, D. Effects of institutional changes on land use: Agricultural land abandonment during the transition from state-command to market-driven economies in Post-Soviet Eastern Europe. Environ. Res. Lett. 2012, 7, 024021. [Google Scholar] [CrossRef]
- Pătru-Stupariu, I.; Angelstam, P.; Elbakidze, M.; Huzui, A.; Andersson, K. Using forest history and spatial patterns to identify potential high conservation value forests in Romania. Biodivers. Conserv. 2013, 22, 2023–2039. [Google Scholar] [CrossRef]
- Kolecka, N.; Kozak, J. Wall-to-wall parcel-level mapping of agricultural land abandonment in the Polish Carpathians. Land 2019, 8, 129. [Google Scholar] [CrossRef] [Green Version]
- Sobala, M.; Myga-Piątek, U.; Szypuła, B. Assessment of Changes in a Viewshed in the Western Carpathians Landscape as a Result of Reforestation. Land 2020, 9, 430. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Information | Category | Percentage of Interviewees (n = 25) |
|---|---|---|
| Type of interviewees | Livestock farmers | 80 |
| Administrative employees | 20 | |
| Age | Age < 30 | 16 |
| 30 ≤ Age < 50 | 36 | |
| Age ≥ 50 | 48 | |
| Years farming | YF < 5 | 8 |
| 5 ≤ YF < 15 | 40 | |
| YF ≥ 15 | 52 | |
| Type of farming | Transhumant | 32 |
| Non-transhumant | 60 | |
| Stabled | 8 | |
| Type of livestock | Cattle | 48 |
| Sheep | 40 | |
| Horses | 12 | |
| Farming in protected areas | Yes | 56 |
| No | 44 | |
| Farming as main activity | Yes | 60 |
| No | 40 | |
| Common Agricultural Policy (CAP) direct payments | Yes | 60 |
| No | 40 | |
| CAP Rural Development Funds | Yes | 40 |
| No | 60 | |
| CAP income support | Yes | 60 |
| No | 40 | |
| Duration of CAP funds | No funds | 36 |
| Years of funding < 10 | 32 | |
| Years of funding ≥ 10 | 32 |
| Trait Category | Trait | Description | Grazed % | Ungrazed % |
|---|---|---|---|---|
| Raunkiær Life Form | Ch (Chamaephytes) | Plants with persistent shoot systems between 25 and 50 cm from the soil | 29.4 | 16.7 |
| G (Geophytes) | Plants with a periodic reduction of the shoot system to soil-embedded organs | 11.8 | 16.7 | |
| H (Hemicryptophytes) | Plants with a periodic reduction of the shoot system to the ground surface | 29.4 | 16.7 | |
| T (Therophytes) | Plants with shoot and root systems that die after production, with one-year life cycles | 29.4 | 50.0 | |
| Life Span | annual | Plants that complete their life cycle in one year | 35.3 | 50.0 |
| pluriennial | Plants that complete their life cycles in up to two years | 23.5 | 33.3 | |
| perennial | Plants that complete their life cycle in more than two years | 41.2 | 16.7 | |
| Dispersal Vector | anemochory | Wind-dispersed plants | 17.6 | 16.7 |
| autochory | Self-dispersed plants | 11.8 | 8.3 | |
| polychory | Plants dispersed by multiple vectors | 58.8 | 66.7 | |
| zoochory | Animal-dispersed plants | 11.8 | 8.3 | |
| Pollen Vector | insect | 76.5 | 58.3 | |
| wind | 17.6 | 25.0 | ||
| selfing-insect | 5.9 | 16.7 | ||
| Animal Dispersal | Yes | (zoochory + polychory) | 70.6 | 75.0 |
| No | 29.4 | 25.0 |
| Statement | 1—Disagree | 2—Neutral | 3—Agree | Mean | SD |
|---|---|---|---|---|---|
| Pastoralism and its traditions are important for personal and cultural identity | 0% | 40% | 60% | 2.6 | 0.50 |
| Traditional pastoralism is strongly linked to the landscape | 0% | 24% | 72% | 2.76 | 0.44 |
| Livestock farming has changed in the past decades | 0% | 32% | 68% | 2.68 | 0.48 |
| Land management has changed in the past decades | 0% | 32% | 68% | 2.68 | 0.48 |
| Rural de-population and pastoralism decline are directly linked | 0% | 28% | 72% | 2.72 | 0.48 |
| Hired help is necessary for pastoralism | 0% | 44% | 56% | 2.44 | 0.51 |
| Pastoralism provides sustainable income without agri-environmental schemes | 64% | 36% | 0% | 1.36 | 0.49 |
| Agri-environmental schemes are adequate to support local pastoralism | 84% | 16% | 0% | 1.16 | 0.48 |
| Schemes and regulations have an impact on production and commercialization | 0% | 36% | 64% | 2.64 | 0.50 |
| Traditional pastoralism has future prospects | 32% | 48% | 20% | 1.88 | 0.73 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Troiano, C.; Buglione, M.; Petrelli, S.; Belardinelli, S.; De Natale, A.; Svenning, J.-C.; Fulgione, D. Traditional Free-Ranging Livestock Farming as a Management Strategy for Biological and Cultural Landscape Diversity: A Case from the Southern Apennines. Land 2021, 10, 957. https://doi.org/10.3390/land10090957
Troiano C, Buglione M, Petrelli S, Belardinelli S, De Natale A, Svenning J-C, Fulgione D. Traditional Free-Ranging Livestock Farming as a Management Strategy for Biological and Cultural Landscape Diversity: A Case from the Southern Apennines. Land. 2021; 10(9):957. https://doi.org/10.3390/land10090957
Chicago/Turabian StyleTroiano, Claudia, Maria Buglione, Simona Petrelli, Sofia Belardinelli, Antonino De Natale, Jens-Christian Svenning, and Domenico Fulgione. 2021. "Traditional Free-Ranging Livestock Farming as a Management Strategy for Biological and Cultural Landscape Diversity: A Case from the Southern Apennines" Land 10, no. 9: 957. https://doi.org/10.3390/land10090957