
Diallyl Sulfide Attenuation of Carcinogenesis in Mammary Epithelial Cells through the Inhibition of ROS Formation, and DNA Strand Breaks


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Line, Chemicals, and Reagents
2.2. Cell Culture
2.3. Cell Treatments
2.4. Cell Harvesting
2.5. MTS Assay for Determination of Cell Viability
2.6. Bromodeoxyuridine (BrdU) Cell Proliferation (Chemiluminescent) Assay
2.7. Flow Cytometry for Analysis of Cell Cycle
2.8. Aqueous Peroxide Detection
2.9. Single Cell Gel Electrophoresis (Comet Assay)
2.10. 8-Hydroxy-2-deoxyguanosine (8-OHdG) Detection
2.11. Capillary Electrophoresis Western Analysis
2.12. Statistical Analysis
3. Results
3.1. DAS Inhibits BaP-Induced Cell Proliferation Using MTS and BrdU Proliferation Assays
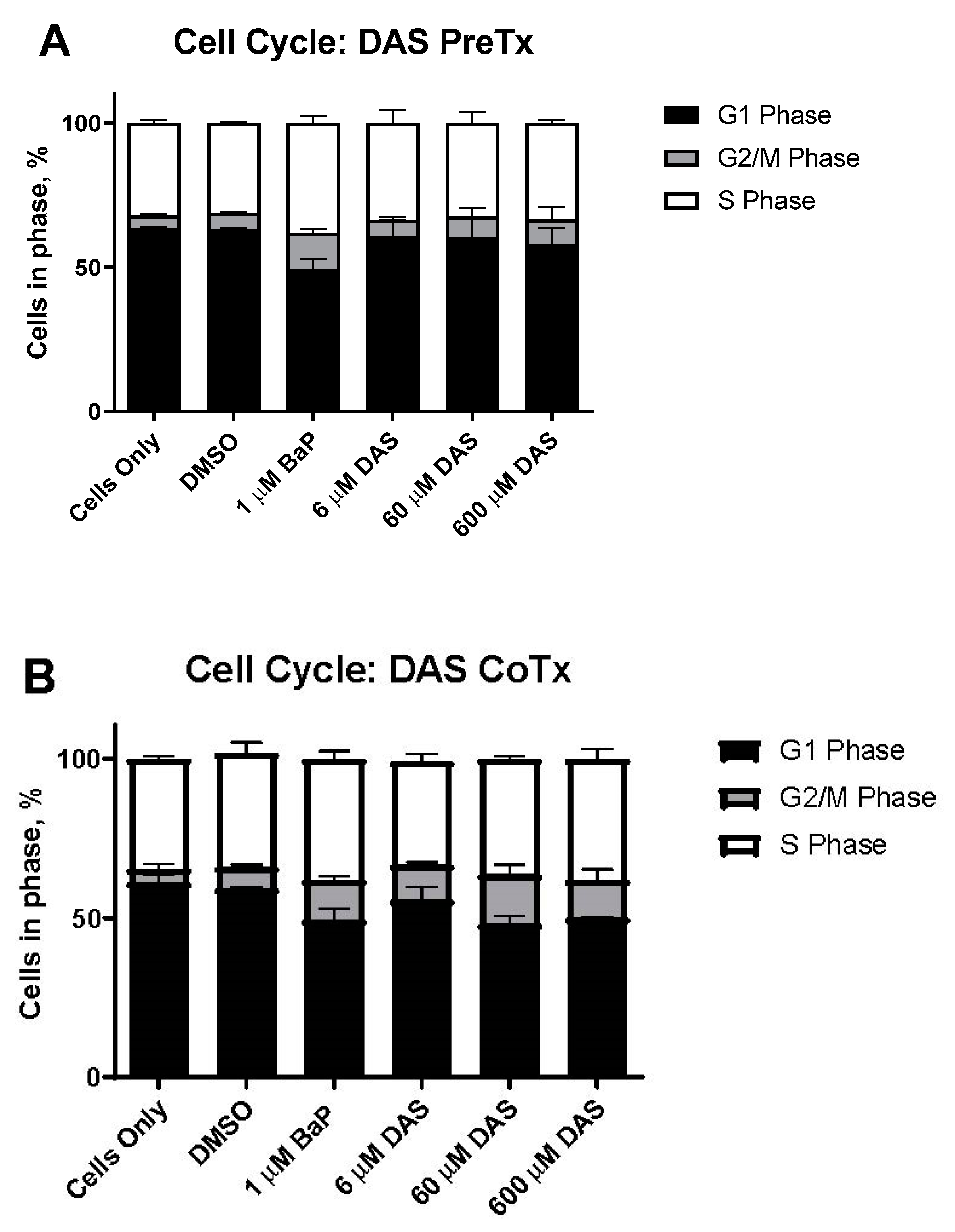
3.2. DAS Prevents BaP-Exposed MCF-10A Cells from Arresting at S and G2/M
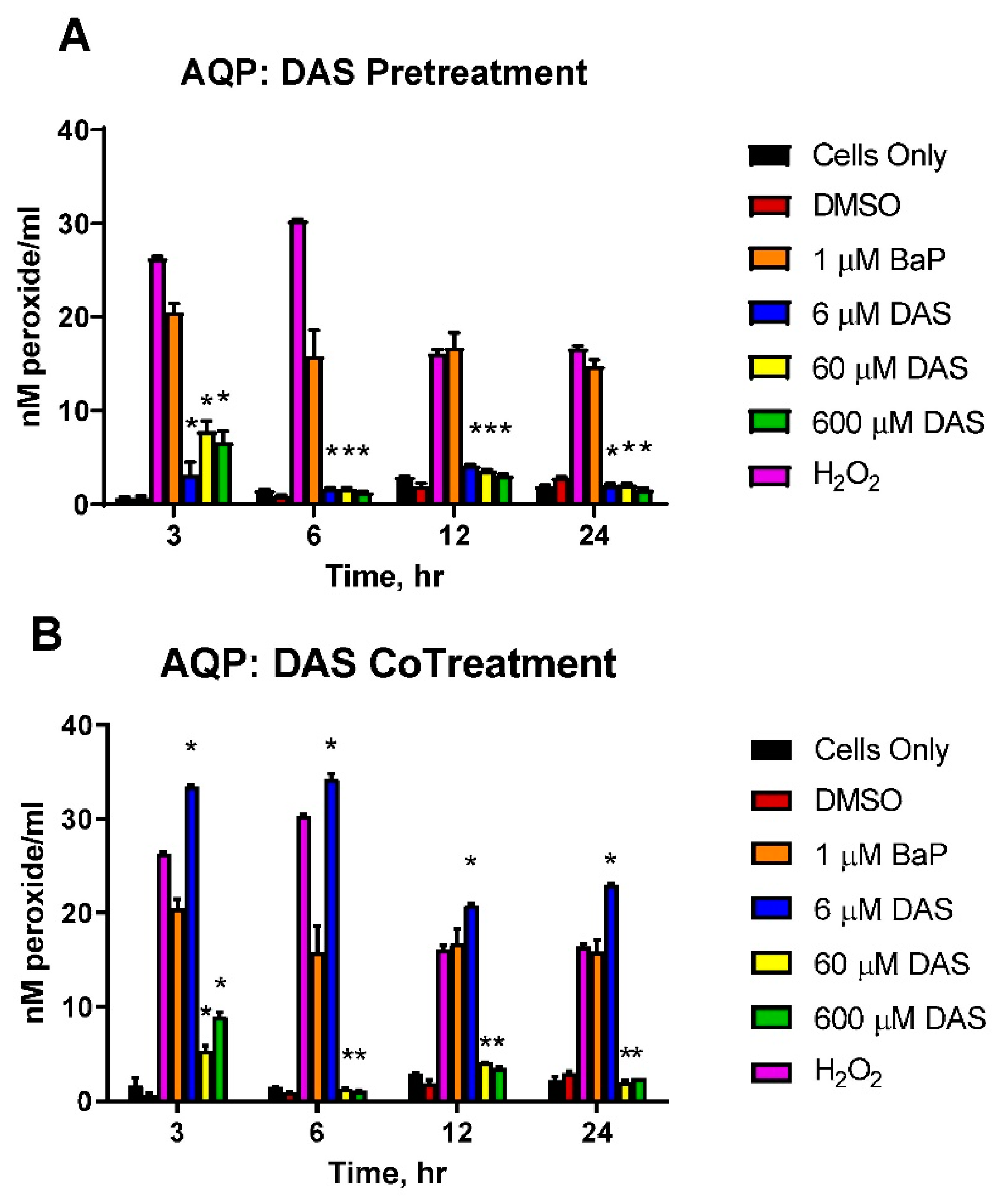
3.3. Reduction in the Accumulation of ROS in BaP-Treated Cells by DAS
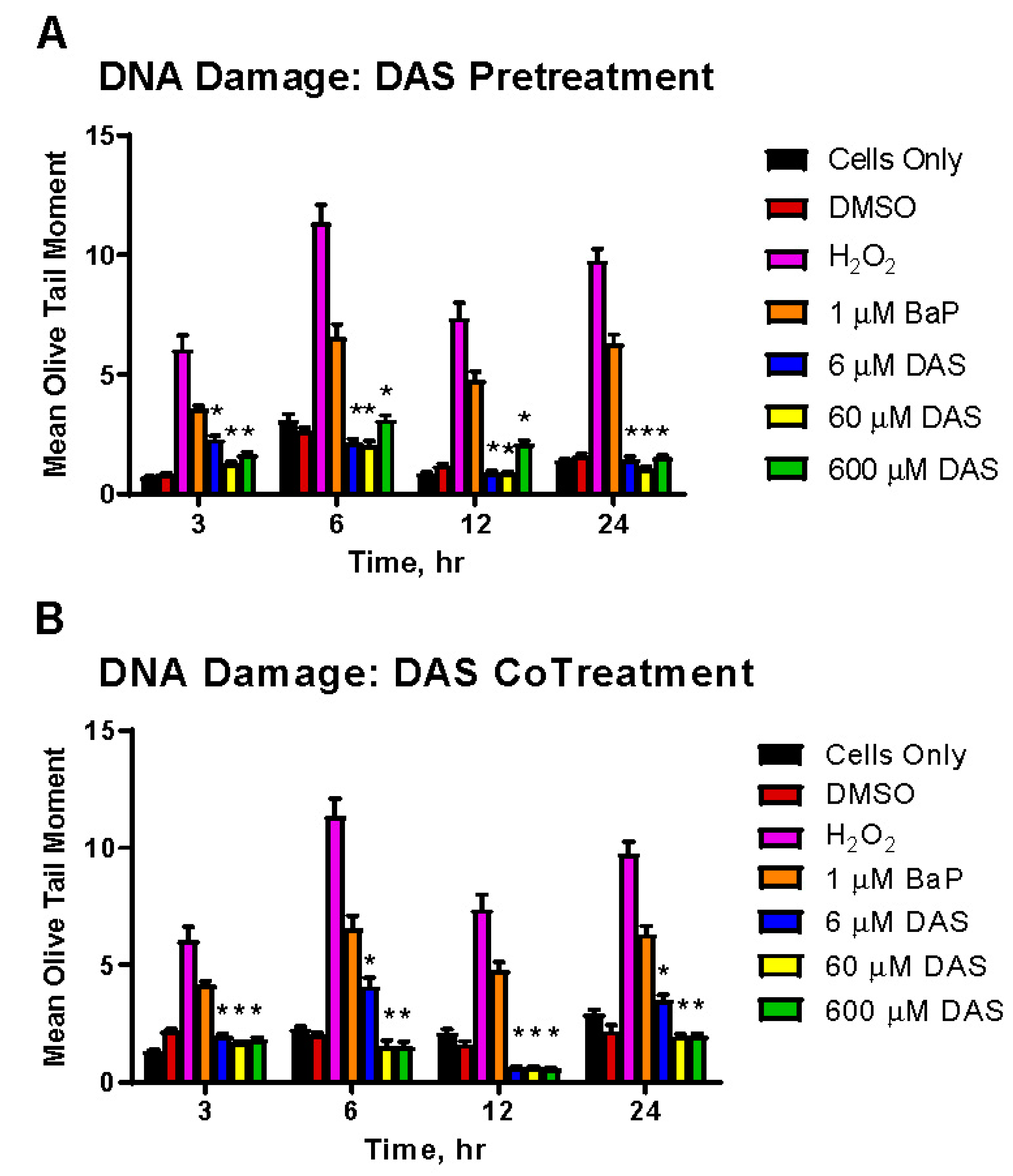
3.4. Inhibition of BaP-Induced DNA Damage by DAS in MCF-10A Cells
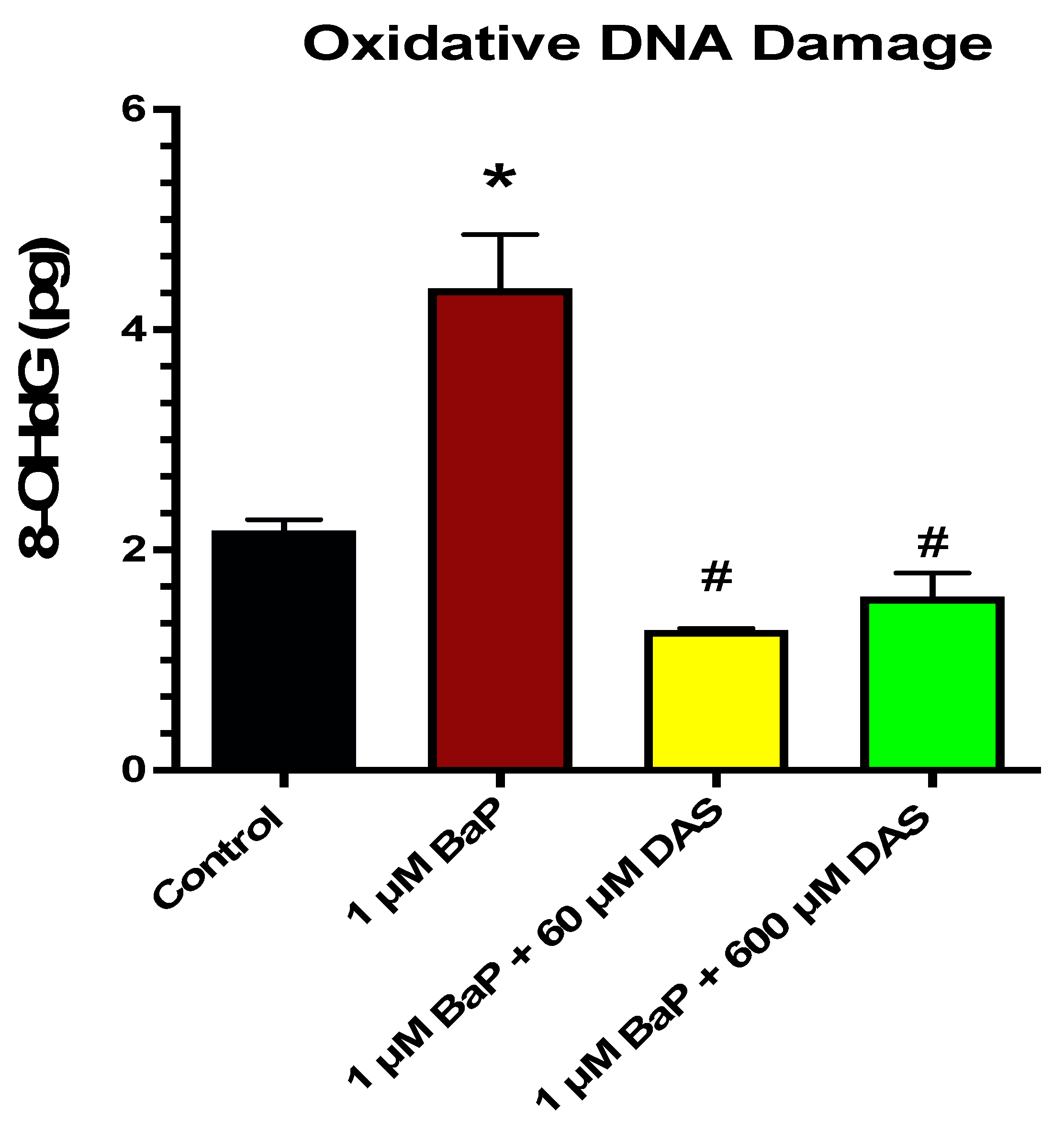
3.5. Inhibition of BaP-Induced Oxidative (8-OHdG) DNA Damage by DAS in MCF-10A Cells
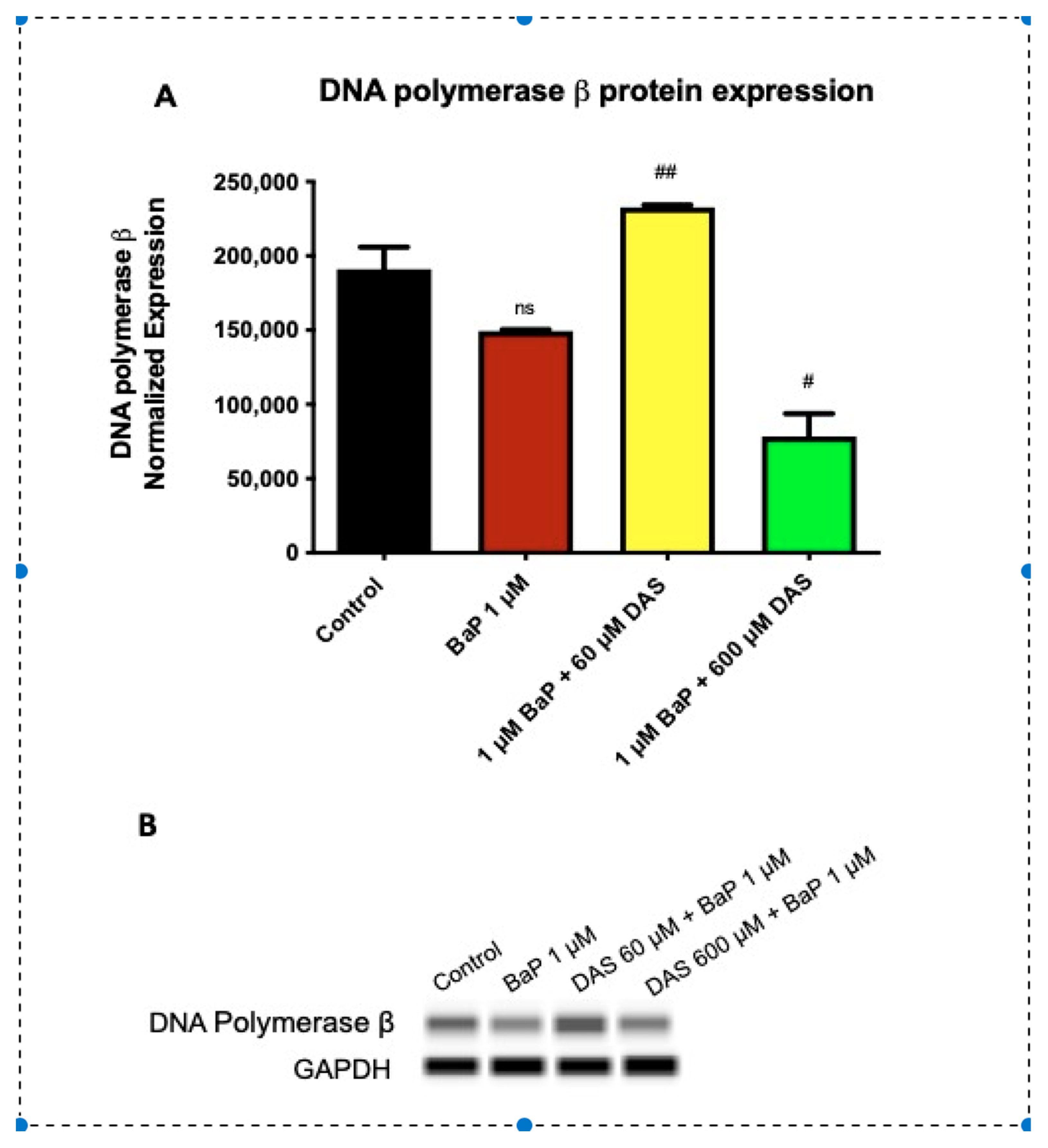
3.6. Increase in DNA Polymerase β (POLβ) Protein Expression by DAS in BaP Treated MCF-10A Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- ACS. Cancer Facts and Figures 2019–2020; American Cancer Society: Atlanta, GA, USA, 2021; pp. 1–44. [Google Scholar]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics 2020. CAN Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Anders, C.K.; Carey, L.A. Biology, Metastatic Patterns, and Treatment of Patients with Triple-Negative Breast Cancer. Clin. Breast Cancer 2009, 9 (Suppl. 2), S73–S81. [Google Scholar] [CrossRef]
- Mendonca, P.; Alghamdi, S.; Messeha, S.; Soliman, K.F.A. Pentagalloyl glucose inhibits TNF-α-activated CXCL1/GRO-α expression and induces apoptosis-related genes in triple-negative breast cancer cells. Sci. Rep. 2021, 11, 5649. [Google Scholar] [CrossRef]
- Cena, H.; Calder, P.C. Defining a Healthy Diet: Evidence for The Role of Contemporary Dietary Patterns in Health and Disease. Nutrients 2020, 12, 334. [Google Scholar] [CrossRef] [Green Version]
- Trio, P.Z.; You, S.; He, X.; He, J.; Sakao, K.; Hou, D.X. Chemopreventive functions and molecular mechanisms of garlic organosulfur compounds. Food Funct. 2014, 5, 833–844. [Google Scholar] [CrossRef]
- Omar, S.H.; Al-Wabel, N.A. Organosulfur compounds and possible mechanism of garlic in cancer. Saudi Pharm. J. 2010, 18, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Ip, C.; Lisk, D.J.; Stoewsand, G.S. Mammary cancer prevention by regular garlic and selenium-enriched garlic. Nutr. Cancer 1992, 17, 279–286. [Google Scholar] [CrossRef]
- Schaffer, E.M.; Liu, J.Z.; Green, J.; Dangler, C.A.; Milner, J.A. Garlic and associated allyl sulfur components inhibit N-methyl-N-nitrosourea induced rat mammary carcinogenesis. Cancer Lett. 1996, 102, 199–204. [Google Scholar] [CrossRef]
- Rao, P.S.; Midde, N.M.; Miller, D.D.; Chauhan, S.; Kumar, A.; Kumar, S. Diallyl Sulfide: Potential Use in Novel Therapeutic Interventions in Alcohol, Drugs, and Disease Mediated Cellular Toxicity by Targeting Cytochrome P450 2E1. Curr. Drug Metab. 2015, 16, 486–503. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.-H.; Wu, C.-M.; Liou, Y.-C. Volatile Compounds from Garlic. J. Agric. Food Chem. 1989, 37, 725–730. [Google Scholar] [CrossRef]
- Minami, T.; Boku, T.; Inada, K.; Morita, M.; Okazaki, Y. Odor components of human breath after ingestion of grated raw garlic. J. Food Chem. 1989, 54, 763–765. [Google Scholar] [CrossRef]
- Shukla, Y.; Arora, A.; Singh, A. Antitumorigenic potential of diallyl sulfide in Ehrlich ascites tumor bearing mice. Biomed. Environ. Sci. 2002, 15, 41–47. [Google Scholar]
- Surh, Y.J.; Lee, R.C.; Park, K.K.; Mayne, S.T.; Liem, A.; Miller, J.A. Chemoprotective effects of capsaicin and diallyl sulfide against mutagenesis or tumorigenesis by vinyl carbamate and N-nitrosodimethylamine. Carcinogenesis 1995, 16, 2467–2471. [Google Scholar] [CrossRef]
- Tadi, P.P.; Teel, R.W.; Lau, B.H. Organosulfur compounds of garlic modulate mutagenesis, metabolism, and DNA binding of aflatoxin B1. Nutr. Cancer 1991, 15, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Hageman, G.J.; van Herwijnen, M.H.; Schilderman, P.A.; Rhijnsburger, E.H.; Moonen, E.J.; Kleinjans, J.C. Reducing effects of garlic constituents on DNA adduct formation in human lymphocytes in vitro. Nutr. Cancer 1997, 27, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Green, M.; Wilson, C.; Newell, O.; Sadrud-Din, S.; Thomas, R. Diallyl sulfide inhibits diethylstilbesterol-induced DNA adducts in the breast of female ACI rats. Food Chem. Toxicol. 2005, 43, 1323–1331. [Google Scholar] [CrossRef]
- Wilson, C.; Aboyade-Cole, A.; Newell, O.; Darling-Reed, S.; Oriaku, E.; Thomas, R. Diallyl sulfide inhibits PhIP-induced DNA strand breaks in normal human breast epithelial cells. Oncol. Rep. 2007, 17, 807–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aboyade-Cole, A.; Darling-Reed, S.; Oriaku, E.; McCaskill, M.; Thomas, R. Diallyl sulfide inhibits PhIP-induced cell death via the inhibition of DNA strand breaks in normal human breast epithelial cells. Oncol. Rep. 2008, 20, 319–323. [Google Scholar]
- Nkrumah-Elie, Y.M.; Reuben, J.S.; Hudson, A.M.; Taka, E.; Badisa, R.; Ardley, T.; Israel, B.; Sadrud-Din, S.Y.; Oriaku, E.T.; Darling-Reed, S.F. The attenuation of early benzo(a)pyrene-induced carcinogenic insults by diallyl disulfide (DADS) in MCF-10A cells. Nutr. Cancer 2012, 64, 1112–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nkrumah-Elie, Y.M.; Reuben, J.S.; Hudson, A.; Taka, E.; Badisa, R.; Ardley, T.; Israel, B.; Sadrud-Din, S.Y.; Oriaku, E.; Darling-Reed, S.F. Diallyl trisulfide as an inhibitor of benzo(a)pyrene-induced precancerous carcinogenesis in MCF-10A cells. Food Chem. Toxicol. 2012, 50, 2524–2530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Block, E. The chemistry of garlic and onions. Sci. Am. 1985, 252, 114–119. [Google Scholar] [CrossRef]
- Gued, L.R.; Thomas, R.D.; Green, M. Diallyl sulfide inhibits diethylstilbestrol-induced lipid peroxidation in breast tissue of female ACI rats: Implications in breast cancer prevention. Oncol. Rep. 2003, 10, 739–743. [Google Scholar]
- Thomas, R.D.; Green, M.R.; Wilson, C.; Sadrud-Din, S. Diallyl sulfide inhibits the oxidation and reduction reactions of stilbene estrogens catalyzed by microsomes, mitochondria and nuclei isolated from breast tissue of female ACI rats. Carcinogenesis 2004, 25, 787–791. [Google Scholar] [CrossRef] [Green Version]
- Wargovich, M.J.; Woods, C.; Eng, V.W.; Stephens, L.C.; Gray, K. Chemoprevention of N-nitrosomethylbenzylamine-induced esophageal cancer in rats by the naturally occurring thioether, diallyl sulfide. Cancer Res. 1988, 48, 6872–6875. [Google Scholar]
- Siriwardhana, N.; Wang, H.C. Precancerous carcinogenesis of human breast epithelial cells by chronic exposure to benzo[a]pyrene. Mol. Carcinog. 2008, 47, 338–348. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Shukla, Y. Antitumour activity of diallyl sulfide on polycyclic aromatic hydrocarbon-induced mouse skin carcinogenesis. Cancer Lett. 1998, 131, 209–214. [Google Scholar] [CrossRef]
- Singh, A.; Shukla, Y. Antitumour activity of diallyl sulfide in two-stage mouse skin model of carcinogenesis. Biomed. Environ. Sci. 1998, 11, 258–263. [Google Scholar]
- Sheen, L.Y.; Wu, C.C.; Lii, C.K.; Tsai, S.J. Effect of diallyl sulfide and diallyl disulfide, the active principles of garlic, on the aflatoxin B(1)-induced DNA damage in primary rat hepatocytes. Toxicol. Lett. 2001, 122, 45–52. [Google Scholar] [CrossRef]
- Wu, P.P.; Chung, H.W.; Liu, K.C.; Wu, R.S.; Yang, J.S.; Tang, N.Y.; Lo, C.; Hsia, T.C.; Yu, C.C.; Chueh, F.S.; et al. Diallyl sulfide induces cell cycle arrest and apoptosis in HeLa human cervical cancer cells through the p53, caspase- and mitochondria-dependent pathways. Int. J. Oncol. 2011, 38, 1605–1613. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.A.; Cha, Y.Y.; Park, M.S.; Kim, J.M.; Lim, Y.C. Diallyl sulfide induces growth inhibition and apoptosis of anaplastic thyroid cancer cells by mitochondrial signaling pathway. Oral Oncol. 2010, 46, e15–e18. [Google Scholar] [CrossRef]
- Wang, H.C.; Yang, J.H.; Hsieh, S.C.; Sheen, L.Y. Allyl sulfides inhibit cell growth of skincancer cells through induction of DNA damage mediated G2/M arrest and apoptosis. J. Agric. Food Chem. 2010, 58, 7096–7103. [Google Scholar] [CrossRef]
- Truong, D.; Hindmarsh, W.; O’Brien, P.J. The molecular mechanisms of diallyl disulfide and diallyl sulfide induced hepatocyte cytotoxicity. Chem. Biol. Interact. 2009, 180, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Sriram, N.; Kalayarasan, S.; Ashokkumar, P.; Sureshkumar, A.; Sudhandiran, G. Diallyl sulfide induces apoptosis in Colo 320 DM human colon cancer cells: Involvement of caspase-3, NF-kappaB, and ERK-2. Mol. Cell Biochem. 2008, 311, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.S.; Ham, Y.A.; Choi, J.H.; Kim, J. Effects of allyl sulfur compounds and garlic extract on the expression of Bcl-2, Bax, and p53 in non small cell lung cancer cell lines. Exp. Mol. Med. 2000, 32, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, S.; Banik, N.L.; Patel, S.J.; Ray, S.K. Garlic compounds induced calpain and intrinsic caspase cascade for apoptosis in human malignant neuroblastoma SH-Sy5Y cells. Apoptosis 2007, 12, 671–684. [Google Scholar] [CrossRef]
- Kim, Y.A.; Xiao, D.; Xiao, H.; Powolny, A.A.; Lew, K.L.; Reilly, M.L.; Zeng, Y.; Wang, Z.; Singh, S.V. Mitochondria-mediated apoptosis by diallyl trisulfide in human prostate cancer cells is associated with generation of reactive oxygen species and regulated by Bax/Bak. Mol. Cancer Ther. 2007, 6, 1599–1609. [Google Scholar] [CrossRef] [Green Version]
- Powolny, A.A.; Singh, S.V. Multitargeted prevention and therapy of cancer by diallyl trisulfide and related Allium vegetable-derived organosulfur compounds. Cancer Lett. 2008, 269, 305–314. [Google Scholar] [CrossRef] [Green Version]
- Ansar, S.; Iqbal, M.; AlJameil, N. Diallyl sulphide, a component of garlic, abrogates ferric nitrilotriacetate-induced oxidative stress and renal damage in rats. Hum. Exp. Toxicol. 2014, 33, 1209–1216. [Google Scholar] [CrossRef]
- Xiao, D.; Srivastava, S.K.; Lew, K.L.; Zeng, Y.; Hershberger, P.; Johnson, C.S.; Trump, D.L.; Singh, S.V. Allyl isothiocyanate, a constituent of cruciferous vegetables, inhibits proliferation of human prostate cancer cells by causing G2/M arrest and inducing apoptosis. Carcinogenesis 2003, 24, 891–897. [Google Scholar] [CrossRef]
- Xiao, D.; Herman-Antosiewicz, A.; Antosiewicz, J.; Xiao, H.; Brisson, M.; Lazo, J.S.; Singh, S.V. Diallyl trisulfide-induced G(2)-M phase cell cycle arrest in human prostate cancer cells is caused by reactive oxygen species-dependent destruction and hyperphosphorylation of Cdc 25C. Oncogene 2005, 24, 6256–6268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, D.; Pinto, J.T.; Gundersen, G.G.; Weinstein, I.B. Effects of a series of organosulfur compounds on mitotic arrest and induction of apoptosis in colon cancer cells. Mol. Cancer Ther. 2005, 4, 1388–1398. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.C.; Chung, J.G.; Tsai, S.J.; Yang, J.H.; Sheen, L.Y. Differential effects of allyl sulfides from garlic essential oil on cell cycle regulation in human liver tumor cells. Food Chem. Toxicol. 2004, 42, 1937–1947. [Google Scholar] [CrossRef]
- Jeffy, B.D.; Chen, E.J.; Gudas, J.M.; Romagnolo, D.F. Disruption of cell cycle kinetics by benzo[a]pyrene: Inverse expression patterns of BRCA-1 and p53 in MCF-7 cells arrested in S and G2. Neoplasia 2000, 2, 460–470. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Qi, Y.; Chen, Q.; Yang, D.; Tang, S.; Jin, X.; Gao, J.; Fu, J.; Zhou, Z.; Wang, J.; et al. Cyclin A is essential for the p53-modulated inhibition from benzo(a)pyrene toxicity in A549 cells. Toxicology 2009, 256, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Leadon, S.A.; Stampfer, M.R.; Bartley, J. Production of oxidative DNA damage during the metabolic activation of Benzo [a]pyrene in human mammary epithelial cells correlates with cell killing. Proc. Natl. Acad. Sci. USA 1988, 85, 4365–4368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, D.; Cho, Y.; Collins, L.B.; Swenberg, J.A.; Di Giulio, R.T. Effects of benzo[a]pyrene on mitochondrial and nuclear DNA damage in Atlantic killifish (Fundulus heteroclitus) from a creosote-contaminated and reference site. Aquat. Toxicol. 2009, 95, 44–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rekhadevi, P.V.; Diggs, D.L.; Huderson, A.C.; Harris, K.L.; Archibong, A.E.; Ramesh, A. Metabolism of the environmental toxicant benzo(a)pyrene by subcellular fractions of human ovary. Hum. Exp. Toxicol. 2014, 33, 196–202. [Google Scholar] [CrossRef] [Green Version]
- Gollapalle, E.; Wang, R.; Adetolu, R.; Tsao, D.; Francisco, D.; Sigounas, G.; Georgakilas, A.G. Detection of oxidative clustered DNA lesions in X-irradiated mouse skin tissues and human MCF-7 breast cancer cells. Radiat. Res. 2007, 167, 207–216. [Google Scholar] [CrossRef]
- Curtis, C.D.; DThorngren, L.; Nardulli, A.M. Immunohistochemical analysis of oxidative stress and DNA repair proteins in normal mammary and breast cancer tissues. BMC Cancer 2010, 10, 9. [Google Scholar] [CrossRef] [Green Version]
- Matsui, A.; Ikeda, T.; Enomoto, K.; Hosoda, K.; Nakashima, H.; Omae, K.; Watanabe, M.; Hibi, T.; Kitajima, M. Increased formation of oxidative DNA damage, 8-hydroxy-2′-deoxyguanosine, in human breast cancer tissue and its relationship to GSTP1 and COMT genotypes. Cancer Lett. 2000, 151, 87–95. [Google Scholar] [CrossRef]
- Musarrat, J.; Arezina-Wilson, J.; Wani, A.A. Prognostic and aetiological relevance of 8-hydroxyguanosine in human breast carcinogenesis. Eur. J. Cancer 1996, 32, 1209–1214. [Google Scholar] [CrossRef]
- Nyaga, S.G.; Jaruga, P.; Lohani, A.; Dizdaroglu, M.; Evans, M.K. Accumulation of oxidatively induced DNA damage in human breast cancer cell lines following treatment with hydrogen peroxide. Cell Cycle 2007, 6, 1472–1478. [Google Scholar] [CrossRef] [PubMed]
- Atukeren, P.; Yavuz, B.; Soydinc, H.O.; Purisa, S.; Camlica, H.; Gumustas, M.K.; Balcioglu, I. Variations in systemic biomarkers of oxidative/nitrosative stress and DNA damage before and during the consequent two cycles of chemotherapy in breast cancer patients. Clin. Chem. Lab. Med. 2010, 48, 1487–1495. [Google Scholar] [CrossRef] [PubMed]
- Tapiero, H. Influence of alcohol consumption and smoking habits on human health. Biomed. Pharmacother. 2004, 58, 75–76. [Google Scholar] [CrossRef]
- Caruso, J.A.; Reiners, J.J., Jr.; Emond, J.; Shultz, T.; Tainsky, M.A.; Alaoui-Jamali, M.; Batist, G. Genetic alteration of chromosome 8 is a common feature of human mammary epithelial cell lines transformed in vitro with benzo[a]pyrene. Mutat. Res. 2001, 473, 85–99. [Google Scholar] [CrossRef]
- Sigounas, G.; Hairr, J.W.; Cooke, C.D.; Owen, J.R.; Asch, A.S.; Weidner, D.A.; Wiley, J.E. Role of benzo[alpha]pyrene in generation of clustered DNA damage in human breast tissue. Free Radic. Biol. Med. 2010, 49, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, M.V.; Dickey, J.S.; Bonner, W.M.; Sedelnikova, O.A. gamma-H2AX in bystander cells: Not just a radiation-triggered event, a cellular response to stress mediated by intercellular communication. Cell Cycle 2007, 6, 2210–2212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kryston, T.B.; Georgiev, A.B.; Pissis, P.; Georgakilas, A.G. Role of oxidative stress and DNA damage in human carcinogenesis. Mutat. Res. 2011, 711, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Kassie, F.; Mersch-Sundermann, V. Induction of apoptosis in tumor cells by naturally occurring sulfur-containing compounds. Mutat. Res. 2005, 589, 81–102. [Google Scholar] [CrossRef]
- Iciek, M.; Kwiecien, I.; Wlodek, L. Biological properties of garlic and garlic-derived organosulfur compounds. Environ. Mol. Mutagen. 2009, 50, 247–265. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.L.; Cole, K.J.; Williams, J.A.; Millar, B.C.; Harvey, D.; Weaver, G.; Grover, P.L.; Phillips, D.H. Activation of genotoxins to DNA-damaging species in exfoliated breast milk cells. Mutat. Res. 2000, 470, 115–124. [Google Scholar] [CrossRef]
- Yu, F.L.; Bender, W.; Fang, Q.; Ludeke, A.; Welch, B. Prevention of chemical carcinogen DNA binding and inhibition of nuclear RNA polymerase activity by organosulfur compounds as the possible mechanisms for their anticancer initiation and proliferation effects. Cancer Detect. Prev. 2003, 27, 370–379. [Google Scholar] [CrossRef]
- Chattergee, N.; Walker, G.C. Mechanisms of DNA Damage, Repair, and Mutagenesis. Environ. Mol. Mutagen. 2017, 58, 235–263. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Wenjing, H.; Pan, L.; Boldogh, I.; Ba, X. The role of base excision repair enzyme OGG1 in gene expression. Cell. Mol. Life Sci. 2018, 75, 3741–3750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mordukhovich, I.; Beyea, J.; Herring, A.H.; Hatch, M.; Stellman, S.D.; Teltelbaum, S.L.; Richardson, D.B.; Millikan, R.C.; Engel, L.S.; Shantakumar, S.; et al. Polymorphisms in DNA repair genes, traffic-related polycyclic aromatic hydrocarbon exposure and breast cancer incidence. Int. J. Cancer 2016, 139, 310–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Gil, N.; Vidal-Gil, L.; Flores-Bellvor, M.; Maista, R.; Sancho-Pelluz, J.; Diaz-Llopis, M.; Barcia, J.M.; Romero, F.J. Ethanol-Induced Oxidative Stress modifies Inflammation and Angiogenesis Biomarkers in Retinol Pigment Epithelial Cells (ARPE-19): Role of CYP2E1 and its inhibition by Antioxidants. Antioxidants 2020, 9, 776. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.-G.; Yan, H.; Yao, P.; Zheng, W.; Zou, L.-J.; Sang, F.-F.; Li, K.; Sin, X.-F. CYP2E1-dependent hepatotoxicity and oxidative damage after ethanol administration in human primary hepatocytes. World J. Gastroenterol. 2005, 11, 4530–4535. [Google Scholar] [CrossRef]
- Uchiyama, Y.; Takeuchi, P.; Kodera, H.; Sakaguchi, K. Distribution and roles of X-family DNA polymerases in eukaryotes. Biochimie 2009, 9, 165–170. [Google Scholar] [CrossRef]
- Beard, W.A.; Wilson, S.H. Structure and mechanism of DNA polymerase beta. Biochemistry 2014, 53, 2768–2780. [Google Scholar] [CrossRef]
- Souliotis, V.L.; Vlachogiannis, N.I.; Pappa, M.; Argyriou, A.; Ntouros, P.A.; Sfikakis, P.P. DNA damage response and oxidative stress in systemic autoimmunity. Int. J. Mol. Sci. 2020, 21, 55. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Wu, M.; Cui, J.; Chen, C.; Zhang, Z.-Z. Effects of DNA polymerase beta on the genotoxicity and genetic instability induced benzo(a)pyrene. Zhonghue Lao Dong Wei Sheng Zhi Ye Bing Za Zhi 2011, 29, 801–805. (In Chinese) [Google Scholar]
- Wu, M.; Lai, Y.; Zhang, Z. [Effect of DNA polymerase beta on repair of DNA damage induced by benzo(a)pyrene]. Wei Sheng Yan Jiu 2010, 39, 13–18. (In Chinese) [Google Scholar] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Darling-Reed, S.F.; Nkrumah-Elie, Y.; Ferguson, D.T.; Flores-Rozas, H.; Mendonca, P.; Messeha, S.; Hudson, A.; Badisa, R.B.; Tilghman, S.L.; Womble, T.; et al. Diallyl Sulfide Attenuation of Carcinogenesis in Mammary Epithelial Cells through the Inhibition of ROS Formation, and DNA Strand Breaks. Biomolecules 2021, 11, 1313. https://doi.org/10.3390/biom11091313
Darling-Reed SF, Nkrumah-Elie Y, Ferguson DT, Flores-Rozas H, Mendonca P, Messeha S, Hudson A, Badisa RB, Tilghman SL, Womble T, et al. Diallyl Sulfide Attenuation of Carcinogenesis in Mammary Epithelial Cells through the Inhibition of ROS Formation, and DNA Strand Breaks. Biomolecules. 2021; 11(9):1313. https://doi.org/10.3390/biom11091313
Chicago/Turabian StyleDarling-Reed, Selina F., Yasmeen Nkrumah-Elie, Dominique T. Ferguson, Hernan Flores-Rozas, Patricia Mendonca, Samia Messeha, Alicia Hudson, Ramesh B. Badisa, Syreeta L. Tilghman, Tracy Womble, and et al. 2021. "Diallyl Sulfide Attenuation of Carcinogenesis in Mammary Epithelial Cells through the Inhibition of ROS Formation, and DNA Strand Breaks" Biomolecules 11, no. 9: 1313. https://doi.org/10.3390/biom11091313