
Effects of Stocking Density on Growth Performance and Stress Responses of Bester and Bester ♀ × Beluga ♂ Juveniles in Recirculating Aquaculture Systems

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Fish Maintenance
2.2. Water Quality Parameters
2.3. Growth Performance
- Weight gain (WG, %) = [(BWf − BWi)/BWi] × 100, where BWi and BWf are the initial and final average body weight (g) of fish sampled from each tank, and t is the experimental period in each trial (day);
- Specific growth rate (SGR, %/day) = [(lnBWf − lnBWi)/t] × 100;
- Fulton’s condition factor (K, %) = [BWf (g)/(SL (cm)3] × 100, where SL = standard body length;
- Feed conversion ratio (FCR) = FI (g)/BG (g), where FI stands for food consumption (food provided – uneaten food) and BG is biomass gain per tank;
- Protein efficiency ratio (PER) = BG/protein consumed;
- Hepatosomatic index (HSI) = [liver weight (g)/body weight (g)] × 100);
- Viscerosomatic index (VSI) = [visceral weight (g)/body weight (g)] × 100);
- The coefficient of variability (CV) = CvBW (%) = 100 (SD/mean BW), and was calculated for the body weight on the initial (CvBWi) and final (CvBWf) days of the experiment.
2.4. Sampling Protocol and Blood Analysis
2.5. Data Analysis
3. Results
3.1. Trial 1 (T1)
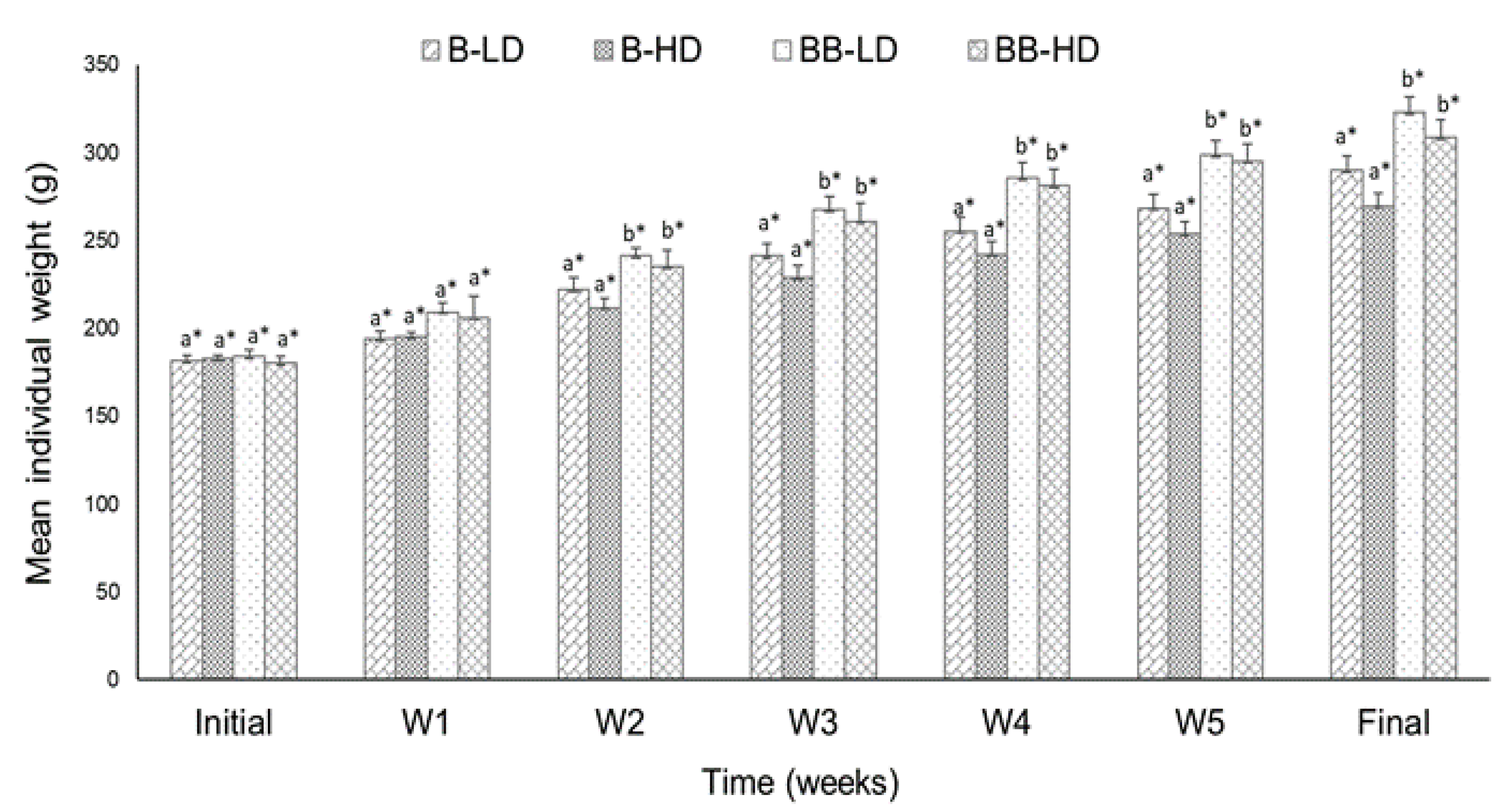
3.1.1. Growth Performance
3.1.2. Hematological Parameters and Blood Indices
3.2. Trial 2 (T2)
3.2.1. Growth Performance
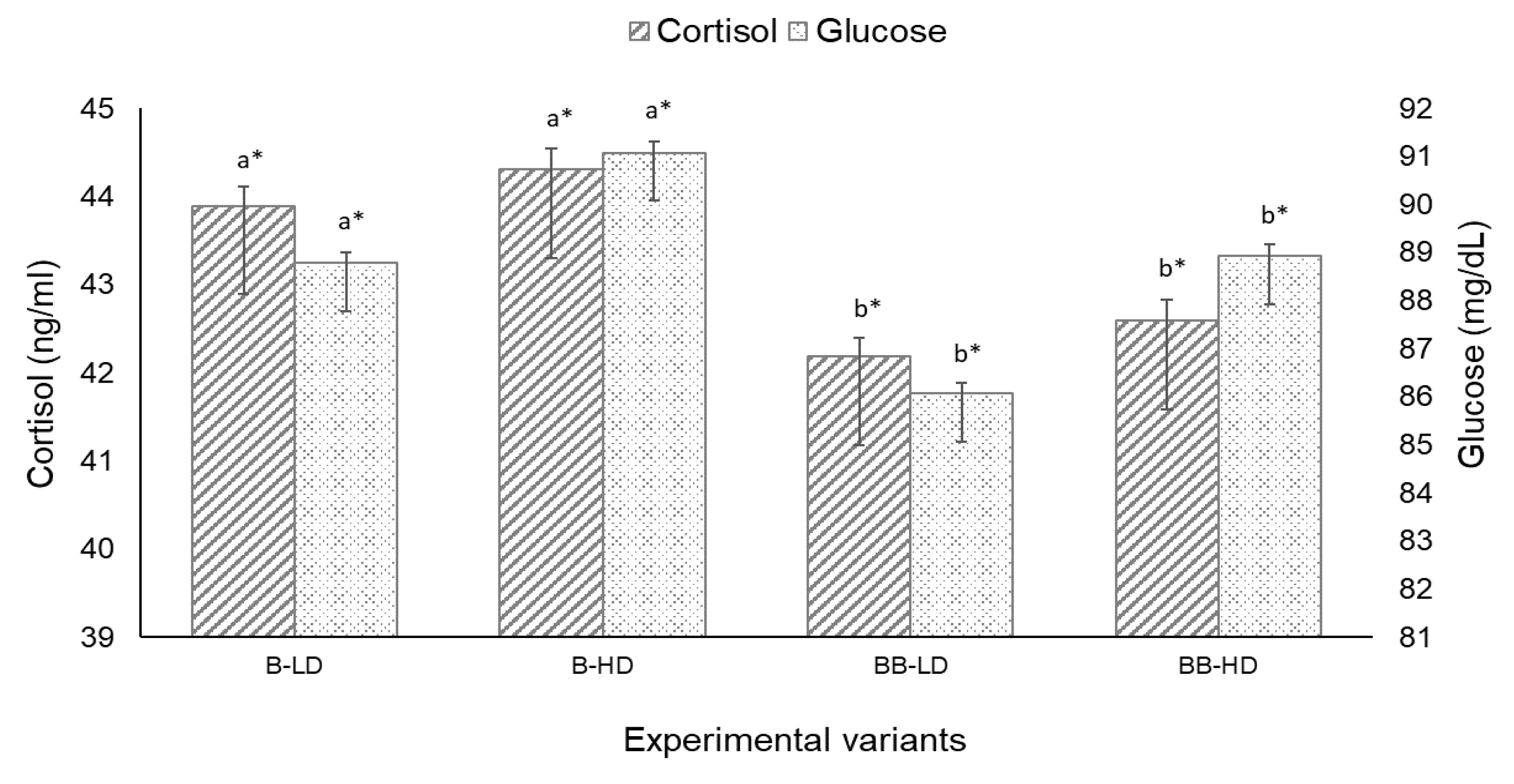
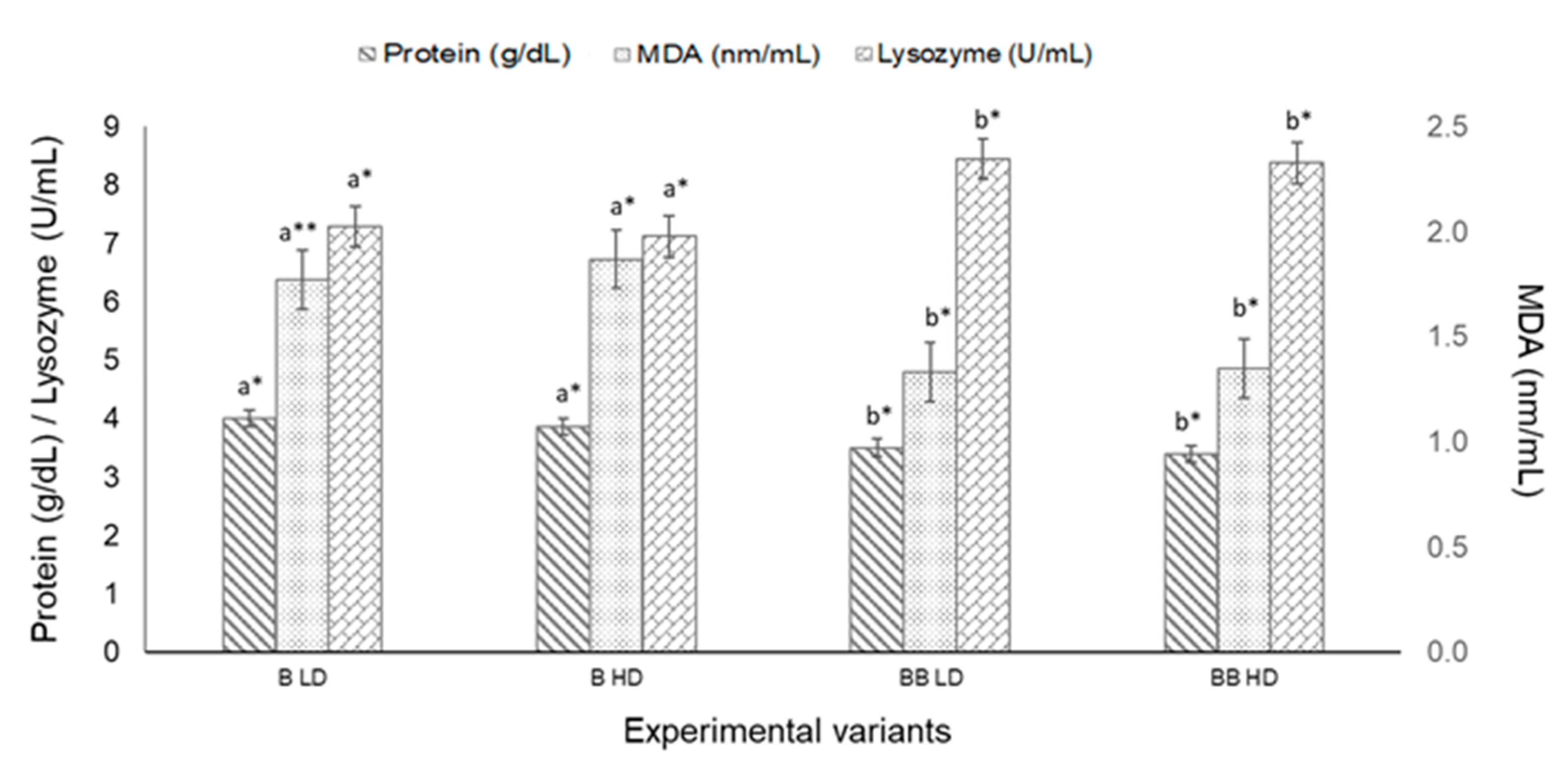
3.2.2. Haematological Parameters and Blood Indices
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, D.; Ma, G.; Sun, D. Sturgeon Culture: Status and Practices. In Aquaculture in China: Success Stories and Modern Trends; Gui, J.F., Tang, Q., Li, Z., Liu, J., De Silva, S.S., Eds.; John Wiley & Sons Publishing: Chichester, UK, 2018; pp. 234–245. [Google Scholar]
- Vasilyeva, L.M.; Elhetawy, A.I.G.; Sudakova, N.V.; Astafyeva, S.S. History, Current Status and Prospects of Sturgeon Aquaculture in Russia. Aquac. Res. 2019, 50, 979–993. [Google Scholar] [CrossRef]
- Vasilyeva, L.M.; Sudakova, N.V. Biological and Technological Bases of Commodity Sturgeon Production; ASU Publishing House: Astrakhan, Russia, 2014; 247p. [Google Scholar]
- Ponomareva, E.N.; Belaya, M.M. Modern Sterlet Acipenser ruthenus Broodstock Management in Southern Russia. World Aquac. 2013, 44, 55–59. [Google Scholar]
- Murray, F.; Bostock, J.; Fletcher, D. Review of Recirculation Aquaculture System Technologies and Their Commercial Application; University of Stirling Aquaculture: Stirling, UK, 2014. [Google Scholar]
- Iguchi, K.; Ogawa, K.; Nagae, M.; Ito, F. The influence of rearing density on stress response and disease susceptibility of ayu (Plecoglossus altivelis). Aquaculture 2003, 220, 515–523. [Google Scholar] [CrossRef]
- Bauer, O.N.; Pugachev, O.N.; Voronin, V.N. Study of Parasites and Diseases of Sturgeons in Russia: A Review. J. Appl. Ichthyol. 2002, 18, 420–429. [Google Scholar] [CrossRef]
- Pankhurst, N.W.; Van Der Kraak, G. Evidence That Acute Stress Inhibits Ovarian Steroidogenesis in Rainbow Trout in Vivo, Through the Action of Cortisol. Gen. Comp. Endocrinol. 2000, 117, 235–237. [Google Scholar] [CrossRef] [PubMed]
- Doroshov, S.I.; Moberg, G.P.; van Eenennaam, J.P. Observations on the Reproductive Cycle of Cultured White Sturgeon, Acipenser transmontanus. Environ. Biol. Fishes 2002, 48, 265–278. [Google Scholar] [CrossRef]
- Bronzi, P.; Chebanov, M.; Michaels, J.T.; Wei, Q.; Rosenthal, H.; Gessner, J. Sturgeon meat and caviar production: Global update 2017. J. Appl. Ichthyol. 2019, 35, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Lopez, A.; Vasconi, M.; Bellagamba, F.; Mentasti, T.; Moretti, V.M. Sturgeon Meat and Caviar Quality from Different Cultured Species. Fishes 2020, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- Chebanov, M.S.; Galich, E.V. Sturgeon Hatchery Manual; Food and Agriculture Organization of the United Nations: Ankara, Turkey, 2011; 338p. [Google Scholar]
- Wuertz, S.; Gaillard, S.; Barbisan, F.; Carle, S.; Congiu, L.; Forlani, A.; Aubert, J.; Kirschbaum, F.; Tosi, E.; Zane, L.; et al. Extensive Screening of Sturgeon Genomes by Random Screening Techniques Revealed No Sex-Specific Marker. Aquaculture 2006, 258, 685–688. [Google Scholar] [CrossRef]
- Baradaran, N.S.; Bahmani, M.; Abdolhay, H.; Hosseini, M.R.; Chakmehduz, F.; Hallajian, A.; Darvishi, S.; Farabi, M.V. Bester (Beluga ♀ × Sterlet ♂) Production and Comparing Their Growth with Beluga in Iran; Iranian Fisheries Science Research Institute: Tehran, Iran, 2009; 55p. [Google Scholar]
- Zharkenov, D.; Pekli, J.; Kirichrko, O.; Zsuga, K.; Sadykulov, T. Cage cultivation of bester in East Kazakhstan. Columella J. Environ. Agric. Sci. 2016, 3, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Burtsev, I.A. Bester in aquaculture. In Sturgeon Stocks and Caviar Trade Workshop; Birstein, V.J., Bauer, A., Kaiser-Pohlmann, A., Eds.; Occasional Paper of the IUCN Species Survival Commission (SSC) No. 17; IUCN: Gland, Switzerland; Cambridge, UK, 1995; ISBN 2-8317-0401-4. [Google Scholar]
- Kazemi, R.; Yarmohammadi, M.; Hallajian, A.; Tizkar, B. Determination of gender and sexual maturity stages of reared great sturgeon (Huso huso) using blood plasma sex steroid ratios. Iran. J. Fish. Sci. 2020, 19, 2185–2198. [Google Scholar] [CrossRef]
- Montero, D.; Izquierdo, M.S.; Tort, L.; Robaina, L.; Vergara, J.M. High stocking density produces crowding stress altering some physiological and biochemical parameters in gilthead seabream, Sparus aurata, juveniles. Fish Physiol. Biochem. 1999, 20, 53–60. [Google Scholar] [CrossRef]
- Conte, F.S. Stress and the Welfare of Cultured Fish. Appl. Anim. Behav. Sci. 2004, 86, 205–223. [Google Scholar] [CrossRef]
- North, B.P.; Turnbull, J.F.; Ellis, T.; Porter, M.J.; Migaud, H.; Bron, J.; Bromage, N.R. The Impact of Stocking Density on the Welfare of Rainbow Trout (Oncorhynchus mykiss). Aquaculture 2006, 255, 466–479. [Google Scholar] [CrossRef]
- Lupatsch, I.; Santos, G.A.; Schrama, J.W.; Verreth, J.A.J. Effect of Stocking Density and Feeding Level on Energy Expenditure and Stress Responsiveness in European Sea Bass Dicentrarchus labrax. Aquaculture 2010, 298, 245–250. [Google Scholar] [CrossRef]
- Turnbull, J.; Bell, A.; Adams, C.; Bron, J.; Huntingford, F. Stocking Density and Welfare of Cage Farmed Atlantic Salmon: Application of a Multivariate Analysis. Aquaculture 2005, 243, 121–132. [Google Scholar] [CrossRef]
- Ni, M.; Wen, H.; Li, J.; Chi, M.; Bu, Y.; Ren, Y.; Zhang, M.; Song, Z.; Ding, H. The Physiological Performance and Immune Responses of Juvenile Amur Sturgeon (Acipenser schrenckii) to Stocking Density and Hypoxia Stress. Fish. Shellfish Immunol. 2014, 36, 325–335. [Google Scholar] [CrossRef]
- Hasanalipour, A.; Eagderi, S.; Poorbagher, H.; Bahmani, M. Effects of Stocking Density on Blood Cortisol, Glucose and Cholesterol Levels of Immature Siberian Sturgeon (Acipenser baerii Brandt, 1869). Turk. J. Fish. Aquat. Sci. 2013, 13, 1–6. [Google Scholar] [CrossRef]
- Rafatnezhad, S.; Falahatkar, B.; Gilani, M.H.T. Effects of Stocking Density on Haematological Parameters, Growth and Fin Erosion of Great Sturgeon (Huso huso) Juveniles. Aquac. Res. 2008, 39, 1506–1513. [Google Scholar] [CrossRef]
- Jodun, W.A.; Millard, M.J.; Mohler, J. The Effect of Rearing Density on Growth, Survival, and Feed Conversion of Juvenile Atlantic Sturgeon. N. Am. J. Aquac. 2002, 64, 10–15. [Google Scholar] [CrossRef]
- Zare, R.; Bahmani, M.; Yavari, V.; Kazimi, R.; Pasha, H.; Pourdehghani, M.; Fazeli, N.; Yooneszade, B.; Nateghi, S.A. The effects of Rearing Density on Growth Performance and Food Conversion Ratio of Siberian Sturgeon (Acipenser baeri Brandt). Asian Fish. Sci. 2009, 22, 107–115. [Google Scholar] [CrossRef]
- Celikkale, M.S.; Memis, D.; Ercan, E.; Cagiltay, F. Growth Performance of Juvenile Russian Sturgeon (Acipenser gueldenstaedtii Brandt & Ratzenburg, 1833) at Two Stocking Densities in Net Cages. J. Appl. Ichthyol. 2005, 21, 14–18. [Google Scholar] [CrossRef]
- Vasilean, I.; Cristea, V.; Sfetcu, L. Influence of stocking density and water parameters on growth of juvenile beluga sturgeon (Huso huso, Linnaeus, 1758). Bull. UASVM Anim. Sci. Biotechnol. 2009, 52, 666–671. [Google Scholar]
- Aalimahmoudi, M.; Bavarsad, S.S.; Moghdani, S. Effect of Different Stocking Densities on Haematological and Biochemical Parameters of Great Sturgeon Juveniles (Huso huso Linnaeus, 1758). Res. Opin. Anim. Vet. Sci. 2015, 5, 348–352. [Google Scholar]
- Irani, A.; Agh, N. Optimization of Stocking Density for Beluga (Huso huso) and Persian Sturgeon (Acipenser persicus) culture. J. Fish. Sci. Technol. 2019, 8, 15–22. [Google Scholar]
- Bayunova, L.; Barannikova, I.; Semenkova, T. Sturgeon stress reactions in aquaculture. J. Appl. Ichthyol. 2002, 18, 397–404. [Google Scholar] [CrossRef]
- Long, L.; Zhang, H.; Ni, Q.; Liu, H.; Wu, F.; Wang, X. Effects of stocking density on growth, stress, and immune responses of juvenile Chinese sturgeon (Acipenser sinensis) in a recirculating aquaculture system. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 219, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Barton, B.A. Stress in Fishes: A Diversity of Responses with Particular Reference to Changes in Circulating Corticosteroids. Integr. Comp. Biol. 2002, 42, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, D.A.; Elliott, R.F.; Donofrio, M.C.; Patrick, S.F. Larval lake sturgeon production and drift behavior in the Menominee and Oconto Rivers, Wisconsin. Ecol. Freshw. Fish 2020, 29, 722–738. [Google Scholar] [CrossRef]
- Falahatkar, B.; Poursaeid, S. Stress Responses of Great Sturgeon Huso huso Subjected to Husbandry Stressors. Aquac. Int. 2013, 21, 947–959. [Google Scholar] [CrossRef]
- Feshalami, M.Y.; Mozanzadeh, M.T.; Amiri, F.; Mortezavizadeh, S.S.; Gisbert, E. Optimal Stocking Density for Beluga, Huso huso, and Ship Sturgeon, Acipenser nudiventris during the Grow-out Phase. J. Appl. Ichthyol. 2019, 35, 303–306. [Google Scholar] [CrossRef]
- Szczepkowski, M.; Szczepkowska, B.; Piotrowska, I. Impact of higher stocking density of juvenile Atlantic sturgeon, Acipenser oxyrinchus Mitchill, on fish growth, oxygen consumption, and ammonia excretion. Fish. Aquat. Life 2011, 19, 59–67. [Google Scholar] [CrossRef]
- Yang, D.G.; Zhu, Y.J.; Luo, Y.P.; Zhao, J.H.; Chen, J.W. Effect of stocking density on growth performance of juvenile Amur Sturgeon (Acipenser schrenckii). J. Appl. Ichtyol. 2009, 27, 541–544. [Google Scholar] [CrossRef]
- Ni, M.; Wen, H.; Li, J.; Chi, M.; Bu, Y.; Ren, Y.; Zhang, M.; Song, Z.; Ding, H. Effects of stocking density on mortality, growth and physiology of juvenile Amur sturgeon (Acipenser schrenckii). Aquac. Res. 2014, 47, 1596–1604. [Google Scholar] [CrossRef]
- Shaluei, F.; Hedayati, A.; Jahanbakhshi, A.; Baghfalaki, M. Physiological Responses of Great Sturgeon (Huso huso) to Different Concentrations of 2-Phenoxyethanol as an Anesthetic. Fish Physiol. Biochem. 2012, 38, 1627–1634. [Google Scholar] [CrossRef] [PubMed]
- Svoboda, Z.; Pravola, D.; Palackova, J. Unified Methods of Haematological Examination of Fish; Research Institute of Fish Culture and Hydrobiology: Vodnary, Czechoslovakia, 1991. [Google Scholar]
- Hesser, E.F. Methods for routine fish hematology. Progress. Fish Cult. 1960, 22, 164–171. [Google Scholar] [CrossRef]
- Haney, D.C.; Hursh, D.A.; Mix, M.C.; Winton, J.R. Physiological and Hematological Changes in Chum Salmon Artificially Infected with Erythrocytic Necrosis Virus. J. Aquat. Anim. Health 1992, 4, 48–57. [Google Scholar] [CrossRef]
- Draper, H.H.; Hadley, M. Malondialdehyde Determination as Index of Lipid Peroxidation. Methods Enzymol. 1990, 186, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, L.J. The Effects of Stocking Density on Fish Welfare. Plymouth Stud. Sci. 2010, 4, 372–383. [Google Scholar]
- Aketch, B.O.; Ang’ienda, P.O.; Radull, J.O.; Waindi, E.N. Effect of Stocking Density on the Expression of Glucose Transporter Protein 1 and Other Physiological Factors in the Lake Victoria Nile Tilapia, Oreochromis niloticus (L.). Int. Aquat. Res. 2014, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Saillant, E.; Fostier, A.; Haffray, P.; Menu, B.; Laureau, S.; Thimonier, J.; Chatain, B. Effects of Rearing Density, Size Grading and Parental Factors on Sex Ratios of the Sea Bass (Dicentrarchus labrax L.) in Intensive Aquaculture. Aquaculture 2003, 221, 183–206. [Google Scholar] [CrossRef]
- Ellis, T.; North, B.; Scott, A.P.; Bromage, N.R.; Porter, M.; Gadd, D. The Relationships between Stocking Density and Welfare in Farmed Rainbow Trout. J. Fish Biol. 2002, 61, 493–531. [Google Scholar] [CrossRef]
- Ashagrie, G.; Abebe, G.; Mengistou, S. Effect of stocking density on the growth performance and yield of Nile tilapia (Oreochromis niloticus L., 1758) in a cage culture system in Lake Kuriftu, Ethiopia. Aquac. Res. 2008, 39, 1450–1460. [Google Scholar] [CrossRef]
- Aksungur, N.; Aksungur, M.; Akbulut, B. Effects of Stocking Density on Growth Performance, Survival and Food Conversion Ratio of Turbot (Psetta Maxima) in the Net Cages on the Southeastern Coast of the Black Sea. Turk. J. Fish. Aquat. Sci. 2007, 7, 147–152. [Google Scholar]
- Lambert, Y.; Dutil, J.D. Food Intake and Growth of Adult Atlantic Cod (Gadus morhua L.) Reared under Different Conditions of Stocking Density, Feeding Frequency and Size-Grading. Aquaculture 2001, 192, 233–247. [Google Scholar] [CrossRef]
- Sloman, K.A.; Wilson, L.; Freel, J.A.; Taylor, A.C.; Metcalfe, N.B.; Gilmour, K.M. The effects of increased flow rates on linear dominance hierarchies and physiological function in brown trout, Salmo trutta. Can. J. Zool. 2002, 80, 1221–1227. [Google Scholar] [CrossRef]
- Clauss, T.M.; Dove, A.D.; Arnold, J.E. Hematologic disorders of fish. Vet. Clin. N. Am. Exot. Anim. Pract. 2008, 11, 445–462. [Google Scholar] [CrossRef] [PubMed]
- Fajfer, S.; Meyers, L.; Willman, G.; Carpenter, T.; Hansen, M. Growth of Juvenile Lake Sturgeon Reared in Tanks at Three Densities. N. Am. J. Aquac. 1999, 61, 331–335. [Google Scholar] [CrossRef]
- Mohler, J.W.; King, M.K.; Farrell, P.R. Growth and Survival of First-Feeding and Fingerling Atlantic Sturgeon under Culture Conditions. N. Am. J. Aquac. 2000, 62, 174–183. [Google Scholar] [CrossRef]
- Gülten, K.; Ferit, R.; Mevlüt, K. Growth Performance and Feed Conversion Efficiency of Siberian Sturgeon Juveniles (Acipenser baeri) Reared in Concrete Raceways. Turk. J. Vet. Anim. Sci. 2000, 4, 435–442. [Google Scholar]
- Schram, E.; Van der Heur, J.W.; Kamstra, A.; Verdegen, M.C.J. Stocking Density—Dependent Growth of Dover Sole (Solea solea). Aquaculture 2006, 252, 339–347. [Google Scholar] [CrossRef]
- Tolussi, C.E.; Hilsdorf, A.W.; Canneppele, D.; Moreira, R.G. The Effect of Stocking Density in Physiological Parameters and Growth of the Endangered Teleost Specie Piabanba bryconinsignis. Aquaculture 2010, 310, 221–228. [Google Scholar] [CrossRef]
- Docan, A.; Dediu, L.; Crețu, M.; Mogodan, A. Plasma Biochemical Responses of Hybrid Bester Juveniles Reared at Different Stocking Densities. Aquac. Aquar. Conserv. Legis. 2017, 10, 1085–1090. [Google Scholar]
- Kolman, R.; Kapusta, A. Food characteristic and feeding management on sturgeon with a special focus on Siberian sturgeon. In The Siberian Sturgeon (Acipenser baerii, Brandt, 1869); Williot, P., Nonnotte, G., Chebanov, M., Eds.; Springer: Cham, Switzerland, 2018; Volume 2, pp. 75–85. [Google Scholar]
- Mommsen, T.P.; Vijayan, M.M.; Moon, T.W. Cortisol in teleosts: Dynamics, mechanisms of action, and metabolic regulation. Rev. Fish Biol. Fish. 1999, 9, 211–268. [Google Scholar] [CrossRef]
- Mazeaud, M.M.; Mazeaud, F.; Donaldson, E.M. Primary and Secondary Effects of Stress in Fish: Some New Data with General Review. Trans. Am. Fish. Soc. 1977, 106, 201–212. [Google Scholar] [CrossRef]
- Polakof, S.; Panserat, S.; Soengas, J.L.; Moon, T.W. Glucose Metabolism in Fish: A Review. J. Comp. Physiol. B 2012, 182, 1015–1045. [Google Scholar] [CrossRef]
- Marco, P.; McKenzie, D.J.; Mandich, A.; Bronzi, P.; Cataldi, E.; Cataudella, S. Influence of Sampling Conditions on Blood Chemistry Values of Adriatic Sturgeon Acipenser naccarii (Bonaparte, 1836). J. Appl. Ichthyol. 1999, 15, 73–77. [Google Scholar] [CrossRef]
- Li, D.; Liu, Z.; Xie, C. Effect of Stocking Density on Growth and Serum Concentrations of Thyroid Hormones and Cortisol in Amur Sturgeon, Acipenser schrenckii. Fish. Physiol. Biochem. 2012, 38, 511–520. [Google Scholar] [CrossRef]
- Hamlin, H.J.; Moore, B.C.; Edwards, T.M.; Larkin, I.L.V.; Boggs, A.; High, W.J.; Main, K.L.; Guillette, L.J. Nitrate-Induced Elevations in Circulating Sex Steroid Concentrations in Female Siberian Sturgeon (Acipenser baeri) in Commercial Aquaculture. Aquaculture 2008, 281, 118–125. [Google Scholar] [CrossRef]
- Belanger, J.M.; Son, J.H.; Laugero, K.D.; Moberg, G.P.; Doroshov, S.I.; Lankford, S.E.; Cech, J.J. Effects of Short-Term Management Stress and ACTH Injections on Plasma Cortisol Levels in Cultured White Sturgeon, Acipenser transmontanus. Aquaculture 2001, 203, 165–176. [Google Scholar] [CrossRef]
- Semenkova, T.B.; Bayunova, L.V.; Boev, A.A.; Dyubin, V.P. Effects of Stress on Serum Cortisol Levels of Sturgeon in Aquaculture. J. Appl. Ichthyol. 1999, 15, 270–272. [Google Scholar] [CrossRef]
- Lucas, A. Physical concepts of bioenergetics. In Bioenergetics of Aquatic Animals; Lucas, A., Ed.; Taylor & Francis: London, UK, 1996; 169p. [Google Scholar] [CrossRef]
- Iwama, G.K.; Vijayan, M.M.; Forsyth, R.B.; Ackerman, P.A. Heat shock proteins and physiological stress in fish. Am. Zool. 1999, 39, 901–909. [Google Scholar] [CrossRef] [Green Version]
- Wedemeyer, G.A.; Barton, B.A.; McLeay, D.J. Stress and acclimation. In Methods for Fish. Biology; Schreck, C.B., Moyle, P.B., Eds.; American Fisheries Society: Bethesda, MD, USA, 1990; pp. 491–527. [Google Scholar]
- Vazzana, M.; Cammarata, M.; Cooper, E.L.; Parrinello, N. Confinement stress in sea bass (Dicentrarchus labrax) depress peritoneal leucocyte cytotoxicity. Aquaculture 2002, 210, 231–243. [Google Scholar] [CrossRef]
- Pickering, A.D. Stress and Fish.; Academic Press: New York, NY, USA; London, UK, 1981; p. 367. [Google Scholar]
- Jeney, G.; Jeney, Z. Application of Immunostimulants for Modulation of the Non-Specific Defense Mechanisms in Sturgeon Hybrid: Acipenser ruthenus × A. baerii. J. Appl. Ichtyol. 2002, 18, 416–419. [Google Scholar] [CrossRef]
- Atamanalp, M.; YanÝk, T.; HaliloÛlu, I.; Aras, M.S. Alterations in the Hematological Parameters of Rainbow Trout (Oncorhynchus mykiss) Exposed to Cypermethrin. Isr. J. Aquac. Bamidgeh 2002, 54, 99–103. [Google Scholar]
- Hrubec, T.C.; Smith, S.A. Hematology of fish. In Schalm’s Veterinary Hematology, 5th ed.; Feldman, B.F., Zinkl, J.G., Jain, M.C., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2000; pp. 1120–1125. [Google Scholar]
- Falahatkar, B.; Bagheri, M.; Efatpanah, I. The Effect of Stocking Densities on Growth Performance and Biochemical Indices in New Hybrid of Leuciscus aspius♀ × Rutilus frisii♂. Aquac. Rep. 2019, 15, 200–207. [Google Scholar] [CrossRef]
- Chester, A.; Glomski, J.T.; Chainani, M. The Phylogenetic Odyssey of the Erythrocyte. III. Fish, The Lower Vertebrate Experience. Histol. Histopathol. 1992, 7, 501–528. [Google Scholar]
- Chesneau, V. Immunology in Sturgeons with a Focus on the Siberian Sturgeon Mechanisms, Responses to Stress and Stimulation. In The Siberian Sturgeon (Acipenser baerii, Brandt, 1869); Williot, P., Nonnotte, G., Chebanov, M., Eds.; Springer: Cham, Switzerland, 2018; Volume 2, pp. 373–402. [Google Scholar]
- Aksakal, E.; Ekinci, D.; Erdoğan, O.; Beydemir, Ş.; Alım, Z.; Ceyhun, S.B. Increasing Stocking Density Causes Inhibition of Metabolic–Antioxidant Enzymes and Elevates MRNA Levels of Heat Shock Protein 70 in Rainbow Trout. Livest. Sci. 2011, 141, 69–75. [Google Scholar] [CrossRef]
- Karataş, T.; Kocaman, E.M. Comparison of Paraoxonase Activity, Malondialdehyde and High Density Lipoprotein Levels in Cultuvated Normal and Albino Rainbow Trout Reared in the Same Conditions. Kafkas Univ. Vet. Fak. Derg. 2012, 18, 87–90. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, G.; Nie, Z.; Li, Q.; Shao, N.; Xu, P. Effect of Stocking Density on Growth, Serum Biochemical Parameters, Digestive Enzymes Activity and Antioxidant Status of Largemouth Bass, Micropterus salmoides. Pak. J. Zool. 2019, 51, 1509–1517. [Google Scholar] [CrossRef]
- Andrade, T.; Afonso, A.; Pérez-Jiménez, A.; Oliva-Teles, A.; de lasHeras, V.; Mancera, J.M.; Serradeiro, R.; Costas, B. Evaluation of Different Stocking Densities in a Senegalese Sole (Solea senegalensis) Farm: Implications for Growth, Humoral Immune Parameters and Oxidative Status. Aquaculture 2015, 438, 6–11. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, W.; Du, X.; Jiang, J.; Wang, C.; Wang, X.; Zhang, Q.; He, Y. Molecular Characterization and Functional Analysis of the GATA4 in Tongue Sole (Cynoglossus semilaevis). Comp. Biochem. Physiol. B. Biochem. Mol. Biol. 2016, 193, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Coeurdacier, J.L.; Dutto, G.; Gasset, E.; Blancheton, J.-P. Is Total Serum Protein a Good Indicator for Welfare in Reared Sea Bass (Dicentrarchus labrax)? Aquat. Living Resour. 2011, 24, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Naderi, M.; Keyvanshokooh, S.; Salati, A.P.; Ghaedi, A. Effects of chronic high stocking density on liver proteome of rainbow trout (Oncorhynchus mykiss). Fish. Physiol. Biochem. 2017, 43, 1373–1385. [Google Scholar] [CrossRef] [PubMed]
- Munteanu, G.; Bogatu, D. Tratat de ihtiopatologie; Excelsior Art: Timişoara, Romania, 2003; 27p, ISBN 973-592-085-9. [Google Scholar]
- Leatherland, J.F.; Cho, C.Y. Effect of Rearing Density on Thyroid and Interrenal Gland Activity and Plasma and Hepatic Metabolite Levels in Rainbow Trout, Salmo gairdneri Richardson. J. Fish Biol. 1987, 27, 583–592. [Google Scholar] [CrossRef]
- Vijayan, M.M.; Ballantyne, J.S.; Leatherland, J.F. High Stocking Density Alters the Energy Metabolism of Brook Charr, Salvelinus fontinalis. Aquaculture 1990, 88, 371–381. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameter | Bester (B) | Bester × Beluga (BB) | ||
|---|---|---|---|---|
| LD | HD | LD | HD | |
| Initial density (kg/m2) | 2 | 4 | 2 | 4 |
| Final density (kg/m2) | 3.1 | 5.9 | 3.5 | 6.7 |
| IW (g) | 181.47 ± 1.06 a* | 182.48 ± 0.94 a* | 183.87 ± 1.68 a* | 180.10 ± 0.31 a* |
| IL (cm) | 27.62 ± 0.58 a* | 27.54 ± 0.43 a* | 26.21 ± 0.54 a* | 26.00 ± 0.49 a* |
| FW (g) | 289.73 ± 1.93 a** | 269.42 ± 6.06 a* | 322.63 ± 6.66 b** | 308.60 ± 4.47 b* |
| FL (cm) | 34.60 ± 0.36 a* | 33.10 ± 0.40 a* | 32.76 ± 0.68 a* | 31.93 ± 1.25 a* |
| WG (%) | 59.66 ± 1.99 a** | 47.62 ± 2.16 a* | 75.56 ± 1.78 b* | 71.30 ± 3.11 b* |
| CvBWi (%) | 5.72 | 4.87 | 9.2 | 6.9 |
| CvBWf (%) | 18.81 | 16.39 | 29.12 | 24.03 |
| SGR (%/day) | 1.23 ± 0.03 a** | 1.02 ± 0.04 a* | 1.48 ± 0.04 b* | 1.41 ± 0.07 b* |
| FCR | 0.85 ± 0.08 a** | 0.97 ± 0.07 a* | 0.74 ± 0.09 b** | 0.85 ± 0.11 b* |
| PER | 2.17 ± 0.01a** | 1.91 ± 0.01 a* | 2.88 ± 0.01 b** | 2.18 ± 0.01 b* |
| K | 0.87 ± 0.07 a* | 0.85 ± 0.03 a* | 0.92 ± 0.18 b* | 0.90 ± 0.07 b* |
| VSI (%) | 6.72 ± 0.19 a** | 6.03 ± 0.22 a* | 6.91 ± 0.29 b* | 6.89 ± 0.31 b* |
| HSI (%) | 2.21 ± 0.19 a** | 1.82 ± 0.15 a* | 2.13 ± 0.17 b* | 1.98 ± 0.14 b* |
| Parameter | Bester (B) | Bester × Beluga (BB) | ||
|---|---|---|---|---|
| LD | HD | LD | HD | |
| RBC (106/mL) | 0.43 ± 0.10 b** | 0.58 ± 0.13 a* | 0.44 ± 0.06 b** | 0.62 ± 0.14 b* |
| PCV (%) | 29.16 ± 3.12 a* | 29.10 ± 2.09 a* | 22.52 ± 3.90 b** | 20.87 ± 3.30 b* |
| Hb (g/dL) | 6.57 ± 0.57 a** | 6.17 ± 0.60 a* | 5.96 ± 0.98 b* | 5.86 ± 0.51 b* |
| MCV (fl) | 707.65 ± 136.77 a** | 520.59 ± 94.39 a* | 524.56 ± 114.93 b** | 351.07 ± 85.35 b* |
| MCH (pg) | 159.39 ± 29.31 a** | 110.29 ± 21.17 a* | 139.14 ± 29.59 b** | 98.12 ± 16.00 b* |
| MCHC (g/dL) | 22.68 ± 2.43 a** | 21.18 ± 0.79 a* | 26.63 ± 2.56 b** | 28.55 ± 3.27 b* |
| Parameter | SD1 | SD2 | SD3 | SD4 | Equation | R2 | p |
|---|---|---|---|---|---|---|---|
| Initial density (kg/m2) | 5 | 8 | 10 | 12 | - | - | - |
| IW (g) | 327.50 ± 7.12 | 329.55 ± 5.26 | 326.85 ± 8.24 | 323.86 ± 4.56 | - | - | - |
| CvBWi (%) | 9.55 | 8.14 | 9.98 | 10.89 | - | - | - |
| IL (cm) | 33.25 ± 0.87 | 33.38 ± 0.98 | 33.28 ± 1.23 | 33.167 ± 1.45 | - | - | - |
| Final density (kg/m2) | 9 | 13 | 16 | 20 | - | - | - |
| FW (g) | 608.35 ± 10.17 | 546.50 ± 12.56 | 526.35 ± 13.89 | 523.21 ± 12.23 | y = 2.312x2 − 49.070x + 800.930 | 0.99 | 0.008 |
| CvBWf (%) | 22.76 | 24.77 | 26.72 | 31.72 | y = 1.217x + 15.841 | 0.89 | 0.048 |
| FL (cm) | 39.13 ± 1.17 | 39.04 ± 0.89 | 38.44 ± 0.95 | 39.23 ± 1.35 | y = 0.025x2 − 0.432x + 40.805 | 0.33 | 0.770 |
| WG (%) | 85.75 ± 3.13 | 65.83 ± 2.77 | 61.03 ± 4.22 | 61.55 ± 5.83 | y = 0.612x2 − 13.427x + 134 | 0.99 | 0.037 |
| SGR (%/day) | 1.63 ± 0.11 | 1.33 ± 0.15 | 1.25 ± 0.43 | 1.26 ± 0.34 | y = 0.009x2 − 0.205x + 2.383 | 0.99 | 0.046 |
| FCR | 0.91 ± 0.04 | 0.94 ± 0.02 | 1.11 ± 0.04 | 1.15 ± 0.05 | y = 0.037x + 0.69 | 0.88 | 0.038 |
| PER | 2.03 ± 0.08 | 1.96 ± 0.07 | 1.67 ± 0.10 | 1.61 ± 0.07 | y = −0.066x + 2.402 | 0.89 | 0.050 |
| K | 0.92 ± 0.03 | 0.91 ± 0.01 | 0.91 ± 0.03 | 0.91 ± 0.01 | y = 0.0004x2 − 0.008x + 0.951 | 0.74 | 0.098 |
| VSI (%) | 7.19 ± 0.43 | 6.96 ± 0.29 | 6.12 ± 0.04 | 4.67 ± 0.07 | y = −0.071x2 + 0.858x + 4.687 | 0.99 | 0.008 |
| HSI (%) | 2.92 ± 0.14 | 2.91 ± 0.68 | 2.61 ± 0.95 | 2.23 ± 0.98 | y = −0.021x2 + 0.270x + 2.118 | 0.84 | 0.040 |
| Parameter | SD1 | SD2 | SD3 | SD4 | Equation | R2 | p |
|---|---|---|---|---|---|---|---|
| RBC (106/mL) | 0.52 ± 0.10 | 0.69 ± 0.11 | 0.82 ± 0.29 | 0.86 ± 0.39 | y = 0.050x + 0.280 | 0.97 | 0.015 |
| PVC (%) | 21.95 ± 6.63 | 21.86 ± 1.74 | 23.04 ± 1.86 | 24.90 ± 4.47 | y = 0.108x2 − 1.425x + 26.343 | 0.99 | 0.033 |
| Hb (g/dL) | 6.08 ± 0.68 | 6.28 ± 2.70 | 6.24 ± 0.53 | 6.69 ± 0.40 | y = 0.013x2 − 0.144x + 6.4984 | 0.86 | 0.366 |
| MCV (fl) | 430.95 ± 126.70 | 325.27 ± 49.33 | 301.75 ± 78.18 | 323.37 ± 112.75 | y = 5.060x2 − 101.510x + 812.27 | 0.99 | 0.021 |
| MCH (pg) | 116.74 ± 17.66 | 74.31 ± 39.48 | 81.75 ± 21.71 | 87.54 ± 26.73 | y = 2.110x2 − 39.602x + 260.87 | 0.83 | 0.201 |
| MCHC (g/dL) | 29.34 ± 9.90 | 23.71 ± 11.78 | 27.15 ± 2.35 | 27.53 ± 4.86 | y = 0.293x2 − 5.109x + 47.235 | 0.67 | 0.781 |
| MDA (g/dL) | 1.03 ± 0.19 | 0.88 ± 0.11 | 0.97 ± 0.07 | 1.37± 0.13 | y = 0.026x2 − 0.4002x + 2.378 | 0.99 | 0.009 |
| Protein (g/dL) | 4.57 ± 0.23 | 4.11 ± 0.08 | 3.87 ± 0.07 | 4.07 ± 0.08 | y = 0.025x2 − 0.509x + 6.498 | 0.95 | 0.023 |
| Glucose (mg/dL) | 88.14 ± 0.79 | 88.98 ± 0.51 | 89.64 ± 0.88 | 90.81 ± 1.06 | y = 0.370x + 86.147 | 0.96 | 0.089 |
| Cortisol (ng/mL) | 42.48 ± 0.21 | 42.51 ± 0.27 | 43.82 ± 0.24 | 43.91 ± 0.43 | y = 0.020x2 − 0.106x + 42.425 | 0.81 | 0.431 |
| Lysozyme (U/mL) | 10.50 ± 0.65 | 9.23 ± 0.22 | 8.81 ± 0.21 | 8.42 ± 0.25 | y = −0.295x + 11.824 | 0.95 | 0.032 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dediu, L.; Docan, A.; Crețu, M.; Grecu, I.; Mogodan, A.; Maereanu, M.; Oprea, L. Effects of Stocking Density on Growth Performance and Stress Responses of Bester and Bester ♀ × Beluga ♂ Juveniles in Recirculating Aquaculture Systems. Animals 2021, 11, 2292. https://doi.org/10.3390/ani11082292
Dediu L, Docan A, Crețu M, Grecu I, Mogodan A, Maereanu M, Oprea L. Effects of Stocking Density on Growth Performance and Stress Responses of Bester and Bester ♀ × Beluga ♂ Juveniles in Recirculating Aquaculture Systems. Animals. 2021; 11(8):2292. https://doi.org/10.3390/ani11082292
Chicago/Turabian StyleDediu, Lorena, Angelica Docan, Mirela Crețu, Iulia Grecu, Alina Mogodan, Marilena Maereanu, and Lucian Oprea. 2021. "Effects of Stocking Density on Growth Performance and Stress Responses of Bester and Bester ♀ × Beluga ♂ Juveniles in Recirculating Aquaculture Systems" Animals 11, no. 8: 2292. https://doi.org/10.3390/ani11082292