
HMGA1, Moonlighting Protein Function, and Cellular Real Estate: Location, Location, Location!

1
Vall d’Hebron Institute of Oncology (VHIO), 08035 Barcelona, Spain
2
Department of Medicine, Division of Hematology, The Johns Hopkins University School of Medicine, Baltimore, MD 21205, USA
3
Departments of Medicine (Hematology), Oncology, Pathology and Institute of Cellular Engineering, The Johns Hopkins University School of Medicine, Baltimore, MD 21205, USA
4
Pathobiology, Cellular and Molecular Medicine and Human Genetics Graduate Programs, The Johns Hopkins University School of Medicine, Baltimore, MD 21205, USA
5
Centro de Investigación Biomédica en Red de Cáncer (CIBERONC), 28029 Madrid, Spain
*
Authors to whom correspondence should be addressed.
Biomolecules 2021, 11(9), 1334; https://doi.org/10.3390/biom11091334
Submission received: 30 July 2021
/
Revised: 27 August 2021
/
Accepted: 1 September 2021
/
Published: 9 September 2021
(This article belongs to the Special Issue HMG Proteins from Molecules to Disease)
Abstract
:The gene encoding the High Mobility Group A1 (HMGA1) chromatin remodeling protein is upregulated in diverse cancers where high levels portend adverse clinical outcomes. Until recently, HMGA1 was assumed to be a nuclear protein exerting its role in cancer by transcriptionally modulating gene expression and downstream signaling pathways. However, the discovery of an extracellular HMGA1-RAGE autocrine loop in invasive triple-negative breast cancer (TNBC) cell lines implicates HMGA1 as a “moonlighting protein” with different functions depending upon cellular location. Here, we review the role of HMGA1, not only as a chromatin regulator in cancer and stem cells, but also as a potential secreted factor that drives tumor progression. Prior work found that HMGA1 is secreted from TNBC cell lines where it signals through the receptor for advanced glycation end products (RAGE) to foster phenotypes involved in tumor invasion and metastatic progression. Studies in primary TNBC tumors also suggest that HMGA1 secretion associates with distant metastasis in TNBC. Given the therapeutic potential to target extracellular proteins, further work to confirm this role in other contexts is warranted. Indeed, crosstalk between nuclear and secreted HMGA1 could change our understanding of tumor development and reveal novel therapeutic opportunities relevant to diverse human cancers overexpressing HMGA1.
{kind=link}
{kind=link}
{kind=link}
1. Introduction
The diverse functions orchestrated by a cell depend on the action of thousands of proteins working in different cellular compartments at a given time. Until recently, it was thought that most proteins perform one function within a single cellular compartment. However, emerging evidence reveals that an increasing number of proteins break this “one protein-one compartment rule”, thus amplifying the impact that a single protein has on cell fate and behavior [1,2]. Indeed, the term “moonlighting proteins” was established to describe proteins derived from a single gene that perform multiple functions [3]. For example, an enzyme completes a chemical modification based on its specific catalytic site while it performs a different function through protein–protein interactions. A recent study in yeast highlights this concept by revealing that catalytically inactive enzymes can rescue growth. Because the enzymes were engineered to inactivate their catalytic function, these results unveil unexpected functions that are distinct from the enzymatic activity [4].
Another class of moonlighting proteins include multi-compartmental proteins that function in a well-established, so-called “canonical compartment” in addition to a previously unknown, “non-canonical compartment” [1,2,5,6]. Shuttling between such compartments could immerse the protein in distinct local environments (pH, redox, cofactors, PTMs) and thereby allow the protein to function differently in each space. Subcellular localization of proteins was presumed to be dictated primarily by the presence of signal peptides that mediate transfer or tropism toward a cellular organelle. Surprisingly, however, the advent of improved technologies for genome-wide proteomics reveal proteins in unexpected locations [2]. The special case of moonlighting proteins with both intra- and extracellular functions indicates that they are secreted proteins, which could include unconventional pathways that allow for their secretion [5,6,7,8]. While many secreted proteins harbor a characteristic N-terminal sequence that allows for entry and secretion via the endoplasmic reticulum (ER)-Golgi apparatus, novel cellular pathways for secreting proteins have been recently discovered, as detailed next [9].
Unexpected but exciting results from the laboratory of one of the authors (JV) recently uncovered evidence that the High Mobility Group A1 (HMGA1) chromatin regulator is also secreted from aggressive breast cancer cell lines when cultured in vitro [10]. In this review, we will consider the implications of HMGA1 working in different compartments during tumorigenesis. We will also discuss how the interplay between the nuclear and extracellular functions of HMGA1 could impact cancer biology and possibly normal development.
2. Unconventional Protein Secretion (UPS)
Secreted proteins typically contain an N-terminal signal peptide that allows for translocation into the ER. After protein folding, these proteins leave the ER encapsulated in coat protein complex II (COPII)-coated vesicles that link them to the Golgi apparatus. Once they reach the Golgi apparatus, they are sorted and re-encapsulated in vesicles that eventually fuse with the plasma membrane for delivery of vesicular cargo into the extracellular space [11,12]. However, some proteins lacking the N-terminal signal peptide are capable of secretion through unconventional protein secretion (UPS) pathways [13]. The precise molecular mechanisms underlying these pathways are only beginning to emerge, although they appear to be heterogeneous and depend upon the specific secreted protein. Despite these vagaries, a useful classification for such pathways include: (1) direct secretion as soluble proteins, and (2) proteins secreted as extracellular vesicles.
2.1. Unconventional Secretion: UPS Pathways (I–IV)
UPS pathways rely on different cellular mechanisms to facilitate the secretion of proteins while bypassing the ER-Golgi apparatus [9]. Type I and Type II UPS pathways are based on the direct translocation of intracellular proteins across the plasma membrane. Proteins that use Type I UPS are self-translocated through the membrane. The Type I UPS is used for example by the Fibroblast growth factor 2 (FGF2) [14,15,16,17] and the HIV trans-activator of transcription (tat) protein. The HIV-tat protein has dual functions, including enhancing transcription of HIV genes in infected cells within the nucleus, and, upregulating inflammatory genes in uninfected neighboring cells as a secreted signaling factor [18]. By contrast, Type II proteins reach the extracellular compartment through ATP-binding cassette transporter (ABC transporters) which secrete only lipidated peptides and proteins. The yeast pheromone a-factor and Schizosaccharomyces pombe m-factor are both exported by the ABC transporter, Mam1 [19]. Type III UPS proteins first enter into the lumen of intracellular vesicles that ultimately fuse with the plasma membrane releasing their cargoes into the extracellular space. In this case, both lysosomes and autophagosomes are used by Type III proteins for secretion. Notable examples of Type III proteins include two acyl coenzyme A—binding proteins: Acba (Dictyostelium discoideum) [20] and Acb1 (Saccharomyces cerevisiae) [21,22], and interleukin-1β (IL-1β) (Rubartelli et al.). Type III UPS proteins also includes those proteins that are released into the extracellular compartment as vesicles, known as extracellular vesicles (EVs) [23]. Most cells are able to release two types of EVs: exosomes that originate from multivesicular bodies (MVB) [24], and microvesicles generated by exocytosis of the plasma membrane [25]. Finally, a group of transmembrane proteins use the Type IV UPS to reach the plasma membrane bypassing the Golgi apparatus. The best characterized example in this category is the cystic fibrosis transmembrane conductance regulator (CFTR) [26].
2.2. Role of Unconventional Secretion in Cancer Biology
Emerging evidence indicates that unconventional secreted proteins participate in diverse biologic processes. However, most relate to responses to cellular stress. From yeast and parasites to mammalian cells, UPS proteins mediate signals from environmental stimuli. In yeast, the redox enzymes detoxifying superoxide radicals, Superoxide Dismutase 1 (SOD1) and Acb1, are secreted upon nutrient starvation [21]. The bacterial parasite, Dictyostelium discoideum, also secretes Acba, an orthologue of Acb1, upon nutrient starvation to mediate spore formation. The human orthologue of SOD1 is also unconventionally secreted from cells using a mechanism similar to that observed in yeast [27]. Mechanical stress has also been linked to UPS. During the embryogenesis of Drosophila melanogaster, for example, mechanical stress upregulates the gene encoding Golgi Re-Assembly and Stacking Protein (GRASP). GRASP mediates the unconventional secretion of aPS1 integrin subunit, which is required in the establishment of the follicular epithelium of the Drosophila developing oocyte [28]. GRASP also mediates the unconventional secretion of Acba [20], suggesting that it may play a wider role orchestrating unconventional protein secretion pathways.
In human biology, inflammation is a common link between UPS and stress. For example, inflammation triggers the secretion of many downstream mediators and unconventionally secreted proteins, including caspase-1 and IL-1β [29,30]. Caspase 1 enzymatically converts the inactive form of IL-1β, pro-IL-1β, to its active form, IL-1β, and increases in IL-1β are associated with aging and diverse cancers. When pathogens or tissue damage induces inflammatory stress, the pattern recognition receptors, such as Toll-like receptors, and RAGE, become activated, after which pro-IL-1β is processed by caspase 1 and secreted through UPS [31,32]. Caspase 1 is also required for the unconventional secretion of cytoplasmic proteins, including galectins, the macrophage migration inhibitory factor (MIF), and FGF2 [29,33]. Recent work highlighting inflammation in fostering aberrant growth and precancerous lesions in diverse tissues also implicates USP proteins in pre-cancerous neoplasia and progression to frank malignancy. The potential role of UPS in tumor biology is further underscored by recent mass spectrometry analyses of secreted proteins which reveal that cancer cells frequently release factors via UPS pathways [2]. While only a handful of UPS proteins have been validated using orthogonal methods, the role of unconventional secretion is emerging as a critical feature of cancer cells that could be modulated in therapy [34,35]. Since metastatic tumor cells encounter varied stressors when leaving their primary site to establish a tumor in another tissue, this could coordinate of myriad of UPS factors, not only at the primary site, but also within the metastatic niches.
3. High Mobility Group Proteins and Cancer
High mobility group (HMG) proteins are a diverse group of basic proteins which are named for their ability to migrate rapidly (thus high mobility) through polyacrylamide gel by virtue of their low molecular weights [36]. HMG proteins are classified into three families: HMGB, HMGN, and HMGA. All HMG proteins are basic, 10–20 kD proteins which harbor an acidic carboxyl terminus and modulate chromatin structure [37,38,39]. Each family, however, is distinguished by unique DNA and/or nucleosome binding motifs. For example, HMGB family members (HMGB1, HMGB2, HMGB3, HMGB4) are defined by 2 HMG-box motifs that mediate binding to DNA without sequence specificity [39,40,41]. They are the most abundant HMG proteins and were the first HMG proteins to be identified as secreted factors where they function in mediating signals involved in sensing injury, DNA damage, immune activation, inflammation, and cancer. HMGB1 can be passively released from necrotic cells by crossing permeabilized membranes once released from the chromatin [42]. HMGB1 is also actively secreted from damaged cells using an autophagy-dependent secretion (Type III UPS pathway) [40,41]. Indeed, knockdown of key autophagy proteins, including ATG5 and ATG7, abrogates the secretion of HMGB1 [43]. Once in the extracellular space, HMGB1 triggers an inflammatory response. HMGN proteins are found only in vertebrates and include five members: HMGN1, HMGN2, HMGN3, HMGN4, HMGN5 [44]. They lack an HMG box, but contain a positively charged, nucleosome-binding “N” domain that mediates specific binding to nucleosomes [45,46]. HMGN proteins are also be released from cells. For example, extracellular HMGN1 functions as an “alarmin“ or factor that activates the immune [47,48]. In contrast to other HMG proteins, HMGA proteins are defined by three AT-hook motifs that mediate binding to the minor groove of B-form DNA at AT-rich regions. This family includes: (1) HMGA1a/HMGA1b isoforms, encoded by the HMGA1 gene (human chr 6p21) through alternatively spliced mRNA; (2) HMGA1c, the product of a rare splice variant found in testes; (3) HMGA2, a protein highly homologous to HMGA1, but encoded by the HMGA2 gene (chr 12q15) [49,50]. HMGA proteins lack an HMG box and appear to bind to DNA with sequence specifically [51,52].
HMGA proteins are abundant during embryogenesis, but absent or barely detectable postnatally in most differentiated, mature tissues [53]. In embryonic stem cells, HMGA1 induces transcriptional networks involved in self-renewal and pluripotency [54]. HMGA1 becomes aberrantly re-expressed in most aggressive human tumors, where high levels portend poor differentiation status and adverse clinical outcomes [55,56,57,58]. Intriguingly, HMGA1 proteins are among a 13 gene signature encoding transcriptional regulators in embryonic stem cells [59]. Importantly, this signature portends adverse outcomes in brain, bladder, and breast cancers. When overexpressed in lymphoid cells of transgenic mice, Hmga1 induces aggressive leukemia by up-regulating transcriptional networks active in rapidly proliferating stem cells, poorly differentiated cancer cells, and inflammation [60,61]. While mechanisms driving HMGA1 expression in cancer are incompletely understood, growth factors, cancer-associated mutations, including Kras or mutant Apc, and oncogenic transcription factors, such as cMYC, up-regulate HMGA1 in specific contexts, demonstrating that diverse oncogenic pathways converge on HMGA1 to induce its expression [62,63,64,65,66,67]. In intestinal stem cells, HMGA1 also amplifies Wnt signals from the stroma and epithelial niches by inducing expression of genes encoding both Wnt agonist receptors and Wnt effectors, such as cMyc and Sox9 [68]. Together, these findings suggest that HMGA1 could foster tumor progression through both cell-intrinsic and cell-extrinsic or stromal interactions. As presented next, recent work suggests that HMGA1 is secreted via UPS and could thereby drive tumorigenesis and stem cell properties via direct interactions with the stroma.
4. Extracellular Oncogenic Role of HMGA1
Until recently, HMGA1 was assumed to be a nuclear protein exerting its role in cancer by transcriptionally modulating different signaling pathways. However, the laboratory of one of the authors (JV) recently uncovered exciting evidence implicating HMGA1 in extracellular oncogenic function in triple-negative breast cancer (TNBC) cell lines [10]. The secreted form of HMGA1 was mechanistically linked to its role in tumor invasion and metastasis since blocking the extracellular HMGA1 reduced the metastatic burden in a xenograft model of TNBC. This same study also found that an extracellular localization of HMGA1 in human TNBC primary tumors is associated with the presence of distant metastasis in TNBC patients.
4.1. HMGA1 Secretion and Casein Kinase 2 (CK2)
The Villanueva laboratory discovered that HMGA1 is secreted in a regulated fashion from an invasive TNBC breast cancer cell line (MD-MBA-231) [10]. While secretion did not appear to be mediated by extracellular vesicles, the precise UPS pathway employed in this setting remains unknown. However, CK2 was implicated in HMGA1 secretion in these studies. CK2 is known to phosphorylate three serine residues (99, 102, and 103) at the C-terminal region of HMGA1 [69]. In invasive TNBC cells, either inhibiting CK2 or mutating the three HMGA1 serines phosphorylated by CK2 resulting in decreased HMGA1 secretion [10]. Notably, CK2 is linked to the unconventional secretion of other intracellular proteins, including the HMGB1 protein [70]. This study also showed that inhibiting CK2 pharmacologically disrupts invasion in a TNBC cell line. Further, mutation of the three serines (99, 102, 103) in the HMGA1 C-terminus disrupts invasive properties of the TNBC cell line. Intriguingly, CK2 inhibitors were also found to decrease tumor cell invasion in glioblastoma cell lines (GBM) [71,72,73,74]. A recent study using organotypic GBM experimental models also found that the transcriptional repressor interferon regulatory factor 3 (IRF3) decreases invasion in GBM cells [71]. Because CK2 negatively regulates IRF3, inhibiting CK2 up-regulates IRF3, and suppresses invasiveness in GBM cells [71]. Another study showed that either silencing HMGA1 expression or mutation of a CK2 phosphorylation site—Serine 102—of HMGA1 enhanced efficacy of gefitinib in resistant NSCLC cells through reactivation of the downstream signaling of EGFR [75]. However, it remains unknown whether extracellular HMGA1 contributed to drug resistance in cells.
4.2. Receptor for Advanced Glycation End Products (RAGE)
Results from the Villanueva study indicate that HMGA1 mediates its extracellular signaling via the receptor for advanced glycation end products (RAGE receptor) by acting as a ligand [10]. This effect is pERK-dependent and results in enhancing migration, invasion, and metastasis. RAGE is a mammalian pattern recognition receptor from the immunoglobulin superfamily, which is highly expressed during embryogenesis, but rarely expressed in adult healthy tissue [76,77]. The exception is the lung, where RAGE is constitutively expressed in alveolar cells. In the lung, RAGE fosters interactions between adjacent cells, alveolar epithelial cells, and the basal membrane [78]. The protein level of RAGE is tightly regulated by the presence of its ligands, which include advanced glycation end-products (AGEs), proteins of the S100 family, fibrillar proteins, such as amyloid-beta, and HMGB1. More recent data suggests that HMGA1 also regulates RAGE activity and protein levels. Because multiple ligands interact with RAGE, which in turn, signals through diverse pathways, the downstream cellular consequences of RAGE activation are complex. Upon ligand binding to the V domain of RAGE, the receptor oligomerizes, which allows the interaction with different intracellular effectors including Diaphanous 1 (mDIA) [77,79,80]. Activation of RAGE modulates diverse signaling pathways, including nuclear factor kappa-light-chain-enhancer of activated B cells (NF-kB), mitogen-activated protein kinase (MAPK), the Janus kinase (JAK)-signal transducer, and activator of transcription (STAT) [77,79].
Despite the broad array of ligands and pathways regulated, most of the cellular effects induced by RAGE are linked to inflammation [80,81]. RAGE is involved in the resolution of acute inflammation in diverse settings, including wound healing, immune adaptative responses, and nerve regeneration [82]. However, sustained activation of RAGE is directly linked to chronic inflammation, and mediates deleterious effects in common aging-associated diseases such as diabetes, neurodegenerative diseases, and arthritis [77,80]. Although RAGE has not been extensively studied in cancer, many RAGE ligands, such as S100 proteins, are known to foster tumor growth and metastatic progression [10,83,84,85]. Furthermore, increasing evidence underscores the role of chronic inflammation tumor initiation beyond the intestinal tract and colon [86]. Together, these findings implicate RAGE in cancer and suggest that an HMGA1-RAGE autocrine loop could contribute to tumor progression (Figure 1). This is particularly exciting since receptors and secreted factors can be targeted in therapy more readily than most transcription factors.
5. Reconciling HMGA1 Functions across Compartments
The existence of an active, secreted HMGA1 fundamentally changes the current paradigm for HMGA1 function in tumorigenesis. Given the promise of targeting secreted factors, this intriguing discovery warrants further research. Based on existing knowledge, we discuss the potential consequences of cross-talk between the nuclear and extracellular HMGA1 in cancer biology.
5.1. Integrating the Inflammatory Response
HMGA1 is linked to inflammatory processes in diverse contexts [61]. In diverse solid tumors and hematologic malignancy, HMGA1 directly induces expression of genes involved in mediating inflammatory processes. For example, as published by one of the authors (LR) and many others, HMGA1 directly induces expression not only of inflammatory mediators, such as interferons, cytokines, and chemokines, but also of receptors that activate inflammatory responses. HMGA1 was first shown to induce expression of Interferon-β (IFNβ) by recruiting an enhanceosome to key regulatory regions upstream of the coding region, and other cytokines and inflammatory mediators were subsequently linked to HMGA1 transactivation. In Hmga1 transgenic mice, for example, Hmga1 directly induces the Signal Transduction and Activator of Transcription 3 (STAT3) gene with leukemic transformation [87]. STAT3 functions in inflammation, angiogenesis, and tumor progression, and both HMGA1 and STAT3 are up-regulated and co-expressed in hematologic malignancy [88]. Of note, JAK inhibitors are used clinically in hematologic malignancy, although it is not clear whether they alter HMGA1 function [89]. Gene expression studies from the transgenic lymphoid tumor model and other models revealed diverse genes involved in inflammation.
The female Hmga1 transgenic mice also develop uterine tumors, in part, by inducing the gene encoding cyclo-oxygenase-2 (COX2). Indeed, COX2 inhibitors (COXibs) repressed uterine tumor growth in the Hmga1 females and in xenografts of human uterine sarcomas in immunosuppressed mice [90]. HMGA1 and COX2 are also up-regulated and co-expressed in human uterine sarcomas (leiomyosarcomas) [91] in addition to pancreatic tumors [92]. Like STAT3, COX2 is a pleiotropic factor involved in tumor initiation, progression, and inflammatory signaling [49] (Figure 1) and COX-2 inhibitors decrease the incidence of gastrointestinal tumors in mice and humans as well as a subset of tumors arising in other tissues. Intriguingly, HMGA1 was first linked to COX2 expression in vascular endothelium in the setting of hypoxia [93] suggesting that this pathway may function in diverse settings.
Inflammatory mediators also induce HMGA1 expression, such as viral infection, or lipopolysaccharide (LPS), which also activate other inflammatory cytokines. HMGA1 could therefore serve as a hub and feed-forward loop to enforce HMGA1 overexpression and amplify inflammatory signals. Following viral infection or LPS, for example, HMGA1 orchestrates the assembly of NF-κB subunits, p50 and p65, to the IFNβ promoter, inducing IFNβ expression and secretion [61]. Intriguingly, many regulatory regions of HMGA1 transcriptional targets harbor NF-κB binding sites, suggesting that these factors frequently collaborate to modulate gene expression. Because NF-κB is a well-established regulator of inflammation, this interaction further links HMGA1 to inflammatory signals. HMGA1 also modulates the inflammatory response following acute lung injury by facilitating the binding of NF-κB and TNF-α to regulatory elements of the E-selectin and P-selectin genes [94]. Selectins are adhesion molecules that promote transmigration of leukocytes across the endothelial surface into tissues. Studies with selectin genes showed that blocking DNA binding by HMGA1 disrupts recruitment of NF-κB to selectin promoters, and thereby decreases the inflammatory response. Based on the link between inflammation and cancer, HMGA1- NF-κB complexes, also known as the HMGA1 enhanceosome, are likely to contribute neoplastic transformation [61].
While intranuclear HMGA1 contributes to inflammation by modulating expression of cytokines, chemokines, and their receptors, the existence of extracellular-HMGA1 (eHMGA1) opens many avenues for further HMGA1 interactions that could drive both inflammation and cancer. Since eHMGA1 functions through RAGE, and RAGE signals through NF-κB, this provides yet another mechanism whereby HMGA1 could amplify NF-kB inflammatory cytokines. While it is not yet clear whether eHMGA1 occurs more broadly in human cancer and activates NF-κB through RAGE, it should be studied further given the immense clinical implications.
5.2. Epithelial-to-Mesenchymal Plasticity (EMP)
HMGA1 also regulates genes involved in plasticity and an epithelial-mesenchymal transitions. Since EMT is not a binary (E/M) process but involves multiple partial states that give rise to a high degree of cell state plasticity, Epithelial-Mesenchymal plasticity (EMP) [95] is a more accurate definition of the process previously known as EMT. From development to wound healing and acute inflammation, EMP is essential for life [96]. However, during tumor initiation, progression, and cancer therapy, tumor cells hijack EMP networks to thrive [97]. Prior studies found that HMGA1 regulates mesenchymal genes in colon cancer cells, such as Vimentin (VIM), N-cadherin (CDH2), E-cadherin (CDH1), and Fibronectin (FN1) [98,99,100,101]. This is of interest, since both nuclear and eHMGA1 could collaborate in promoting epithelial-to-mesenchymal plasticity (EMP). Prior work from diverse models, including TNBC, indicate that tumor cells hijack EMP networks to thrive during tumor initiation, progression, and cancer therapy [97]. TNBC cells, where extracellular HMGA1 was first described, are characterized by a de-differentiated, mesenchymal like state, which appears to foster tumor initiation and progression. Indeed, those tumor cells with more mesenchymal features frequently localize to the tumor invasive front [102,103]. Thus, regulators of EMP genes appear to play fundamental roles in tumor progression and development and both intranuclear and eHMGA1 could function in these pathways.
In addition to cancer, EMP plays a key role in embryogenesis. During embryonic development, cellular plasticity is required to generate the specialized cell types that will constitute tissues and organs. Morphogenesis occurring during embryogenesis relies on the dynamic interconversion between epithelial and mesenchymal cells, i.e., EMP. HMGA1 is widely expressed during embryogenesis in different tissues, and its loss causes developmental abnormalities. For example, a recent paper found that HMGA1 is required for neural crest formation [104]. Neural crest cells are a population of cells from the neuroectoderm, originally derived from the embryonic ectoderm, which undergo EMP as they acquire mesenchymal properties. These cells are highly migratory and, after delamination, they migrate and generate different specific cell types [105]. HMGA1 is first involved in the specification of the neural plate border through the action of Pax7. Second, HMGA1 modulates Wnt signaling pathways and the Slug transcription factor to foster cranial neural crest emigration and migration. Postnatally, neural crest-derived cells can undergo oncogenic transformation, and not surprisingly, HMGA1 is overexpressed in at least a subset of tumors derived from neural crest cells. Thus, HMGA1 may induce EMP signals in a tightly regulated fashion during development, while this process becomes de-regulated during tumorigenesis.
Because HMGA1 is linked to EMP in cancer and development, it is plausible that crosstalk between the nuclear and the extracellular forms of HMGA1 occur with plasticity. One potential multitasking protein, NUMB [106], could also participate in maintaining plasticity together with HMGA1. NUMB is expressed by neural progenitor cells, neuroblasts, and stem cells where it is asymmetrically segregated in cells. This asymmetry allows the daughter cells containing Numb to acquire a different cell state than the daughter cells lacking it [107]. A recent study also found that HMGA1 downregulates NUMB in brain tumor stem cells, which could maintain the tumor cells in a more de-differentiated, stem-like state and contribute to tumor progression. Another recent study showed that the mice deficient in RAGE develop defective muscle regeneration due to defective myoblast cell division [108]. These correlative findings suggest that NUMB, RAGE, nuclear HMGA1, and extracellular HMGA1 could cooperate in regulating stem cell fate and plasticity.
Because RAGE may mediate crosstalk between nuclear and eHMGA1 with inflammation, this interaction could also participate in EMP. Indeed, RAGE is linked to EMP in tumor cells originating from different tissues. In breast and lung cancer cells, for example, lysophosphatidic acid (LPA) drives EMP through its interaction with RAGE [109]. Another study implicated RAGE and the NF-kB pathway in EMP in breast cancer [110]. In this study, miR-185-5p was overexpressed and served to repress RAGE, thereby reversing EMP properties in MDA-MB-231 cells [111]. In the prostate cancer cell line, PC-3, silencing RAGE also repressed EMT markers, including matrix metalloproteases, disrupting cell migration and invasion [112]. RAGE is also involved in fibrotic diseases marked by aberrant EMP, such as nasal polyp formation [113]. Another recent study showed that the HMGB1-RAGE signaling mediates EMP in upper airway epithelial cells [114]. In the renal tubular cells, another study found that release of HMGB1 up-regulated connective tissue growth factor (CTGF) and transforming growth factor β (TGFβ) to drive renal fibrosis in a RAGE-dependent fashion [115]. Thus, RAGE functions not only in development, inflammation and cancer, but also in fibrosis.
6. Conclusions and Future Directions
The discovery of an extracellular HMGA1-RAGE autocrine loop in invasive TNBC cell lines implicates HMGA1 as a moonlighting protein with different functions depending upon cellular location. Prior studies of HMGA1 in cancer and development focused on its role in the nucleus as a regulator of chromatin structure and function. Indeed, HMGA1 proteins have been described as architectural transcription factors for this reason. Because HMGA1 is overexpressed in most aggressive tumors where it functions in tumor progression, further study of its potential roles in the extracellular space promise to provide important insight relevant to diverse tumors. The unconventional secretion of HMGA1 warrants further study since this location provides an exciting avenue for therapeutic targeting. If secreted HMGA1 is a common feature in settings beyond TNBC cells, this could have important implications in cancer biology [35].
Key research questions that remain unanswered include the following (see Figure 2):
- (1)
- What factors dictate the location of HMGA1?
- (2)
- Are posttranslational modifications involved (similar to those described for HMGB1)?
- (3)
- What triggers secretion together with CK2 or independent of CK2?
- (4)
- Which UPS pathway leads to HMGA1 secretion?
- (5)
- Do receptors other than RAGE mediate eHMGA1 function?
- (6)
- How does intranuclear HMGA1 collaborate with eHMGA1?
- (7)
- Does eHMGA1 function in embryonic development
- (8)
- Is HMGA1 secreted in vivo?
- (9)
- Do diverse tumors secrete HMGA1?
In closing, the prospect that HMGA1 functions in the extracellular space by signaling through RAGE and possibly other cell surface receptors opens many exciting avenues for both future study and therapeutic targeting. Further elucidation of secreted proteins and their function in noncanonical settings should provide insight relevant to normal growth and development in addition to what goes awry with neoplastic transformation and tumor progression. Similar to real estate investments, a key feature of moonlighting protein function is location, location, location!
Author Contributions
Conceptualization, L.R. and J.V.; writing original draft, M.P., L.R., and J.V.; writing, review, editing, and literature search, M.P., L.R., and J.V.; supervision, L.R. and J.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by a grant from Instituto de Salud Carlos III through the project “PI19/01292” (Co-funded by European Regional Development Fund/European Social Fund “A way to make Europe”/“Investing in your future”).
Acknowledgments
We thank Servier for providing design medical art for figures (smart.servier.com). Servier Medical Art by Servier is licensed under a Creative Commons Attribution 3.0 Unported License.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Smalheiser, N.R. Proteins in unexpected locations. Mol. Biol. Cell 1996, 7, 1003–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, G.S.; Overall, C.M. Proteomic identification of multitasking proteins in unexpected locations complicates drug targeting. Nat. Rev. Drug Discov. 2009, 8, 935–948. [Google Scholar] [CrossRef]
- Jobin, X.P.G.; Solis, X.N.; Machado, Y.; Bell, X.P.A.; Rai, S.K.; Kwon, N.H.; Kim, S.; Overall, C.M.; Butler, G.S. Moonlighting matrix metalloproteinase substrates: Enhancement of proinflammatory functions of extracellular tyrosyl-tRNA synthetase upon cleavage. J. Biol. Chem. 2020, 295, 2186–2202. [Google Scholar] [CrossRef] [Green Version]
- Espinosa-cantú, A.; Ascencio, D.; Herrera-basurto, S.; Xu, J.; Roguev, A. Protein Moonlighting Revealed by Noncatalytic Phenotypes of Yeast Enzymes. Genetics 2018, 208, 419–431. [Google Scholar] [CrossRef] [Green Version]
- Arnoys, E.J.; Wang, J.L. Dual localization: Proteins in extracellular and intracellular compartments. Acta Histochem. 2007, 109, 89–110. [Google Scholar] [CrossRef]
- Eustace, B.K.; Sakurai, T.; Stewart, J.K.; Yimlamai, D.; Unger, C.; Zehetmeier, C.; Lain, B.; Torella, C.; Henning, S.W.; Beste, G.; et al. Functional proteomic screens reveal an essential extracellular role for hsp90α in cancer cell invasiveness. Nat. Cell Biol. 2004, 6, 507–514. [Google Scholar] [CrossRef]
- Greenberg, Y.; King, M.; Kiosses, W.; Ewalt, K.; Yang, X.; Schimmel, P.; Reader, J.; Tzima, E. The novel fragment of tyrosyl tRNA synthetase, mini-TyrRS, is secreted to induce an angiogenic response in endothelial cells. FASEB J. 2008, 22, 1597–1605. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Kim, H.; Min, Y.; Choi, E.; Shin, Y.; Park, B.; Lee, S.; Kim, S. Human lysyl-tRNA synthetase is secreted to trigger proinflammatory response. Proc. Natl. Acad. Sci. USA 2005, 102, 6356–6361. [Google Scholar] [CrossRef] [Green Version]
- Rabouille, C.; Malhotra, V.; Nickel, W. Diversity in unconventional protein secretion. J. Cell Sci. 2012, 125, 5251–5255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méndez, O.; Peg, V.; Pujals, M.; Fern, Y.; Matres, A.; Valeri, M.; Gregori, J.; Villarreal, L.; Arribas, J.; Villanueva, J. Extracellular HMGA1 Promotes Tumor Invasion and Metastasis in Triple-Negative Breast Cancer. Clin. Cancer Res. 2018, 24, 6367–6382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osborne, A.; Rapoport, T.; van den Berg, B. Protein translocation by the Sec61/SecY channel. Annu. Rev. Cell Dev. Biol. 2005, 21, 529–550. [Google Scholar] [CrossRef]
- Lee, M.C.S.; Miller, E.A.; Goldberg, J.; Orci, L.; Schekman, R. Bi-directional protein transport between the ER and Golgi. Annu. Rev. Cell Dev. Biol. 2004, 20, 87–123. [Google Scholar] [CrossRef] [Green Version]
- Nickel, W.; Seedorf, M. Unconventional mechanisms of protein transport to the cell surface of eukaryotic cells. Annu. Rev. Cell Dev. Biol. 2008, 24, 287–308. [Google Scholar] [CrossRef] [PubMed]
- Zehe, C.; Engling, A.; Wegehingel, S.; Schäfer, T.; Nickel, W. Cell-surface heparan sulfate proteoglycans are essential components of the unconventional export machinery of FGF-2. Proc. Natl. Acad. Sci. USA 2006, 103, 15479–15484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nickel, W. The unconventional secretory machinery of fibroblast growth factor 2. Traffic 2011, 12, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, T.; Zentgraf, H.; Zehe, C.; Brügger, B.; Bernhagen, J.; Nickel, W. Unconventional Secretion of Fibroblast Growth Factor 2 Is Mediated by Direct Translocation across the Plasma Membrane of Mammalian Cells. J. Biol. Chem. 2004, 279, 6244–6251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backhaus, R.; Zehe, C.; Wegehingel, S.; Kehlenbach, A.; Schwappach, B.; Nickel, W. Unconventional protein secretion: Membrane translocation of FGF-2 does not require protein unfolding. J. Cell Sci. 2004, 117, 1727–1736. [Google Scholar] [CrossRef] [Green Version]
- Ajasin, D.; Eugenin, E.A. HIV-1 Tat: Role in Bystander Toxicity. Front. Cell. Infect. Microbiol. 2020, 10. [Google Scholar] [CrossRef]
- Christensen, P.U.; Davey, J.; Nielsen, O. The Schizosaccharomyces pombe mam1 gene encodes an ABC transporter mediating secretion of M-factor. Mol. Gen. Genet. MGG 1997, 255, 226–236. [Google Scholar] [CrossRef]
- Cabral, M.; Anjard, C.; Malhotra, V.; Loomis, W.F.; Kuspa, A. Unconventional secretion of AcbA in Dictyostelium discoideum through a vesicular intermediate. Eukaryot. Cell 2010, 9, 1009–1017. [Google Scholar] [CrossRef] [Green Version]
- Duran, J.; Anjard, C.; Stefan, C.; Loomis, W.; Malhotra, V. Unconventional secretion of Acb1 is mediated by autophagosomes. J. Cell Biol. 2010, 188, 527–536. [Google Scholar] [CrossRef] [Green Version]
- Manjithaya, R.; Anjard, C.; Loomis, W.F.; Subramani, S. Unconventional secretion of Pichia pastoris Acb1 is dependent on GRASP protein, peroxisomal functions, and autophagosome formation. J. Cell Biol. 2010, 188, 537–546. [Google Scholar] [CrossRef] [Green Version]
- Stahl, P.D.; Raposo, G. Extracellular Vesicles: Exosomes and Microvesicles, Integrators of Homeostasis. Physiology 2019, 34, 169–177. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef] [PubMed]
- Souza-schorey, C.D.; Clancy, J.W. Tumor-derived microvesicles: Shedding light on novel microenvironment modulators and prospective cancer biomarkers. Genes Dev. 2012, 26, 1287–1299. [Google Scholar] [CrossRef] [Green Version]
- Yoo, J.S.; Moyer, B.D.; Bannykh, S.; Yoo, H.M.; Riordan, J.R.; Balch, W.E. Non-conventional Trafficking of the Cystic Fibrosis Transmembrane Conductance Regulator through the Early Secretory Pathway. J. Biol. Chem. 2002, 277, 11401–11409. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Garcia, D.; Brouwers, N.; Duran, J.M.; Mora, G.; Curwin, A.J.; Malhotra, V. A diacidic motif determines unconventional secretion of wild-type and ALS-linked mutant SOD1. J. Cell Biol. 2017, 216, 2691–2700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giuliani, F.; Grieve, A.; Rabouille, C. Unconventional secretion: A stress on GRASP. Curr. Opin. Cell Biol. 2011, 23, 498–504. [Google Scholar] [CrossRef]
- Nickel, W.; Rabouille, C. Mechanisms of regulated unconventional protein secretion. Nat. Rev. Mol. Cell Biol. 2008, 10, 148–155. [Google Scholar] [CrossRef]
- Ogura, Y.; Sutterwala, F.S.; Flavell, R.A. The Inflammasome: First Line of the Immune Response to Cell Stress. Cell 2006, 126, 659–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardaway, A.L.; Podgorski, I. IL-1β, RAGE and FABP4: Targeting the dynamic trio in metabolic inflammation and related pathologies. Future Med. Chem. 2013, 5, 1089–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Q.; You, H.; Li, X.; Liu, T.; Wang, P.; Wang, B. HMGB1 promotes the synthesis of pro-IL-1β and pro-IL-18 by activation of p38 MAPK and NF-κB through receptors for advanced glycation end-products in macrophages. Asian Pac. J. Cancer Prev. 2012, 13, 1365–1370. [Google Scholar] [CrossRef] [Green Version]
- Keller, M.; Ru, A.; Werner, S.; Beer, H. Active Caspase-1 Is a Regulator of Unconventional Protein Secretion. Cell 2008, 132, 818–831. [Google Scholar] [CrossRef] [Green Version]
- Jiyoon, K.; Heon Yung, G.; Min Goo, L. Unconventional protein secretion – new insights into the pathogenesis and therapeutic targets of human diseases. J. Cell Sci. 2018, 131, jcs213686. [Google Scholar] [CrossRef] [Green Version]
- Mendez, O.; Pérez, J.; Soberino, J.; Racca, F.; Cortes, J.; Villanueva, J. Clinical Implications of Extracellular HMGA1 in Breast Cancer. Int. J. Mol. Sci. 2019, 20, 5950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodwin, G.H.; Sanders, C.; Johns, E.W. A New Group of Chromatin-Associated Proteins with a High Content of Acidic and Basic Amino Acids. Eur. J. Biochem. 1973, 38, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Reeves, R. Nuclear functions of the HMG proteins. Biochim. Biophys. Acta - Gene Regul. Mech. 2010, 1799, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Hock, R.; Furusawa, T.; Ueda, T.; Bustin, M. HMG chromosomal proteins in development and disease. Trends Cell Biol. 2007, 17, 72–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerlitz, G.; Hock, R.; Ueda, T.; Bustin, M. The dynamics of HMG protein-chromatin interactions in living cells. Biochem. Cell Biol. 2009, 87, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.H.; Kwak, M.S.; Lee, B.; Shin, J.M.; Aum, S.; Park, I.H.; Lee, M.G.; Shin, J.S. Secretory autophagy machinery and vesicular trafficking are involved in HMGB1 secretion. Autophagy 2020. [Google Scholar] [CrossRef]
- Wang, Z.; Zhou, H.; Zheng, H.; Zhou, X.; Shen, G.; Teng, X.; Liu, X.; Zhang, J.; Wei, X.; Hu, Z.; et al. Autophagy-based unconventional secretion of HMGB1 by keratinocytes plays a pivotal role in psoriatic skin inflammation. Autophagy 2021, 17, 529–552. [Google Scholar] [CrossRef] [PubMed]
- Scaffidi, P.; Misteli, T.; Bianchi, M.E. Release of chromatin protein HMGB1 by necrotic cells triggers inflammation. Nature 2002, 418, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Thorburn, J.; Horita, H.; Redzic, J.; Hansen, K.; Frankel, A.E.; Thorburn, A. Autophagy regulates selective HMGB1 release in tumor cells that are destined to die. Cell Death Differ. 2009, 16, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Postnikov, Y.; Bustin, M. Regulation of chromatin structure and function By HMGN proteins. Biochim. Biophys. Acta - Gene Regul. Mech. 2010, 1799, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Catez, F.; Brown, D.T.; Misteli, T.; Bustin, M. Competition between histone H1 and HMGN proteins for chromatin binding sites. EMBO Rep. 2002, 3, 760–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.H.; West, K.L.; Rubinstein, Y.; Bergel, M.; Postnikov, Y.V.; Bustin, M. Chromosomal protein HMGN1 enhances the acetylation of lysine 14 in histone H3. EMBO J. 2005, 24, 3038–3048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Tewary, P.; de la Rosa, G.; Wei, F.; Oppenheim, J.J. The alarmin functions of high-mobility group proteins. Physiol. Behav. 2010, 1799, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Park, I.A.; Heo, S.-H.; Song, I.H.; Kim, Y.-A.; Park, H.S.; Bang, W.S.; Park, S.Y.; Jo, J.-H.; Lee, H.J.; Gong, G. Endoplasmic reticulum stress induces secretion of high-mobility group proteins and is associated with tumor-infiltrating lymphocytes in triple-negative breast cancer. Oncotarget 2016, 7, 59957–59964. [Google Scholar] [CrossRef] [Green Version]
- Resar, L.M.S. The High Mobility Group A1 Gene: Transforming Inflammatory Signals into Cancer? Cancer Res. 2010, 70, 436–439. [Google Scholar] [CrossRef] [Green Version]
- Fusco, A.; Fedele, M. Roles of HMGA proteins in cancer. Nat. Rev. Cancer 2007, 7, 899–910. [Google Scholar] [CrossRef]
- Banks, G.C.; Mohr, B.; Reeves, R. The HMG-I(Y) A · T-hook peptide motif confers DNA-binding specificity to a structured chimeric protein. J. Biol. Chem. 1999, 274, 16536–16544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geierstanger, B.H.; Volkman, B.F.; Kremer, W.; Wemmer, D.E. Short Peptide Fragments Derived from HMG-I/Y Proteins Bind Specifically to the Minor Groove of DNA. Biochemistry 1994, 33, 5347–5355. [Google Scholar] [CrossRef]
- Chiappetta, G.; Avantaggiato, V.; Visconti, R.; Fedele, M.; Battista, S.; Trapasso, F.; Merciai, B.M.; Fidanza, V.; Giancotti, V.; Santoro, M.; et al. High level expression of the HMGI (Y) gene during embryonic development. Oncogene 1996, 13, 2439–2446. [Google Scholar] [PubMed]
- Shah, S.N.; Kerr, C.; Cope, L.; Zambidis, E.; Liu, C.; Hillion, J.; Belton, A.; Huso, D.L.; Resar, L.M.S. HMGA1 Reprograms Somatic Cells into Pluripotent Stem Cells by Inducing Stem Cell Transcriptional Networks. PLoS ONE 2012, 7, e48533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hristov, A.C.; Cope, L.; Reyes, M.D.; Singh, M.; Iacobuzio-Donahue, C.; Maitra, A.; Resar, L.M.S. HMGA2 protein expression correlates with lymph node metastasis and increased tumor grade in pancreatic ductal adenocarcinoma. Mod. Pathol. 2009, 22, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Di Cello, F.; Kowalski, J.; Hristov, A.C.; Tsai, H.L.; Bhojwani, D.; Meyer, J.A.; Carroll, W.L.; Belton, A.; Resar, L.M.S. HMGA1 overexpression correlates with relapse in childhood B-lineage acute lymphoblastic leukemia. Leuk. Lymphoma 2013, 54, 2565–2567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hristov, A.C.; Cope, L.; Di Cello, F.; Reyes, M.D.; Singh, M.; Hillion, J.A.; Belton, A.; Joseph, B.; Schuldenfrei, A.; Iacobuzio-Donahue, C.A.; et al. HMGA1 correlates with advanced tumor grade and decreased survival in pancreatic ductal adenocarcinoma. Mod. Pathol. 2010, 23, 98–104. [Google Scholar] [CrossRef]
- Gorbounov, M.; Carleton, N.M.; Asch-Kendrick, R.J.; Xian, L.; Rooper, L.; Chia, L.; Cimino-Mathews, A.; Cope, L.; Meeker, A.; Stearns, V.; et al. High mobility group A1 (HMGA1) protein and gene expression correlate with ER-negativity and poor outcomes in breast cancer. Breast Cancer Res. Treat. 2020, 179, 25–35. [Google Scholar] [CrossRef]
- Ben-Porath, I.; Thomson, M.W.; Carey, V.J.; Ge, R.; Bell, G.W.; Regev, A.; Weinberg, R.A. An embryonic stem cell-like gene expression signature in poorly differentiated aggressive human tumors. Nat. Genet. 2008, 40, 499–507. [Google Scholar] [CrossRef]
- Xu, Y.; Sumter, T.F.; Bhattacharya, R.; Tesfaye, A.; Fuchs, E.J.; Wood, L.J.; Huso, D.L.; Resar, L.M.S. The HMG-I oncogene causes highly penetrant, aggressive lymphoid malignancy in transgenic mice and is overexpressed in human leukemia. Cancer Res. 2004, 64, 3371–3375. [Google Scholar] [CrossRef] [Green Version]
- Schuldenfrei, A.; Belton, A.; Kowalski, J.; Talbot, C.C.; Di Cello, F.; Poh, W.; Tsai, H.L.; Shah, S.N.; Huso, T.H.; Huso, D.L.; et al. HMGA1 drives stem cell, inflammatory pathway, and cell cycle progression genes during lymphoid tumorigenesis. BMC Genomics 2011, 12, 549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeves, R.; Beckerbauer, L. HMGI/Y proteins: Flexible regulators of transcription and chromatin structure. Biochim. Biophys. Acta - Gene Struct. Expr. 2001, 1519, 13–29. [Google Scholar] [CrossRef]
- Lanahan, A.; Williams, J.B.; Sanders, L.K.; Nathans, D. Growth factor-induced delayed early response genes. Mol. Cell. Biol. 1992, 12, 3919–3929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holth, L.T.; Thorlacius, A.E.; Reeves, R. Effects of epidermal growth factor and estrogen on the regulation of the HMG-I/Y gene in human mammary epithelial cell lines. DNA Cell Biol. 1997, 16, 1299–1309. [Google Scholar] [CrossRef]
- Cleynen, I.; Huysmans, C.; Sasazuki, T.; Shirasawa, S.; Van De Ven, W.; Peeters, K. Transcriptional control of the human high mobility group A1 gene: Basal and oncogenic Ras-regulated expression. Cancer Res. 2007, 67, 4620–4629. [Google Scholar] [CrossRef] [Green Version]
- Wood, L.J.; Mukherjee, M.; Dolde, C.E.; Xu, Y.; Maher, J.F.; Bunton, T.E.; Williams, J.B.; Resar, L.M.S. HMG-I/Y, a New c-Myc Target Gene and Potential Oncogene. Mol. Cell. Biol. 2000, 20, 5490–5502. [Google Scholar] [CrossRef] [Green Version]
- Giannini, G.; Cerignoli, F.; Mellone, M.; Massimi, I.; Ambrosi, C.; Rinaldi, C.; Dominici, C.; Frati, L.; Screpanti, I.; Gulino, A. High mobility group A1 is a molecular target for MYCN in human neuroblastoma. Cancer Res. 2005, 65, 8308–8316. [Google Scholar] [CrossRef] [Green Version]
- Xian, L.; Georgess, D.; Huso, T.; Cope, L.; Belton, A.; Chang, Y.T.; Kuang, W.; Gu, Q.; Zhang, X.; Senger, S.; et al. HMGA1 amplifies Wnt signalling and expands the intestinal stem cell compartment and Paneth cell niche. Nat. Commun. 2017, 8, 15008. [Google Scholar] [CrossRef] [Green Version]
- Palvimo, J.; Linnala-Kankkunen, A. Identification of sites on chromosomal protein HMG-I phosphorylated by casein kinase II. Elsevier Sci. Publ. 1989, 257, 101–104. [Google Scholar] [CrossRef] [Green Version]
- de Abreu da Silva, I.C.; Carneiro, V.C.; de Moraes Maciel, R.; da Costa, R.F.M.; Furtado, D.R.; de Oliveira, F.M.B.; da Silva-Neto, M.A.C.; Rumjanek, F.D.; Fantappié, M.R. CK2 Phosphorylation of Schistosoma mansoni HMGB1 Protein Regulates Its Cellular Traffic and Secretion but Not Its DNA Transactions. PLoS ONE 2011, 6, e23572. [Google Scholar] [CrossRef]
- Pencheva, N.; de Gooijer, M.C.; Vis, D.J.; Wessels, L.F.A.; Würdinger, T.; van Tellingen, O.; Bernards, R. Identification of a Druggable Pathway Controlling Glioblastoma Invasiveness. Cell Rep. 2017, 20, 48–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitt, B.M.; Boewe, A.S.; Götz, C.; Philipp, S.E.; Urbschat, S.; Oertel, J.; Menger, M.D.; Laschke, M.W.; Ampofo, E. Ck2 activity mediates the aggressive molecular signature of glioblastoma multiforme by inducing nerve/glial antigen (Ng)2 expression. Cancers 2021, 13, 1678. [Google Scholar] [CrossRef] [PubMed]
- Nitta, R.T.; Gholamin, S.; Feroze, A.H.; Agarwal, M.; Cheshier, S.H.; Mitra, S.S.; Li, G. Casein kinase 2α regulates glioblastoma brain tumor-initiating cell growth through the β-catenin pathway. Oncogene 2015 3428 2014, 34, 3688–3699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; McFarland, B.C.; Drygin, D.; Yu, H.; Bellis, S.L.; Kim, H.; Bredel, M.; Benveniste, E.N. Targeting protein kinase CK2 suppresses prosurvival signaling pathways and growth of glioblastoma. Clin. Cancer Res. 2013, 19, 6484–6494. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Pan, S.; Tsai, C.; Kuo, T.; Hsu, Y. Phosphoproteomics Reveals HMGA1, a CK2 Substrate, as a Drug-Resistant Target in Non-Small Cell Lung Cancer. Nat. Publ. Gr. 2017, 7, 44021. [Google Scholar] [CrossRef] [Green Version]
- Sessa, L.; Gatti, E.; Zeni, F.; Antonelli, A.; Catucci, A.; Koch, M.; Pompilio, G.; Fritz, G.; Raucci, A.; Bianchi, M.E. The Receptor for Advanced Glycation End-products (RAGE) is only present in mammals, and belongs to a family of Cell Adhesion Molecules (CAMs). PLoS ONE 2014, 9, e86903. [Google Scholar] [CrossRef]
- Teissier, T.; Boulanger, É. The receptor for advanced glycation end-products (RAGE) is an important pattern recognition receptor (PRR) for inflammaging. Biogerontology 2019, 20, 279–301. [Google Scholar] [CrossRef]
- Demling, N.; Ehrhardt, C.; Kasper, M.; Laue, M.; Knels, L.; Rieber, E.P. Promotion of cell adherence and spreading: A novel function of RAGE, the highly selective differentiation marker of human alveolar epithelial type I cells. Cell Tissue Res. 2006, 323, 475–488. [Google Scholar] [CrossRef]
- Xie, J.; Méndez, J.D.; Méndez-Valenzuela, V.; Aguilar-Hernández, M.M. Cellular signalling of the receptor for advanced glycation end products (RAGE). Cell. Signal. 2013, 25, 2185–2197. [Google Scholar] [CrossRef]
- Hudson, B.I.; Lippman, M.E. Targeting RAGE Signaling in Inflammatory Disease. Annu. Rev. Med. 2018, 69, annurev. [Google Scholar] [CrossRef]
- Sparvero, L.J.; Asafu-Adjei, D.; Kang, R.; Tang, D.; Amin, N.; Im, J.; Rutledge, R.; Lin, B.; Amoscato, A.A.; Zeh, H.J.; et al. RAGE (Receptor for advanced glycation endproducts), RAGE ligands, and their role in cancer and inflammation. J. Transl. Med. 2009, 7, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palanissami, G.; Paul, S.F.D. RAGE and Its Ligands: Molecular Interplay Between Glycation, Inflammation, and Hallmarks of Cancer—A Review. Horm. Cancer 2018, 9, 295–325. [Google Scholar] [CrossRef]
- Chen, M.C.; Chen, K.C.; Chang, G.C.; Lin, H.; Wu, C.C.; Kao, W.H.; Teng, C.L.J.; Hsu, S.L.; Yang, T.Y. RAGE acts as an oncogenic role and promotes the metastasis of human lung cancer. Cell Death Dis. 2020, 11, 265. [Google Scholar] [CrossRef]
- Chen, R.C.; Yi, P.P.; Zhou, R.R.; Xiao, M.F.; Huang, Z.B.; Tang, D.L.; Huang, Y.; Fan, X.G. The role of HMGB1-RAGE axis in migration and invasion of hepatocellular carcinoma cell lines. Mol. Cell. Biochem. 2014, 390, 271–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, T.; Li, X.; Hua, Z.; Ma, J.; Wu, X.; Liu, Z.; Chen, H.; Cui, Z. S100A7 promotes the migration, invasion and metastasis of human cervical cancer cells through epithelial-mesenchymal transition. Oncotarget 2017, 8, 24964–24977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Body-Malapel, M.; Djouina, M.; Waxin, C.; Langlois, A.; Gower-Rousseau, C.; Zerbib, P.; Schmidt, A.M.; Desreumaux, P.; Boulanger, E.; Vignal, C. The RAGE signaling pathway is involved in intestinal inflammation and represents a promising therapeutic target for Inflammatory Bowel Diseases. Mucosal Immunol. 2019, 12, 468–478. [Google Scholar] [CrossRef]
- Resar, L.; Hillion, J.; Dhara, S.; Sumter, T.F.; Mukherjee, M.; Di Cello, F.; Belton, A.; Turkson, J.; Jaganathan, S.; Cheng, L.; et al. The HMGA1a-STAT3 axis: An “Achilles Heel” for Hematopoietic Malignancies Overexpressing HMGA1a? Blood 2008, 112, 3810. [Google Scholar] [CrossRef]
- Hillion, J.; Belton, A.M.; Shah, S.N.; Turkson, J.; Jing, N.; Tweardy, D.J.; Di Cello, F.; Huso, D.L.; Resar, L.M.S. Nanoparticle delivery of inhibitory signal transducer and activator of transcription 3 G-quartet oligonucleotides blocks tumor growth in HMGA1 transgenic model of T-cell leukemia. Leuk. Lymphoma 2014, 55, 1194–1197. [Google Scholar] [CrossRef] [Green Version]
- Arcaini, L.; Cazzola, M. Benefits and risks of JAK inhibition. Blood 2018, 132, 675–676. [Google Scholar] [CrossRef] [Green Version]
- Di Cello, F.; Hillion, J.; Kowalski, J.; Ronnett, B.M.; Aderinto, A.; Huso, D.L.; Resar, L.M.S. Cyclooxygenase inhibitors block uterine tumorigenesis in HMGA1a transgenic mice and human xenografts. Mol. Cancer Ther. 2008, 7, 2090–2095. [Google Scholar] [CrossRef] [Green Version]
- Tesfaye, A.; Di Cello, F.; Hillion, J.; Ronnett, B.M.; Elbahloul, O.; Ashfaq, R.; Dhara, S.; Prochownik, E.; Tworkoski, K.; Reeves, R.; et al. The High-Mobility Group A1 Gene Up-Regulates Cyclooxygenase 2 Expression in Uterine Tumorigenesis. Cancer Res. 2007, 67, 3998–4004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillion, J.; Smail, S.; Di Cello, F. The HMGA1-COX-2 axis: A key molecular pathway and potential target in pancreatic adenocarcinoma. Pancreatology 2012, 12, 372–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, Y.S.; Xu, Q.; Schmedtje, J.F. Hypoxia induces high-mobility-group protein I(Y) and transcription of the cyclooxygenase-2 gene in human vascular endothelium. Circ. Res. 1998, 83, 295–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baron, R.M.; Lopez-Guzman, S.; Riascos, D.F.; Macias, A.A.; Layne, M.D.; Cheng, G.; Harris, C.; Chung, S.W.; Reeves, R.; von Andrian, U.H.; et al. Distamycin A inhibits HMGA1-binding to the P-selectin promoter and attenuates lung and liver inflammation during murine endotoxemia. PLoS ONE 2010, 5, e10656. [Google Scholar] [CrossRef] [Green Version]
- Nieto, M.A.; Huang, R.Y.Y.J.; Jackson, R.A.A.; Thiery, J.P.P. Emt: 2016. Cell 2016, 166, 21–45. [Google Scholar] [CrossRef] [Green Version]
- Nieto, M.A. Epithelial Plasticity: A Common Theme in Embryonic and Cancer Cells. Science 2013, 342. [Google Scholar] [CrossRef] [Green Version]
- Beerling, E.; Seinstra, D.; de Wit, E.; Kester, L.; van der Velden, D.; Maynard, C.; Schäfer, R.; van Diest, P.; Voest, E.; van Oudenaarden, V.; et al. Plasticity between Epithelial and Mesenchymal Report Plasticity between Epithelial and Mesenchymal States Unlinks EMT from Metastasis-Enhancing Stem Cell Capacity. Cell Rep. 2016, 14, 2281–2288. [Google Scholar] [CrossRef] [Green Version]
- Belton, A.; Gabrovsky, A.; Bae, Y.K.; Reeves, R.; Iacobuzio-Donahue, C.; Huso, D.L.; Resar, L.M.S. HMGA1 induces intestinal polyposis in transgenic mice and drives tumor progression and stem cell properties in colon cancer cells. PLoS ONE 2012, 7, e30034. [Google Scholar] [CrossRef] [Green Version]
- Pegoraro, S.; Ros, G.; Piazza, S.R.; Sommaggio, R.; Ciani, Y.; Rosato, A.; Sgarra, R.; Del Sal, G.; Manfioletti, G. HMGA1 promotes metastatic processes in basal-like breast cancer regulating EMT and stemness. Oncotarget 2013, 4, 1293–1308. [Google Scholar] [CrossRef] [Green Version]
- Jin, G.H.; Shi, Y.; Tian, Y.; Cao, T.T.; Mao, Y.; Tang, T.Y. HMGA1 accelerates the malignant progression of gastric cancer through stimulating EMT. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 3642–3647. [Google Scholar] [CrossRef]
- Shah, S.N.; Cope, L.; Poh, W.; Belton, A.; Roy, S. HMGA1: A Master Regulator of Tumor Progression in Triple-Negative Breast Cancer Cells. PLoS ONE 2013, 8, e63419. [Google Scholar] [CrossRef] [Green Version]
- Scheel, C.; Eaton, E.; Li, S.; Chaffer, C.; Reinhardt, F.; Kah, K.; Bell, G.; Guo, W.; Rubin, J.; Richardson, A.L.; et al. Paracrine and autocrine signals induce and maintain mesenchymal and stem cell states in the breast. Cell 2011, 145, 926–940. [Google Scholar] [CrossRef] [Green Version]
- Guen, V.J.; Chavarria, T.E.; Kröger, C.; Ye, X.; Weinberg, R.A.; Lees, J.A. EMT programs promote basal mammary stem cell and tumor-initiating cell stemness by inducing primary ciliogenesis and Hedgehog signaling. Proc. Natl. Acad. Sci. USA 2017, 114, E10532–E10539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandhi, S.; Hutchins, E.J.; Maruszko, K.; Park, J.H.; Thomson, M.; Bronner, M.E. Bimodal function of chromatin remodeler Hmga1 in neural crest induction and Wnt- dependent emigration. Elife 2020, 9, e57779. [Google Scholar] [CrossRef] [PubMed]
- Bronner, M.E.; LeDouarin, N.M. Development and evolution of the neural crest: An overview. Dev. Biol. 2012, 366, 2–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, S.; Johnston, S.; Johnston, S. PARtitioning Numb. EMBO Rep. 2007, 8, 233–235. [Google Scholar] [CrossRef] [Green Version]
- Puca, F.; Tosti, N.; Federico, A.; Kuzay, Y.; Pepe, A.; Morlando, S.; Savarese, T.; D’Alessio, F.; Colamaio, M.; Sarnataro, D.; et al. HMGA1 negatively regulates NUMB expression at transcriptional and post transcriptional level in glioblastoma stem cells. Cell Cycle 2019, 18, 1446–1457. [Google Scholar] [CrossRef] [PubMed]
- Riuzzi, F.; Sorci, G.; Sagheddu, R.; Donato, R. HMGB1-RAGE regulates muscle satellite cell homeostasis through p38-MAPK- and myogenindependent repression of Pax7 transcription. J. Cell Sci. 2012, 125, 1440–1454. [Google Scholar] [CrossRef] [Green Version]
- Rashmi, R.; Nitish, J.; Satyendra Kumar, S.; Sunita, S.; Vivek, R. Lysophosphatidic acid-RAGE axis promotes lung and mammary oncogenesis via protein kinase B and regulating tumor microenvironment. Cell Commun. Signal. 2020, 18, 170. [Google Scholar] [CrossRef]
- Yin, C.; Li, H.; Zhang, B.; Liu, Y.; Lu, G.; Lu, S.; Sun, L.; Qi, Y.; Li, X.; Chen, W. RAGE-binding S100A8/A9 promotes the migration and invasion of human breast cancer cells through actin polymerization and epithelial-mesenchymal transition. Breast Cancer Res. Treat. 2013, 142, 297–309. [Google Scholar] [CrossRef]
- Yin, C.; Zhang, G.; Sun, R.; Pan, X.; Wang, X.; Li, H.; Sun, Y. miR-185-5p inhibits F-actin polymerization and reverses epithelial mesenchymal transition of human breast cancer cells by modulating RAGE. Mol. Med. Rep. 2018, 2621–2630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Shao, S.; Han, D.; Xu, Y.; Jiao, D.; Wu, J.; Yang, F.; Ge, Y.; Shi, S.; Li, Y.; et al. High mobility group box 1 promotes the epithelial-to-mesenchymal transition in prostate cancer PC3 cells via the RAGE/NF-KB signaling pathway. Int. J. Oncol. 2018, 53, 659–671. [Google Scholar] [CrossRef] [PubMed]
- Könnecke, M.; Burmeister, M.; Pries, R.; Böscke, R.; Bruchhage, K.; Ungefroren, H.; Klimek, L.; Wollenberg, B. Epithelial-Mesenchymal Transition in Chronic Rhinosinusitis: Differences Revealed Between Epithelial Cells from Nasal Polyps and Inferior Turbinates. Arch. Immunol. Ther. Exp. (Warsz). 2017, 65, 157–173. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-C.; Statt, S.; Wu, R.; Chang, H.-T.; Liao, J.-W.; Wang, C.-N.; Shyu, W.-C.; Lee, C.-C. High mobility group box 1-induced epithelial mesenchymal transition in human airway epithelial cells. Sci. Rep. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, M.; Liu, H.; Zhang, D.; Liu, Y.; Wang, C.; Liu, F.; Chen, J. HMGB1 Enhances the AGE-Induced Expression of CTGF and TGF-β via RAGE-Dependent Signaling in Renal Tubular Epithelial Cells. Am. J. Nephrol. 2015, 41, 257–266. [Google Scholar] [CrossRef]
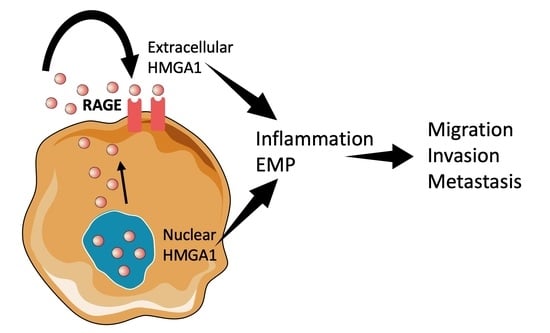
Figure 1.
HMGA1 was recently discovered in different cellular compartments during tumorigenesis. The scheme illustrates known molecular mechanisms that mediate HMGA1 function in cancer.
Figure 1.
HMGA1 was recently discovered in different cellular compartments during tumorigenesis. The scheme illustrates known molecular mechanisms that mediate HMGA1 function in cancer.

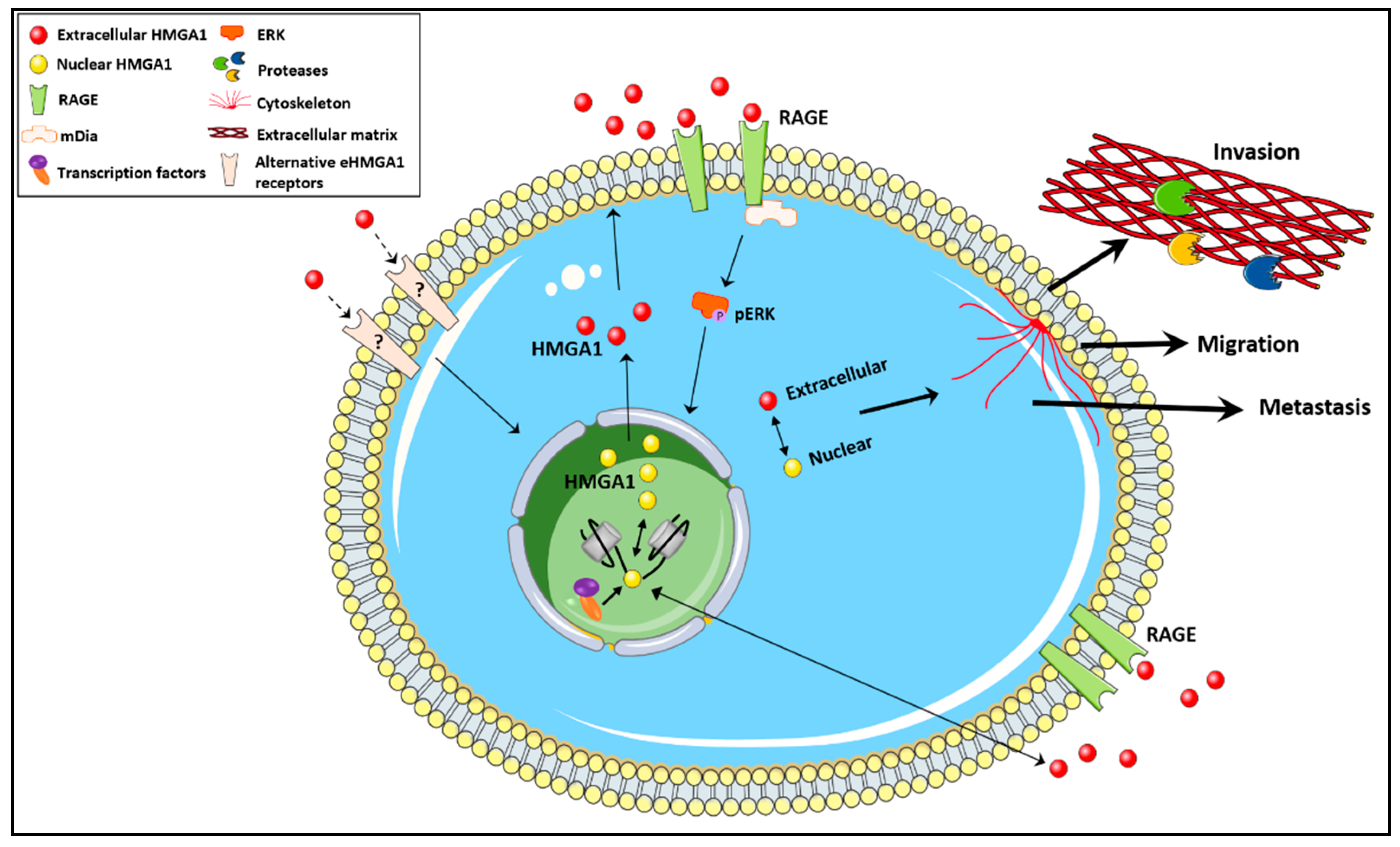
Figure 2.
The unconventional secretion of HMGA1 warrants further studies to understand the role of extracellular HMGA1 in cancer biology. Future research should answer questions related to: the molecular mechanism driving the subcellular location of HMGA1, the factors controlling HMGA1 secretion, whether eHMGA1 mediates an oncogenic function by binding to receptors other than RAGE, whether there is a cross-talk between the nuclear and extracellular HMGA1, and how the equilibrium between the two forms of HMGA1 dictates the role of HMGA1 in cancer biology.
Figure 2.
The unconventional secretion of HMGA1 warrants further studies to understand the role of extracellular HMGA1 in cancer biology. Future research should answer questions related to: the molecular mechanism driving the subcellular location of HMGA1, the factors controlling HMGA1 secretion, whether eHMGA1 mediates an oncogenic function by binding to receptors other than RAGE, whether there is a cross-talk between the nuclear and extracellular HMGA1, and how the equilibrium between the two forms of HMGA1 dictates the role of HMGA1 in cancer biology.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pujals, M.; Resar, L.; Villanueva, J. HMGA1, Moonlighting Protein Function, and Cellular Real Estate: Location, Location, Location! Biomolecules 2021, 11, 1334. https://doi.org/10.3390/biom11091334
AMA Style
Pujals M, Resar L, Villanueva J. HMGA1, Moonlighting Protein Function, and Cellular Real Estate: Location, Location, Location! Biomolecules. 2021; 11(9):1334. https://doi.org/10.3390/biom11091334
Chicago/Turabian StylePujals, Mireia, Linda Resar, and Josep Villanueva. 2021. "HMGA1, Moonlighting Protein Function, and Cellular Real Estate: Location, Location, Location!" Biomolecules 11, no. 9: 1334. https://doi.org/10.3390/biom11091334
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.