
Abstract
Novel trajectories of food production, urbanism, and inter-regional trade fueled the emergence of numerous complex Iron Age polities in central and southern Africa. Renewed research and re-dating efforts in Botswana, Zimbabwe, and along the Swahili Coast are transforming models for how inter-regional interaction spheres contributed to these patterns. While societies in present-day Zambia played an important role in the trade of copper, ivory, gold, and other resources between central and southern Africa, little is known about lifeways during the rise of social complexity in this region. This paper reports the results of re-excavation at Kalundu Mound on the Batoka Plateau of southern Zambia, one of the iconic mound sites of the Iron Age “Kalomo Culture.” New radiocarbon dates were combined with the original dates in a series of Bayesian models, indicating that previous chronologies for the site are not reliable and that the mound site likely developed rapidly from AD 1190 to 1410. Archaeobotanical, zooarchaeological, and paleo-proteomic analyses of excavated materials suggests a broad subsistence base combining wild and domesticated species, including the first reported evidence for finger millet (Eleusine coracana) in the region. Considering these findings, it is necessary to re-evaluate the temporal context of the Kalomo site-group, and to also systematically reinvestigate the systems of exchange and subsistence that supported Later Iron Age complexity.
Résumé
De nouvelles orientations de production alimentaire, d'urbanisme et de commerce interrégional ont contribué à l'émergence de nombreux régimes politiques complexes de l'âge de fer en Afrique centrale et australe. La reprise des recherches et un nouveau travail sur la datation au Botswana, au Zimbabwe et le long de la côte swahilie sont en train de transformer notre vision de la manière dont les sphères d'interaction interrégionale ont contribué à ces tendances. Alors que les sociétés actuelles en Zambie ont joué un rôle important dans le commerce du cuivre, de l'ivoire, de l'or et d'autres ressources entre l'Afrique centrale et australe, on en sait peu sur les modes de vie pendant l'essor de la complexité sociale dans cette région. Cet article rapporte les résultats de nouvelles fouilles à Kalundu Mound, sur le plateau de Batoka, au sud de la Zambie, l'un des sites de buttes emblématiques de la « culture de Kalomo » de l'âge de fer. De nouvelles datations au carbone ont été combinées avec les datations originales dans une série de modèles bayésiens, indiquant que les chronologies précédentes du site ne sont pas fiables et que ce site de butte s’est probablement développé rapidement de 1190 à 1410 après JC. Les analyses paléobotaniques, archéozoologiques et paléoprotéomiques des matériaux fouillés suggèrent une base de subsistance plus large combinant des espèces sauvages et domestiquées, y compris les premières traces signalées de mil éleusine (Eleusine coracana) dans la région. Compte tenu de ces résultats, il est nécessaire de réévaluer le cadre temporel du groupe de sites de Kalomo et de réexaminer méthodiquement les systèmes d'échange et de subsistance qui ont soutenu la complexité de l'âge du fer ultérieur.
Similar content being viewed by others


Introduction
The Later Iron Age (LIA) of southern Africa is characterized by the emergence of major polities and expansive trade networks connecting the interior to Indian Ocean markets. Major sites with significant architecture like Great Zimbabwe, Bosutswe, and Mapungubwe are among the most iconic monuments associated with the social complexity that manifested during this period (c. AD 900–1650) (Caton-Thompson, 1931; Chirikure et al., 2013a, b, Chirikure, 2014; Denbow et al., 2008; Huffman, 2009a; 2015; Pikirayi, 2002; Sinclair et al., 1993). Trade networks and processes of sociopolitical change were mediated by diverse forager, farmer, and pastoralist communities. The interactions between these communities in the LIA fundamentally shaped linguistic, cultural, and economic legacies of pre- and post-colonial southern Africa (de Luna, 2006; Fagan, 1969; Musonda, 1987, 2013a, 2013b; Ndlovu, 2011). It is increasingly clear that peoples in the “peripheries” played an important role in the broader heterarchical networks and were themselves affected by the trajectories of social complexity underway across the region (Chirikure, et al., 2018a; Klehm, 2017; Mukwende et al., 2018; Pikirayi & Pwiti, 1999).
The Lower Zambezi Valley, connecting Zambia, Zimbabwe, and Mozambique to the Indian Ocean, was a significant avenue for trade and population interactions during the Iron Age but has been less well studied than the major settlements further south. Increased interactions in trade networks across the Indian Ocean are evidenced in the appearance of glass beads, Asian food crops, and other trade goods in the coastal and southeastern African interior over the last one thousand years (Boivin et al., 2014; Chirikure, 2014; Denbow et al., 2015). Increasing Portuguese involvement in southern African trade in the sixteenth and seventeenth centuries made the immediate Lower Zambezi Valley particularly important, reflected in the increased evidence of commercial wealth during this period (de Luna, 2017, p. 1090). Even in earlier centuries, the communities of this region played a critical role in facilitating the movement of copper from northern Zambia into southern trade networks and were also involved in the trade of gold, ivory, and iron artifacts (de Luna, 2012, 2017; Kusimba et al., 2017; Nikis & Livingstone Smith, 2017; Swan, 2007).
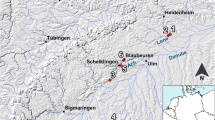
Within this increasingly globalized world of the second millennium AD, a phenomenon of large mound sites began to appear in the Lower Zambezi and Kafue River Valleys (Derricourt, 1976; Fagan, 1967; Vogel, 1970; Watt, 1980). The best known of these mound site entities is the “Kalomo Culture” marked by globular comb-stamped pottery, human and animal figurine traditions, and particular traditions of crouched burials. Sites exhibiting “Kalomo” features are primarily situated on the Batoka Plateau on the north side of the Zambezi River (all such sites extend across northern Zimbabwe) (Fagan, 1967, p. 35–37; Vogel, 1970; Watt, 1980) (Fig. 1). The Kalomo Culture is often associated with larger sites containing elaborate burials like Kalundu Mound and Isamu Pati. Smaller sites with Kalomo style ceramics have also been documented around the Batoka Plateau and Kafue Flats (Vogel, 1970). Based on current research, these sites seem to have been occupied by semi-sedentary agro-pastoralists of the LIA.

Location of Kalundu Mound relative to other Kalomo period sites on and around the Batoka Plateau, southern Zambia.
Kalundu Mound is one of the largest Kalomo Culture sites, covering an area of approximately 137 m by 106 m (or ~14,500 sq. m) that gradually slopes to a maximum height of almost 3 m above the modern ground surface (Inskeep, 1962, p. 138). Brian Fagan (1967, p. 58) estimated that around 130 people could have lived on the mound at a time. However, little is known about how the mound was constructed or how different parts of the mound were used through time. These sites and other LIA mounds in Zambia appear to be tells, formed by concentrated human occupation through the Iron Age as evidenced by the remains of houses, grain bins, storage pits, and dense archaeological debris (Fagan, 1967).
Fagan’s analysis of the stratigraphy at Kalundu Mound left open the possibility that Kalomo Culture populations rotated between mound sites on the Batoka Plateau as part of a fallow-cycle (Fagan, 1967, p. 42). Hence, he proposed that the “mounding” of the sites may have resulted from deliberate construction efforts to raise villages above potential flood levels (Fagan, 1967, p. 43). These hypotheses have implications for land use and the degree of labor organization and social complexity exhibited during the LIA of southern Africa. Existing radiometric dates for the Kalomo Culture were primarily generated in the 1960s, with additional dates from other sites like Matobo from the 1980s (e.g., AD 890±115, N-596 and AD 800±115, N-597; Watt, 1980, p. 84). These dates were obtained on unidentified specimens or derived from pooled samples of multiple charcoal specimens across a layer or context. The potential for old-wood or aquatic reservoir effects impedes the temporal resolution necessary to advance discussions on how the development of these mound sites relates to broader social, economic, and environmental changes.
Here, we report the initial results of renewed excavations at the Kalomo site of Kalundu Mound on the Batoka Plateau of southern Zambia, carried out in 2017 in collaboration with the Livingstone Museum and National Heritage Conservation Commission of Zambia. Kalundu is the first excavated Kalomo site (Inskeep, 1962) and appears to capture the longest known temporal sequence for the Kalomo Culture. The goal of the 2017 excavations was to generate a higher resolution 14C chronology for the site and, by extension, the Kalomo Culture. Collecting bone and archaeobotanical samples of short-lived plants from distinct stratigraphic contexts at Kalundu Mound for chronometric dating allowed us to build a more precise understanding of the occupation phases of the site by applying Bayesian chronological modeling to the extant and new AMS 14C assays. In the process, we also generated small zooarchaeological, proteomic zooarchaeology-by-mass-spectrometry (ZooMS), archaeobotanical, ceramic, and ornamental datasets that shed new light on the development of LIA societies in the region.
Situating Kalundu Mound Within the Regional Archaeology
Iron Age archaeology in southern Africa has long demonstrated diverse pathways to social complexity. Those studies challenge ideas derived from European and Near Eastern case studies about the role of “urbanism” (sensu stricto), institutionalized inequality, and land-use patterns — in generating and maintaining complexity. Despite its importance, research into the Iron Age archaeology of present-day Zambia progressed slowly in the last century. Excavations at Gokomere Mission (Schofield, 1948), Kalambo Falls (Clark et al., 1969), and Machili Forest Station (Clark & Fagan, 1965) provided the first evidence for the Early Iron Age in southern-central Africa. Many of these sites were ephemeral scatters and had limited preservation of bone or macrobotanicals, but with 14C dates (some of the first from Africa) of close to 50 cal BC, the evidence was strong enough to refute earlier ideas that the Iron Age in southern Africa began only half a century before European intrusion (Fagan, 1993).
Excavations by Summers (1958) and Robinson (1959) and investigations of the ceramic sequences by Schofield (1948) marked major advances in Iron Age archaeology south of the Zambezi. In contrast, the Iron Age of what was then Northern Rhodesia (Zambia) remained spotty and without real chronological anchors. However, the Iron Age research in present-day Zambia began to gain serious momentum when the road construction on the Batoka Plateau in 1956 revealed that some of the many hills dotting the region are archaeological mounds densely packed with pottery and animal bones.
Investigating what would become known as Kalundu Mound fell to then Keeper of Archaeology at the Livingstone Museum, Ray Inskeep. Recognizing the importance of this site, Inskeep excavated a 40-m-long trench oriented roughly north-south through the center of the mound. Pottery found in the stratified sequence reflected wholly new styles for Inskeep to develop a rough seriation. Inskeep, and later Brian Fagan (who expanded excavations in the 1960s), obtained 14C dates from the sequence and developed the first Iron Age sequence for the southern Central African highlands. With the Kalundu sequence as a foundation, Inskeep, Fagan, and others were able to identify and excavate several other stratified mound sites on the Batoka Plateau (Fagan, 1993). The stratigraphy at Kalundu became the basis for defining the Kalomo Culture and a reference point for its chronology. Not only was there a rich Iron Age in Zambia, but discoveries of copper ornaments, glass beads, and other exotic items proved that people living at these sites were participating in vast exchange networks that extended hundreds of kilometers to the north, south, and east of the Zambezi.
In subsequent years, Iron Age archaeology in Zambia enjoyed a brief boom, aided by the increasing pace of colonial damming projects necessitating rapid salvage. Massive survey and excavation projects in the Middle and Lower Zambezi Valley (Inskeep, 1962; Vogel, 1971; Watt, 1980), the Itezhi-Tezhi Basin (Derricourt, 1985), the Mulungushi Drainage (Robertson, 2000), and later the Upper Zambezi (Katanekwa, 1978, 1979) established key Iron Age sequences. These projects were critical in building regional chronologies and understanding the development of the cultural landscape from prehistoric times to the arrival of modern agro-pastoral communities (Phillipson, 1968, 1974).
Most of these early food-producing lifeways appear to have spread primarily through river valleys and only later permeated the hinterlands. A “western” tradition of channel-decorated pottery appears along the upper Zambezi only after ca. AD 1, along with evidence for domesticated livestock at the site of Salumano (and possibly Lusu Rapids) (Katanekwa, 1979). Another “central” tradition is known from the site of Muteteshi in the Mulungushi Drainage near Kabwe, where punctate motifs and horizons yielding two domesticated sorghum seeds were dated to ca. 50 BC–AD 150 (Robertson, 2000). Finally, a broad “southern” tradition of channeled and comb-stamp decorated ceramics is documented for several sites around Livingstone on the lower Zambezi River at a slightly later date (after ca. AD 350). Unlike the earlier sites, which appear largely ephemeral, sites dating from roughly AD 450 to 950 show a pronounced increase in daga (daub) from structures and large features indicative of longer-term villages. Smelting sites and kilns also increase dramatically by this time.
How these LIA traditions relate to each other and the Early Iron Age traditions of Central and eastern Africa remain unresolved, with multiple conflicting interpretations (Katanekwa, 2016, p. 323–326; also see Derricourt, 1976). There are also diverse proposals for the mechanism of spread and development of the Iron Age, involving varying degrees of population spread, local adoption, and farmer-forager interaction (Holden, 2006; Huffman, 1970, 1989; Musonda, 1987; Robertson, 2000; see also Crowther et al., 2018). Certainly by ca. AD 950, agro-pastoral, iron-producing economies were established across several environmental zones in Zambia, with increasing interconnectivity and participation in regional exchange. It is during this Later Iron Age phase that complex polities developed in southern Africa. Kalundu Mound became important during this period.
Kalundu Mound and the Kalomo Culture
Environmental Setting and Regional History
Kalundu is situated on the granitic Batoka Plateau, which historically was dominated by miombo woodlands characterized by Brachystegia and Julbernardia. Many of the Kalomo Culture sites are located near the interface between these woodlands and the surrounding low-elevation open grasslands where Hyparrhenia and Loudetia simplex thrive. During and after the rainy season (November to April), the average annual rainfall of 500–800 mm is enough to turn portions of the grasslands into waterlogged pans or dambos. Further afield, grasslands grade into open Acacia savannas interspersed with patches of bush. Tsetse flies, a vector for trypanosomiasis (African Sleeping Sickness), are absent from the highlands but were historically common at lower elevations in the region. While trypanosomiasis affects humans, it is the cause of a high mortality rate for cattle and can be devastating to herds. Despite this, the grasslands supported dense human settlements and large domesticated and wild herds thousands of years before European colonialism.
The use of this resource-rich landscape by ethnohistoric Tonga peoples has formed the basis for most speculation on prehistoric Iron Age subsistence economies. The Tonga arrived and settled in southern Zambia during the first half of the second millennium AD, about the time most other Bantu-speaking groups began arriving in the region as a part of large-scale migrations. Tonga-speaking peoples identify as either “Plains Tonga” of the Gwembe Valley or “Plateau Tonga” who occupy highlands along the Zambezi River, including the Batoka Plateau. Geographic ranges of the Tonga overlap with that of the Iron Age Kalomo, and many Kalomo sites have evidence of historic Tonga occupations in the uppermost horizons. While this has led to some speculation that the Tonga descended from the Iron Age Kalomo populations, this is not yet established from archaeological or archaeogenetic data.
Both the Plains and Plateau Tonga historically practiced mixed agro-pastoralism (Scudder, 1962). Most Tonga identify as cattle herders and place strong social and cultural value on keeping cattle herds. Management of cattle among the Tonga was similar to that of eastern African herders, involving minimal consumption of cattle outside of weddings, funerals, or other ritual events or major social gatherings. The Tonga kept some goat and sheep in addition to cattle, and the remainder of the diet consisted of milk products, sorghum (replaced by maize over the last century), finger millet, and wild foods, including fruits and root vegetables (Scudder, 1962, p. 80–88). Early European records discuss the Batoka plateau as having no trees, likely reflecting forest clearing practices for grazing lands and making charcoal for iron production. Historic Tonga also cleared the bush around shallow wetlands near streams and rivers (dambo ecologies) to reduce risks from trypanosomiasis, foot-rot, and other livestock diseases.
Recent decades have seen a shift toward intensive agriculture in much of the research area. Today, most Kalomo period sites on the Batoka Plateau are under cultivation, and non-African crops like maize, potato, and, less commonly, wheat are grown on them. Soils associated with Kalomo sites are known to be particularly rich. They were being mined for use in agriculture in nearby areas on the Batoka Plateau, at least by the early twentieth century AD, and this continues into the present.
The 1957 Excavations
Excavations at Kalundu Mound were first carried out in 1957. Raymond Inskeep excavated four trenches, each about a meter wide, south of the new road that exposed archaeological occurrences (Inskeep, 1962). Three of the trenches (I, III, and IV) were eventually joined together to form a total length of 42 m (see Fig. 2). Trench II was in line with the others but placed at the very edge of the mound and produced a thin (25.4 cm) horizon with little archaeological material overlying sterile subsoils. The main trench on top of the mound found a much thicker set of archaeological horizons, reaching approximately 1.7 m down to a sterile “old ground surface” (Inskeep, 1962). In a few locations, large pits with archaeological materials were found on this surface.

Location of 2017 excavations relative to 1957 excavations at Kalundu Mound.
Inskeep (1962) cautiously identified three main stratigraphic units (Zones I–III), although some of these divisions were not clearly identifiable in later expansions to the trenches by Fagan (1967; see Fig. 3). Inskeep’s Zone I was a “hard brownish earth” (Inskeep, 1960, p. 138) with small quantities of pottery and daga, and sporadic ephemeral features like post holes and shallow pits. Being near the surface, there was also evidence of animal disturbance and more recent cultivation into this horizon. Zone II, when visible, was a layer of light-gray soft ashy sediment with abundant potsherds, bones, and charcoal. There was little evidence of features at this level. Below Zone II, Inskeep encountered a compacted brown horizon that he called Zone III. Arbitrary spits within Zone III had markedly different densities of artifacts, suggesting discontinuous archaeological deposition. Despite high quantities of daga relative to Zone II, the only features detected were a single post hole and several pits dug into the sterile sub-soils overlying granitic bedrock. Pits varied in shape and size. Most are 5–45 cm deep, but one extended 1.8 m and was filled with archaeological materials.

Comparison of recorded stratigraphic profiles and radiocarbon date locations from the 2017 excavations reported here (left) and Inskeep’s (1957) excavations (right) at Kalundu Mound (see Fig. 2 for trench locations). Sample locations for 1960s radiocarbon dates approximated based on reported depths (Fagan, 1967, p. 41).
Ceramics are the most abundant materials from the 1950s excavations and the later work at Gundu, Kalomo, and Isamu Pati. These helped establish a rough seriation whereby the earlier ladder-and-groove and punctate motifs gave way to bordered roulette and triangular motifs (Katanekwa, 2005, 2016). Iron artifacts, tuyere fragments, and slag were present in low proportions throughout the sequence. Several ostrich eggshells and aquatic shell beads were found, but only a single glass bead was recovered from Zone I. Presumed evidence for agriculture was restricted to a single grindstone.
The 1960–1962 Excavations
Expanded excavations and subsequent analyses at several Kalomo sites by Brian Fagan (1967) added new stratigraphic information, 14C dates, and vital subsistence data to the emerging picture of the Kalomo Culture. The recovery of small quantities of sorghum (Sorghum bicolor) at Kalundu and Isamu Pati confirmed the agricultural components of Kalomo economies. Fagan (1967, p. 63) also reports macrobotanical evidence for cultivation or gathering of fruits, including figs (identified by leaf imprints in clay); and mushuk/loquot (Uapaca kirkiana), mabula (Parinari curatellifolia/cork tree), and Duiker tree (Pseudolachnostylis maprouneifolia), all of which were represented by charred plant remains. All of these fruits are still eaten by people in the region today.
Faunal analyses confirmed the presence of domesticated livestock, along with a diverse array of wild fauna. Most faunal identifications were based on complete or nearly complete mandibles and teeth. However, sample sizes are small, with significant potential for behavioral or taphonomic biases in the representation of specific species and age cohorts. This scenario also limits potential interpretations of butchery practices.
Cattle appeared to be the most important domesticated animal at Kalomo Culture sites. Fagan and colleagues identified what appear to be caprines but in small quantities, and they were unable to definitively determine if these were goats or sheep (Fagan, 1967, p. 197). Small-stock were less common than cattle on the Batoka plateau through pre- and post-colonial periods, possibly due to ecological and zoonotic restrictions (Scudder, 1962, p. 164). Based on tooth eruption patterns from the larger sample of mandibles at Isamu Pati, cattle were killed at various ages, but more commonly as mature adults (Fagan, 1967, p. 66). This may reflect forms of herd management that were oriented around milk production instead of culling younger individuals for meat. Identifiable elements for cattle are predominantly lower-limb bones, especially phalanges (see Fagan, 1967, p. 196). In addition, the remains of seven domesticated dogs (including one nearly complete skeleton, two almost complete skulls, and several mandible fragments) were found at Isamu Pati, but none were identified from Kalundu (see Degerbol, 1967). Finally, Fagan reports chickens (Gallus gallus) as identified by Mr. A. Wetmore, from nearly all archaeological horizons at both Kalundu and Isamu Pati. However, these may represent wild guinea fowl given recent zooarchaeological analyses that emphasize the difficulty and complexity of differentiating wild and domesticated fowl (MacDonald, 1992; Prendergast et al., 2017; Woldekiros & D’Andrea, 2017).
Both Kalundu and Isamu Pati yielded evidence that hunting remained an important dimension to Kalomo subsistence. Duiker appears to be the most commonly hunted animal at Kalundu (Fagan, 1967, p. 71–72). More detailed faunal data is available for Isamu Pati, where duiker constitute 34% of identified mandibles, followed by grysbuck (17%), waterbuck (13%), oribi (12%), buffalo (8%), and reedbuck (4%). Only one or two mandible fragments represented each of the other bovids like impala and kudu, and species such as zebra, warthog, bush pig, hartebeest, elephant, and rhinoceros. Of rodents, several species of Muridae and Viverridae are present only as post-crania, and Fagan (1967, p. 71) reports black rat (Rattus rattus) from Isamu Pati. Some carnivore species like leopard, lion, and hyena were identified in small quantities at Isamu Pati but not in Kalundu. Conversely, Kalundu produced a small proportion of fish remains, whereas none was recovered from Isamu Pati. Based on patterns at Isamu Pati, Gundu, and Matobo, there is evidence that wild fauna was less common relative to cattle in the uppermost strata (Fagan, 1963, p. 176, 1978; Vogel, 1970, p. 84; Watt, 1980).
Available age estimates for wild fauna from Kalundu and Isamu Pati show a bias toward hunting adult or older individuals, except for grysbok, where there is a slight bias toward immature and adolescent individuals (Fagan, 1967, p. 76). Many of the mandibles selected for these analyses were excavated from pit features possibly associated with house structures. Multiple complete mandibles in pits are conspicuous and might signal ritual, rather than subsistence, behaviors. Only a more comprehensive analysis of the fauna from the site can help determine the accurate representation of wild animal species and, by extension, the intensity of LIA hunting in southern Zambia.
The 1962 excavations at Kalundu did not produce any additional trade beads ( n=1 from the 1959 cuttings). At Isamu Pati, 29 glass beads and a small collection of shell beads were recovered from the upper horizons, possibly indicating different levels of engagement in trade among Kalomo sites, but only in the later phases.
The 2017 Excavations
In August–September of 2017, we carried out small test excavations at several sites in southern Zambia to refine the chronology for the development of food-producing economies during the Iron Age and collect new subsistence data, particularly archaeobotanical and faunal remains. In collaboration with the Livingstone Museum, the National Heritage Conservation Commission, and the current landowners, we opened a small 1-×-3-m trench at the National Monument site of Kalundu Mound (see Fig. 2). Original excavations at Kalundu Mound were placed entirely to the south of the road under construction in 1959. So we placed this trench, oriented E-W, slightly to the north of the road in an area that was used for cassava cultivation during Inskeep's excavations, but is now used principally for growing maize.
Excavations followed a single-context recording method wherein each distinct stratum or depositional episode receives a unique context number. Contexts were excavated in 5–10-cm spits until a new context was encountered to more accurately record spatial relationships for artifacts recovered from the sieves. Arbitrary subdivisions of contexts are identified with decimal values (e.g., context 3 may contain three 10-cm spits labeled “3,” “3.1,” “3.2”). Otherwise, the goal was to record the precise location of artifacts and faunal specimens over 2 cm in size, as well as features and sample locations with a total station. Two extensions (0.5 × 1 m and 0.5 × 0.5 m) were added to reveal a large stone feature in the upper horizons. Only the westernmost 1 sq. m. reached the basal interface with sterile eroding regolith labeled as the “old ground surface” by Inskeep (1962).
Flotation was carried out on bulk sediment samples from three contexts (3, 5, and 8; 10 liters each) at Kalundu, and a single hearth (context 6; 3 liters), producing 38.8 g of flot in total. Several larger seeds and endocarp (nutshell) fragments were also hand collected during excavation (8.0 g in total). The flots were sieved into size fractions (>2mm, 1–2mm, 0.5–1mm, and <0.5mm) and sorted with the aid of a low-power Olympus SZ61 stereozoom microscope (6–45×). Where necessary, higher power images were obtained using a Hitachi TM3030 desktop scanning electron microscope (SEM).
Due to the highly fragmentary nature of the assemblage, 102 specimens of animal bone taken from all contexts were sampled for identification of collagen proteins, using zooarchaeology by mass spectrometry (ZooMS) (Bradfield et al., 2019; Brown et al., 2020; Buckley et al., 2009; Welker et al., 2015, 2016). These included a random selection of bone fragments that were otherwise only identifiable to broad taxa size classes from all horizons (see expanded methods in ESM 1). Identifications were made using published spectra and the African Wild Bovid ZooMS Database (Janzen et al., 2021).
Results
Stratigraphy
The stratigraphic sequence in the 2017 test trench largely matched the archetypal contexts/layers described by Inskeep (1962, p. 188–189) in both the physical qualities of sediment and relative thickness of horizons (Fig. 3). Each horizon appears to be thinner in our excavations than those reported by Inskeep, likely because our test trench was placed at a slightly lower northern part of the mound.
Context 0 represents the upper 20 cm of compacted modern surface that had been heavily reworked through hand-plowed maize cultivation. The plow zone was heavily disturbed with substantial rooting, and it yielded fauna and Iron Age potsherds but no historic or recent artifacts. Context 1 was a firm brown sandy silt containing occasional sub-angular quartzite and granite granules and rare pebble-to-cobble-sized sub-angular granites. The layer corresponds to Inskeep’s “hard brownish earth” of Zone I. It had a low density of Kalomo Culture artifacts and faunal remains, as well as small fragments of clay daga.
Context 2 is a grayish brown sandy silt with moderate sub-rounded and sub-angular granules of quartzite and granite. There was a marked increase in the density of artifacts, fauna (including several large-sized bovid teeth), and daga in this context. After 20 cm, the context terminated at a stratigraphic interface with a loose gray horizon.
At a depth of 40 cm below the surface, we encountered a large stone-and-daga feature on the eastern side of the trench (Fig. 4). The feature emerged as having a linear form running roughly northwest-to-southeast and was 50–60 cm wide. The feature’s appearance coincided with a diffuse change in sediment, so the trench was divided into context 2 (to the east and outside of the feature) and context 3 (inside and to the west of the feature). The entirety of the context 3 feature rests on the same horizontal plane as the start of context 2. It comprises 2–3 courses of densely packed stones and large daga fragments mixed with potsherds. It extends from the north wall of the trench, takes a slight turn to the west, and ends abruptly after about 1 m. The “corner” of the feature is the highest and most heavily built-up. This may be a huge and dense “grain bin,” a common phenomenon at Kalomo Culture sites. Some remains of sorghum were found associated with this feature.

Stone, daga, and pottery feature (context 3) within context 2 in Trench 1 at Kalundu Mound (Bottom-left scale is 50 cm, north arrow scale is 35 cm)
Context 4 begins with a transition from context 2/3 into a soft, loose, brownish gray sandy silt heavily disturbed by rooting. This continued for about 60 cm with little noticeable change. However, there is a slight increase in the clay component of the sediment with depth. We also encountered dense cultural material throughout the horizon, all of which is consistent with Inskeep’s description for “Zone II.” The only exception is that we documented large amounts of daga in the trench (occasionally found in small concentrations, whereas Inskeep noted a decrease in daga from Zones I to II (Inskeep, 1962, p. 140)). The soft, powdery texture of the sediment is consistent with what has been described for decayed dung deposits in southern Kenya (Marshall et al., 2018), and so this horizon may have been similarly derived.
Context 4 gradually transitions into darker sediment with an increasingly brown hue. The matrix and inclusions remain consistent, but with a marked drop in evidence of rooting and burrowing, as well as a decrease in material culture. Based on these differences, we labeled this 20-cm horizon context 5, which is the equivalent of Inskeep’s Zone II, Spit VI. Within context 5, at 125 cm below the surface, there was a probable hearth feature which was labeled context 6. The feature was a visibly darkened circular patch 25 cm in diameter and 10 cm deep, with abundant charcoal within the matrix.
Context 7 begins at the base of context 5 with a mottled grayish-brown horizon containing high proportions of charcoal and artifacts. Sub-angular granitic inclusions from granule to small cobble sizes were common throughout the context; and they increased with depth, signaling increasing proximity to the granite-rich subsoils described by Inskeep (1962, p. 140). This certainly relates to the upper portions (Spits VII and VIII) of Inskeep’s Zone III. Excavations caught just the edge of a linear burning feature, visible in the profile as a 5-cm-thick and 65-cm-long band of dense charcoal and burnt bone. As only 3–5 cm of the feature extended into the actual trench, it was not given a separate context designation. The density of material culture varied between 10-cm spits within this horizon, possibly indicative of periodic occupations. Inskeep describes a gradual change into less mottled and more yellow sediment within Zone III. However, we found this to be more abrupt and began context 8 at this transition. Other than color, a reduction in the density of charcoal, and the continued increase in granitic inclusions, there were no other differences between contexts 7 and 8.
Only half a square meter was excavated further down through a somewhat diffuse transition into a heavily compacted darker yellow-brown sediment with only a few ceramic sherds and very little charcoal. There was an absence of fauna beyond this sediment transition. This is likely the “old ground” surface described by Inskeep (1962, p. 140) that overlays the bedrock. Inskeep encountered numerous pit features and evidence of post-holes beginning at this context and extending into the old ground surface. In some cases, these pits and postholes extend over 2 m into the bedrock (Inskeep, 1962, p. 140). However, none were observed in the 2017 excavation unit.
Chronology
The conventional 14C dates for Kalundu continue to be used in regional studies. However, taking these dates at face value is problematic because of significant advances in radiocarbon dating practice and standards. Dates were initially obtained from large samples of pooled charcoals without taxonomic identification that might evaluate old carbon effects. Additionally, ultra-filtration and other pretreatments that minimize contamination issues were not available. The resulting dates have very large error margins, meaning that they have low accuracy and low precision. To address these issues, we obtained six new 14C dates for the Kalundu sequence, using modern AMS dating methods. The dates are from three bone fragment samples and three identifiable macrobotanicals to avoid any potential old carbon sources. All bone collagen extraction and radiocarbon processing took place at the Waikato Radiocarbon Dating Laboratory in New Zealand using established methods (see Waikato Radiocarbon Dating Laboratory, 2017).
The calibrated age ranges of the six new 14C dates indicate that the mound formation occurred between c. AD 1180 and AD 1400 at the 95.4% confidence level (before modeling; Table 1, Fig. 5, but see Table 2 and Fig. 6 for modeled date ranges). These dates indicate no significant inversions, so we feel confident that they accurately represent the age of the three major mound formation/deposition phases we identified during excavations. However, the calibrated 14C age ranges of our new dates appear to drastically differ from the extant Kalomo dates and suggest a later and more rapid accumulation of Kalundu Mound (Table 1; Fig. 5). Since our excavations did not find a deep subterranean pit like those recorded in Inskeep’s larger trench, we could not run new dates related to a pre-mound occupation phase that had been identified in earlier work at the site (Fagan, 1967). Ceramics from these lower contexts are dominated by triangular stamp motifs with a sharp transition to more classic Later Iron Age Kalomo styles in the initial mound deposits, supporting a hypothesis that the pre-mound pits represent earlier habitations. Given the similarities between the earlier stamp-and-ladder designs and early Iron Age pottery from Central Zambia, dated to roughly 50 cal BC (Robertson, 2000), the original date for the base of the largest pit, cal AD 249–605 (1650 ± 90 BP, SR-65; Table 1), may accurately represent the timing of these deposits.

Calibrated age ranges of 14C dates from Kalundu Mound (Brackets under the distributions denote the 68.2% and 95.4% age ranges. The plus sign represents the median of the calibrated date range).

The structure and results of Model 3. The posterior density estimates (modeled dates) are shaded in dark gray. Notes: The plus symbols on the distributions shaded in light gray represent the original calibrations of each 14C date. A: values represent the agreement index of individual dates, while the Amodel value represents the overall model agreement. C: values denote convergence. O: values represent the posterior/prior probability that a date is an outlier. Example: assay WK-477775 had a prior probability of being an outlier set to 5% while the posterior (modeled) probability was 4%, strongly indicating this assay is not an outlier.
Any adjustment to the Kalundu Mound chronology would impact the interpretation of the Kalomo Culture sequence, the pace of landscape transformation on the Batoka Plateau, and the expansion of Kalomo ceramics (Huffman, 1982, 1989). It was, therefore, necessary to assess how well, if at all, original and new radiocarbon dates could be reconciled to form a more precise chronology for the site. We arranged our 14C assays and the earlier dates in a series of Bayesian chronological models in the OxCal v4.4.2 software package (Bronk Ramsey, 2009a, 2020) using SHCal20 (Hogg et al. 2020) to statistically build a more precise chronology of mound formation at Kalundu. Bayesian statistics are increasingly being used to quantitatively assess the temporal range of archaeological contexts and histories (Bayliss, 2009, 2015; Bronk Ramsey, 1995, 2009a, 2009b; Buck & Meson, 2015; Litton & Buck, 1995). While we do not have the room to cover the history and specifics of the method here, we point the reader to the excellent review and best-practice articles that have recently been published (Bayliss, 2015; Buck & Meson, 2015; Hamilton & Kenney, 2015; Hamilton & Krus, 2018).
The Bayesian chronological models that we developed for Kalundu Mound were organized according to the stratigraphic observations from both the original and recent excavations of the site. Because the Gulbenkian laboratory in what was then Rhodesia, where the original dates were run, is no longer operational, there is limited information available on specific protocols and standards that may have affected these dates. As a result, we used standard assumptions for averaging the effects of lumped charcoal samples. These investigations identified three primary units of mound formation and evidence for site use on a pre-mound ground surface. We arranged the 14C dates in our models according to these phases of human activity. For all models, we assumed the phases of mound formation were sequential. Sequential models leave open the possibility that the boundaries between phases could have encompassed an abandonment or gap of time between them. We present the results of our models below. The names and ranges of all posterior density estimates (modeled dates) are presented in italics and at both the 68% and 95% levels of probability (rounded to the nearest five years). This is described as the best practice for reporting the results of Bayesian chronological models (Bayliss, 2015; Hamilton & Krus, 2018). When we discuss specific function commands in OxCal, the Courier font is used to denote the Chronological Query Language (CQL2) software code underlying the program. All models were run with and without the bone collagen assays used as terminus post-quem (TPQ) dates for their corresponding horizon. Each model that we created to test our assumptions of the Kalundu Mound chronology incorporated Difference and Interval functions to examine both the duration of mound formation and potential periods of abandonment.
We ran four different models operating under slightly different possibilities for the mound formation, and we focus here on the results of our most parsimonious model (Model 3). Details of the other models; explanation of how we arrived at our preference for Model 3, including a model run for sensitivity analysis; and discussion of the time-scales for mound formation phases and times between phases of mound formation are presented in detail in Electronic Supplementary Materials (ESM 1). Associated tables and visualizations for the alternative models are provided in ESM 2-7.
Chronological Modeling Results
Our primary model takes advantage of the aggressive wood charcoal Outlier_Model function (Bronk Ramsey, 2009b; Dee & Bronk Ramsey, 2014). This was applied to dates from the Gulbenkian laboratory and the single date from UCLA because they are from unidentified charred material and, most likely, assays on a pooled sample of charred materials. The remaining dates were subjected to the general Outlier_Model to test the probability of their positions within our model. Convergence values for this model are above 95% and we estimate that after an initial period of pre-mound formation use of the site, formation of Kalundu Mound began between cal AD 1240 and 1275 and ended between cal AD 1325 and 1410 (see Fig. 6 and Table 3; note 68% CI dates are considered more probable). The total duration of site use is estimated using the Difference function as lasting 60–240 years (95% probability; Fig. 7; Site Duration) and likely 75–195 years (68% probability).

Probability distribution for the duration of site occupation at Kalundu as derived from Model 3.
While we feel that Model 3 provides the best chronological fit for our data, we explored the possibility that dates on bone collagen could be considered terminus post quem (TPQ) dates as a form of sensitivity analysis using the After function. This acknowledges the possibility that the 14C date on a disarticulated bone might represent the animal’s time of death that predates its deposition within archaeological deposits. This was done for dates from bone samples for two reasons. First, bones can resist decay better than paleobotanical remains and so may remain on a surface longer. Second, faunal elements were found in high frequency in pits and features at the Kalomo site of Isamu Pati, possibly indicating a special function or curation of these pieces before deposition (Fagan, 1967, p. 47–48). The bone preservation in Kalundu Mound was excellent, with a well-preserved cortical surface on all samples submitted for 14C analyses. There is no evidence that the bone samples submitted for dating were part of ritual caches like those at other Kalomo sites. Nevertheless, we include Model 4 to determine how much variability such taphonomic processes might introduce and to provide this as a baseline for the types of considerations that should be made for re-dating of Kalomo period sites. The results of this model are presented in ESM 5 and ESM 6. Model 4 calculates overall longer ranges for most phases at both the 68% and 95% confidence intervals relative to Model 3, which is especially apparent in the 50–100-year shift later in time of the maximum estimates for the end of each mound formation phase. Conversely, start-date estimates for each phase do not vary much between Models 3 and 4. The increased uncertainties of Model 3 are worth noting, but additional radiocarbon dates from the sequence would be needed for a more thorough comparison of Models 3 and 4.
Based on consideration of all models and available taphonomic data, observations of stratigraphy, and material culture associations, we feel that the results of our primary model (Model 3) present the most accurate revised chronology of site use and mound formation at Kalundu. The precision of this model is anchored in our new dates and their strong association with the mound stratigraphy, but this should continue to be tested as additional dates are generated. These results are different from the earlier radiocarbon chronology. They imply that the mound formation process occurred quickly, cyclically, and was much later than previously proposed.
Chronology Summary
Given the resolution of the Bayesian modeling of the radiocarbon dates from Kalundu Mound, we propose a modeled date range of cal AD 1190–1410 for site formation (see Table 3 for specific estimated ranges). The use of modeling allows for greater confidence in this date range as opposed to considering only the un-modeled calibrated dates (see un-modeled vs. modeled dates in Table 2). Patterns observed in modeling the occupational duration of different layers/phases are more consistent with a few temporally discrete site formation periods, not with continuous formation. This evidence supports the hypotheses that Kalundu Mound was periodically abandoned (or at least occupied in a lower density) between major phases, possibly in relation to broader practices of agricultural fallowing proposed by Fagan (1967, p. 43–44).
While we encountered issues attempting to rectify the original and new 14C dates in our initial models, we were able to quantitatively test whether some dates were outliers. Given that the earlier dates were most likely run on pooled sets of multiple charcoal samples very early in the development of 14C technology, the flagging of those earlier dates as outliers is not surprising. This result forces us to reconsider the early 14C data from other Kalomo sites, in addition to other long-held assumptions about the chronology of regional culture groups based on ceramics or other material culture. The results of our modeling also make it clear that additional projects that focus on recovering more datable samples from secure contexts at Kalundu and other sites on the Batoka Plateau will increase the chronological resolution and integrity of this poorly understood region.
The new chronometric data we present here is more precise than the earlier dates because they are based on bone and plant-based 14C samples. However, the continued reassessment of archaeological materials from the Kalomo group will be a good test for this assertion. Nevertheless, it is clear that the Kalomo components of Kalundu Mound formed much more recently in time and much more quickly than was previously believed. If the uppermost dates in the late fourteenth and early fifteenth centuries AD are accurate, the LIA Kalomo group would move much closer in time to, if not overlap with, the origins of material culture and linguistic signals of the ethnohistoric Tonga peoples who continue to live in the area today. The Tonga cattle-herding strategies likely have their origins in the LIA, and these findings support claims that the Tonga are, at least in part, descendants of the Kalomo peoples who were present in southern Zambia by at least the early twelfth century.
Subsistence
Faunal Remains
Excavations yielded a small assemblage of animal bone weighing 1,465.9 g, of which 1,280.2 g (531 specimens) was identifiable. However, due to high rates of fragmentation and generally poor preservation, a large proportion of the specimens are identifiable to only the broadest taxonomic categories (Table 4, see also ESM 8). There is no clear distribution of taxa across contexts. Rather, diagnostic taxa occur in small numbers throughout the stratigraphic sequence. Mammals dominate the assemblage, and most are medium- to large-sized. The large-sized mammals identified based on morphology were domesticated cattle (n=7) and caprines (n=2). The small mammals include leporids (rabbits or hares; n=9), gerbils (n=1), and unidentified rodents (n=2). Non-mammalian taxa are relatively rare, including fragmentary bivalves (n=12), birds (n=5), turtle carapace fragments (n=5), and a mandible of a large varanid (cf. Nile Monitor) (n=1).
Given the poor preservation across the sites, we applied ZooMS to 102 fragmentary specimens to improve the resolution of faunal exploitation at Kalundu (Janzen et al., 2021). Identifications aided by ZooMS are presented along with all zooarchaeological identifications in ESM 9. Collagen peptide mass fingerprints for 35 fragments matched published marker profiles for Bos when specimen size was also considered. ZooMS data also expanded the faunal inventory to include elephant (n=1), canids (n=2), and suids, as well as several wild bovid taxa, including members of the Cephalophinae (n=9), Tragelaphini (n=2), and Hippotraginae (n=2). There are also some spectra with markers consistent with Leporidae and Mustelidae (Janzen et al., 2021). However, these are diverse families, and the ZooMS markers for native African species from these families have not been studied. Therefore, these identifications should be considered tentative. Fourteen fragments did not have adequate collagen preservation for definitive identification.
Most of the faunal assemblage (1,160 g) is unburned. Burned material is limited to 83 specimens (119.2 g) distributed throughout the stratigraphic sequence. Only one specimen is calcined, indicating a level of thermal alteration beyond what is typically seen in culinary processing. Out of 436 specimens (365 with good preservation and 71 with medium cortical preservation), evidence for culinary processing occurs on 36, and mostly (28 specimens) from contexts 4.1 to 8. All evidence for culinary processing occurs on medium- to large-sized mammals. This is unsurprising, given that the overwhelming majority of specimens from this site are classified as such, and larger animals require more processing. Post-depositional surface modifications include rodent and carnivore gnawing as well as some gastric etching. Carnivore damage and gastric etching are primarily confined to the lower contexts; 25 of the total 29 specimens that exhibit those modifications are from context 4.1 or deeper in the stratigraphy. It is likely that this damage was from dogs rather than wild canids, as the faunal remains were recovered from domestic contexts. Carnivore gnawing also occurs on two specimens that exhibit butchery modifications.
Macrobotanical Remains
Here we report only the results of plant remains >1mm in size, though a brief scan of the smaller fraction suggests these account for the majority of identifiable seeds and other economic remains. In total, 3,901 charred remains were recorded (Table 5), the vast majority being charred wood (charcoal) (n= 3,222, ~83%), the identification of which is ongoing. The non-charcoal component includes the remains of crops such as finger millet (Eleusine coracana), sorghum (Sorghum bicolor) and possibly pearl millet (cf. Pennisetum glaucum), wild grasses including wild finger millet (Eleusine africana/indica), and wild fruits/nuts and legumes (Figs. 8 and 9). Seed density was highest in context 6 (hearth; 3.7 seeds/liter) but relatively low in all other contexts (0.7–1.5 seeds/liter). In contrast, contexts 6 and 8 produced the lowest densities of charred wood fragments by weight (0.14 g and 0.12 g/liter, respectively).

Examples of crops and wild relatives from Kalundu. (A) Sorghum bicolor; (B) cf. Pennisetum glaucum; (C) indet Panicoid grass; (D) Eleusine coracana; (E) Eleusine africana/indica.

Examples of Parinari sp. endocarp from Kalundu (A–D) and comparative reference material (E–F), showing morphological and anatomical features used for identification. Notes: (A) Large, near-complete fragment with preserved outer fibrous mesocarp layer and inner hard, dense endocarp layer (context 8); (B) cross section of septum showing two inner seed locules and distinctive layers of globular phytoliths (silica cells; indicated by arrows) that form under the locule walls; note also the distinctive swirling schlerenchyma tissue in the central portion of the endocarp which, in the absence of the phyolith layer, enabled a cf. Parinari identification on small fragments (context 3); (C) exposed surface of phytolith layer under the seed locule wall, with negative honeycomb-like pockmarking visible where phytoliths have degraded or been dislodged (context 3); (D) cross section of large endocarp fragment with two seed locules (context 8); (E) cross section of modern Parinari curatellifolia endocarp (charred), showing globular phytolith layer under the locule wall (arrows) (cf. Fig 13B); (F) modern Parinari curatellifolia endocarp (charred), showing globular phytolith layer (cf. Fig. 13C).
The degree of fragmentation was also high overall; the 1–2-mm fraction (charcoal and endocarp only) weighed just over half as much as the >2-mm fraction (3.80 g and 6.88 g, respectively), but contained almost six times the number of fragments (n=3,157 and 556 respectively). Fragmentation is likely the result of trampling and other on-site activities, as well as post-depositional processes such as bioturbation, given that all samples also contained some fine rootlets and occasional uncharred seeds. Overall, these data underline the need for relatively large-scale bulk sampling to obtain sufficiently representative botanical assemblages from Iron Age sites (Crowther et al., 2018). However, given that archaeobotanical analyses from Zambia have been very limited, with finds rarely subjected to direct radiocarbon dating (cf. Fagan et al., 1969, p. 81, 85), this study has nonetheless yielded significant insights into the agricultural choices of early farmers in this region.
All floated contexts from Kalundu yielded crop remains, including eight finger millet/cf. finger millet grains in contexts 3, 5, 6, and 8; a sorghum grain in context 3 directly dated to cal AD 1320–1415 (606±15 BP, Wk-47773); two sorghum glumes from context 5; and a possible pearl millet grain in context 6 (Fig. 8). Notably under-represented in the Kalundu assemblage was pearl millet (Pennisetum glaucum), a common cultivar in southern Zambia today. Its presence was indicated only by a single tentatively identified grain in context 6. Although this specimen has the distinctive club shape of domesticated Pennisetum, it is much smaller than domestic varieties. However, diminutive forms of domesticated pearl millet have been recorded archaeobotanically elsewhere in eastern Africa (Crowther et al., 2014; Walshaw, 2005). The possible explanations for the smaller size include harvesting immature grains, discard of small basal grains from the inflorescence shaft, and introgression from wild Pennisetum. Domesticated in West Africa around 2500 BC, pearl millet has not been recorded at any archaeological site in Zambia to date, though its presence is common at late first/early second millennium AD sites across southeast Africa (see Boivin et al., 2014; Crowther et al., 2018; Fuller et al., 2021). Hence, its absence in Kalundu Mound may be a sampling issue. Further analysis of the <1-mm fraction may determine if its presence can be detected more confidently through chaff (involucre bases) rather than seeds.
The most ubiquitous plant material found, aside from charred wood, was Parinari sp. (Chrysobalanaceae) endocarp (Fig. 9). Parinari occurred in all four floated contexts, while large fragments were also hand collected from three contexts. From the floated assemblages, a total of 363 fragments (2.95 g; including those identified as cf. Parinari, see Fig. 9C for details) were recovered at a rate of 12 fragments (~0.1g)/liter, which is relatively high given that charcoal density was just 0.24g/liter. This volume hints at Parinari’s overall importance as a food source to the site’s inhabitants. Parinari was most abundant in context 3 (0.17 g/liter) and decreased with depth to just 0.01 g/liter in context 8; a similar pattern of decrease in charcoal density was also observed, suggesting taphonomic factors may have affected preservation with depth. Six species of Parinari are found across tropical Africa, but P. curatellifolia (mabula plum), a common miombo woodland tree, is considered the most important for its fruit (Jud et al., 2016). The vitamin C–rich plum-like fruit ripens around October–November and can be made into a porridge or syrup called “kachasu,” which can then be fermented into a beer. The oil-rich seeds contained within the endocarp can be eaten raw or used as fuel (Smith & Allen, 2004). The Parinari endocarps were also recovered along with burnt gourd fragments from the nearby Kalomo site of Matobo (Watt, 1980).
Beans from a large-seeded (~1–2 cm) legume, tentatively identified as cf. Elephantorrhiza sp. (Fabaceae) (n=16), were found in three contexts: 5, 7, and 8. Radiocarbon dates on two of the seeds, from contexts 5 and 8, returned similar dates: cal AD 630–715 (850 ± 17 BP, Wk-47774) and cal AD 675–765 (851 ± 15 BP, Wk-47775). Elephantorrhiza is a genus of nine species restricted to sub-equatorial Africa, several of which are of minor local importance as sources of tannin, dye, and medicine, mainly from the bark of their rhizome. The seeds can be roasted, ground, and brewed to create a coffee-like beverage (Jansen, 2005). Further reference collection analyses are required to confirm this identification.
A few common weeds related to an agricultural landscape were also present, although in low frequency and abundance. These include a single example of each of the Aizoaceae and Boraginaceae families and wild finger millet (Eleusine africana/indica—owing to the degraded nature of the seed, it was not possible to determine which of the two wild species was represented). The low frequency of weed seeds is not surprising given the preference across sub-Saharan Africa for hand harvesting of millets by cutting individual heads at the stalks, thus avoiding collecting weeds. Finding wild finger millet in an assemblage that is also abundant with the domesticated variety is also not unexpected, given that this wild relative is a dominant weed in Africa and often invades E. coracana fields. Because of the similar growth and morphology of both the wild and domesticated finger millet varieties, it is difficult to weed out the wild varieties in the early stages of crop production.
Material culture
Ceramics
A total of 2,389 ceramic fragments were recovered from the test trench. Of these, only a small proportion (n=54, 2.2%) were decorated (Fig. 10). Rim forms and decorations match typical styles known for Kalomo Culture sites dominated by rounded vessels with thin walls, little to no shoulders, and a narrow band of comb-stamp decoration around the rim (equivalent to Fagan’s Class III). The next most common pot type is globular with sharply everted rims, with narrow bands of decoration along the rim (Fagan’s Class VI). Only a few rim sherds exhibit shoulder morphologies equivalent to Fagan’s Class I or II pottery. Comb-stamping is the dominant decorative motif, usually in the form of a single band formed by two borderlines with diagonal comb-stamping between them. Variations of this include bands without borderlines or where one borderline was incised rather than comb-stamped.

Typical decorative motifs on potsherds from Kalundu Mound—(a, c, h) bands of comb-stamp impressions, (b, i) incised/“thumbnail” impression motifs, (d, f) comb-stamp impressions forming triangles, (e) comb-stamp herringbone motif, (g) indeterminate punctate motif.
The second most common decorative motifs are bands of simple incised horizontal or diagonal dashes. Only a few sherds reflect other styles, including a single band of low-angled zig-zags (usually comb-stamped but in one specimen made with fine incisions), comb-stamped triangle motifs, and incised crosshatching. Five sherds exhibit external burnishing. Again, these styles are generally consistent with observations made by Inskeep (1962, p. 142–149), Fagan (1967, p. 94–17), and Katanekwa (2016, p. 87–89) for the Kalundu Mound. Many styles—especially the punctate motifs associated with earlier horizons—are not represented in this collection, likely a reflection of the limited size of the excavation.
Small Finds
Twelve beads were recovered during our excavations at Kalundu Mound. Of these, seven were made from ostrich eggshell (OES), one was made from stone, and four were Indian Ocean glass trade beads. The stone bead and all but one of the OES beads were recovered from context 1, whereas glass beads were predominantly found near the bottom of the sequence in context 8. All the glass beads are turquoise in color with diameters from 3.2 to 5.5 mm, consistent in morphology with K2-Indo-Pacific style beads dating to the late tenth to early thirteenth centuries (Koleini et al., 2016; Wood, 2011). Geochemical analyses of the trade beads to confirm this identification are forthcoming. Recovery of many beads from a 1-×-3-m test trench is significant as only a single glass bead was identified from the 1957 and 1962 excavations. Either there is spatial variation in the distribution of beads at Kalundu, or the difference is due to recovery. As three of the four glass beads found in 2017 were recovered in heavy fractions from flotation samples, the latter possibility seems more likely. The result fits the patterns of glass bead recovery on the eastern African coast by the Sealinks project, where high rates of flotation were correlated with unprecedented rates of recovery of stone, glass, and other beads, as well as shell bead production debris (Crowther et al., 2016; d’Errico et al., 2020).
Discussion
Kalundu Mound and the other Kalomo sites are frequently drawn upon for answering major archaeological and linguistic questions on the Iron Age of southeastern Africa (de Luna, 2016; de Luna & Fleisher, 2019; Fagan, 2017; Huffman, 1982, 1989, 2010; Musonda, 2013a, 2013b; Phillipson, 1970, 1975; Steele, 2006). Based on the modeling of new dates calibrated to the SHCal2020 curve, we propose a modeled site chronology of AD 1190–1410. This chronology helps test hypotheses regarding the formation of the mound site and the relationship between the Kalomo group and broader social and subsistence transitions in Iron Age south-central Africa.
Site Formation
The Kalomo Culture is associated with stratified mound sites in the Zambezi and Kafue basins that continued to be occupied across these regions into the modern era. There are two possibilities for how these mound sites formed. Kalomo mounds were thought to result from deliberate construction to raise habitations within flood plains or were created through concentrated continuous or successive occupations (Fagan, 1967, p. 43–44). There is not a clear divide between these processes; for example, initial unintentional mound formation can shift into a deliberate process if the benefits of raising settlement height are recognized. Mound formation processes reflect changing settlement patterns and seemingly a shift toward increasing sedentism. Different models of the intentionality of mound creation have implications in terms of social and labor organization and the trajectories of human-environmental interactions associated with the origins of social complexity in southern Zambia. Closer examination of site formation from economic and geophysical perspectives helps reconstruct exactly how these relationships developed.
The Kalundu Mound stratigraphy reveals 3–4 primary depositional periods, each separated by abandonment intervals of 20–45 years (see ESM 4-7). We base our chronology on Model 3 presented above, given the specific taphonomic conditions and stratigraphic observations during excavations. However, future work at Kalundu or other Kalomo period sites may reveal that an alternative model may be more appropriate.
The lowest levels of the mound (context 7, Inskeep’s Zone III) overlaying Inskeep’s “old ground surface” (equivalent to context 8) were heavily mottled and rich in fauna and material culture. There was no evidence of basket loading or other indicators of intentional construction in these earliest phases of mound formation. Heavy mottling suggests bioturbation and soil development occurring over a longer period. It is possible that taphonomic processes could obscure signs of intentional mound construction. However, the detection of numerous intact features in these and previous excavations at Kalundu makes this unlikely. Archaeological materials, including daga, are consistent with an incidental origin of mound deposits derived from human habitations.
The next major sediment package comprises a pale fine ashy substrate that was very heavily bioturbated (context 4, Inskeep’s Zone II). These sediments are identical in appearance and texture to the deposits of decayed dung documented at Pastoral Neolithic sites in Kenya and Tanzania (Marshall et al., 2018; Grillo et al., 2018). Ongoing geoarchaeological and micromorphological analyses will determine if this layer derives from animal dung deposits. These sediments are known to be nutrient-rich and are being mined for fertilizer in the surrounding areas of the site. Detection of this layer in the 1960–1962 and 2017 excavation areas indicates that it covers a large portion of the mound, with a maximum linear extent of approximately 42 m. Such a size would be excessive for a cattle corral. So it may represent a major burning phase, which would be consistent with the density of burnt daga we encountered in 2017 (but which were not associated with this layer by Inskeep (1967)).
Finally, the uppermost strata (contexts 1–2, Inskeep’s Zone I) begin with a stable surface with human occupation overlaying the ash layer of context 4/Zone II, as evidenced by the detection of an intact-rock and daga feature (context 3) at the base of this horizon. Inskeep (1962, p. 142) detected multiple pits and post-hole features in this horizon. Compared to the earlier deposits, these upper horizons reflect more intensive or longer-term habitation of the mound than the lower horizons.
Altogether, the evidence suggests Kalundu Mound formed through multiple occupational episodes rather than intentional construction. Inskeep’s Zones I, II, and III appear appropriate in demarcating three phases of occupation: initial mound formation, ash/dung accumulation or burning phase, and later occupations, respectively. These phases are likely the result of separate settlement episodes. It is interesting that the stratigraphy is so consistent across the earlier and new excavations, possibly reflecting larger synchronic occupations instead of palimpsests of multiple asynchronous events. These indicate that the mound sites were used with a broader landscape-level strategy of settlement rotation in the Batoka Plateau to permit the fallowing of agricultural fields (Fagan, 1967, p. 44). The intensity of occupation needed to form these shallow mounds across southern Zambia signals important transitions in settlement structure and land-use strategies during the Later Iron Age. More work is required to understand how this phenomenon is related to broader trends toward social complexity in southern Africa.
Iron Age Agriculture at Kalundu
The finding of finger millet from Kalundu is particularly significant, being the first archaeological example in Zambia and one of just a few in southeastern Africa. Domesticated in eastern Africa, probably somewhere in the region between Uganda and Ethiopia (Giblin & Fuller, 2011), the spread of finger millet into southeastern African has been recorded from only three sites in Mozambique, South Africa, and Zimbabwe, all dating to the seventh through eleventh century AD (Maggs & Ward, 1984; Pwiti, 1996; Sinclair, 1982). In Zambia, finger millet is usually grown in the country’s northern parts with a higher level of rainfall. Indeed, unlike other millets, it is generally regarded as a more upland-adapted crop best grown at higher (cooler) elevations and higher humidity. The relatively fertile soils and higher altitude of the Batoka Plateau (Kalundu is at ~1,200 m asl; finger millet grows best between c. 1,000 and 2,000 m asl) may have afforded more favorable growing conditions for finger millet than other drier regions of southern Zambia, where sorghum, pearl millet, and maize are preferred today. The Batoka Plateau region receives about 700–800 mm of rainfall annually, the lower limit of finger millet’s preference, though it will tolerate as little as 500 mm (Fuller, 2014). Finger millet grows best on rich soil, so it is usually the first crop grown in the chitemene rotation system used in northern Zambia today, followed by other millets and legumes. The grain is highly versatile and can be used to make porridge (nsima), bread, malt, and beer, while its straw is a highly nutritious animal fodder. Unlike sorghum, finger millet and pearl millet store exceptionally well. Hence, the grain bins found at the nearby Kalomo Culture site of Isamu Pati were likely for millet rather than sorghum (Oliver & Fagan, 1975, p. 99).
Sorghum is a major cultivar in Zambia today and has been widespread across sub-Saharan Africa since the mid-late first millennium AD. Carbonized sorghum grains were recovered at Kalundu during Fagan’s earlier campaign (Fagan, 1967, p. 62), and at numerous other Zambian Iron Age sites, including Isamu Pati, located just a few kilometers from Kalundu (Fagan, 1966; see also Robertson, 2000). Sorghum grains from Ingombe Ilede were directly dated to the seventh century AD (Fagan et al., 1969, p. 81, 85), predating the Kalundu find by several centuries. Considering that re-dating at Kalundu and Ingombe Ilede (McIntosh & Fagan, 2017) have significantly shifted the chronologies of both sites forward in time, it is now necessary to re-evaluate agricultural chronologies for the region.
While both sorghum and finger millet are known to have been important Iron Age crops, the recovery of Parinari and cf. Elephantorrhiza sp. from Kalundu shed light on the potential importance of wild plant use in the Zambian Iron Age. Likewise, morphological analyses of faunal remains from Kalundu indicate that wild resources were an important supplement to the cattle-based economies assumed for the Batoka Plateau. The animal resource base appears to have been somewhat wide, including smaller wild species in addition to the larger wild grazers identified from previous studies of tooth-rows from the site. Additionally, this study identified aquatic resources like the turtle, Nile Monitor, and bivalves transported to the anthropogenically elevated mound site. It is not clear if the remains of elephant and hippopotamus/rhinoceros represent subsistence, craft production, or other activities. There is a need for more extensive flotation and re-analysis of fauna to re-assess trajectories of subsistence change related to the shifts in settlement size and density, and increasing social complexity, in the Later Iron Age of southern Africa.
Chronology for Kalundu Mound and the Kalomo Group
The new 14C dates reported here offer a revised chronology for the Kalundu Mound site. Multiple models demonstrate that these new dates cannot easily be reconciled with the original dates and shift the timing of the mound development to cal AD 1190–1410. The formation of Kalundu Mound appears to have been more rapid and Kalomo occupation much later than previously thought. There is evidence that the basal layers at Kalundu and Isamu Pati are dominated by pottery types now recognized as the Early Iron Age channeled and punctate variants (Katanekwa, 2016, p. 218–221; Robinson et al., 1963). There is a sharp stratigraphic discontinuity between these and the Later Iron Age comb-stamp and paneled Kalomo styles. This would suggest the early dates at Kalomo sites reflect unrelated earlier occupations and not an early origin for the subsistence and cultural patterns associated with the Kalomo Culture.
Recent re-dating of elaborate burials at the nearby Ingombe Ilede suggests this site had emerged as a regionally important trading hub in the late fifteenth century AD, as opposed to the fourteenth/early fifteenth century previously proposed (McIntosh & Fagan, 2017). The analysis of the style, geology, and geochemistry of beads in these burials by Robertshaw et al. (2016) indicated some tentative similarities with beads from the Kalomo site of Isamu Pati. The styles of copper bangles and ceramics from Ingombe Ilede are also similar to those excavated at Isamu Pati (de Luna, 2017; Fagan, 1967). Other sites with Kalomo pottery like Simbusenga, Zambezi Farm, and Simonga also have dates in the range of thirteenth through fifteenth century AD (Vogel, 1971, 1973). The most parsimonious explanation for these data is that Ingombe Ilede emerged out of the existing social and economic systems, providing a model of more continuous socioeconomic change in the Zambezi Valley.
Analysis of the ceramics from Kalomo Mound led Phillipson (1968) to conclude that the pre-mound horizons dated by Inskeep and Fagan to cal AD 300–500 did reflect a separate phenomenon from the later Kalomo occupations. Early dates from Kalomo sites like Ingombe Ilede, Gundu, Matobo, and Kamangoza are solidly associated with typical Kalomo style ceramics (Fagan, 1978; Huffman, 1989; Vogel, 1970; Watt, 1980). Based on these dates, Vogel (1970) proposed AD 800 for the beginning of Kalomo. However, when subjected to modern calibration, all of these “early” dates fall as recently as cal AD 1150–1350, much closer to the lower Kalundu Mound dates reported here. These differences between previous bulk charcoal dates and newer AMS dates from Kalundu Mound (on bone and botanicals) and Ingombe Illede (on fibers) strongly indicate a systemic problem with the reliability of pre-AMS dates. The fact that two dates from a large burnt log at Matobo both fall before cal AD 1000 (Watt, 1980) supports the proposition that old-wood may be a significant problem impacting previous chronologies. Our Bayesian modeling efforts, like those that have recently refined the chronologies for Great Zimbabwe and Khami group sites in Zimbabwe (Chirikure et al., 2013a, b), demonstrate that high-resolution modeling with AMS dates, with greater sampling hygiene protocols, is necessary to understand the development of these complex stratified LIA sites.
Implications for Regional Trajectories of Social Complexity
An early thirteenth century AD origin for the Kalomo mound sites would make them contemporaneous with parallel trends toward increased settlement intensity and social complexity further south in Zimbabwe and Botswana. Mapungubwe was in decline at this time, as economic power shifted toward the Kingdom of Zimbabwe, creating cascading changes to trade networks that reached into the Zambezi. It is also around the twelfth century AD that there is evidence for dramatic social and ecological change at sites like Bosutswe, Khami, and Toutswe (Chirikure et al., 2018a,b; Denbow et al., 2008). The transition away from typical Kalomo material patterns around cal AD 1400 would likewise align with the major social changes across Zambezia, including the end of the Woolandale, Malumba, Lose, and Toutswe phenomena to the south (Chirikure, 2014; Klehm, 2017).
The regions to the north are less well understood, although the re-dating of the Iron Age occurrences in central and northern Zambia may also shift chronologies of those sites into the LIA. The Copperbelt of northern Zambia, presumably supplying the copper being traded through the Batoka Plateau, seems to reflect a different archaeological pattern. Surveys of 14 LIA sites, dating to after c. AD 110 in the Luano area near the Kafue River, indicated sites of this period are typically smaller (< 1.4 ha) than the preceding Early or Middle Iron Age sites in the region (Blisson, 1992, p. 241). In addition, LIA sites are more often along streams or on the margins of dambo wetlands, with evidence for a shift to more dispersed land use after c. AD 1500 (Blisson, 1992). While few habitations have been excavated in the Upemba Depression further north into the Democratic Republic of Congo, extensive work on burial sites may suggest a similar pattern of de-centralization ca. AD 1200–1500. There is a marked reduction in quantities of grave goods (especially markers of elite status) and simpler pottery than in the preceding periods (Dlamini, 2014, p. 12–34; de Maret, 1979). It is only after AD 1500 that these northern regions show evidence of Indian Ocean trade goods and likely an increasing copper trade (de Maret, 1999). This may indicate that the Copperbelt and proximate regions benefited less from LIA trade than the middlemen until the more intensified forms of trade associated with social changes of the fifteenth and sixteenth centuries AD discussed above. Given current evidence, re-dating indicates that social trajectories in the Zambezi Valley are consistent with the regional models for broad LIA synchronic interaction spheres characterized by “malleable hierarchies of control and situational heterarchies” (Chirikure et al., 2018a, p. 45). New research is needed to better understand how societies on the Batoka Plateau interacted with other polities during this period and how they may have exercised local agency within political and trade relationships.
Major socio-economic transitions in the second millennium AD were tied to the increasing entanglement of southeastern Africa and the Zambezi Valley with Swahili coast and interior trade networks. An increased prevalence of copper artifacts and other trade goods in Kalomo-period sites indicate that the inhabitants of these LIA sites were more engaged in (or even played a more active role in encouraging) trade between the Zambezi Valley and regions to the south on the one hand, and Greater Zambia on the other (Fagan, 1967; see also Wilmsen, 2017). Recovery of four glass trade beads from flotation samples (from the 2017 1-×-3-m trench) does reflect more integration in the Indian Ocean and Zambezi Valley trade networks than was evident from the original excavations where only one glass bead was recovered from a 42-m trench. The rise of Ingombe Ilede as a trade center in the following centuries may be due to its inhabitants managing to wrest control of this network when the presence of Portuguese traders increased demand for goods like ivory and copper (Killick, 2017; de Luna, 2016, 2017; Wilmsen, 2017).
While access to globalized trade networks was important in the development of early Zambian mound sites like Kalundu, there were likely several factors that contributed to this trajectory. A modeled occupation duration range of cal AD 1185–1405 for the Kalomo would also situate the development of these mound sites in the period of an exceptionally high frequency of southern hemisphere ENSO events that have been correlated with major drought phases in southeastern Africa (Huffman, 2008, 2009b, 2010; Moy et al., 2002). It is not yet clear how much these stresses impacted Iron Age lifeways. However, the intensification of cattle herding evident for the Kalomo period has been associated at earlier times and elsewhere in Africa with attempts to maintain predictable access to animal protein (for subsistence or ritual purposes) during periods of increasing climatic uncertainty (Marshall & Hildebrand, 2002).
As trade fueled wealth differentiation among individuals or communities, the social and economic value of cattle also increased. With evidence across southern and eastern Africa for control of trade networks shifting between particular regions, if not individual sites (e.g., Anderson, 2009; Huffman et al., 2018; Killick, 2017; Klehm, 2017), there would also have been incentives for the kind of reduced mobility on the Batoka Plateau that resulted in habitation mound accumulation. The tension between growing reliance on cattle herding and decreased mobility would have had ecological implications like increased erosion and re-distribution of soil nutrients like nitrogen and phosphorus (e.g., Marshall et al., 2018). The land clearance for grazing and agricultural intensification may explain the early European observations that the Batoka Plateau was devoid of trees. The archaeological evidence of decreased wild fauna through the Kalomo sequences also lends weight to the open vegetation scenario. It is not clear how people responded to environmental stress and degradation, but one form of resilience was increased reliance on regional trade and exchange. Pending further archaeological testing, this model suggests that the early manifestations of social complexity in southern Zambia were not caused by any single condition but rather the accumulated influence of multiple entangled local and regional forces.
Conclusions
Excavations at Kalundu Mound were instrumental in the development of Iron Age archaeology in southeastern Africa. Under the inspiration of Desmond Clark, and through the work of Ray Inskeep, Brian Fagan, David Phillipson, Joseph Vogel, Robin Derricourt, and others, the Zambezi and Kafue basins became—for a time—a center for archaeological research into late precolonial history in sub-Saharan Africa. Kalundu and other Kalomo sites have continued to play an important role in the ongoing discourse concerning major putative demographic, economic, and cultural changes in southern Africa. Renewed archaeological research at Kalundu reveals issues of chronology and subsistence that require significant revision, with implications for the broader socio-economic, interaction, and complexity models appropriate for understanding Kalomo Culture data.
One of the important contributions of these test excavations was to provide new 14C dates for Kalundu Mound. The new dates indicate that site formation began later than previously believed and accumulated more quickly through a series of habitation episodes. The revised dates suggest that the development of Kalomo Culture traditions was temporally associated with the broader shifts toward social complexity in Zimbabwe and Botswana and a period of climatic instability. However, Bayesian modeling demonstrates that even this sample of dates does not produce a definitive chronology. Considering that very few Iron Age sites in southeastern Africa have several 14C dates, much less direct dates on non-wood materials run with modern methods, many of the existing site chronologies are likely problematic. The foodways associated with early mound sites in Zambia also require new analyses that use a combination of zooarchaeology, ZooMS, and enhanced archaeobotanical recovery methods. Archaeobotanical data from Kalundu is particularly important because it demonstrates that while the Kalomo group is associated with herding-focused economies, the population maintained a significant plant cultivation component and continued to use wild food resources. In addition, enhanced recovery of small trade beads through flotation holds the potential to reshape our understanding of how connected Kalomo period sites were to broader southern African and Indian Ocean trade networks.
As a type site for the Kalomo Culture, Kalundu Mound is important to the ongoing debates on the unique trajectories of social and political complexity in southeastern Africa (Katanekwa, 2016; Kusimba et al., 2017). The results reported here highlight the need to revisit Kalomo sites with modern excavation protocols, recovery strategies, and laboratory techniques. Integrating higher resolution data for chronologies (e.g., Chirikure et al., 2013a, b), trade, subsistence, and environmental and climatic change from multiple Kalomo period sites will aid the development of more nuanced models that consider the role of local dynamics in shaping large-scale social change (after de Luna et al. 2012; Klehm, 2017; Stahl, 2002). Advancing our understanding of pre-colonial histories in Zambia will, in turn, support better reconstruction of how these sites and their occupants contributed to the rich cultural heritage of the region.
References
Anderson, M. S. (2009). Marothodi: The historical archaeology of an African capital. Atikkam Media.
Bayliss, A. (2009). Rolling out revolution: Using radiocarbon dating in archaeology. Radiocarbon, 51(1), 123–147.
Bayliss, A. (2015). Quality in Bayesian chronological models in archaeology. World Archaeology, 47(4), 677–700.
Blisson, S. (1992). A survey of Late Stone Age and Iron Age sites at Luano, Zambia. World Archaeology, 24(2), 234–248.
Boivin, N., Crowther, A., Prendergast, M., & Fuller, D. Q. (2014). Indian Ocean food globalization and Africa. African Archaeological Review, 31(4), 547–581.
Bradfield, J., Forssman, T., Spindler, L., & Antonites, A. R (2019). Identifying the animal species used to manufacture bone arrowheads in South Africa. Archaeological and Anthropological Sciences, 11, 2419–2434.
Bronk Ramsey, C. (1995). Radiocarbon calibration and analysis of stratigraphy: The OxCal Program. Radiocarbon, 37(2), 425–430.
Bronk Ramsey, C. (2009a). Bayesian analysis of radiocarbon dates. Radiocarbon, 51(1), 337–360.
Bronk Ramsey, C. (2009b). Dealing with outliers and offsets in radiocarbon dating. Radiocarbon, 51(3), 1023–1045.
Brown, S., Hebestreit, S., Wang, N., Boivin, N., Douka, K., & Richter, K. (2020). Zooarchaeology by mass spectrometry (ZooMS) for bone material - Acid insoluble protocol [WWW Document]. https://doi.org/10.17504/protocols.io.bf43jqyn
Buckley, M., Collins, M., Thomas-Oates, J., & Wilson, J. C. (2009). Species identification by analysis of bone collagen using matrix-assisted laser desorption/ionisation time-of-flight mass spectrometry. Rapid Communications in Mass Spectrometry, 23, 3843–3854.
Buck, C. E. & Meson, B. (2015). On being a good Bayesian. World Archaeology, 47(4), 1–18. doi:https://doi.org/10.1080/00438243.2015.1053977
Caton-Thompson, G. (1931). The Zimbabwe culture: Ruins and reactions. Oxford University Press.
Chirikure, S. (2014). Land and sea links: 1500 years of connectivity between southern Africa and the Indian Ocean rim regions, AD 700 to 1700. African Archaeological Review, 31(4), 705–724.
Chirikure, S., Manyanga, M., Pikirayi, I., & Pollard, M. (2013a). New pathways of sociopolitical complexity in Southern Africa. African Archaeological Review, 30(4), 339–366.
Chirikure, S., Pollard, M., Manyanga, M., & Bandama, F. (2013b). A Bayesian chronology for Great Zimbabwe: Re-threading the sequence of a vandalised monument. Antiquity, 87(337), 854–872.
Chirikure, S., Mukwende, T., Moffett, A. J., Nyamushosho, R. T., Bandama, F., & House, M. (2018a). No big brother here: Heterarchy, Shona political succession and the relationship between Great Zimbabwe and Khami, southern Africa. Cambridge Archaeological Journal, 28(1), 45–66.
Chirikure, S., Nyamushosho, R., Bandama, F., & Dandara, C. (2018b). Elites and commoners at Great Zimbabwe: Archaeological and ethnographic insights on social power. Antiquity, 92(364), 1056–1075.
Clark, J. D., Cormack, J., & Chin, S. (1969). Kalambo Falls prehistoric site, volume 3, the earlier cultures: Middle and Earlier Stone Age. Cambridge University Press.
Clark, J. D., & Fagan, B. M. (1965). Charcoals, sands, and channel decorated pottery from Northern Rhodesia. American Anthropologist, 67(2), 354–371.
Crowther, A., Horton, M. C., Kotarba-Morley, A., Prendergast, M., Quintana Morales, E., Wood, M., Shipton, C., Fuller, D. Q., Tibesasa, R., Mills, W., & Boivin, N. (2014). Iron Age agriculture, fishing and trade in the Mafia Archipelago, Tanzania: New evidence from Ukunju Cave. Azania: Archaeological Research in Africa, 49, 21–44.
Crowther, A., Faulkner, P., Prendergast, M. E., Quintana Morales, E. M., Horton, M., Wilmsen, E., Kotarba-Morley, A. M., Christie, A., Petek, N., Tibesasa, R., & Douka, K. (2016). Coastal subsistence, maritime trade, and the colonization of small offshore islands in eastern African prehistory. The Journal of Island and Coastal Archaeology, 11(2), 211–237.
Crowther, A., Prendergast, M. E, Fuller, D.Q., & Boivin N. L. (2018). Subsistence mosaics, forager-farmer interactions and the transition to food production in eastern Africa. Quaternary International, 489, 101–120.
Dee, M. W., & Bronk Ramsey, C. (2014). High-precision Bayesian modeling of samples susceptible to inbuilt age. Radiocarbon, 56(1), 83–94. https://doi.org/10.2458/56.16685.
Degerbøl, M. (1967). Dogs from the Iron Age (c. AD 950-1000) in Zambia with remarks on dogs from primitive cultures. In B. M. Fagan (Ed.), Iron Age cultures in Zambia (Kalomo and Mangila) (pp. 178–207). Chatto & Windus.
de Luna, K. M. (2012). Surveying the boundaries of historical linguistics and archaeology: Early settlement in South Central Africa. African Archaeological Review, 29, 209–251.
de Luna, K. M. (2016). Collecting food, cultivating people: Subsistence and society in Central Africa. Yale University Press.
de Luna, K. M. (2017). Rethinking Ingombe Ilede and Its Hinterland. Antiquity, 91(358), 1089–1091.
de Luna, K. M., Fleisher, J. B., & McIntosh, S. K. (2012). Thinking across the African past: Interdisciplinarity and early history. African Archaeological Review, 29(2), 75–94.
de Luna, K. M., & Fleisher, J. B. (2019). Speaking with substance. Springer.
Denbow, J. (1990). Congo to Kalahari: Data and hypotheses about the political economy of the western stream of the Early Iron Age. African Archaeological Review, 8(1), 139–175.
Denbow, J., Smith, J., Ndobochani, N. M., Atwood, K., & Miller, D. (2008). Archaeological excavations at Bosutswe, Botswana: Cultural chronology, paleo-ecology and economy. Journal of Archaeological Science, 35(2), 459–480.
Denbow, J., Klehm, K., & Dussubieux, L. (2015). The glass beads of Kaitshàa and early Indian Ocean trade into the far interior of southern Africa. Antiquity, 89, 361–377.
Derricourt, R. M. (1976). Archaeology in Zambia: An historical outline. African Social Research, 21, 31–50.
Derricourt, R. M. (1985). Man on the Kafue: The archaeology and history of the Itezhitezhi area of Zambia. Ethnographica.
d’Errico, F., Martí, A. P., Shipton, C., Le Vraux, E., Ndiema, E., Goldstein, S., Petraglia, D., & Boivin, N. (2020). Trajectories of cultural innovation from the Middle to Later Stone Age in Eastern Africa: Personal ornaments, bone artifacts, and ocher from Panga ya Saidi, Kenya. Journal of Human Evolution, 141, 102737.
Dlamini, N. (2014). The early inhabitants of the Upemba Depression, the Democratic Republic of Congo. Ph.D. dissertation, University of Cape Town, Cape Town.
Fagan, B. M. (1966). The Iron Age of Zambia. Current Anthropology, 7(4), 453–462.
Fagan, B. M. (1967). Iron Age cultures in Zambia (Kalomo and Kangila). Chatto & Windus.
Fagan, B. M. (1969). Early trade and raw materials in South Central Africa. The Journal of African History, 10, 1–13.
Fagan, B. M. (1978). Gundu and Ndonde, Basanga and Mwanamaimpa. Azania, 13(1), 127–134.
Fagan, B. M. (1993). Ray Inskeep and the Kalomo culture. The South African Archaeological Bulletin, 48(158), 115–117.
Fagan, B. M., Phillipson, D. W., & Daniels, S. G. H. (1969). Iron Age cultures in Zambia, 2. Chatto & Windus.
Fuller, D. Q. (2014). Finger millet, origins and development of. In C. Smith (Ed.), Encyclopedia of Global Archaeology. Springer.
Fuller, D. Q., Barron, A., Champion, L., Dupuy, C., Commelin, D., Raimbault, M., & Denham, T. (2021). Transition from wild to domesticated pearl millet (Pennistum glaucum) revealed in ceramic temper at three Middle Holocene sites in Northern Mali. African Archaeological Review. https://doi.org/10.1007/s10437-021-09428-8.
Giblin, J., & Fuller, D. Q. (2011). First and second millennium AD agriculture in Rwanda: Archaeobotanical finds and radiocarbon dates from seven sites. Vegetation History and Archaeobotany, 20, 253–265.
Hamilton, W. D., & Kenney, J. (2015). Multiple Bayesian modelling approaches to a suite of radiocarbon dates from ovens excavated at Ysgol yr Hendre, Caernarfon, North Wales. Quaternary Geochronology, 25, 72–82. https://doi.org/10.1016/j.quageo.2014.10.001.
Hamilton, W. D., & Krus, A. M. (2018). The myths and realities of Bayesian chronological modeling revealed. American Antiquity, 83(2), 187–203. https://doi.org/10.1017/aaq.2017.57.
Hogg, A. G., Heaton, T. J., Hua, Q., Palmer, J. G., Turney, C. S., Southon, J., et al. (2020). SHCal20 Southern Hemisphere calibration, 0–55,000 years cal BP. Radiocarbon, In Press. https://doi.org/10.1017/RDC.2020.59.
Holden, C. J. (2006). The spread of Bantu languages, farming, and pastoralism in sub-equatorial Africa. In C. P. Lipo (Ed.), Mapping our ancestors (pp. 247–268). Routledge.
Huffman, T. N. (1970). The Early Iron Age and the spread of the Bantu. The South African Archaeological Bulletin, 25(97), 3–21.
Huffman, T. N. (1982). Archaeology and ethnohistory of the African Iron Age. Annual Review of Anthropology, 11(1), 133–150.
Huffman, T. N. (1989). Ceramics, settlements and Late Iron Age migrations. The African Archaeological Review, 7(1), 155–182.
Huffman, T. N. (2008). Climate change during the Iron Age in the Shashe-limpopo basin, southern Africa. Journal of Archaeological Science, 35(7), 2032–2047.
Huffman, T. N. (2009a). Mapungubwe and Great Zimbabwe: The origin and spread of social complexity in southern Africa. Journal of Anthropological Archaeology, 28(1), 37–54.
Huffman, T. N. (2009b). A cultural proxy for drought: Ritual burning in the Iron Age of southern Africa. Journal of Archaeological Science, 36(4), 991–1005.
Huffman, T. N. (2015). Social complexity in southern Africa. African Archaeological Review, 232(1), 71–91.
Huffman, T. N., Manyanga, M., Tarduno, J. A., & Watkeys, M. K. (2018). The archaeological context of recent archaeomagnetic research in Zimbabwe. Southern African Humanities, 31(1), 205–221.
Inskeep, R. R. (1962). Some Iron Age sites in Northern Rhodesia. The South African Archaeological Bulletin, 17(67), 136–180.
Jansen, C. M. (2005). Elephantorrhiza elephantina. In P. C. M. Jansen and D. Cardon (Eds.), Plant resources of tropical Africa 3: dyes and tannins (pp. 75-76). PROTA Foundation, Backhuys Publishers, Wageningen, The Netherlands.
Janzen, A., Richter, K. K., Mwebi, O., Brown, S., Onduso, V., Gatwiri, F., Ndiema, E., Katongo, M., Goldstein, S. T., Douka, K., & Boivin, N. (2021). Distinguishing African bovids using zooarchaeology by mass spectrometry (ZooMS): New peptide markers and assessments of Iron Age economies in Zambia. PLoS One, 16(5), e0251061. https://doi.org/10.1371/journal.pone.0251061.
Jud, N. A., Nelson, C. W., & Herrera, F. (2016). Fruits and wood of Parinari from the early Miocene of Panama and the fossil record of Chrysobalanaceae. American Journal of Botany, 103(2), 277–289.
Katanekwa, N. M. (1978). Some Early Iron Age sites from the Machili Valley of South Western Zambia. Azania, 13(1), 135–166.
Katanekwa, N. M. (1979). Namakala and Nanga Sites and the chronology of the Early Iron Age in Southern & South-West Zambia. The South African Archaeological Bulletin, 34(130), 120–122.
Katanekwa, N. M. (2005). Ingombe Ilede. In Shillington, K. (Ed.), Encyclopedia of African history Volume 2 (pp. 679–680). Fitzroy Dearborn.
Katanekwa, N. M. (2016). The prehistory of the 73+ Bantu languages and Bantu language groups of Zambia 3000 BC to 1600 AD. Published by author.
Killick, D. (2017). Tracing Ingombe Ilede’s trade connections. Antiquity, 91(358), 1087–1088.
Klehm, C. E. (2017). Local dynamics and the emergence of social inequality in Iron Age Botswana. Current Anthropology, 58(5), 604–633.
Koleini, F., Prinsloo, L. C., Biemond, W. M., Colomban, P., Ngo, A. T., Boeyens, J. C., & Van der Ryst, M. M. (2016). Towards refining the classification of glass trade beads imported into Southern Africa from the 8th to the 16th century AD. Journal of Cultural Heritage, 19, 435–444.
Kusimba, C. M., Kim, N. C. & Kusimba, S. B. (2017). Trade and state formation in ancient East African coast and southern Zambezia. In R. J. Chacon and R. G. Mendoza (Eds.), Feast, famine or fighting? (pp. 61-89). Springer International.
Litton, C. D., & Buck, C. E. (1995). The Bayesian approach to the interpretation of archaeological data. Archaeometry, 37(1), 1–24.
Maggs, T., & Ward, V. (1984). Early Iron Age sites in the Muden area of Natal. Annals of the Natal Museum, 26, 105–140.
de Maret, P. (1979). Luba roots: The first complete Iron Age sequence in Zaire. Current Anthropology, 20(1), 233–235.
de Maret, P. (1999). The power of symbols and the symbols of power through time: Probing the Luba past. In S. K. McIntosh (Ed.), Beyond chiefdoms: Pathways to complexity in Africa (pp. 151–165). Cambridge University Press.
Marshall, F. B., & Hildebrand, E. (2002). Cattle before crops: The beginnings of food production in Africa. Journal of World Prehistory, 16(2), 99–143.
Marshall, F. B., Reid, R. E., Goldstein, S., Storozum, M., Wreschnig, A., Hu, L., Kiura, P., Shahack-Gross, R., & Ambrose, S. H. (2018). Ancient herders enriched and restructured African grasslands. Nature, 561(7723), 387.
MacDonald, K. C. (1992). The domestic chicken (Gallus gallus) in sub-Saharan Africa: A background to its introduction and its osteological differentiation from indigenous fowls (Numidinae and Francolinus sp.). Journal of Archaeological Science, 19, 303–318.
McIntosh, S. K., & Fagan, B. M. (2017). Re-dating the Ingombe Ilede burials. Antiquity, 91(358), 1069–1077.
Moy, C. M., Seltzer, G. O., Rodbell, D. T., & Anderson, D. M. (2002). Variability of El Niño/Southern Oscillation activity at millennial timescales during the Holocene epoch. Nature, 420(6912), 162–165.
Mukwende, T., Bandama, F., Chirikure, S., & Nyamushosho, R. T. (2018). The chronology, craft production and economy of the Butua capital of Khami, southwestern Zimbabwe. Azania: Archaeological Research in Africa, 53(4), 477–506.
Musonda, F. B. (1987). The significance of pottery in Zambian Later Stone Age contexts. African Archaeological Review, 5(1), 147–158.
Musonda, F. B. (2013a). Looking back and looking forward: Iron Age studies in Southern Zambia and their contribution to the understanding of the Tonga. In L. Cliggett & V. Bond (Eds.), Tonga timeline: Appraising sixty years of multidisciplinary research in Zambia and Zimbabwe (pp. 3–24). Lembani Trust.
Musonda, F. B. (2013b). Ethnicity and ethical issues in archaeological interpretation: The case of Chakeluka Iron Age site, Lusaka, Zambia. South African Archaeological Bulletin, 68(197), 52–62.
Ndlovu, N. (2011). Management versus preservation: Archaeological heritage management in a transforming South Africa. Conservation and Management of Archaeological Sites, 13(2–3), 123–133.
Nikis, N., & Livingstone Smith, A. (2017). Copper, trade and polities: Exchange networks in Southern Central Africa in the 2nd Millennium CE. Journal of Southern African Studies, 43(5), 895–911.
Oliver, R., & Fagan, B. M. (1975). Africa in the Iron Age c. 500 B.C. to A.D. 1400. Cambridge University Press.
Phillipson, D. W. (1968). The Early Iron Age in Zambia-regional variants and some tentative conclusions. Journal of African History, 9(2), 191–211.
Phillipson, D. W. (1970). Notes on the later prehistoric radiocarbon chronology of eastern and southern Africa. The Journal of African History, 11(1), 1–5.
Phillipson, D. W. (1974). Iron Age history and archaeology in Zambia. Journal of African History, 15(1), 1–25.
Phillipson, D. W. (1975). The chronology of the Iron Age in Bantu Africa. The Journal of African History, 16(3), 321–342.
Pikirayi, I. (2002). The Zimbabwe culture: Origins and decline of southern Zambezian states. Rowman Altamira.
Pikirayi, I., & Pwiti, G. (1999). States, traders, and colonists: Historical archaeology in Zimbabwe. Historical Archaeology, 33(2), 73–89.
Prendergast, M. E., Buckley, M., Crowther, A., Frantz, L., Eager, H., Lebrasseur, O., Hutterer, R., Hulme-Beaman, A., Van Neer, W., Douka, K., & Veall, M. A. (2017). Reconstructing Asian faunal introductions to eastern Africa from multi-proxy biomolecular and archaeological datasets. PloS one, 12(8), e0182565.
Pwiti, G. (1996). Settlement and subsistence of prehistoric farming communities in the mid-Zambezi Valley, northern Zimbabwe. South African Archaeological Bulletin, 51, 3–6.
Robertshaw, P., Wood, M., Melchiorre, E., Popelka-Filkoff, R., & Glascock, M. (2010). Southern African glass beads: Chemistry, glass sources, and patterns of trade. Journal of Archaeological Science, 17, 1898–1912.
Robertson, J. H. (2000). Early Iron Age archaeology in central Zambia. Azania: Archaeological Research in Africa, 35(1), 147–182.
Robinson, K. (1959). Khami ruins: Report on excavations undertaken for the commission for the preservation of natural and historical monuments and relics, Southern Rhodesia, 1947–1955. Cambridge University Press.
Robinson, K., Shee, J. C., & Barnard, K. H. (1963). Further excavations in the Iron Age deposits at the Tunnel site, Gokomere Hill, Southern Rhodesia. The South African Archaeological Bulletin, 18(72), 155–171.
Russell, T., Silva, F., & Steele, J. (2014). Modelling the spread of farming in the Bantu-speaking regions of Africa: An archaeology-based phylogeography. PLoS One, 9(1), e87854.
Schofìeld, J. F. (1948). Primitive pottery: An introduction to South African ceramics, prehistoric and protohistoric. South African Archaeological Society Handbook Series no. 3. Cape.
Scudder, T. (1962). The ecology of the Gwembe Tonga. Manchester University Press.
Sinclair, P. (1982). Chibuene: An early trading city in southern Mozambique. Paideuma, 28, 149–164.
Sinclair, P. J., Pikirayi, I., Pwiti, G., & Soper, R. (1993). Urban trajectories on the Zimbabwean plateau. In B. Andah, A. Okpoko, T. Shaw, & P. Sinclair (Eds.), The archaeology of Africa: Food, metals and towns (pp. 705–731). Routledge.
Smith, P., & Allen, Q. (2004). Field guide to the trees and shrubs of the Miombo Woodlands. University of Chicago Press.
Summers, R. F. (1958). Inyanga: Prehistoric settlements in Southern Rhodesia. Cambridge University Press.
Stahl, A. (2002). Political economic mosaics: Archaeology of the last two millennia in tropical sub-Saharan Africa. Annual Review of Anthropology, 33, 145–172.
Swan, L. (2007). Economic and ideological roles of copper ingots in prehistoric Zimbabwe. Antiquity, 81, 999–1012.
Huffman, T. N. (2010) Intensive El Niño and the Iron Age of South-eastern Africa. Journal of Archaeological Science 37(10), 2572–2586.
Vogel, J. O. (1970). The Kalomo culture of southern Zambia: Some notes toward a reassessment. Zambia Museums Journal, 1, 77–88.
Vogel, J. O. (1971). Kumadzulo, an early Iron Age village site in southern Zambia. Oxford University Press.
Vogel, J. O. (1973). Some Early Iron Age sites in southern and western Zambia. Azania, 8(1), 25–54.
Walshaw, S. C. (2005). Swahili urbanization, trade and food production: Botanical perspectives from Pemba Island, Tanzania, A.D. 600-1500. Ph.D. dissertation. Washington University in St. Louis.
Watt, E. (1980). Excavations at Matobo Iron Age site, Kalomo, Zambia. Zambia Museums Journal, 5, 79–85.
Welker, F., Soressi, M., Rendu, W., Hublin, J.-J., & Collins, M. (2015). Using ZooMS to identify fragmentary bone from the Late Middle/Early Upper Palaeolithic sequence of Les Cottés, France. Journal of Archaeological Science, 54, 279–286.
Welker F., Hajdinjak, M., Talamo, S., Jaouen, K., Dannemann, M., David, F., Julien, M., Meyer, M., Kelso, J., Barnes, I., Brace, S., Kamminga, P., Fisher, R., Kessler, B. M., Stewart, J. R., Pääbo, S., Collins, M. J. & Hublin, J. J. (2016). Palaeoproteomic evidence identifies archaic hominins associated with the Châtelperronian at the Grotte du Renne. Proceedings of the National Academy of Sciences, 113, 11162–11167.
Waikato Radiocarbon Dating Laboratory. (2017). AMS Processing Technical Report. Radiocarbon Dating Laboratory, University of Waikato, NZ. https://radiocarbondating.com/__data/assets/pdf_file/0010/387712/Waikato-Radiocarbon-Dating-Laboratory-AMS-Processing-Technical-Report-2017.pdf. Accessed 02/03/2019.
Wilmsen, E. N. (2017). Baubles, bangles and beads: Commodity exchange between the Indian Ocean region and interior Southern Africa during 8th–15th Centuries CE. Journal of Southern African Studies, 43(5), 913–926.
Woldekiros, H. S., & D’Andrea, A. C. (2017). Early evidence for domestic chickens (Gallus gallus domesticus) in the Horn of Africa. International Journal of Osteoarchaeology, 27(3), 329–341.
Wood, M. (2011). A glass bead sequence for southern Africa. Journal of African Archaeology, 9, 67–84.
Wood, M. (2016). Glass beads from pre-European contact sub-Saharan Africa: Peter Francis’s work revisited and updated. Archaeological Research in Asia, 6, 65–80.
Acknowledgements
Archaeological excavations were carried out under a research permit issued by the National Heritage Conservation Commission of Zambia (NHCC), and the permission for all sample exports was granted under the Export Permit of Relics (#NHCC/CB/17/25/9/2017). We are grateful for the assistance and support of the NHCC, especially Mr. Macmillan Mudenda and Mr. Richard Mbombo, for facilitating this research. We are also indebted to Dr. George Mudenda and the staff of the Livingstone Museum for generous help throughout this research project. Finally, we thank Dr. Francis Musonda, Dr. Brian Fagan, Dr. Kathryn de Luna, and Dr. Lawrence Barham for discussions, advice, and support.
Funding
Open Access funding enabled and organized by Projekt DEAL. The Max Planck Society provided funding for this project.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Archaeological Time Period: Iron Age (c.1200–400 BP)
County and Region: Zambia, southern-central Africa
Supplementary Information
ESM 1:
Supplemental text. (DOCX 27 kb)
ESM 2:
Results of Model 1. These results are depicted visually in ESM 5a. Note: the -5 error on the Difference commands are an artifact of the modeling. (XLSX 15 kb)
ESM 3:
Results of Model 2. These results are depicted visually in ESM 5b. (XLSX 15 kb)
ESM 4:
Results of Model 3. Note: the -5 error on the Difference commands are an artifact of the modeling. (XLSX 14 kb)
ESM 5:
Results of Model 4. These results are depicted visually in ESM 5c. (XLSX 14 kb)
ESM 6:
Structure and results of Models 1, 2, and 4; (a) The structure and results of Model 1. The plus symbols on the distributions shaded in light gray represent the original calibrations of each 14C dates. The posterior density estimates (modeled dates) are shaded in dark gray. A: values represent the agreement index of individual dates, while the Amodel value represents the overall model agreement. C: values denote convergence; (b) The structure and results of Model 2. Dates with green distributions represent dates that pass the general Outlier_Model analysis applied to this model. Red distributions mark dates that the outlier analysis flagged as possible outliers. O: values represent the posterior/prior probability that a date is an outlier. Example: assay SR-65 had a prior probability of being an outlier set to 5% while the posterior (modeled) probability was 100%, strongly indicating this assay is an outlier; (c) The structure and results of Model 4. (PDF 517 kb)
ESM 7:
Duration of Site Occupation from Model 4. (TIF 1533 kb)
ESM 8:
Expanded table of zooarchaeological identifications with peptide identifications of faunal samples tested using Zooarchaeology by Mass Spectrometry (ZooMS). (XLSX 57 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Goldstein, S.T., Crowther, A., Henry, E.R. et al. Revisiting Kalundu Mound, Zambia: Implications for the Timing of Social and Subsistence Transitions in Iron Age Southern Africa. Afr Archaeol Rev 38, 625–655 (2021). https://doi.org/10.1007/s10437-021-09440-y
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10437-021-09440-y