
Abstract
Several endemic corona viruses (eCoVs) have been reported to be the most common etiologic agents for the seasonal common cold and also cause pneumonia. These eCoVs share extensive sequence homology with SARS-CoV-2, and immune responses to eCoVs can cross-react with SARS-CoV-2 antigens. Based on such cross-reactivity of antigens among eCoVs, the IgG antibodies against the spike protein (SP) of severe acute respiratory syndrome coronavirus (SARS-CoV) were isolated from pig serum using magnetic beads immobilized with SARS-CoV SP and a protein-A column. The selectivity of the isolated antibodies was tested using different types of antigens, such as SARS-CoV-2 nucleoprotein (NP), influenza A virus (Beijing type), influenza B virus (Tokio and Florida types), human hepatitis B virus surface antigen (HBsAg), and bovine serum albumin (BSA). From the selectivity test, the anti-SP antibodies isolated from pig serum had sufficient selectivity to other kinds of viral antigens, and the apparent binding constant of the isolated antibodies was approximately 1.5 × 10–8 M from the surface plasmon resonance (SPR) measurements. Finally, the isolated anti-SP antibodies were applied to the immunoassay of SP using competitive immunoassay configuration. The feasibility of the detection as well as the quantitative analysis of the SARS-CoV viral culture fluid was determined using four viral culture samples, namely, SARS-CoV, SARS-CoV-2, MERS-CoV, and CoV-229E.
Similar content being viewed by others


1 Introduction
Humans are reported to be infected by alpha- and beta-coronaviruses (CoVs). Especially, beta-CoVs have caused acute respiratory diseases, such SARS-CoV-2, SARS-CoV, MERS-CoV [1,2,3]. Usually, the different human CoVs, such as CoV strains of OC43, HKU1, NL63, CoV-229E are known to be among the most common etiologic agents for the seasonal common cold and also cause pneumonia [4, 5]. Such endemic CoVs (eCoVs) have been reported to share extensive sequence homology with SARS-CoV-2, and immune responses to eCoVs can cross-react with SARS-CoV-2 antigens [6,7,8,9,10]. Recently, the anti-SARS-CoV-2 nucleoprotein (NP) was isolated from pig serum using human NP immobilized on microbeads [11,12,13,14]. Isolating the anti-NP antibodies from pig serum was considered feasible based on the similarity in amino acid sequences between human NP and the NPs of other CoVs. From such homology in amino acid sequences, the isolated anti-NP antibodies could specifically detect NPs, and the viral culture of SARS-CoV-2 could be distinguished from those of SARS-CoV, MERS-CoV, and CoV-229E. In this work, anti-SARS-CoV spike protein (SP) antibodies were isolated from pig serum based on such cross-reactivity of antigens among eCoVs using human SP.
This work aimed to isolate antibodies against SARS-CoV SP from pig serum. Among four major structural proteins from the CoV genome such as SP, envelope protein, membrane protein, and NP [15], the SARS-CoV SP is a large protein with a molecular weight of 139.1 kDa, which is composed of 1,255 amino acids. The SP is classified as a group of class I viral fusion glycoproteins, and most of the protein appears on the viral surface. The SARS-CoV SP is known to be composed of two subunits: the S1 subunit recognizes and binds to the angiotensin-converting enzyme 2 (ACE2) receptor and the S2 subunit mediates the fusion of host and viral membranes [16]. From such reasons, the SARS-CoV SP has been chosen as an important target for vaccines which induce the production of antibodies to block binding as well as fusion of CoVs into host cells. [17, 18].
The feasibility of isolating anti-SP antibodies was also considered on the basis of the homology of amino acid sequences between human SP and the SP of other CoVs [19]. For example, pigs have been reported to be infected by alpha- and beta-CoVs [20], such as the porcine epidemic diarrhea virus (PEDV; alpha-CoV), transmissible gastroenteritis virus (TGEV; alpha-CoV), and porcine hemagglutinating encephalomyelitis virus (PHEV; beta-CoV) [1, 21]. As the first step, the amino acid homology between SARS-CoV SP and several kinds of porcine CoVs SP that infected pigs was analyzed in comparison with PEDV (alpha-CoV), TGEV (alpha-CoV), and PHEV (Beta-CoV) using the Basic Local Alignment Search Tool (BLAST, https://blast.ncbi.nlm.nih.gov). As summarized in Table 1, the SP from these CoV strains had a high identity and similarity in amino acid sequences compared to SARS-CoV SP. The SP from PHEV (beta-CoV) with 1349 amino acid residues was found to have 37% identity and 54% similarity to SARS-CoV SP (Table 2). The other two pig-infecting alpha-CoVs were found to have SPs with an identity and similarity of 31% and 48% for TGEV (Table 3) and 35% and 52% for PEDV (Table 4), respectively. This sequence analysis demonstrates the highly homologous amino acid sequences of SPs between porcine CoVs and SARS-CoV [1, 22].
In this work, the isolation of anti-SP antibodies from pig serum was performed using magnetic beads immobilized with human SP. The selectivity to SP and specific binding properties of the isolated antibodies were then analyzed using immunoassays. Using the surface plasmon resonance (SPR) biosensor, the apparent binding constant (KD) of the isolated anti-SP antibodies was estimated. Finally, the isolated anti-SP antibodies were used to differentiate the viral culture of SARS-CoV from that of SARS-CoV-2, MERS-CoV, and CoV-229E using a competitive assay configuration.
2 Results and Discussion
2.1 Isolation of Anti-SP Antibodies from Pig Serum
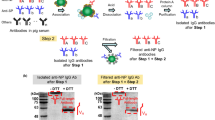
The anti-SP antibodies were isolated from pig serum using magnetic beads immobilized with SARS-CoV SP. As shown in Fig. 1a, the magnetic beads were mixed with pig serum, and the proteins bound to the magnetic beads were dissociated using treatment with acid. The dissociated proteins were considered to contain anti-SP antibodies. The antibody fraction was selectively isolated from the dissociated proteins using a protein-A column. The isolated antibody fraction showed an immunoglobulin protein band at the molecular weight of 150 kDa before reduction with dithiothreitol (DTT) (Fig. 1b). Fragments of the immunoglobulin were observed for heavy chains at a molecular weight of 50 kDa and for light chains at a molecular weight of 25 kDa after the disulfide bonds were reduced by treatment with DTT. These results showed that antibodies (IgGs) were successfully isolated from pig serum using magnetic beads immobilized with SARS-CoV SP. For the calculation of yield of antibodies from pig serum, the total protein in pig serum from Sigma-Aldrich Korea (Seoul, Korea) was estimated to be 87.2 ± 5.8 mg from pig serum (1 mL) (n = 5). And, the adsorbed protein to the magnetic beads with the immobilized SARS-CoV SP was estimated to be 48.4 ± 13.0 µg from pig serum (1 mL) (n = 5). Finally, the isolated anti-SP antibodies (IgGs) through protein-A column were estimated to be 13.8 ± 3.4 µg from pig serum (1 mL) (n = 5). Therefore, the yield of the isolated anti-SP antibodies was calculated to be 0.015 ± 0.004% (n = 5).

Isolation of anti-SP antibodies from pig-serum. a The anti-SP antibodies were isolated using magnetic beads with covalently immobilized SP. The isolation was carried out by (1) binding of proteins to the magnetic beads, (2) dissociation of proteins from magnetic bead using acid treatment, (3) isolation of antibody (IgG) fraction using protein-A column. b SDS-PAGE of the absorbed proteins (from acid treatment step) and isolated anti-SP antibodies under non-reducing condition without dithiotreitol (DTT) and reducing conditions with DTT
2.2 Properties of Anti-SP Antibodies from Pig Serum
The isolated anti-SP antibodies from pig serum were considered polyclonal antibodies composed of antibodies with different epitopes of SP. The binding properties of the isolated anti-SP antibodies were estimated using the immobilized SARS-CoV SP (Fig. 2a). The bound antibodies were quantified using secondary antibodies and the chromogenic reaction with 3,3’,5,5’-tetramethylbenzidine (TMB) [23, 24]. The isolated anti-SP antibodies in the concentration range of 6.8 ng/mL – 5.0 µg/mL were used to immobilize the SPs. The SARS-CoV-2 NP solution in the same concentration range was used as a negative control. The isolated anti-SP antibody showed quantitative binding to the immobilized SP, and the negative control showed nearly baseline level binding over the whole concentration range (Fig. 2b). These results showed that the isolated polyclonal antibodies had specific and quantitative binding properties to SARS-CoV SP. The selectivity of the isolated anti-SP antibodies was tested using different types of antigens, such as SARS-CoV SP, SARS-CoV-2 NP, influenza A virus (Beijing type), influenza B virus (Tokio and Florida types), human hepatitis B virus surface antigen (hHBsAg), and bovine serum albumin (BSA). As shown in Fig. 2c, the isolated anti-SP antibodies were observed to bind remarkably more to SARS-CoV SP in comparison to other proteins. These results showed that the isolated anti-SP antibodies had a significantly high selectivity and could be used for the specific detection of SARS-CoV SP. As previously reported [11,12,13], the anti-NP antibodies were also isolated from pig serum using magnetic beads immobilized with SARS-CoV-2 NP. When the same antigens in Fig. 2c were used for the selectivity test of the anti-NP antibodies, NP showed a remarkably higher response in comparison with other antigens, including SARS-CoV SP, as shown in Fig. 2d. These results showed that the anti-SP antibodies isolated from pig serum had high selectivity for other kinds of viral antigens.

Binding properties of the isolated anti-SP antibodies. a Configuration of the binding activity test using immobilized SP on microplate. b Comparison of binding activity of the isolated anti-SP antibodies to SARS-CoV SP and SARS-CoV-2 NP. The selectivity tests of the isolated c anti-SP antibodies and d anti-NP antibodies against different types of antigens, such as SARS-CoV-2 NP, influenza A virus (Beijing type), influenza B virus (Tokio and Florida types), human hepatitis B virus surface antigen (HBsAg), and bovine serum albumin (BSA)
The binding constants of the isolated anti-SP antibodies were estimated using an SPR biosensor. SP was immobilized on the Au chip of the SPR biosensor (Fig. 3a). To estimate the KD of the isolated anti-SP antibodies, the SPR response was monitored during the continuous flow of the isolated anti-SP antibody solution at a flow rate of 15 μL/min. The isolated anti-SP antibody solutions in the concentration range of 2.5 nM – 66.7 nM were used for the SPR measurements (Fig. 3b). As the isolated anti-SP antibodies were a mixture of antibodies (polyclonal antibodies), the KD could not be estimated for each antibody. However, the KD of the isolated anti-SP antibodies was approximately 1.5 × 10–8 M from the SPR measurements, which could be used to compare binding properties with other antibodies. The anti-NP antibody isolated from pig serum was carried out as shown in Fig. 3c. When anti-NP antibody solutions at the same concentration range as that in Fig. 3b were used for the binding assay to the SARS-CoV SP, anti-NP antibody showed an approximately 5-fold lower SPR response in comparison to anti-SP antibodies. These results showed that the anti-SP antibodies isolated from pig serum had sufficiently high specificity to SP.

Estimation of binding constant (KD) of the isolated antibodies using SPR biosensor. a Configuration for the SPR measurement using immobilized isolated antibodies on Au-chip. The SPR measurement was carried out under the continuous flow condition for the monitoring of association and dissociation of the antibodies. The sensorgrams of SPR measurements according to the concentration of isolated b anti-SP antibodies and c anti-NP antibodies against immobilized SP
2.3 Immunoassay of CoV Viral Fluids with Anti-SP Antibodies from Pig Serum
The isolated anti-SP antibodies were used for the detection of SPs in the samples. In this immunoassay, the SPs were immobilized on magnetic beads and mixed with samples, as well as a known concentration of the isolated anti-SP antibodies (Fig. 4a). The number of antibodies bound to the magnetic beads was quantified using a secondary antibody and the chromogenic reaction of TMB. In the case of negative samples without SPs, the isolated anti-SP antibodies could only bind to the SPs on the magnetic beads. In the case of positive samples with SPs (target antigens), the mixed antibodies could bind to the SPs on the magnetic beads as well as the SPs in the sample. Therefore, the number of antibodies in the positive samples on the magnetic beads was always lesser than that of the negative samples. The difference between the number of antibodies on the magnetic beads could be correlated with the number of SPs (target antigens) in the samples. As the isolated anti-SP antibodies consisted of different kinds of antibodies (polyclonal antibodies), it was very difficult to select antibodies with different epitopes for the formation of sandwich complexes. Such a competitive test could be effectively used for the detection of SPs in a sample even without the formation of a sandwich complex that requires two different types of antibodies, which should have different epitopes for the binding of antibodies. A competitive assay configuration was applied for the detection of SPs in the standard samples. Standard samples were prepared in the SP concentration range of 0.02 μg/mL– 20 μg/mL in phosphate-buffered saline (PBS), and the samples of SARS-CoV-2 NP were also prepared as a negative control (Fig. 4b). For the standard samples, the competitive assay could be used for the quantitative detection of SPs, and the signal for the negative control was observed to be at the baseline level in comparison with a blank sample (only with PBS).

Competitive immunoassay of SP in viral culture fluids of SARS-CoV-2, SARS-CoV, MERS-CoV, CoV strain 229E using the isolated anti-SP antibodies. a Configuration of the competitive immunoassay using magnetic beads with immobilized SP and the isolated anti-SP antibody solution at a known concentration. b Assay results of SARS-CoV SP and SARS-CoV-2 NP in standard samples using the competitive immunoassay. c Assay results of four kinds of CoV viral culture fluids using the competitive immunoassay
The competitive assay was applied to four viral culture samples: SARS-CoV, SARS-CoV-2, MERS-CoV, and CoV-229E. The viral culture fluids of four CoVs were diluted from 10-fold (dilution factor of 10%) to 4 × 104-fold (dilution factor of 0.04%) (Fig. 4c). For example, the dilution factor of culture fluid at 1% corresponded to a 100-fold dilution. The viral culture fluids of SARS-CoV showed a much higher response than those of the other three types of viral culture fluid in the whole dilution range. In particular, the viral culture fluids of SARS-CoV-2 and CoV-229E showed a much lower response than that of SARS-CoV. Quantitative analysis was deemed possible for the viral culture fluids of SARS-CoV at a dilution range of 10-fold – 4 × 104-fold. These results showed that the detection and quantitative analysis of viral culture fluid of SARS-CoV were feasible using the anti-SP antibodies isolated from pig serum.
3 Conclusion
The anti-SP antibodies were isolated from pig serum using magnetic beads immobilized with SARS-CoV SP and a protein-A column. For the calculation of yield of antibodies from pig serum, the total protein in pig serum was estimated to be 87.2 ± 5.8 mg from pig serum (1 mL) (n = 5). And, the adsorbed protein to the magnetic beads with the immobilized SARS-CoV SP was estimated to be 48.4 ± 13.0 µg from pig serum (1 mL) (n = 5). Finally, the isolated anti-SP antibodies (IgGs) through protein-A column were estimated to be 13.8 ± 3.4 µg from pig serum (1 mL) (n = 5). Therefore, the yield of the isolated anti-SP antibodies was calculated to be 0.015 ± 0.004% (n = 5). The selectivity of the isolated anti-SP antibodies was tested using different types of antigens, such as SARS-CoV SP, SARS-CoV-2 NP, influenza A virus (Beijing type), influenza B virus (Tokio and Florida types), hHBsAg, and BSA. From the selectivity test, the anti-SP antibodies isolated from pig serum had a high enough selectivity to other kinds of viral antigens, and the KD of the isolated anti-SP antibodies was approximately 1.5 × 10–8 M from the SPR measurements. The isolated anti-SP antibodies were applied to the immunoassay of SP using a competitive immunoassay configuration. In this immunoassay, the SPs were immobilized on magnetic beads and mixed with samples, as well as a known concentration of the isolated anti-SP antibodies. The competitive assay was applied to four kinds of viral culture samples, namely, SARS-CoV, SARS-CoV-2, MERS-CoV, and CoV-229E. The detection and quantitative analysis of the viral culture fluid of SARS-CoV were deemed to be feasible using the anti-SP antibodies isolated from pig serum.
4 Materials and Methods
4.1 Materials
The anti-pig IgG antibody labeled with horseradish peroxidase (ab102135) was purchased from Abcam (Cambridge, UK). M-280 tosyl-activated dynabeads (diameter of 2.8 µm, 14203) were purchased from Invitrogen Co. (Carlsbad, CA, USA). Pig serum (porcine serum, P9783-500 mL), BSA (A2153), and other chemicals were purchased from Sigma-Aldrich Korea (Seoul, Korea). The bicinchoninic acid (BCA) assay kit (23227) and TMB reagent (34021) was purchased from Thermo Fisher Scientific (Waltham, MA, USA). The SARS-CoV SP and SARS-CoV-2 NP were supplied by Optolane Inc (Seongnam, Korea). Influenza virus antigens (R86280 [A/Beijing/262/95], R86250 [B/Tokio/53/99]), R01247 [B/Florida/07/04], were purchased from Meridian Life Science Inc (Memphis, TN, USA). Culture fluids of SARS-CoV-2, SARS-CoV, MERS-CoV, and CoV-229E were obtained from Zeptometrix (Buffalo, NY, USA).
4.2 Purification of Anti-SP Antibodies
Isolated anti-SP (or NP) antibodies against SARS-CoV SP (or SARS-CoV-2 NP) were isolated from pig serum as follows. First, the pig serum (1 mL) was added to magnetic beads (5 mg) coated with SARS-CoV SP (20 µg). The reaction tube was agitated by mild shaking using a programmable digital rotator for 1 h (RT-10, Daehan Scientific Co, Wonju, Korea). Next, the absorbed proteins bound to the magnetic beads were dissociated by treatment with 1 mL of 0.1 M glycine–HCl buffer (pH 2.7) for 30 s, and then, 30 μL of 1 M Tris–HCl buffer (pH 11) was added for neutralization. The anti-SP antibodies were purified using a protein-A column without other proteins. Finally, the product was concentrated using an Amicon filter with a cutoff pore size of 10 kDa. The yield of anti-SP antibodies from 1 mL of pig serum was determined using a BCA kit according to the manufacturer’s instructions.
4.3 The Binding Affinity Analysis of the Anti-SP Antibodies
The binding properties of the isolated anti-SP antibodies were estimated using a Maxisorp microplate, as shown in Fig. 2a. For the microplate binding assay, the SARS-CoV SP (20 μg/mL, 100 μL) was incubated in a microplate at 4 °C for overnight. The SARS-CoV-2 NP solution in the same concentration range was used as a negative control (Fig. 2b). After washing with 0.1% PBST and PBS, each well was blocked with BSA (5 mg/mL, 150 μL) for 1 h. After washing, the isolated anti-SP antibodies (100 µL) in the concentration range of 6.8 ng/mL – 5.0 µg/mL were used to immobilize the SPs. Then, the HRP-labeled secondary antibody (10 ng/mL, 100 µL) was added and the solution was incubated for 1 h. After washing, the chromogenic reaction was carried out using the TMB reagent for 3 min. Then, each well was quenched with 2 M sulfuric acid (100 μL) and the optical density was measured at 450 nm using a Versamax Microplate Reader (Molecular Devices, Sunnyvale, CA, USA).
The selectivity of the isolated anti-SP antibodies was tested using different types of antigens, namely, SARS-CoV SP, SARS-CoV-2 NP, influenza A virus (Beijing type), influenza B virus (Tokio and Florida types), hHBsAg, and BSA. (Fig. 2c and 2d). Each antigen (20 μg/mL, 100 μL) was incubated in a microplate at 4 °C for overnight. After washing with 0.1% PBST and PBS, each well was blocked with BSA (5 mg/mL, 150 μL) for 1 h. After washing, the isolated anti-SP antibodies (5 µg/mL, 100 µL) were treated with the immobilized antigen. After washing, the HRP-labeled secondary antibody (10 ng/mL, 100 µL) was added and the solution was incubated for 1 h. The chromogenic reaction step was performed under the same conditions.
4.4 SPR Measurement
The KD of isolated anti-SP antibodies were estimated using an SPR biosensor from i-ClueBio (Seongnam, Korea) (Fig. 3b and 3c). Au chips for SPR measurements were prepared using the following procedure. First, BK-7 glass (10 × 10 mm2, thickness of 0.5 mm) was deposited with (1) a 2-nm adhesive layer of titanium and (2) 48-nm gold layer by sputtering. Next, the SPR chip was incubated with SARS-CoV SP solution (20 µg/mL, 100 µL) for overnight. After washing with 0.1% PBST and PBS, as an association step, anti-SP antibody solution at a concentration range of 2.5 nM– 66.7 nM was injected into the flow cell for 9 min, and as a dissociation step, PBS was injected into the flow cell for 8 min. The pumping rate was set as 15 µg/mL/min.
4.5 Standard Curve Fitting
SPR signals were fitted using the four-parameter logistic equation [25, 26]:
where a is the maximum and d is the minimum SPR signal (RU); c is the concentration of SARS-CoV SP; and b is Hill’s slope of plot.
4.6 Competitive Assay of Anti-SP Antibodies
A competitive assay configuration was applied for the detection of SPs in the standard samples. As shown in Fig. 4, magnetic beads (5 mg/mL, 10 μL) with immobilized SARS-CoV SP, isolated anti-SP antibodies (5 μg/mL, 10 μL) and SARS-CoV SP (range of 0.02 μg/mL – 20 μg/mL, 180 μL) were mixed at the same time. The SARS-CoV-2 NP solution in the same concentration range was used as a negative control. After a 1 h reaction with the washing step, HRP-labeled anti-pig IgG antibody solution (10 ng/mL, 100 μL) was incubated for 1 h. After washing, a chromogenic reaction was performed using the TMB reagent for 3 min. Then, each well was quenched with 2 M sulfuric acid (100 μL) and the optical density was measured using a microplate reader.
The competitive assay was applied to four viral culture samples: SARS-CoV, SARS-CoV-2, MERS-CoV, and CoV-229E. The viral culture fluids of four CoVs were diluted from 10-fold (dilution factor of 10%) to 4 × 104-fold (dilution factor of 0.04%).
References
Belouzard, S., Millet, J.K., Licitra, B.N., Whittaker, G.R.: Mechanisms of coronavirus cell entry mediated by the viral spike protein. Viruses 4, 1011–1033 (2012). https://doi.org/10.3390/v4061011
Chen, Y., Liu, Q., Guo, D.: Emerging coronaviruses: genome structure, replication, and pathogenesis. J. Med. Virol. 92, 418–423 (2020). https://doi.org/10.1002/jmv.25681
Elfiky, A.A.: Anti-HCV, nucleotide inhibitors, repurposing against COVID-19. Life Sci. (2020). https://doi.org/10.1016/j.lfs.2020.117477
Sariol, A., Perlman, S.: Lessons for COVID-19 immunity from other coronavirus infections. Immunity (2020). https://doi.org/10.1016/j.immuni.2020.07.005
Jain, S., et al.: Community-acquired pneumonia requiring hospitalization among US adults. New Engl. J. Med. 373, 415–427 (2015). https://doi.org/10.1056/NEJMoa1500245
Braun, J., et al.: SARS-CoV-2-reactive T cells in healthy donors and patients with COVID-19. Nature 587, 270–274 (2020). https://doi.org/10.1038/s41586-020-2598-9
Grifoni, A., et al.: Targets of T cell responses to SARS-CoV-2 coronavirus in humans with COVID-19 disease and unexposed individuals. Cell 181, 1489–1501 (2020). https://doi.org/10.1016/j.cell.2020.05.015
Le Bert, N., et al.: SARS-CoV-2-specific T cell immunity in cases of COVID-19 and SARS, and uninfected controls. Nature 584, 457–462 (2020). https://doi.org/10.1038/s41586-020-2550-z
Wec, A.Z., et al.: Broad neutralization of SARS-related viruses by human monoclonal antibodies. Science 369, 731–736 (2020). https://doi.org/10.1126/science.abc7424
Sagar, M., et al.: Recent endemic coronavirus infection is associated with less-severe COVID-19. J. Clin. Investig. (2021). https://doi.org/10.1172/JCI143380
Bong, J.-H., et al.: Pig Sera-derived Anti-SARS-CoV-2 Antibodies in Surface Plasmon Resonance Biosensors. BioChip. J. 14, 358–368 (2020). https://doi.org/10.1007/s13206-020-4404-z
Bong, J.-H., et al.: Competitive Immunoassay of SARS-CoV-2 Using Pig Sera-Derived Anti-SARS-CoV-2 Antibodies. BioChip. J. 15, 100–108 (2021). https://doi.org/10.1007/s13206-021-00011-6
Jung, J., et al.: Anti-SARS-CoV-2 Nucleoprotein Antibodies Derived from Pig Serum with a Controlled Specificity. BioChip. J. 15,195–203, (2021). https://doi.org/10.1007/s13206-021-00019-y
Song, S., et al.: Sensitivity Improvement in Electrochemical Immunoassays Using Antibody Immobilized Magnetic Nanoparticles with a Clean ITO Working Electrode. BioChip. J. 14, 308–316 (2020). https://doi.org/10.1007/s13206-020-4309-x
Ghodake, G.S., et al.: Biological characteristics and biomarkers of novel SARS-CoV-2 facilitated rapid development and implementation of diagnostic tools and surveillance measures. Biosens. Bioelectron. (2021). https://doi.org/10.1016/j.bios.2021.112969
Wong, S.K., Li, W., Moore, M.J., Choe, H., Farzan, M.: A 193-amino acid fragment of the SARS coronavirus S protein efficiently binds angiotensin-converting enzyme 2. J. Biol. Chem. 279, 3197–3201 (2004). https://doi.org/10.1074/jbc.C300520200
Krammer, F.: SARS-CoV-2 vaccines in development. Nature 586, 516–527 (2020). https://doi.org/10.1038/s41586-020-2798-3
Amanat, F., Krammer, F.: SARS-CoV-2 vaccines: status report. Immunity (2020). https://doi.org/10.1016/j.immuni.2020.03.007
Du, L., et al.: The spike protein of SARS-CoV—a target for vaccine and therapeutic development. Nat. Rev. Microbiol. 7, 226–236 (2009). https://doi.org/10.1038/nrmicro2090
Monchatre-Leroy, E., et al.: Identification of alpha and beta coronavirus in wildlife species in France: bats, rodents, rabbits, and hedgehogs. Viruses 9, 364 (2017). https://doi.org/10.3390/v9120364
Kim, J.-M., et al.: Identification of Coronavirus Isolated from a Patient in Korea with COVID-19. Osong Public Health Res. Perspect. 11, 3 (2020). https://doi.org/10.24171/j.phrp.2020.11.1.02
Sun, C., et al.: SARS-CoV-2 and SARS-CoV spike-RBD structure and receptor binding comparison and potential implications on neutralizing antibody and vaccine development. Biorxiv. (2020). https://doi.org/10.1101/2020.02.16.951723
Kim, H.-R., et al.: An On-chip Chemiluminescent Immunoassay for Bacterial Detection using in Situ-synthesized Cadmium Sulfide Nanowires with Passivation Layers. BioChip. J. 14, 268–278 (2020). https://doi.org/10.1007/s13206-020-4305-1
Jung, J., et al.: Screening of Fv Antibodies with Specific Binding Activities to Monosodium Urate and Calcium Pyrophosphate Dihydrate Crystals for the Diagnosis of Gout and Pseudogout. ACS Appl. Bio mater. 4, 3388–3397 (2021). https://doi.org/10.1021/acsabm.0c01680
Hasani, A., et al.: Recent progress of two-dimensional materials and metal–organic framework-based taste sensors. J. Korean Ceram. Soc. 57, 353–367 (2020). https://doi.org/10.1007/s43207-020-00047-8
Kim, J.H., et al.: No Tilt Angle Dependence of Grain Boundary on Mechanical Strength of Chemically Deposited Graphene Film. J. Korean Ceram. Soc. 56, 506–512 (2019). https://doi.org/10.4191/kcers.2019.56.5.11
Acknowledgements
This work was supported by the National Research Foundation of Korea [grant number NRF-2020R1A2B5B01002187 and NRF-2020R1A5A101913111].
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Jung, J., Bong, JH., Kim, TH. et al. Isolation of Antibodies Against the Spike Protein of SARS-CoV from Pig Serum for Competitive Immunoassay. BioChip J 15, 396–405 (2021). https://doi.org/10.1007/s13206-021-00033-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13206-021-00033-0